
Adherence to MIND Diet, Genetic Susceptibility, and Incident Dementia in Three US Cohorts

 ,
, 
Abstract
1. Introduction
2. Methods
2.1. Study Populations
2.2. Diet Assessment and MIND Adherence Score
2.3. Cognitive Assessments
2.4. Dementia Outcomes
2.5. Genetic Data
2.6. Other Covariates
2.7. Final Analytical Samples
2.8. SNP-Selection and Calculation of Genetic Susceptibility Scores (GS)
2.9. Statistical Analysis
3. Results
3.1. Baseline Characteristics of Study Populations
3.2. Main Effects of MIND Adherence and Genetic Factors on Incident Dementia and Cognitive Decline
3.3. Gene × MIND Interactions for Incident Dementia and Cognitive Decline
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Association. Trajectory Report. Available online: https://www.alz.org/help-support/resources/publications/trajectory_report (accessed on 1 September 2020).
- Aridi, Y.; Walker, J.; Wright, O. The association between the Mediterranean dietary pattern and cognitive health: A systematic review. Nutrients 2017, 9, 674. [Google Scholar] [CrossRef]
- Morris, M.C. Nutritional determinants of cognitive aging and dementia. Proc. Nutr. Soc. 2012, 71, 1–13. [Google Scholar] [CrossRef]
- Jansen, I.; Savage, J.; Watanabe, K.; Bryois, J.; Williams, D.; Steinberg, S.; Sealock, J.; Karlsson, I.; Hägg, S.; Athanasiu, L. Genome-wide meta-analysis identifies new loci and functional pathways influencing Alzheimer’s disease risk. Nat. Genet. 2019, 51, 404–413. [Google Scholar] [CrossRef]
- Desikan, R.S.; Schork, A.J.; Wang, Y.; Thompson, W.K.; Dehghan, A.; Ridker, P.M.; Chasman, D.I.; McEvoy, L.K.; Holland, D.; Chen, C.-H.; et al. Polygenic overlap between c-reactive protein, plasma lipids, and Alzheimer disease. Circulation 2015, 131, 2061–2069. [Google Scholar] [CrossRef][Green Version]
- Chen, J.; Xie, C.; Zhao, Y.; Li, Z.; Xu, P.; Yao, L. Gene expression analysis reveals the dysregulation of immune and metabolic pathways in Alzheimer’s disease. Oncotarget 2016, 7, 72469–72474. [Google Scholar] [CrossRef]
- Sureda, A.; Bibiloni, M.D.M.; Julibert, A.; Bouzas, C.; Argelich, E.; Llompart, I.; Pons, A.; Tur, J.A. Adherence to the Mediterranean diet and inflammatory markers. Nutrients 2018, 10, 62. [Google Scholar] [CrossRef]
- Grosso, G.; Mistretta, A.; Frigiola, A.; Gruttadauria, S.; Biondi, A.; Basile, F.; Vitaglione, P.; D’Orazio, N.; Galvano, F. Mediterranean diet and cardiovascular risk factors: A systematic review. Crit. Rev. Food Sci. Nutr. 2014, 54, 593–610. [Google Scholar] [CrossRef]
- Toledo, E.; Wang, D.D.; Ruiz-Canela, M.; Clish, C.B.; Razquin, C.; Zheng, Y.; Guasch-Ferre, M.; Hruby, A.; Corella, D.; Gomez-Gracia, E.; et al. Plasma lipidomic profiles and cardiovascular events in a randomized intervention trial with the Mediterranean diet. Am. J. Clin. Nutr. 2017, 106, 973–983. [Google Scholar]
- Nishida, Y.; Ito, S.; Ohtsuki, S.; Yamamoto, N.; Takahashi, T.; Iwata, N.; Jishage, K.-I.; Yamada, H.; Sasaguri, H.; Yokota, S. Depletion of vitamin E increases amyloid beta accumulation by decreasing its clearances from brain and blood in a mouse model of Alzheimer disease. J. Biol. Chem. 2009, 284, 33400–33408. [Google Scholar] [CrossRef]
- Calon, F.; Lim, G.P.; Yang, F.; Morihara, T.; Teter, B.; Ubeda, O.; Rostaing, P.; Triller, A.; Salem, N.; Ashe, K.H.; et al. Docosahexaenoic acid protects from dendritic pathology in an Alzheimer’s disease mouse model. Neuron 2004, 43, 633–645. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative stress and inflammation: What polyphenols can do for us? Oxidative Med. Cell. Longev. 2016, 2016, 9. [Google Scholar] [CrossRef]
