
Association of Polygenic Variants with Type 2 Diabetes Risk and Their Interaction with Lifestyles in Asians

 , and
, and 
Abstract
:1. Introduction
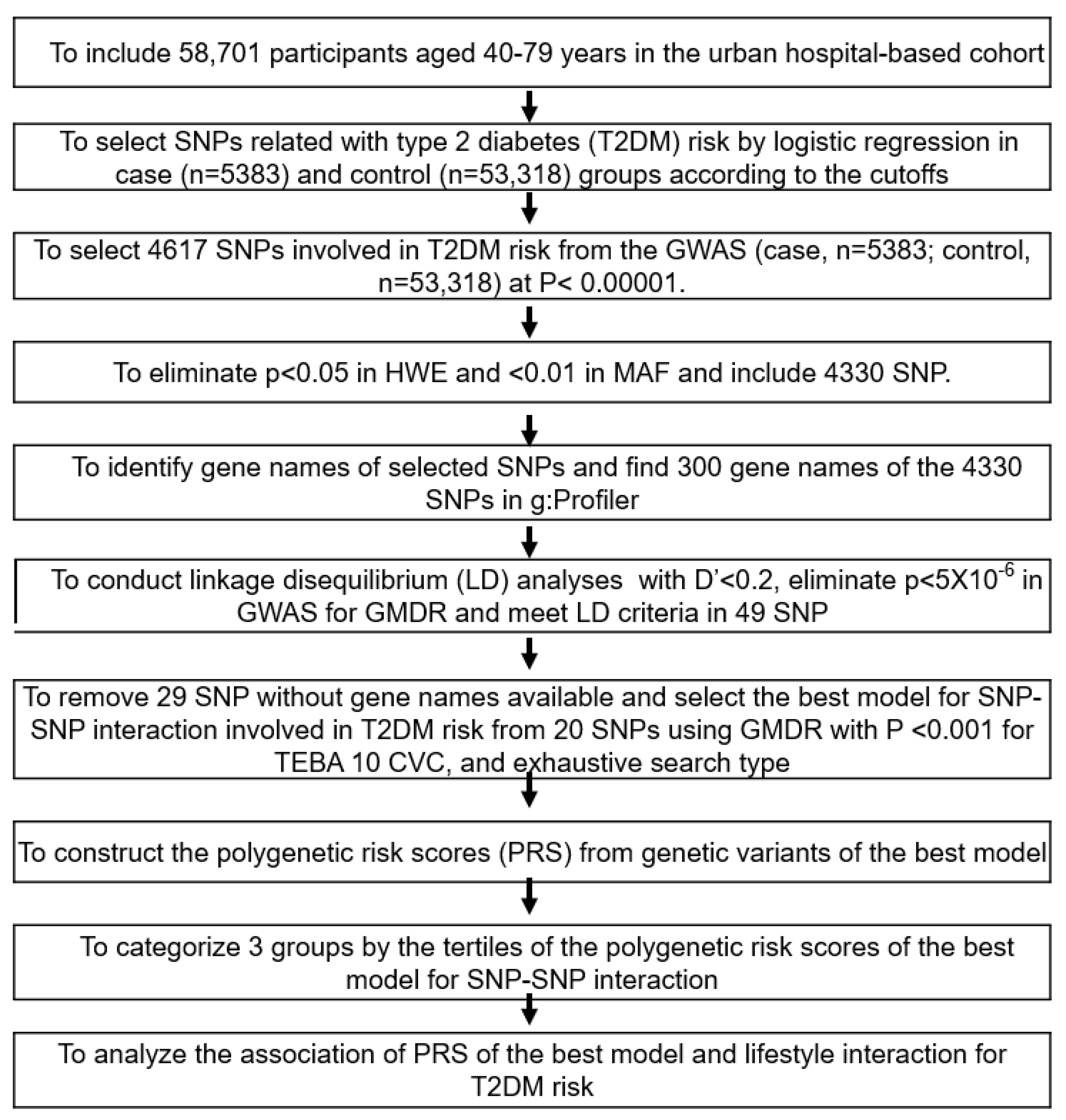
2. Materials and Methods
2.1. Subjects
2.2. Demographic, Anthropometric, and Biochemical Parameters of the Participants
2.3. T2DM Definition
2.4. Estimation of Usual Food Intake Using a Semi-Quantitative Food Frequency Questionnaire (SQFFQ)
2.5. Dietary Patterns by Principal Component Analysis (PCA)
2.6. Genotyping Using a Korean Chip and Quality Control
2.7. Genotype-Tissue Expression (GTEx) of Genetic Mutations and Distribution of Identified Tissue/Organ-Specific Expressed SNPs
2.8. Selection of the Genetic Variants That Influence the T2DM Risk and the Best Model with SNP-SNP Interactions
2.9. Statistical Analysis
3. Results
3.1. Demographic and Lifestyle Characteristics
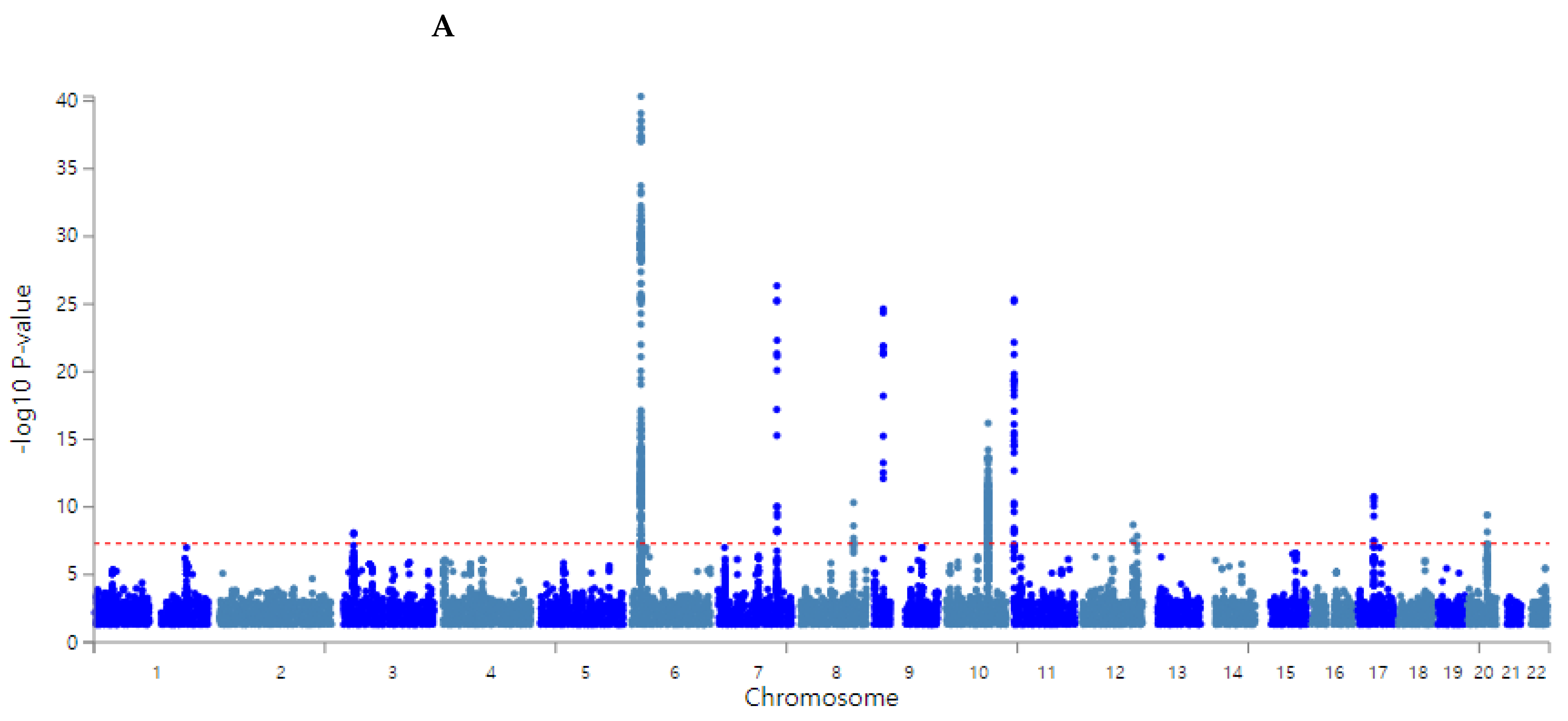
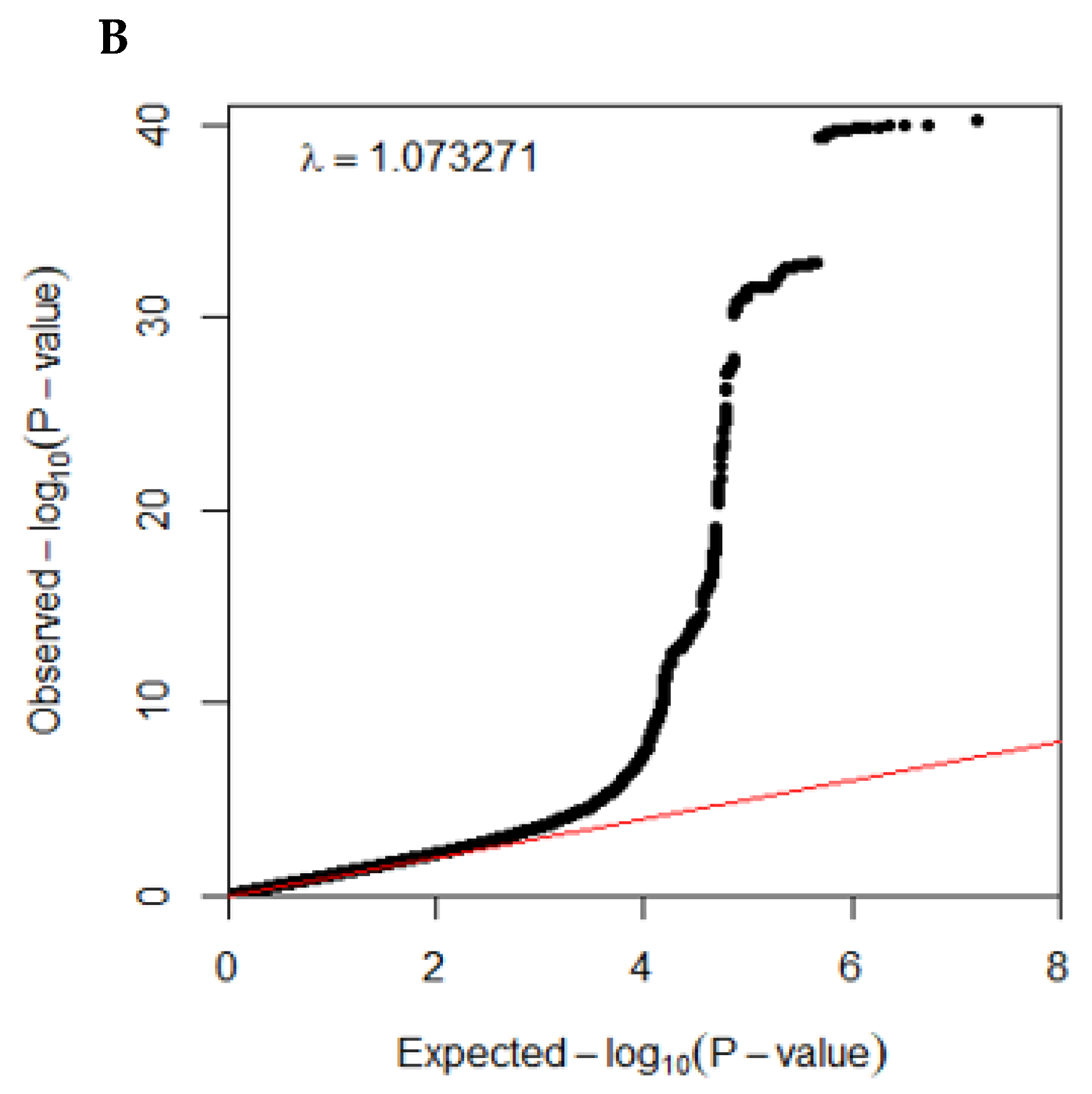
3.2. Polygenetic Variants with Their Interaction Related to the T2DM Risk
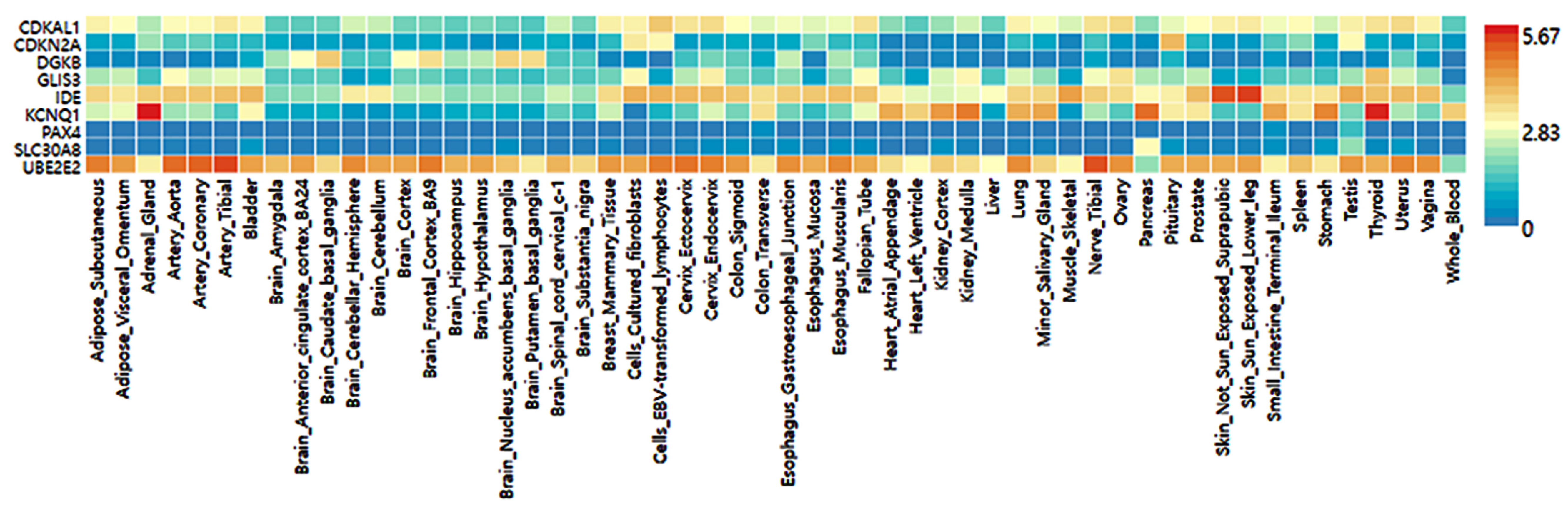
3.3. GTEx and Frequency of Tissue/organ-Specific Expression
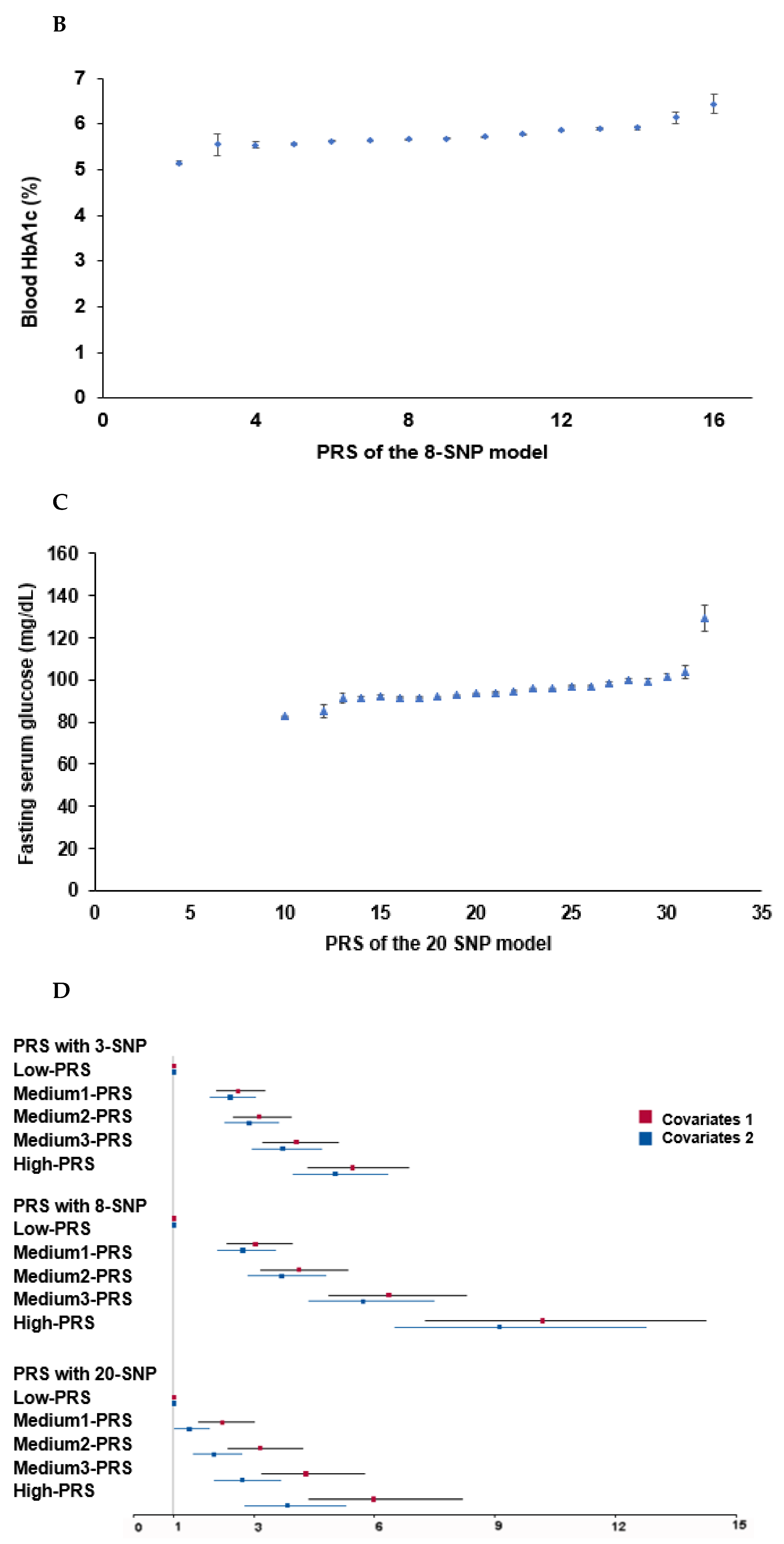
3.4. Metabolism Related to the Genetic Variants for T2DM Risk
3.5. Interaction of PRS with Lifestyle Factors to Influence T2DM Risk
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Inaishi, J.; Saisho, Y. Ethnic Similarities and Differences in the Relationship between Beta Cell Mass and Diabetes. J. Clin. Med. 2017, 6, 113. [Google Scholar] [CrossRef] [Green Version]
- Gujral, U.P.; Pradeepa, R.; Weber, M.B.; Narayan, K.V.; Mohan, V. Type 2 diabetes in South Asians: Similarities and differences with white Caucasian and other populations. Ann. N. Y. Acad. Sci. 2013, 1281, 51–63. [Google Scholar] [CrossRef]
- Loh, M.; Zhang, W.; Ng, H.K.; Schmid, K.; Lamri, A.; Tong, L.; Ahmad, M.; Lee, J.-J.; Ng, M.C.Y.; Petty, L.E.; et al. Identification of genetic effects underlying type 2 diabetes in South Asian and European populations. Commun. Biol. 2022, 5, 329. [Google Scholar] [CrossRef] [PubMed]
- Spracklen, C.N.; Horikoshi, M.; Kim, Y.J.; Lin, K.; Bragg, F.; Moon, S.; Suzuki, K.; Tam, C.H.T.; Tabara, Y.; Kwak, S.-H.; et al. Identification of Type 2 Diabetes Loci in 433,540 East Asian Individuals. Nature 2020, 582, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Mambiya, M.; Shang, M.; Wang, Y.; Li, Q.; Liu, S.; Yang, L.; Zhang, Q.; Zhang, K.; Liu, M.; Nie, F.; et al. The Play of Genes and Non-genetic Factors on Type 2 Diabetes. Front. Public Health 2019, 7, 349. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ahn, J.; Lee, B.-K. Very-Low-Fat Diets May Be Associated with Increased Risk of Metabolic Syndrome in the Adult Population. Clin. Nutr. 2016, 35, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ahn, J.; Kim, N.-S.; Lee, B.-K. High Carbohydrate Diets Are Positively Associated with the Risk of Metabolic Syndrome Irrespective to Fatty Acid Composition in Women: The KNHANES 2007–2014. Int. J. Food Sci. Nutr. 2016, 68, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kang, S. Alcohol, Carbohydrate, and Calcium Intakes and Smoking Interactions with APOA5 rs662799 and rs2266788 were Associated with Elevated Plasma Triglyceride Concentrations in a Cross-Sectional Study of Korean Adults. J. Acad. Nutr. Diet. 2020, 120, 1318–1329.e1. [Google Scholar] [CrossRef]
