
An Ecological Validity Model for the Prevention of Obesity: Non-Nutritive Sweetener Consumption in Rats and the Effects of Switching from Sugar-Sweetened to Diet Beverages



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sample Size
2.3. Apparatus
2.4. Test Drinks
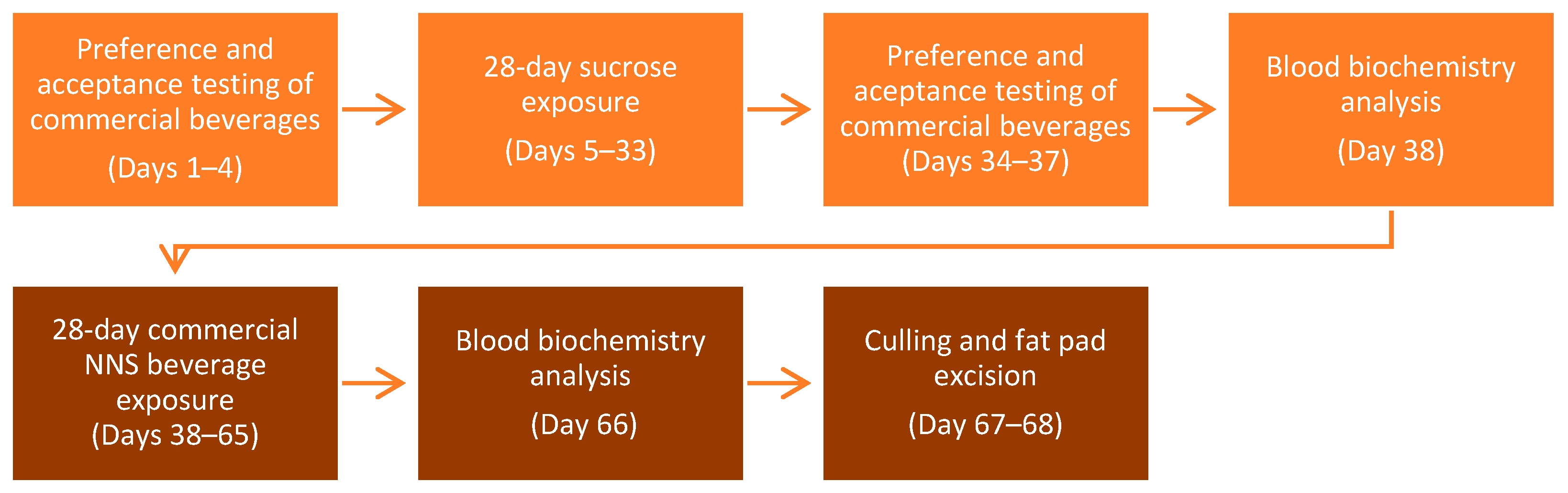
2.5. General Procedures
2.5.1. Phase 1
2.5.2. Phase 2
2.6. Data Analysis
3. Results
3.1. Two-Bottle Preference Tests
3.1.1. Test 1 Pre-Sucrose Exposure (Days 1–2)
3.1.2. Test 2 Post-Sucrose Exposure (Days 34–35)
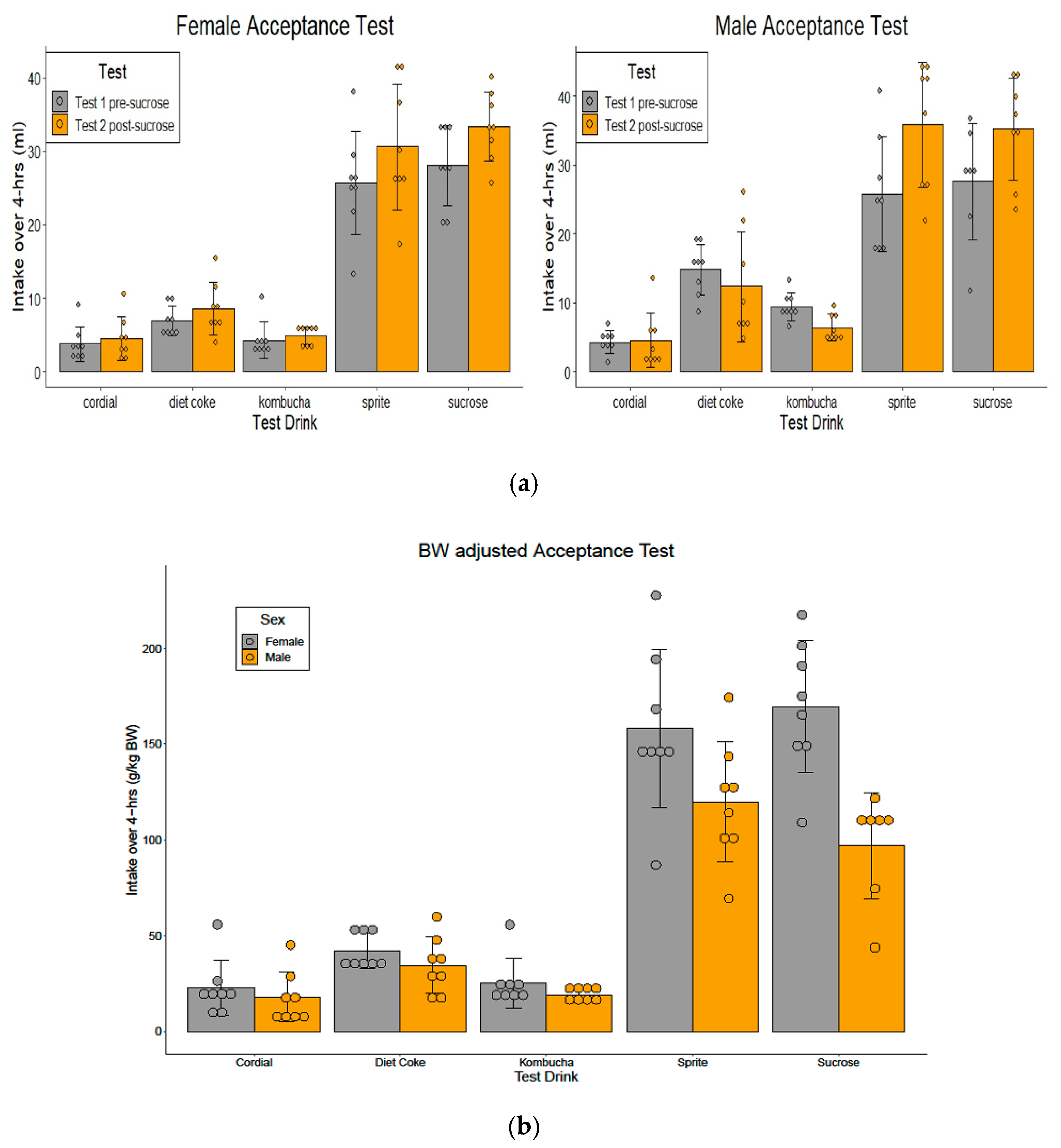
3.2. Four-Hour Acceptance Testshis
3.2.1. Test 1 Pre-Sucrose Exposure (Days 3–4)
3.2.2. Test 2 Post-Sucrose Exposure (Days 36–37)
3.3. Body Weight and Weight Gain
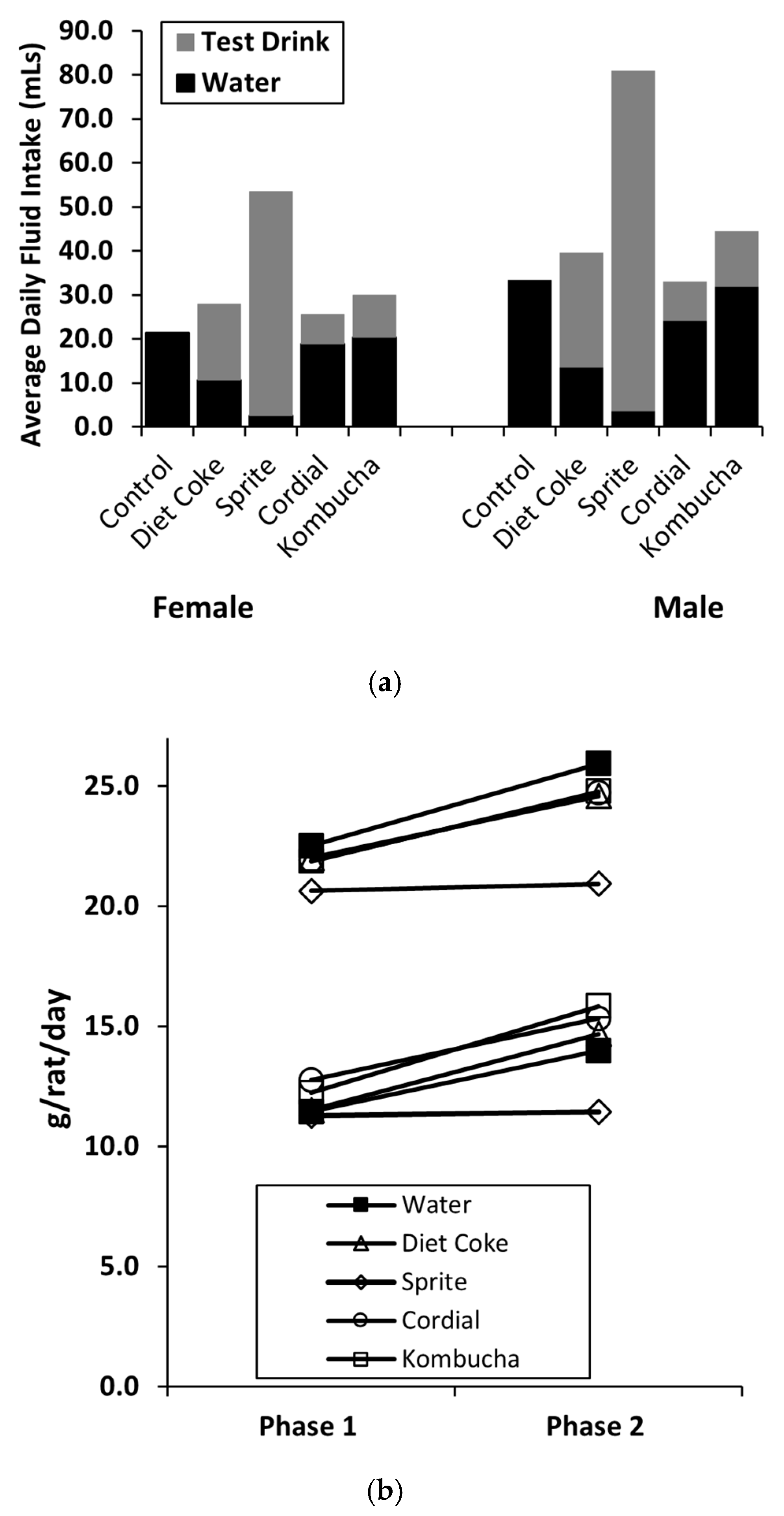