- Lim, S.Y.; Suzuki, H. Effect of dietary docosahexaenoic acid and phosphatidylcholine on maze behavior and fatty acid composition of plasma and brain lipids in mice. Int. J. Vitam. Nutr. Res. 2000, 70, 251–259. [Google Scholar] [CrossRef]
- Evans, D.A.; Bennett, D.A.; Wilson, R.S.; Bienias, J.L.; Morris, M.C.; Scherr, P.A.; Hebert, L.E.; Aggarwal, N.; Beckett, L.A.; Joglekar, R.; et al. Incidence of Alzheimer disease in a biracial urban community: Relation to apolipoprotein e allele status. Arch. Neurol. 2003, 60, 185–189. [Google Scholar] [CrossRef]
- Rajan, K.B.; Weuve, J.; Barnes, L.L.; Wilson, R.S.; Evans, D.A. Prevalence and incidence of clinically diagnosed Alzheimer’s disease dementia from 1994 to 2012 in a population study. Alzheimer’s Dement. 2019, 15, 1–7. [Google Scholar] [CrossRef]
- Bennett, D.A.; Schneider, J.A.; Buchman, A.S.; Barnes, L.L.; Boyle, P.A.; Wilson, R.S. Overview and findings from the rush memory and aging project. Curr. Alzheimer Res. 2012, 9, 646–663. [Google Scholar] [CrossRef]
- Shumaker, S.A.; Legault, C.; Rapp, S.R.; Thal, L.; Wallace, R.B.; Ockene, J.K.; Hendrix, S.L.; Jones, B.N., III; Assaf, A.R.; Jackson, R.D.; et al. Estrogen plus progestin and the incidence of dementia and mild cognitive impairment in postmenopausal women: The women’s health initiative memory study: A randomized controlled trial. JAMA 2003, 289, 2651–2662. [Google Scholar] [CrossRef]
- Anderson, G.L.; Limacher, M. Effects of conjugated equine estrogen in postmenopausal women with hysterectomy: The women’s health initiative randomized controlled trial. J. Am. Med. Assoc. 2004, 291, 1701–1712. [Google Scholar]
- Rapp, S.R.; Espeland, M.A.; Shumaker, S.A.; Henderson, V.W.; Brunner, R.L.; Manson, J.E.; Gass, M.L.; Stefanick, M.L.; Lane, D.S.; Hays, J.; et al. Effect of estrogen plus progestin on global cognitive function in postmenopausal women: The women’s health initiative memory study: A randomized controlled trial. JAMA 2003, 289, 2663–2672. [Google Scholar] [CrossRef]
- Rossouw, J.E.; Anderson, G.L.; Prentice, R.L.; LaCroix, A.Z.; Kooperberg, C.; Stefanick, M.L.; Jackson, R.D.; Beresford, S.A.A.; Howard, B.V.; Johnson, K.C.; et al. Risks and benefits of estrogen plus progestin in healthy postmenopausal women: Principal results from the women’s health initiative randomized controlled trial. J. Am. Med. Assoc. 2002, 288, 321–333. [Google Scholar]
- Espeland, M.A.; Rapp, S.R.; Shumaker, S.A.; Brunner, R.; Manson, J.E.; Sherwin, B.B.; Hsia, J.; Margolis, K.L.; Hogan, P.E.; Wallace, R.; et al. Conjugated equine estrogens and global cognitive function in postmenopausal women: Women’s health initiative memory study. JAMA 2004, 291, 2959–2968. [Google Scholar] [CrossRef]
- Shumaker, S.A.; Legault, C.; Kuller, L.; Brunner, R.; Manson, J.E.; Sherwin, B.B.; Hsia, J.; Margolis, K.L.; Hogan, P.E.; Wallace, R.; et al. Conjugated equine estrogens and incidence of probable dementia and mild cognitive impairment in postmenopausal women: Women’s health initiative memory study. JAMA 2004, 291, 2947–2958. [Google Scholar] [CrossRef]
- Morris, M.C.; Tangney, C.C.; Wang, Y.; Sacks, F.M.; Bennett, D.A.; Aggarwal, N.T. Mind diet associated with reduced incidence of Alzheimer’s disease. Alzheimer’s Dement. 2015, 11, 1007–1014. [Google Scholar] [CrossRef]