- Zhou, J.-Y.; Song, M.Y.; Park, S. Carbohydrate and Sodium Intake and Physical Activity Interact with Genetic Risk Scores of Four Genetic Variants Mainly Related to Lipid Metabolism to Modulate Metabolic Syndrome Risk in Korean Middle-Aged Adults. Br. J. Nutr. 2019, 122, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Park, S. A Causal Relationship between Vitamin C Intake with Hyperglycemia and Metabolic Syndrome Risk: A Two-Sample Mendelian Randomization Study. Antioxidants 2022, 11, 857. [Google Scholar] [CrossRef]
- Zheng, J.-S.; Luan, J.A.; Sofianopoulou, E.; Imamura, F.; Stewart, I.D.; Day, F.R.; Pietzner, M.; Wheeler, E.; Lotta, L.A.; Gundersen, T.E.; et al. Plasma Vitamin C and Type 2 Diabetes: Genome-Wide Association Study and Mendelian Randomization Analysis in European Populations. Diabetes Care 2020, 44, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Lee, C. Association of Glycosylated Hemoglobin with the Gene Encoding CDKAL1 in the Korean Association Resource (KARE) Study. Hum. Mutat. 2012, 33, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Daily, J.W.; Park, S. Interaction of BDNF rs6265 Variants and Energy and Protein Intake in The Risk for Glucose Intolerance and Type 2 Diabetes in Middle-Aged Adults. Nutrition 2016, 33, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Daily, J.W.; Zhang, X.; Jin, H.S.; Lee, H.J.; Lee, Y.H. Interactions with the MC4R rs17782313 Variant, Mental Stress and Energy Intake and The Risk of Obesity in Genome Epidemiology Study. Nutr. Metab. 2016, 13, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Park, S. An Inverse Relation between Hyperglycemia and Skeletal Muscle Mass Predicted by Using a Machine Learning Approach in Middle-Aged and Older Adults in Large Cohorts. J. Clin. Med. 2021, 10, 2133. [Google Scholar] [CrossRef]
- Park, S.; Kim, C.; Wu, X. Development and Validation of an Insulin Resistance Predicting Model Using a Machine-Learning Approach in a Population-Based Cohort in Korea. Diagnostics 2022, 12, 212. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Ahn, J.; Lee, B.-K. Self-Rated Subjective Health Status Is Strongly Associated with Sociodemographic Factors, Lifestyle, Nutrient Intakes, and Biochemical Indices, but Not Smoking Status: KNHANES 2007–2012. J. Korean Med Sci. 2015, 30, 1279–1287. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2021. Diabetes Care 2020, 44, S15–S33. [Google Scholar] [CrossRef]
- Park, S.; Zhang, X.; Lee, N.R.; Jin, H.-S. TRPV1 Gene Polymorphisms Are Associated with Type 2 Diabetes by Their Interaction with Fat Consumption in the Korean Genome Epidemiology Study. Lifestyle Genom. 2016, 9, 47–61. [Google Scholar] [CrossRef]