3.4. Consumption and Energy Intake
3.4.1. Phase 1: Sucrose and Chow Consumption, Energy Intake
3.4.2. Phase 2: NNS Drink and Chow Consumption, Energy Intake
3.5. Fasting Blood Glucose and Plasma Insulin
3.6. Retroperitoneal Fat Pad Mass
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dinsa, G.D.; Goryakin, Y.; Fumagalli, E.; Suhrcke, M. Obesity and Socioeconomic Status in Developing Countries: A Systematic Review. Obes. Rev. 2012, 13, 1067–1079. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, H.; Roser, M. Obesity. Our World Data 2017. Available online: https://www.ourworldindata.org/obesity (accessed on 28 June 2022).
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 10 March 2022).
- Clinical Guidelines on the Identification, Evaluation, and Treatment of Overweight and Obesity in Adults: Executive Summary. Expert Panel on the Identification, Evaluation, and Treatment of Overweight in Adults. Am. J. Clin. Nutr. 1998, 68, 899–917. [CrossRef] [Green Version]
- Yang, Q.; Zhang, Z.; Gregg, E.W.; Flanders, W.D.; Merritt, R.; Hu, F.B. Added Sugar Intake and Cardiovascular Diseases Mortality Among US Adults. JAMA Intern. Med. 2014, 174, 516. [Google Scholar] [CrossRef] [Green Version]
- Malik, V.S.; Hu, F.B. Sweeteners and Risk of Obesity and Type 2 Diabetes: The Role of Sugar-Sweetened Beverages. Curr. Diabetes Rep. 2012, 12, 195–203. [Google Scholar] [CrossRef] [PubMed]
- WHO Guideline; World Health Organization: Geneva, Switzerland, 2015; ISBN 978-92-4-069422-4/978-92-4-154902-8.