- Teng, E.L.; Chui, H.C. The modified mini-mental state (3ms) examination. J. Clin. Psychiatry 1987, 48, 314–318. [Google Scholar]
- Rapp, S.R.; Legault, C.; Espeland, M.A.; Resnick, S.M.; Hogan, P.E.; Coker, L.H.; Dailey, M.; Shumaker, S.A. Validation of a cognitive assessment battery administered over the telephone. J. Am. Geriatr. Soc. 2012, 60, 1616–1623. [Google Scholar] [CrossRef]
- Ellis, R.J.; Jan, K.; Kawas, C.; Koller, W.C.; Lyons, K.E.; Jeste, D.V.; Hansen, L.A.; Thal, L.J. Diagnostic validity of the dementia questionnaire for Alzheimer disease. Arch. Neurol. 1998, 55, 360–365. [Google Scholar] [CrossRef]
- Bienias, J.L.; Beckett, L.A.; Bennett, D.A.; Wilson, R.S.; Evans, D.A. Design of the Chicago health and aging project (CHAP). J. Alzheimers Dis. 2003, 5, 349–355. [Google Scholar] [CrossRef]
- Bienias, J.L.; Kott, P.S.; Evans, D.A. Applying the delete-a-group jackknife variance estimator to analyses of data from a complex longitudinal survey. In Proceedings of the Annual Meeting of the American Statistical Association, Section on Survey Research Methods; American Statistical Association: Alexandria, VA, USA, 2003; pp. 539–544. [Google Scholar]
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34, 939–944. [Google Scholar] [CrossRef]
- Bennett, D.A.; Schneider, J.A.; Aggarwal, N.T.; Arvanitakis, Z.; Shah, R.C.; Kelly, J.F.; Fox, J.H.; Cochran, E.J.; Arends, D.; Treinkman, A.D.; et al. Decision rules guiding the clinical diagnosis of Alzheimer’s disease in two community-based cohort studies compared to standard practice in a clinic-based cohort study. Neuroepidemiology 2006, 27, 169–176. [Google Scholar] [CrossRef]
- De Jager, P.L.; Shulman, J.M.; Chibnik, L.B.; Keenan, B.T.; Raj, T.; Wilson, R.S.; Yu, L.; Leurgans, S.E.; Tran, D.; Aubin, C.; et al. A genome-wide scan for common variants affecting the rate of age-related cognitive decline. Neurobiol. Aging 2012, 33, 1017.e1–1017.e15. [Google Scholar] [CrossRef]
- Dumitrescu, L.; Mahoney, E.R.; Mukherjee, S.; Lee, M.L.; Bush, W.S.; Engelman, C.D.; Lu, Q.; Fardo, D.W.; Trittschuh, E.H.; Mez, J.; et al. Genetic variants and functional pathways associated with resilience to Alzheimer’s disease. Brain 2020, 143, 2561–2575. [Google Scholar] [CrossRef]
- Hayden, K.M.; Wang, Y.; Beavers, D.; Chen, J.-C.; Espeland, M.A.; Ford, C.N.; Harrington, L.B.; He, K.; Jensen, M.K.; Johnson, K.C.; et al. The mind diet and incident dementia: Findings from the women’s health initiative memory study. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2017, 13, P858. [Google Scholar] [CrossRef]
- Chouraki, V.; Reitz, C.; Maury, F.; Bis, J.C.; Bellenguez, C.; Yu, L.; Jakobsdottir, J.; Mukherjee, S.; Adams, H.H.; Choi, S.H.; et al. Evaluation of a genetic risk score to improve risk prediction for Alzheimer’s disease. J. Alzheimer’s Dis. 2016, 53, 921–932. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, I.; Snively, B.M.; Espeland, M.A.; Shumaker, S.A.; Rapp, S.R.; Goveas, J.S.; Casanova, R.L.; Wactawski-Wende, J.; Manson, J.E.; Rossom, R. A candidate gene study of risk for dementia in older, postmenopausal women: Results from the women’s health initiative memory study. Int. J. Geriatr. Psychiatry 2019, 34, 692–699. [Google Scholar] [CrossRef]