- Ahn, Y.; Kwon, E.; Shim, J.E.; Park, M.K.; Joo, Y.; Kimm, K.; Park, C.; Kim, D.H. Validation and Reproducibility of Food Fre-Quency Questionnaire for Korean Genome Epidemiologic Study. Eur. J. Clin. Nutr. 2007, 61, 1435–1441. [Google Scholar] [CrossRef]
- Steel, R.G.; Torrie, J.H.; Dickey, D.A. Principles and Procedures of Statistics: A Biological Approach; McGraw-Hill: New York, NY, USA, 1997. [Google Scholar]
- Kim, J.O.; Mueller, C.W. Factor Analysis. Statistical Methods and Practical Issues; Sage publications: Thousand Oaks, CA, USA, 1978. [Google Scholar]
- Song, Y.; Joung, H.A. Traditional Korean Dietary Pattern and Metabolic Syndrome Abnormalities. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Yang, H.J.; Kim, M.J.; Hur, H.J.; Kim, S.-H.; Kim, M.-S. Interactions between Polygenic Risk Scores, Dietary Pattern, and Menarche Age with the Obesity Risk in a Large Hospital-Based Cohort. Nutrients 2021, 13, 3772. [Google Scholar] [CrossRef]
- van Woudenbergh, G.J.; Theofylaktopoulou, D.; Kuijsten, A.; Ferreira, I.; van Greevenbroek, M.M.; van der Kallen, C.J.; Schalkwijk, C.G.; Stehouwer, C.D.; Ocké, M.C.; Nijpels, G.; et al. Adapted Dietary Inflammatory Index and Its Association with a Summary Score for Low-Grade Inflammation and Markers of Glucose Metabolism: The Cohort Study on Diabetes and Atherosclerosis Maastricht (CODAM) And the Hoorn Study. Am. J. Clin. Nutr. 2013, 98, 1533–1542. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-K.; Kim, M.K. Relationship of Sodium Intake with Obesity among Korean Children and Adolescents: Korea National Health and Nutrition Examination Survey. Br. J. Nutr. 2016, 115, 834–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Han, B.-G.; Ko, G.E.S.G. Cohort Profile: The Korean Genome and Epidemiology Study (KoGES) Consortium. Int. J. Epidemiol. 2017, 46, e20. [Google Scholar] [CrossRef]
- Rabbee, N.; Speed, T.P. A genotype calling algorithm for affymetrix SNP arrays. Bioinformatics 2005, 22, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jyothi, K.U.; Reddy, B.M. Gene–gene and gene–environment interactions in the etiology of type 2 diabetes mellitus in the population of Hyderabad, India. Meta Gene 2015, 5, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Khera, A.V.; Chaffin, M.; Wade, K.H.; Zahid, S.; Brancale, J.; Xia, R.; Distefano, M.; Senol-Cosar, O.; Haas, M.E.; Bick, A.; et al. Polygenic Prediction of Weight and Obesity Trajectories from Birth to Adulthood. Cell 2019, 177, 587–596.e589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, C.M.; Vassos, E. Polygenic Risk Scores: From Research Tools to Clinical Instruments. Genome Med. 2020, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Xue, A.; Wu, Y.; Zhu, Z.; Zhang, F.; Kemper, K.E.; Zheng, Z.; Yengo, L.; Lloyd-Jones, L.R.; Sidorenko, J.; Wu, Y.; et al. Genome-wide association analyses identify 143 risk variants and putative regulatory mechanisms for type 2 diabetes. Nat. Commun. 2018, 9, 2941. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Akiyama, M.; Ishigaki, K.; Kanai, M.; Hosoe, J.; Shojima, N.; Hozawa, A.; Kadota, A.; Kuriki, K.; Naito, M.; et al. Identification of 28 new susceptibility loci for type 2 diabetes in the Japanese population. Nat. Genet. 2019, 51, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, C.C. Genetic Variants in Promoter Regions Associated with Type 2 Diabetes Mellitus: A Large-Scale Meta-Analysis and Subgroup Analysis. J. Cell Biochem. 2019, 120, 13012–13025. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.C.; Park, K.S.; Oh, B.; Tam, C.H.; Cho, Y.M.; Shin, H.D.; Lam, V.K.; Ma, R.C.; So, W.Y.; Cho, Y.S.; et al. Implication of Genetic Variants Near TCF7L2, SLC30A8, HHEX, CDKAL1, CDKN2A/B, IGF2BP2, and FTO in Type 2 Diabetes and Obesity in 6719 Asians. Diabetes 2008, 57, 2226–2233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, K.-W.; Kim, S.H.; Zhang, X.; Park, S. Interactions among The Variants of Insulin-Related Genes and Nutrients Increase the Risk of Type 2 diabetes. Nutr. Res. 2018, 51, 82–92. [Google Scholar] [CrossRef]
- Aarthy, R.; Aston-Mourney, K.; Mikocka-Walus, A.; Radha, V.; Amutha, A.; Anjana, R.M.; Unnikrishnan, R.; Mohan, V. Clinical features, complications and treatment of rarer forms of maturity-onset diabetes of the young (MODY)—A review. J. Diabetes Its Complicat. 2020, 35, 107640. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association Professional Practice Committee. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes—2022. Diabetes Care 2021, 45, S17–S38. [Google Scholar] [CrossRef]
- Firdous, P.; Nissar, K.; Ali, S.; Ganai, B.A.; Shabir, U.; Hassan, T.; Masoodi, S.R. Genetic Testing of Maturity-Onset Diabetes of the Young Current Status and Future Perspectives. Front. Endocrinol. 2018, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, A.M.; Gannon, M. Molecular Regulation of Pancreatic Beta-Cell Mass Development, Maintenance, and Expansion. J. Mol. Endocrinol. 2007, 38, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Asahara, S.-I.; Inoue, H.; Kido, Y. Regulation of Pancreatic β-Cell Mass by Gene-Environment Interaction. Diabetes Metab. J. 2022, 46, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Asahara, S.-I.; Etoh, H.; Inoue, H.; Teruyama, K.; Shibutani, Y.; Ihara, Y.; Kawada, Y.; Bartolome, A.; Hashimoto, N.; Matsuda, T.; et al. Paternal allelic mutation at the Kcnq1 locus reduces pancreatic β-cell mass by epigenetic modification of Cdkn1c. Proc. Natl. Acad. Sci. USA 2015, 112, 8332–8337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flannick, J.; Thorleifsson, G.; Beer, N.L.; Jacobs, S.B.R.; Grarup, N.; Burtt, N.P.; Mahajan, A.; Fuchsberger, C.; Atzmon, G.; Benediktsson, R.; et al. Loss-of-Function Mutations in SLC30A8 Protect Against Type 2 Diabetes. Nat. Genet. 2014, 46, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.J.; Bruckner, R.J.; Paulo, J.A.; Kazak, L.; Long, J.Z.; Mina, A.I.; Deng, Z.; LeClair, K.B.; Hall, J.A.; Hong, S.; et al. Cdkal1, a Type 2 Diabetes Susceptibility Gene, Regulates Mitochondrial Function in Adipose Tissue. Mol. Metab. 2017, 6, 1212–1225. [Google Scholar] [CrossRef] [PubMed]