- Rajan, K.B.; Barnes, L.L.; Wilson, R.S.; Weuve, J.; McAninch, E.A.; Evans, D.A. Apolipoprotein E genotypes, age, race, and cognitive decline in a population sample. J. Am. Geriatr. Soc. 2019, 67, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Schoenfeld, D. Partial residuals for the proportional hazards regression model. Biometrika 1982, 69, 239–241. [Google Scholar] [CrossRef]
- Rajan, K.B.; Barnes, L.L.; Wilson, R.S.; Weuve, J.; McAninch, E.A.; Evans, D.A. Blood pressure and risk of incident Alzheimer’s disease dementia by antihypertensive medications and APOE ε4 allele. Ann. Neurol. 2018, 83, 935–944. [Google Scholar] [CrossRef]
- Samuelsson, J.; Najar, J.; Wallengren, O.; Kern, S.; Wetterberg, H.; Mellqvist Fässberg, M.; Zetterberg, H.; Blennow, K.; Lissner, L.; Rothenberg, E.; et al. Interactions between dietary patterns and genetic factors in relation to incident dementia among 70-year-olds. Eur. J. Nutr. 2022, 61, 871–884. [Google Scholar] [CrossRef]
- Hossain, S.; Beydoun, M.A.; Weiss, J.; Kuczmarski, M.F.; Evans, M.K.; Zonderman, A.B. Longitudinal associations between dietary quality and Alzheimer’s disease genetic risk on cognitive performance among African American adults. Br. J. Nutr. 2020, 124, 1264–1276. [Google Scholar] [CrossRef]
- Cornelis, M.C.; Hu, F.B. Gene-environment interactions in the development of type 2 diabetes: Recent progress and continuing challenges. Annu. Rev. Nutr. 2012, 32, 245–259. [Google Scholar] [CrossRef]
- Tangney, C.C.; Kwasny, M.J.; Li, H.; Wilson, R.S.; Evans, D.A.; Morris, M.C. Adherence to a Mediterranean-type dietary pattern and cognitive decline in a community population. Am. J. Clin. Nutr. 2011, 93, 601–607. [Google Scholar] [CrossRef]
- Duplantier, S.C.; Gardner, C.D. A critical review of the study of neuroprotective diets to reduce cognitive decline. Nutrients 2021, 13, 2264. [Google Scholar] [CrossRef] [PubMed]
- De Crom, T.O.E.; Mooldijk, S.S.; Ikram, M.K.; Ikram, M.A.; Voortman, T. Mind diet and the risk of dementia: A population-based study. Alzheimer’s Res. Ther. 2022, 14, 8. [Google Scholar] [CrossRef] [PubMed]
- Rovio, S.; Kåreholt, I.; Helkala, E.-L.; Viitanen, M.; Winblad, B.; Tuomilehto, J.; Soininen, H.; Nissinen, A.; Kivipelto, M. Leisure-time physical activity at midlife and the risk of dementia and Alzheimer’s disease. Lancet Neurol. 2005, 4, 705–711. [Google Scholar] [CrossRef]
- Anttila, T.; Helkala, E.-L.; Viitanen, M.; Kåreholt, I.; Fratiglioni, L.; Winblad, B.; Soininen, H.; Tuomilehto, J.; Nissinen, A.; Kivipelto, M. Alcohol drinking in middle age and subsequent risk of mild cognitive impairment and dementia in old age: A prospective population based study. Bmj 2004, 329, 539. [Google Scholar] [CrossRef]
- Laitinen, M.H.; Ngandu, T.; Rovio, S.; Helkala, E.L.; Uusitalo, U.; Viitanen, M.; Nissinen, A.; Tuomilehto, J.; Soininen, H.; Kivipelto, M. Fat intake at midlife and risk of dementia and Alzheimer’s disease: A population-based study. Dement. Geriatr. Cogn. Disord. 2006, 22, 99–107. [Google Scholar] [CrossRef]
- Kivipelto, M.; Rovio, S.; Ngandu, T.; Kåreholt, I.; Eskelinen, M.; Winblad, B.; Hachinski, V.; Cedazo-Minguez, A.; Soininen, H.; Tuomilehto, J.; et al. Apolipoprotein E ɛ4 magnifies lifestyle risks for dementia: A population-based study. J. Cell. Mol. Med. 2008, 12, 2762–2771. [Google Scholar] [CrossRef]
- Barberger-Gateau, P.; Raffaitin, C.; Letenneur, L.; Berr, C.; Tzourio, C.; Dartigues, J.-F.; Alpérovitch, A. Dietary patterns and risk of dementia the three-city cohort study. Neurology 2007, 69, 1921–1930. [Google Scholar] [CrossRef]