- Steinthorsdottir, V.; Thorleifsson, G.; Reynisdóttir, I.; Benediktsson, R.; Jonsdottir, T.; Walters, G.B.; Styrkarsdottir, U.; Gretarsdottir, S.; Emilsson, V.; Ghosh, S.; et al. A Variant in CDKAL1 Influences Insulin Response and Risk of Type 2 Diabetes. Nat. Genet. 2007, 39, 770–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadheel, H.K.; Kaftan, A.N.; Naser, F.H.; Hussain, M.K.; Algenabi, A.H.A.; Mohammad, H.J.; Al-Kashwan, T.A. Association of CDKN2A/B gene polymorphisms (rs10811661 and rs2383208) with type 2 diabetes mellitus in a sample of Iraqi population. Egypt. J. Med. Hum. Genet. 2022, 23, 1–7. [Google Scholar] [CrossRef]
- Plengvidhya, N.; Chanprasert, C.; Chongjaroen, N.; Yenchitsomanus, P.-T.; Homsanit, M.; Tangjittipokin, W. Impact of KCNQ1, CDKN2A/2B, CDKAL1, HHEX, MTNR1B, SLC30A8, TCF7L2, and UBE2E2 on risk of developing type 2 diabetes in Thai population. BMC Med. Genet. 2018, 19, 93. [Google Scholar] [CrossRef]
- Wen, J.; Rönn, T.; Olsson, A.; Yang, Z.; Lu, B.; Du, Y.; Groop, L.; Ling, C.; Hu, R. Investigation of Type 2 Diabetes Risk Alleles Support CDKN2A/B, CDKAL1, and TCF7L2 As Susceptibility Genes in a Han Chinese Cohort. PLoS ONE 2010, 5, e9153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, S.; Jacobs, S.; Zheng, J.; Meidtner, K.; Schwingshackl, L.; Schulze, M.B. Gene-lifestyle interaction on risk of type 2 diabetes: A systematic review. Obes. Rev. 2019, 20, 1557–1571. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Kim, B.C.; Daily, J.W.; Park, S. High genetic risk scores for impaired insulin secretory capacity doubles the risk for type 2 diabetes in Asians and is exacerbated by Western-type diets. Diabetes/Metabolism Res. Rev. 2017, 34, e2944. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Park, C.H.; Jang, J.S. Antecedent Intake of Traditional Asian-Style Diets Exacerbates Pancreatic Beta-Cell Function, Growth and Survival after Western-Style Diet Feeding in Weaning Male Rats. J. Nutr. Biochem. 2006, 17, 307–318. [Google Scholar] [CrossRef]
- Frassetto, L.; Banerjee, T.; Powe, N.; Sebastian, A. Acid Balance, Dietary Acid Load, and Bone Effects-A Controversial Subject. Nutrients 2018, 10, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Moon, S.H.; Kim, H.J. The Korea Biobank Array Project. Public Health Wkly. Rep. Kcdc 2015, 8, 670–674. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Men (n = 20,293) | Women (n = 38,408) | Adjusted ORs and 95% CI | |||
|---|---|---|---|---|---|
| Control (n = 17,656) | T2M (n = 2637) | Control (n = 35,662) | T2DM (n = 2746) | ||
| Age (years) | 55.7 ± 0.06 b | 58.7 ± 0.15 b | 52.3 ± 0.04 c | 55.5 ± 0.14 b *** +++ | 1.875 (1.740–2.022) |
| Education | |||||
| ≤Middle school | 1467 (13.7) | 286 (16.1) ‡‡ | 5887 (21.1) | 851 (34.8) ‡‡‡ | 1 |
| High school | 8103 (75.7) | 1331 (75.1) | 20,360 (72.9) | 1511 (61.8) | 0.723(0.664–0.786) |