- Podewils, L.J.; Guallar, E.; Kuller, L.H.; Fried, L.P.; Lopez, O.L.; Carlson, M.; Lyketsos, C.G. Physical activity, APOE genotype, and dementia risk: Findings from the cardiovascular health cognition study. Am. J. Epidemiol. 2005, 161, 639–651. [Google Scholar] [CrossRef]
- Huang, T.L.; Zandi, P.; Tucker, K.; Fitzpatrick, A.; Kuller, L.; Fried, L.; Burke, G.; Carlson, M. Benefits of fatty fish on dementia risk are stronger for those without APOE ε4. Neurology 2005, 65, 1409–1414. [Google Scholar] [CrossRef]
- Luchsinger, J.A.; Tang, M.X.; Siddiqui, M.; Shea, S.; Mayeux, R. Alcohol intake and risk of dementia. J. Am. Geriatr. Soc. 2004, 52, 540–546. [Google Scholar] [CrossRef]
- Licher, S.; Ahmad, S.; Karamujić-Čomić, H.; Voortman, T.; Leening, M.J.G.; Ikram, M.A.; Ikram, M.K. Genetic predisposition, modifiable-risk-factor profile and long-term dementia risk in the general population. Nat. Med. 2019, 25, 1364–1369. [Google Scholar] [CrossRef] [PubMed]
- Lourida, I.; Hannon, E.; Littlejohns, T.J.; Langa, K.M.; Hyppönen, E.; Kuźma, E.; Llewellyn, D.J. Association of lifestyle and genetic risk with incidence of dementia. JAMA 2019, 322, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Hollingworth, P.; Harold, D.; Sims, R.; Gerrish, A.; Lambert, J.-C.; Carrasquillo, M.M.; Abraham, R.; Hamshere, M.L.; Pahwa, J.S.; Moskvina, V.; et al. Common variants at ABCA7, MS4A6A/MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer’s disease. Nat. Genet. 2011, 43, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.C.; Ibrahim-Verbaas, C.A.; Harold, D.; Naj, A.C.; Sims, R.; Bellenguez, C.; DeStafano, A.L.; Bis, J.C.; Beecham, G.W.; Grenier-Boley, B.; et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat. Genet. 2013, 45, 1452–1458. [Google Scholar] [CrossRef] [PubMed]
- Naj, A.C.; Jun, G.; Beecham, G.W.; Wang, L.S.; Vardarajan, B.N.; Buros, J.; Gallins, P.J.; Buxbaum, J.D.; Jarvik, G.P.; Crane, P.K.; et al. Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1 are associated with late-onset Alzheimer’s disease. Nat. Genet. 2011, 43, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Herold, C.; Hooli, B.V.; Mullin, K.; Liu, T.; Roehr, J.T.; Mattheisen, M.; Parrado, A.R.; Bertram, L.; Lange, C.; Tanzi, R.E. Family-based association analyses of imputed genotypes reveal genome-wide significant association of Alzheimer’s disease with OSBPL6, PTPRG, and PDCL3. Mol. Psychiatry 2016, 21, 1608–1612. [Google Scholar] [CrossRef]
- Lambert, J.C.; Heath, S.; Even, G.; Campion, D.; Sleegers, K.; Hiltunen, M.; Combarros, O.; Zelenika, D.; Bullido, M.J.; Tavernier, B.; et al. Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer’s disease. Nat. Genet. 2009, 41, 1094–1099. [Google Scholar] [CrossRef]
- Jun, G.; Ibrahim-Verbaas, C.A.; Vronskaya, M.; Lambert, J.C.; Chung, J.; Naj, A.C.; Kunkle, B.W.; Wang, L.S.; Bis, J.C.; Bel-lenguez, C.; et al. A novel Alzheimer disease locus located near the gene encoding tau protein. Mol. Psychiatry 2016, 21, 108–117. [Google Scholar] [CrossRef]
- Jun, G.R.; Chung, J.; Mez, J.; Barber, R.; Beecham, G.W.; Bennett, D.A.; Buxbaum, J.D.; Byrd, G.S.; Carrasquillo, M.M.; Crane, P.K.; et al. Transethnic genome-wide scan identifies novel Alzheimer’s disease loci. Alzheimer’s Dement. 2017, 13, 727–738. [Google Scholar] [CrossRef]
- Jiang, Q.; Jin, S.; Jiang, Y.; Liao, M.; Feng, R.; Zhang, L.; Liu, G.; Hao, J. Alzheimer’s Disease Variants with the Genome-Wide Significance are Significantly Enriched in Immune Pathways and Active in Immune Cells. Mol. Neurobiol. 2016, 54, 594–600. [Google Scholar] [CrossRef]
- Kunkle, B.W.; Grenier-Boley, B.; Sims, R.; Bis, J.C.; Damotte, V.; Naj, A.C.; Boland, A.; Vronskaya, M.; Van Der Lee, S.J.; Amlie-Wolf, A.; et al. Genetic meta-analysis of diagnosed Alzheimer’s disease identifies new risk loci and implicates Aβ, tau, immunity and lipid processing. Nat. Genet. 2019, 51, 414–430. [Google Scholar] [CrossRef] [PubMed]


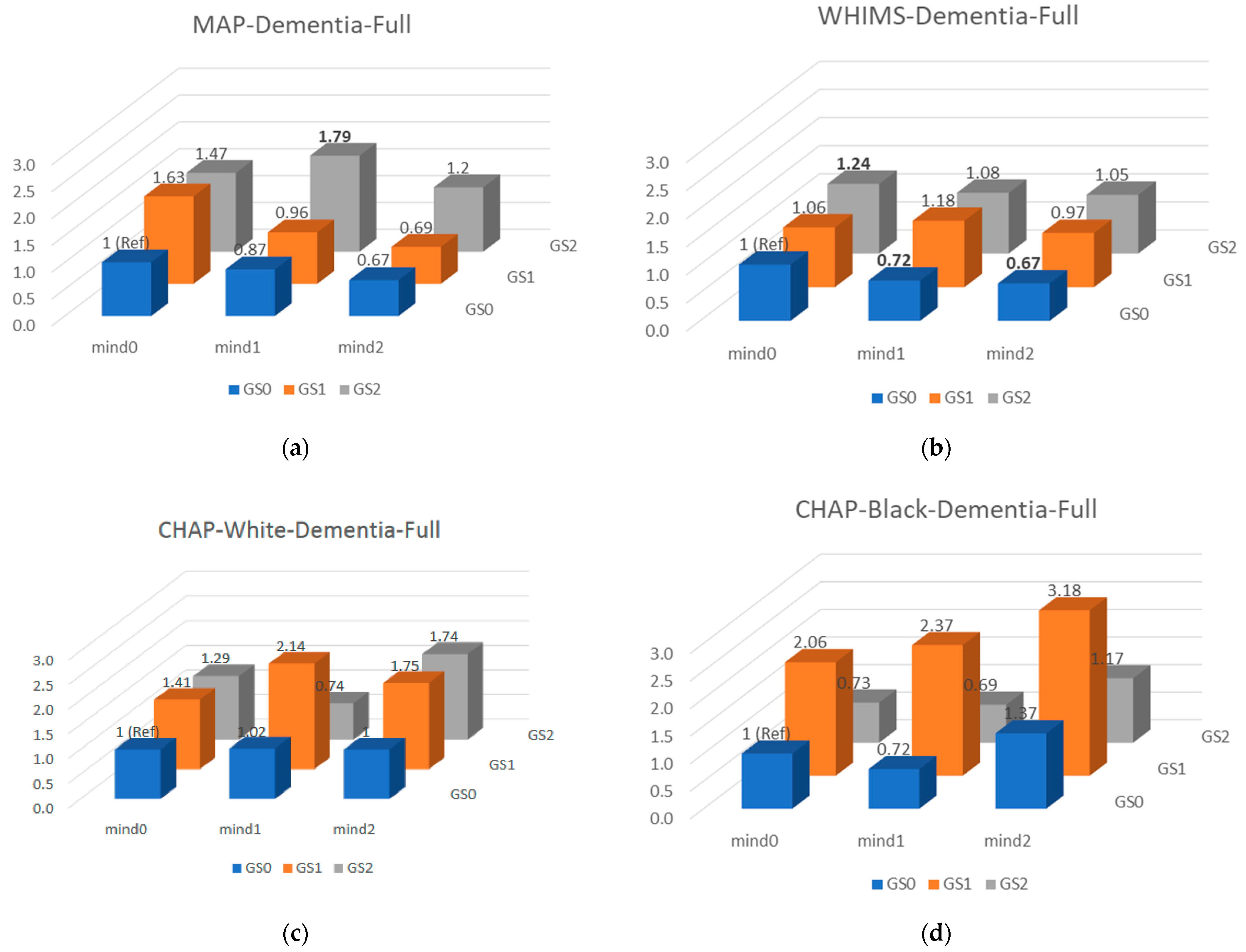{kind=link}
| MAP | WHIMS | CHAP-White | CHAP-Black | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Characteristic | T1 | T2 | T3 | T1 | T2 | T3 | T1 | T2 | T3 | T1 | T2 | T3 |
| 3.5–7.0 | 7.5–8.5 | 9.0–13.0 | 2–5.5 | 6–7 | 7.5–12 | 2–6 | 6.5–7.5 | 8–14 | 2–6 | 6.5–7.5 | 8–12.5 | |
| n = 270 | n = 233 | n = 222 | n= 1485 | n = 1917 | n = 1906 | n = 255 | n = 300 | n = 391 | n = 475 | n = 513 | n = 515 | |
| Age, years | 82.3 ± 7.2 | 82.5 ± 6.5 | 80.3 ± 6.8 | 69.8 ± 3.8 | 70.2 ± 3.85 | 70.3 ± 3.8 | 74.0 ± 6.3 | 74.2 ± 6.3 | 72.2 ± 5.7 | 71.7 ± 4.6 | 71.9 ± 4.5 | 71.1 ± 4.1 |
| Male, n (%) | 74 (27) | 60 (26) | 52 (23) | 0 (0) | 0 (0) | 0 (0) | 123 (48) | 105 (35) | 130 (33) | 218 (46) | 176 (34) | 162 (31) |
| Baseline cognitive function, Z-score | −0.05 ± 0.57 | 0.13 ± 0.50 | 0.30 ± 0.45 | −0.08 ± 1.01 | 0.01 ± 1.01 | 0.04 ± 0.99 | 0.61 ± 0.53 | 0.69 ± 0.52 | 0.81 ± 0.40 | 0.23 ± 0.58 | 0.31 ± 0.60 | 0.40 ± 0.56 |
| 2 Mild cognitive impairment, n (%) | 80 (30) | 56 (24) | 33 (15) | 0 (0) | 0 (0) | 0 (0) | 40 (16) | 41 (21) | 34 (12) | 79 (27) | 89 (31) | 78 (39) |
| Education | ||||||||||||
| College or University degree, n (%) | n/a | n/a | n/a | 318 (21) | 585 (31) | 755 (40) | n/a | n/a | n/a | n/a | n/a | n/a |
| Years of education | 14.3 ± 3.1 | 15.1 ± 2.8 | 15.5 ± 2.7 | n/a | n/a | n/a | 14.1 ± 3.0 | 14.6 ± 3.2 | 15.3 ± 3.3 | 11.3 ± 3.2 | 12.0 ± 3.2 | 12.7 ± 3.2 |
| Hypertension, n (%) | 202 (75) | 168 (72) | 152 (68) | 562 (38) | 756 (39) | 684 (36) | 101 (40) | 131 (44) | 167 (43) | 292 (62) | 324 (63) | 346 (67) |
| Diabetes, n (%) | 46 (17) | 27 (12) | 25 (11) | 105 (7) | 136 (7) | 136 (7) | 31 (12) | 28 (9) | 39 (10) | 102 (21) | 129 (25) | 123 (24) |
| Stroke, n (%) | 38 (14) | 17 (7) | 22 (10) | 23 (2) | 32 (2) | 19 (1) | 19 (7) | 21 (7) | 18 (5) | 41 (9) | 41 (8) | 35 (7) |
| Heart disease, n (%) | 55 (20) | 31 (13) | 38 (17) | 254 (17) | 310 (16) | 324 (17) | 20 (8) | 38 (13) | 55 (14) | 56 (12) | 66 (13) | 50 (10) |
| Current smoker, n (%) | 9 (3) | 2 (1) | 2 (1) | 127 (9) | 123 (6) | 76 (4) | 34 (13) | 25 (8) | 25 (6) | 70 (15) | 65 (13) | 57 (11) |
| BMI, kg/m2 | 27.5 ± 5.2 | 27.3 ± 5.0 | 26.6 ± 5.6 | 29.2 ± 5.7 | 28.5 ± 5.8 | 27.7 ± 5.3 | 26.7 ± 5.3 | 27.1 ± 4.9 | 27.2 ± 4.9 | 28.9 ± 6.2 | 29.4 ± 6.3 | 29.2 ± 5.6 |
| 3 Depression/symptoms, n (%) | 18 (7) | 8 (3) | 3 (1) | 138 (9) | 132 (7) | 109 (6) | 20 (8) | 8 (3) | 14 (4) | 62 (13) | 58 (11) | 47 (9) |
| MIND score | 6.1 ± 0.8 | 8.0 ± 0.4 | 9.9 ± 0.9 | 4.8 ± 0.7 | 6.5 ± 0.4 | 8.3 ± 0.8 | 5.2 ± 0.9 | 7.0 ± 0.4 | 9.1 ± 1.1 | 5.3 ± 0.7 | 7.0 ± 0.4 | 8.8 ± 0.9 |
| Apoe ε4 carriers, n (%) | 57 (21) | 44 (19) | 47 (21) | 187 (13) | 226 (12) | 262 (14) | 63 (25) | 69 (23) | 109 (28) | 173 (36) | 174 (34) | 206 (40) |