| ≥College | 1136 (10.6) | 155 (8.75) | 1698 (6.08) | 84 (3.43) | 0.646 (0.550–0.760) |
| Income | |||||
| ≤USD 2000 | 1338 (7.96) | 268 (10.7) ‡‡‡ | 3677 (11.0) | 495 (19.1) ‡‡‡ | 1 |
| USD 2000–4000 | 7082 (42.1) | 1125 (44.9) | 14,700(43.8) | 1285 (49.7) | 0.846 (0.770–0.928) |
| >USD 4000 | 8389 (49.9) | 1111 (44.4) | 15,163(45.2) | 807 (31.2) | 0.748 (0.676–0.827) |
| BMI (kg/m2) | 24.3 ± 0.02 b | 25.0 ± 0.06 a | 23.5 ± 0.02 c | 25.0 ± 0.06 a *** +++ ### | 1.692 (1.557–1.838) |
| Waist circumferences (cm) | 84.2 ± 0.04 b | 85.2 ± 0.10 a | 78.7 ± 0.03 d | 79.8 ± 0.09 c *** +++ | 1.916 (1.752–2.095) |
| SMI (kg/m2) | 7.33 ± 0.01 a | 7.20 ± 0.01 b | 6.22 ± 0.01 c | 6.06 ± 0.004 d *** +++ ### | 0.768 (0.638–0.924) |
| Fat mass (%) | 23.1 ± 0.03 d | 24.0 ± 0.07 c | 31.0 ± 0.02 b | 32.6 ± 0.07 a *** +++ ### | 1.578 (1.429–1.742) |
| Serum glucose (mg/dL) | 93.3 ± 0.17 c | 133.8 ± 0.40 a | 90.3 ± 0.12 d | 129.1 ± 0.42 b *** +++ ### | |
| HbA1c (%) | 5.52 ± 0.01 b | 7.04 ± 0.02 a | 5.56 ± 0.01 b | 7.08 ± 0.02 a ** +++ | |
| Insulin resistance (%) | 680 (3.65) | 1632 (61.9) ‡‡‡ | 791 (2.22) | 1501 (54.7) ‡‡‡ | 58.81 (51.59–67.04) |
| Men (n = 20,293) | Women (n = 38,408) | Adjusted ORs and 95% CI | |||
|---|---|---|---|---|---|
| Control (n = 17,656) | T2DM (n = 2637) | Control (n = 35,662) | T2DM (n = 2746) | ||
| Energy intake (EER %) 1 | 86.0 ± 0.06 1 b | 85.4 ± 0.13 c | 104 ± 0.05 a | 1040 ± 0.09 a *** ### | 0.958 (0.767–1.196) |
| CHO (En%) 2 | 71.6 ± 0.08 | 71.3 ± 0.17 | 71.7 ± 0.06 | 71.6 ± 0.12 | 0.997 (0.932–1.067) |
| Fat (En%) 3 | 13.9 ± 0.06 a b | 14.2 ± 0.12 a b | 13.9 ± 0.04 b | 14.1 ± 0.09 a ## | 0.987 (0.893–1.091) |
| Protein (En%) 4 | 13.3 ± 0.03 c | 13.4 ± 0.05 b | 13.6 ± 0.02 a | 13.6 ± 0.04 a *** + # | 1.054 (0.978–1.136) |
| Fiber (g) 5 | 14.3 ± 0.07 a | 14.3 ± 0.12 a | 14.8 ± 0.07 b | 14.9 ± 0.08 b *** | 0.740 (0.266–2.054) |
| Calcium (mg) 6 | 383 ± 1.61 c | 385 ± 4.00 c | 475 ± 1.11 a | 463 ± 3.91 b *** # | 0.985 (0.906–1.071) |
| Vitamin C (mg) 7 | 89.3 ± 0.47 c | 89.0 ± 1.16 c | 114.2 ± 0.32 a | 110 ± 1.13 b *** ++ # | 0.918 (0.857–0.983) |
| Vitamin D (ug) 8 | 5.24 ± 0.04 c | 4.98 ± 0.08 d | 7.18 ± 0.04 a | 6.95 ± 0.06 b *** ++ | 0.967 (0.871–1.075) |
| DII (scores) 9 | −18.3 ± 0.12 c | −18.5 ± 0.29 c | −21.6 ± 0.28 a | −20.7 ± 0.08 b *** ++ | 1.109 (1.028–1.195) |
| Flavonoids (mg) 10 | 30.0 ± 0.24 c | 29.9 ± 0.60 d | 43.1 ± 0.17 a | 40.0 ± 0.59 b *** +++ | 0.891 (0.809–0.981) |
| KBD (%) 11 | 6671 (39.6) | 1430(41.7) ‡ | 9165 (30.1) | 2303(28.8) ‡ | 0.958 (0.910–1.008) |
| PBD (%) 11 | 3488 (20.7) | 710 (20.7) | 12,346 (40.6) | 3032(37.9 ‡‡‡ | 0.890 (0.827–0.958) |
| WSD (%) 11 | 8489 (50.4) | 1933 (56.3) ‡‡‡ | 10,333 (34.0) | 2788(34.8) | 1.269 (1.207–1.335) |
| RMD (%) 11 | 5376 (31.9) | 1089(31.7) | 10,370 (34.1) | 2736(34.2) | 1.018 (0.970–1.068) |
| Alcohol (g) 12 | 35.7 ± 0.38 a | 36.6 ± 0.94 a | 5.37 ± 0.26 b | 4.96 ± 0.91 b *** | 0.878 (0.818–0.942) |