| GSAD | 26.6 ± 3.0 | 26.8 ± 2.9 | 26.7 ± 2.8 | 26.9 ± 3.0 | 26.8 ± 3.1 | 26.9 ± 3.0 | 26.5 ± 3.0 | 27.1 ± 3.0 | 27.0 ± 3.0 | 24.4 ± 2.6 | 24.5 ± 2.8 | 24.4 ± 2.6 |
| GSAD-I | 9.6 ± 2.1 | 9.7 ± 2.0 | 9.5 ± 1.9 | 9.7 ± 2.1 | 9.6 ± 2.2 | 9.6 ± 2.1 | 9.6 ± 2.2 | 9.8 ± 2.1 | 9.6 ± 2.2 | 9.6 ± 1.6 | 9.4 ± 1.6 | 9.4 ± 1.7 |
| GSAD-C | 5.5 ± 0.9 | 5.6 ± 0.9 | 5.5 ± 1.0 | 5.52 ± 0.95 | 5.49 ± 0.96 | 5.49 ± 0.97 | 5.5 ± 1.0 | 5.5 ± 0.9 | 5.6 ± 0.9 | 5.0 ± 1.0 | 5.0 ± 1.0 | 5.0 ± 0.9 |
| Model | MAP | WHIMS | CHAP-White | CHAP-Black | ||||
|---|---|---|---|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | OR (95% CI) | p | OR (95% CI) | p | |
| Basic 1 | ||||||||
| MIND T1 | Ref. | Ref. | Ref. | Ref. | ||||
| MIND T2 | 0.70 (0.52, 0.94) | 0.02 | 0.88 (0.80, 0.97) | 0.008 | 0.54 (0.21, 1.43) | 0.22 | 0.82 (0.35, 1.95) | 0.66 |
| MIND T3 | 0.43 (0.30, 0.61) | <0.0001 | 0.78 (0.70, 0.86) | <0.0001 | 0.49 (0.18, 1.33) | 0.16 | 0.97 (0.32, 2.87) | 0.95 |
| Trend | 0.84 (0.77, 0.91) | <0.0001 | 0.93 (0.91, 0.96) | <0.0001 | 0.81 (0.61, 1.09) | 0.17 | 0.99 (0.73, 1.34) | 0.96 |
| Full 2 | ||||||||
| MIND T1 | Ref. | Ref. | Ref. | Ref. | ||||
| MIND T2 | 0.85 (0.62, 1.16) | 0.31 | 0.87 (0.79, 0.97) | 0.008 | 0.87 (0.30, 2.54) | 0.80 | 0.86 (0.36, 2.05) | 0.74 |
| MIND T3 | 0.63 (0.42, 0.92) | 0.02 | 0.80 (0.72, 0.89) | <0.0001 | 1.23 (0.47, 3.18) | 0.68 | 1.48 (0.51, 4.27) | 0.47 |
| Trend | 0.91 (0.83, 1.00) | 0.06 | 0.95 (0.92, 0.97) | <0.0001 | 1.00 (0.81, 1.25) | 0.97 | 1.08 (0.79, 1.48) | 0.61 |
| Model | MAP | CHAP-White | CHAP-Black | |||
|---|---|---|---|---|---|---|
| β (95% CI) | p | β (95% CI) | p | β (95% CI) | p | |
| Basic 1 | ||||||
| MIND T1 | Ref. | Ref. | Ref. | |||
| MIND T2 | 0.009 (−0.01, 0.03) | 0.39 | 0.001 (−0.01, 0.01) | 0.78 | 0.001 (−0.01, 0.01) | 0.78 |
| MIND T3 | 0.04 (0.02, 0.06) | 0.0004 | 0.001(−0.01, 0.01) | 0.77 | −0.001 (−0.01, 0.01) | 0.85 |
| Trend | 0.008 (0.003, 0.01) | 0.002 | 0.0002 (−0.002, 0.003) | 0.89 | 0.001 (−0.002, 0.003) | 0.54 |
| Full 2 | ||||||
| MIND T1 | Ref. | Ref. | Ref. | |||
| MIND T2 | 0.006 (−0.01, 0.02) | 0.50 | 0.0001 (−0.01, 0.01) | 0.99 | 0.0003 (−0.01, 0.01) | 0.95 |
| MIND T3 | 0.03 (0.01, 0.05) | 0.001 | −0.0008(−0.01, 0.01) | 0.89 | −0.003 (−0.01, 0.01) | 0.51 |
| Trend | 0.006 (0.003, 0.01) | 0.002 | −0.0004 (−0.003, 0.002) | 0.78 | −0.00002 (−0.003, 0.003) | 0.99 |
| GS | MAP | WHIMS | CHAP-White | CHAP-Black | ||||
|---|---|---|---|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | OR (95% CI) | p | OR (95% CI) | p | |
| GSAD | ||||||||
| T1 | Ref. | Ref. | Ref. | Ref. | ||||
| T2 | 1.12 (0.80, 1.58) | 0.51 | 1.28 (1.16, 1.41) | <0.001 | 1.55 (0.66, 3.66) | 0.32 | 2.41 (0.82, 7.12) | 0.11 |
| T3 | 1.81 (1.31, 2.51) | 0.0003 | 1.43 (1.30, 1.58) | <0.001 | 1.55 (0.55, 4.34) | 0.41 | 0.77 (0.28, 2.11) | 0.61 |
| Trend | 1.10 (1.05, 1.15) | <0.0001 | 1.05 (1.04, 1.06) | <0.0001 | 1.09 (0.92, 1.30) | 0.32 | 1.06 (0.94, 1.20) | 0.32 |
| GSAD-I | ||||||||
| T1 | Ref. | Ref. | Ref. | Ref. | ||||
| T2 | 1.45 (1.03, 2.02) | 0.03 | 0.99 (0.89, 1.09) | 0.764 | 1.12 (0.50, 2.53) | 0.78 | 1.84 (0.76, 4.46) | 0.18 |
| T3 | 1.43 (1.03, 1.99) | 0.03 | 1.11 (1.01, 1.23) | 0.026 | 1.86 (0.74, 4.71) | 0.19 | 0.66 (0.25, 1.77) | 0.41 |
| Trend | 1.06 (1.00, 1.14) | 0.06 | 1.02 (1.01, 1.04) | 0.014 | 1.15 (0.96, 1.39) | 0.14 | 0.88 (0.73, 1.07) | 0.20 |
| GSAD-C | ||||||||
| T1 | Ref. | Ref. | Ref. | Ref. | ||||
| T2 | 0.86 (0.62, 1.20) | 0.38 | 0.99 (0.90, 1.10) | 0.911 | 1.01 (0.41, 2.45) | 0.99 | 1.29 (0.41, 4.04) | 0.66 |
| T3 | 1.08 (0.79, 1.47) | 0.64 | 1.25 (1.13, 1.37) | <0.0001 | 2.08 (0.82, 5.27) | 0.12 | 0.57 (0.21, 1.55) | 0.27 |
| Trend | 1.06 (0.93, 1.22) | 0.36 | 1.08 (1.03, 1.12) | 0.001 | 1.54 (0.86, 2.74) | 0.15 | 1.06 (0.81, 1.38) | 0.68 |
| Outcome | Interaction Term | MAP | WHIMS | CHAP (White) | CHAP (Black) | Meta-Analysis | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| β (SE) | p | β (SE) | p | β (SE) | p | β (SE) | p | β (SE) | p | ||
| All-cause dementia | GSAD × MIND | −0.001 (0.01) | 0.009 | 0.008 (0.004) | 0.08 | −0.02 (0.04) | 0.63 | 0.02 (0.04) | 0.52 | 0.007 (0.004) | 0.07 |
| GSAD-I × MIND | −0.009 (0.02) | 0.67 | −0.005 (0.006) | 0.47 | −0.02 (0.05) | 0.61 | 0.08 (0.05) | 0.13 | −0.004 (0.006) | 0.44 | |
| GSAD-C × MIND | 0.006 (0.04) | 0.88 | −0.03 (0.01) | 0.07 | −0.07 (0.11) | 0.51 | 0.04 (0.10) | 0.70 | −0.02 (0.01) | 0.02 | |
| APOE × MIND | 0.12 (0.09) | 0.17 | −0.13 (0.03) | <0.0001 | 0.42 (0.22) | 0.05 | −0.08 (0.33) | 0.81 | −0.10 (0.03) | 0.0006 | |
| Alzheimer’s disease | GSAD × MIND | −0.001 (0.01) | 0.004 | n/a | n/a | −0.02 (0.04) | 0.65 | 0.02 (0.04) | 0.55 | −0.0009 (0.009) | 0.92 |
| GSAD-I × MIND | −0.006 (0.02) | 0.76 | n/a | n/a | −0.02 (0.05) | 0.69 | 0.08 (0.06) | 0.12 | −0.0002 (0.02) | 0.99 | |
| GSAD-C × MIND | 0.01 (0.04) | 0.07 | n/a | n/a | −0.05 (0.12) | 0.69 | 0.04 (0.11) | 0.72 | 0.008 (0.04) | 0.83 | |
| APOE × MIND | 0.14 (0.09) | 0.12 | n/a | n/a | 0.44 (0.23) | 0.05 | −0.09 (0.34) | 0.80 | 0.16 (0.08) | 0.04 | |
| Cognitive decline | GSAD × MIND × time | 0.001 (0.0008) | 0.24 | n/a | n/a | 0.001 (0.0004) | 0.23 | −0.0006 (0.0005) | 0.19 | 0.0005 (0.0003) | 0.12 |
| GSAD-I × MIND × time | 0.0008 (0.001) | 0.49 | n/a | n/a | −0.0001 (0.001) | 0.93 | −0.002 (0.0008) | 0.008 | −0.0007 (0.0005) | 0.20 | |
| GSAD-C × MIND × time | 0.003 (0.002) | 0.28 | n/a | n/a | 0.001 (0.001) | 0.57 | −0.003 (0.001) | 0.016 | −0.0006 (0.0007) | 0.40 | |
| APOE × MIND × time | −0.002 (0.006) | 0.69 | n/a | n/a | 0.0001 (0.003) | 0.97 | 0.006 (0.003) | 0.04 | 0.003 (0.002) | 0.21 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, T.H.T.; Beck, T.; Bennett, D.A.; Schneider, J.A.; Hayden, K.M.; Shadyab, A.H.; Rajan, K.B.; Morris, M.C.; Cornelis, M.C. Adherence to MIND Diet, Genetic Susceptibility, and Incident Dementia in Three US Cohorts. Nutrients 2022, 14, 2759. https://doi.org/10.3390/nu14132759
Vu THT, Beck T, Bennett DA, Schneider JA, Hayden KM, Shadyab AH, Rajan KB, Morris MC, Cornelis MC. Adherence to MIND Diet, Genetic Susceptibility, and Incident Dementia in Three US Cohorts. Nutrients. 2022; 14(13):2759. https://doi.org/10.3390/nu14132759
Chicago/Turabian StyleVu, Thanh Huyen T., Todd Beck, David A. Bennett, Julie A. Schneider, Kathleen M. Hayden, Aladdin H. Shadyab, Kumar B. Rajan, Martha Clare Morris, and Marilyn C. Cornelis. 2022. "Adherence to MIND Diet, Genetic Susceptibility, and Incident Dementia in Three US Cohorts" Nutrients 14, no. 13: 2759. https://doi.org/10.3390/nu14132759
APA StyleVu, T. H. T., Beck, T., Bennett, D. A., Schneider, J. A., Hayden, K. M., Shadyab, A. H., Rajan, K. B., Morris, M. C., & Cornelis, M. C. (2022). Adherence to MIND Diet, Genetic Susceptibility, and Incident Dementia in Three US Cohorts. Nutrients, 14(13), 2759. https://doi.org/10.3390/nu14132759