| Exercise (%) 13 | 10,323 (58.7) | 1629 (61.9) ‡‡ | 18,537 (52.2) | 1487 (54.3) ‡ | 1.143 (1.073–1.217) |
| Non-smoking (%) | 5150 (29.2) | 629 (23.9) ‡‡‡ | 34,442 (96.9) | 2618 (95.7) ‡‡‡ | 1 |
| Former smoking | 7541 (42.8) | 1254 (47.7) | 427 (1.2) | 33 (1.21) | 1.289 (1.163–1.430) |
| Smoking | 4919 (27.9) | 745 (28.4) | 664 (1.87) | 85 (3.11) | 1.602 (1.431–1.792) |
| Chr 1 | SNP 2 | Position | Mi 3 | Ma 4 | OR and 95% CI for City 5 | p Value Adjusted 6 | p Value Adjusted 7 | MAF 8 | p Value for HWE 9 | Gene | Functional Consequence |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 3 | rs7631705 | 23632234 | C | T | 0.888 | 8.00 × 10−9 | 0.0395 | 0.3341 | 0.5715 | UBE2E2 | 3_prime_utr |
| 6 | rs35612982 | 20682622 | C | T | 1.342 | 9.35 × 10−39 | 1.27 × 10−8 | 0.4649 | 0.1131 | CDKAL1 | Intron |
| 7 | rs2191349 | 15064309 | G | T | 0.891 | 2.91 × 10−7 | 0.0175 | 0.3233 | 0.04656 | DGKB | Intron |
| 7 | rs61160304 | 127249659 | T | C | 1.492 | 6.34 × 10−26 | 2.11 × 10−7 | 0.0738 | 0.2274 | PAX4 | Downstream |
| 8 | rs13266634 | 118184783 | T | C | 0.853 | 8.22 × 10−12 | 0.00255 | 0.398 | 0.9656 | SLC30A8 | Missense |
| 9 | rs7034200 | 4289050 | A | C | 1.113 | 2.05 × 10−7 | 0.0150 | 0.4064 | 0.3467 | GLIS3 | Nmd transcript |
| 9 | rs10811661 | 22134094 | C | T | 0.797 | 6.33 × 10−24 | 1.08 × 10−6 | 0.4387 | 0.1998 | CDKN2A/B | Non-coding transcript |
| 10 | rs12764758 | 94516663 | T | C | 1.285 | 5.00 × 10−10 | 0.0304 | 0.0586 | 0.3958 | IDE | Intron |
| 11 | rs60808706 | 2857233 | A | G | 0.787 | 6.65 × 10−25 | 8.17 × 10−7 | 0.3913 | 0.2251 | KCNQ1 | Downstream |
| 17 | rs11651052 | 36102381 | A | G | 1.157 | 5.17 × 10−10 | 0.008245 | 0.3009 | 0.4383 | HNF1B | Intron |
| Covariates | Adjusted for Age, Gender, BMI, Education, Income, Income, Area | Adjusted for Age, Gender, BMI, Education, Income, Income, Area, Smoke, Exercise, Alcohol, and Energy Intake | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Models | TRBA | TEBA | p Value | CVC | TRBA | TEBA | p Value | CVC | ||
| CDKAL1_rs35612982 | 0.5411 | 0.5412 | 0.001 | 10 | 0.5411 | 0.5412 | 0.001 | 10 | ||
| CDKN2A/B_rs10811661 plus model 1 | 0.5529 | 0.5493 | 0.001 | 10 | 0.5529 | 0.5493 | 0.001 | 10 | ||
| KCNQ1_rs60808706 plus model 2 | 0.5599 | 0.5589 | 0.001 | 10 | 0.5599 | 0.5589 | 0.001 | 10 | ||
| GLIS3_rs7034200 plus model 3 | 0.5665 | 0.5578 | 0.001 | 9 | 0.5665 | 0.5578 | 0.001 | 9 | ||
| UBE2E2_rs7631705 plus model 4 | 0.5778 | 0.5552 | 0.001 | 8 | 0.5778 | 0.5552 | 0.001 | 8 | ||
| HNF1B_rs11651052 plus model 5 | 0.5985 | 0.5393 | 0.001 | 7 | 0.5985 | 0.5393 | 0.001 | 7 | ||
| SLC30A8_rs13266634 plus model 6 | 0.6395 | 0.5294 | 0.001 | 7 | 0.6395 | 0.5294 | 0.001 | 7 | ||
| PAX4_rs61160304 plus model 7 | 0.706 | 0.5208 | 0.001 | 10 | 0.706 | 0.5208 | 0.001 | 10 | ||
| IDE_rs12764758 plus model 8 | 0.7644 | 0.5215 | 0.001 | 10 | 0.7644 | 0.5215 | 0.001 | 10 | ||
| DGKB_rs2191349 plus model 9 | 0.812 | 0.5218 | 0.001 | 10 | 0.812 | 0.5218 | 0.001 | 10 | ||
| Pathways | No. of Genes 1 | Beta 2 | SD 3 | p Value 4 | p Value Bonferroni 5 | Participating Genes |
|---|---|---|---|---|---|---|
| Regulation of gene expression in endocrine committed neurog3plus progenitor cells | 2 | 1.677 | 0.0229 | 2.92 × 10−15 | 4.45 × 10−11 | PAX6, HNF1α, HNF1β |
| Maturity onset diabetes of the young | 16 | 0.6309 | 0.0243 | 3.88 × 10−13 | 5.92 × 10−09 | PDX1, HNF1β, HNF1a, HNF4α, NeuroD1 |
| Regulation of β-cell development | 24 | 0.4618 | 0.0218 | 8.64 × 10−10 | 1.32 × 10−05 | HNF1β, FGF10, ONECUT3, HNF6, PDX1 |
| Pancreatic endocrine progenitor | 6 | 0.8761 | 0.0207 | 1.12 × 10−09 | 1.72 × 10−05 | |
| Negative regulation of hormone secretion | 34 | 0.3263 | 0.0183 | 6.46 × 10−08 | 9.85 × 10−04 | |
| Negative regulation of insulin secretion | 22 | 0.3953 | 0.0179 | 1.58 × 10−07 | 0.0024 | ADR2α, CRHR2, KLF7, PDE1c, UCP2 |
| Low-PRS (n = 14,420) | Medium-PRS (n = 21,641) | High-PRS (n = 4201) | Gene-Nutrient Interaction p Value | |
|---|---|---|---|---|
| Low energy 1 High energy | 1 | 1.771 (1.505–2.084) 2.237 (1.661–3.012) | 2.960 (2.503–3.502) 3.592 (2.650 4.870 | 0.0002 |
| Low KBD 2 High KBD | 1 | 1.857 (1.611–2.141) 1.961 (1.636–2.351) | 3.005 (2.597–3.477) 3.220 (2.674 3.878) | 0.6555 |
| Low PBD 2 High PBD | 1 | 1.891 (1.638–2.183) 1.917 (1.621–2.266) | 2.903 (2.453–3.508) 3.130 (2.700 3.627) | 0.3048 |
| Low WSD 2 High WSD | 1 | 1.891 (1.638–2.183) 1.963 (1.632–2.361) | 3.130 (2.700–3.627) 3.329 (2.754 4.023) | 0.0347 |
| Low RMD 2 High RMD | 1 | 1.891 (1.638–2.183) 1.995 (1.677–2.374) | 3.130 (2.700–3.627) 3.221 (2.694 3.850) | 0.0715 |
| Low alcohol 3 High alcohol | 1 | 1.867 (1.548–2.253) 1.937 (1.550–2.421) | 3.048 (2.513–3.697) 3.254 (2.588 4.093) | 0.3049 |
| Low exercise 4 High exercise | 1 | 2.229 (1.758–2.827) 1.711 (1.428–2.049) | 3.667 (2.874–4.679) 2.845 (2.363 3.426) | 0.4115 |
| Non-smoking + former smoking Smoking | 1 | 1.835 (1.580–2.131) 3.021 (1.979–4.611) | 3.068 (2.631–3.577) 4.889 (3.154– 7.576) | 0.0006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hur, H.J.; Yang, H.J.; Kim, M.J.; Lee, K.-H.; Kim, M.-S.; Park, S. Association of Polygenic Variants with Type 2 Diabetes Risk and Their Interaction with Lifestyles in Asians. Nutrients 2022, 14, 3222. https://doi.org/10.3390/nu14153222
Hur HJ, Yang HJ, Kim MJ, Lee K-H, Kim M-S, Park S. Association of Polygenic Variants with Type 2 Diabetes Risk and Their Interaction with Lifestyles in Asians. Nutrients. 2022; 14(15):3222. https://doi.org/10.3390/nu14153222
Chicago/Turabian StyleHur, Haeng Jeon, Hye Jeong Yang, Min Jung Kim, Kyun-Hee Lee, Myung-Sunny Kim, and Sunmin Park. 2022. "Association of Polygenic Variants with Type 2 Diabetes Risk and Their Interaction with Lifestyles in Asians" Nutrients 14, no. 15: 3222. https://doi.org/10.3390/nu14153222
APA StyleHur, H. J., Yang, H. J., Kim, M. J., Lee, K.-H., Kim, M.-S., & Park, S. (2022). Association of Polygenic Variants with Type 2 Diabetes Risk and Their Interaction with Lifestyles in Asians. Nutrients, 14(15), 3222. https://doi.org/10.3390/nu14153222