- National Health and Medical Research Council (Australia); Department of Health and Ageing Eat for Health. Providing the Scientific Evidence for Healthier Australian Diets. In Australian Dietary Guidelines; National Health and Medical Research Council: Canberra, Australia, 2013; ISBN 9781864965742 9781864965759. [Google Scholar]
- Bailey, R.; Fulgoni, V.; Cowan, A.; Gaine, P. Sources of Added Sugars in Young Children, Adolescents, and Adults with Low and High Intakes of Added Sugars. Nutrients 2018, 10, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, P.; Ahuja, S.; Sriram, K. Non-Nutritive Sweeteners: Review and Update. Nutrition 2013, 29, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.P.; Williams, K.; Resendez, R.G.; Hunt, K.J.; Hazuda, H.P.; Stern, M.P. Fueling the Obesity Epidemic? Artificially Sweetened Beverage Use and Long-Term Weight Gain. Obesity 2008, 16, 1894–1900. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial Sweeteners Induce Glucose Intolerance by Altering the Gut Microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Swithers, S.E.; Baker, C.R.; Davidson, T.L. General and Persistent Effects of High-Intensity Sweeteners on Body Weight Gain and Caloric Compensation in Rats. Behav. Neurosci. 2009, 123, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Boakes, R.A.; Kendig, M.D.; Martire, S.I.; Rooney, K.B. Sweetening Yoghurt with Glucose, but Not with Saccharin, Promotes Weight Gain and Increased Fat Pad Mass in Rats. Appetite 2016, 105, 114–128. [Google Scholar] [CrossRef]
- Glendinning, J.I.; Hart, S.; Lee, H.; Maleh, J.; Ortiz, G.; Ryu, Y.S.; Sanchez, A.; Shelling, S.; Williams, N. Low-Calorie Sweeteners Cause Only Limited Metabolic Effects in Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R70–R80. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Smith, K.R.; Crouch, A.L.; Sharma, V.; Yi, F.; Vargova, V.; LaMoia, T.E.; Dupont, L.M.; Serna, V.; Tang, F.; et al. High-Dose Saccharin Supplementation Does Not Induce Gut Microbiota Changes or Glucose Intolerance in Healthy Humans and Mice. Microbiome 2021, 9, 11. [Google Scholar] [CrossRef]
- Rogers, P.J.; Hogenkamp, P.S.; de Graaf, C.; Higgs, S.; Lluch, A.; Ness, A.R.; Penfold, C.; Perry, R.; Putz, P.; Yeomans, M.R.; et al. Does Low-Energy Sweetener Consumption Affect Energy Intake and Body Weight? A Systematic Review, Including Meta-Analyses, of the Evidence from Human and Animal Studies. Int. J. Obes. 2016, 40, 381–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morahan, H.L.; Leenaars, C.H.C.; Boakes, R.A.; Rooney, K.B. Metabolic and Behavioural Effects of Prenatal Exposure to Non-Nutritive Sweeteners: A Systematic Review and Meta-Analysis of Rodent Models. Physiol. Behav. 2020, 213, 112696. [Google Scholar] [CrossRef]
- Dunford, E.K.; Coyle, D.H.; Louie, J.C.Y.; Rooney, K.; Blaxland, A.; Pettigrew, S.; Jones, A. Changes in the Presence of Nonnutritive Sweeteners, Sugar Alcohols, and Free Sugars in Australian Foods. J. Acad. Nutr. Diet. 2022, 122, 991–999. [Google Scholar] [CrossRef]
- Best Practice Methodology in the Use of Animals for Scientific Purposes|NHMRC. Available online: https://www.nhmrc.gov.au/about-us/publications/best-practice-methodology-use-animals-scientific-purposes (accessed on 30 March 2022).
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE Guidelines 2.0: Updated Guidelines for Reporting Animal Research. J. Cereb. Blood Flow Metab. 2020, 40, 1769–1777. [Google Scholar] [CrossRef]
- Speakman, J.R. Use of High-Fat Diets to Study Rodent Obesity as a Model of Human Obesity. Int. J. Obes. 2019, 43, 1491–1492. [Google Scholar] [CrossRef] [Green Version]
- Kendig, M.D.; Fu, M.X.; Rehn, S.; Martire, S.I.; Boakes, R.A.; Rooney, K.B. Metabolic and Cognitive Improvement from Switching to Saccharin or Water Following Chronic Consumption by Female Rats of 10% Sucrose Solution. Physiol. Behav. 2018, 188, 162–172. [Google Scholar] [CrossRef]
- Additives. Available online: https://www.foodstandards.gov.au/consumer/additives/additiveoverview/Pages/default.aspx (accessed on 30 March 2022).
- Boakes, R.A.; Rehn, S.; Badolato, C.; Rooney, K.B. Reduced Acceptance of Saccharin Solutions by Rats Previously Consuming More Highly Palatable Solutions. Physiol. Behav. 2020, 218, 112822. [Google Scholar] [CrossRef] [PubMed]
- Rehn, S.; Boakes, R.A.; Badolato, C.J.; Rooney, K.B. Sex Differences in Recovery from Cognitive and Metabolic Impairments Induced by Supplementary Sucrose in Rats. Physiol. Behav. 2021, 239, 113515. [Google Scholar] [CrossRef]
- Sclafani, A.; Bahrani, M.; Zukerman, S.; Ackroff, K. Stevia and Saccharin Preferences in Rats and Mice. Chem. Senses 2010, 35, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Otero-Losada, M.E.; Mc Loughlin, S.; Rodríguez-Granillo, G.; Müller, A.; Ottaviano, G.; Moriondo, M.; Cutrin, J.C.; Milei, J. Metabolic Disturbances and Worsening of Atherosclerotic Lesions in ApoE-/- Mice after Cola Beverages Drinking. Cardiovasc. Diabetol. 2013, 12, 57. [Google Scholar] [CrossRef] [PubMed]
- Otero-Losada, M.; Cao, G.; González, J.; Muller, A.; Ottaviano, G.; Lillig, C.; Capani, F.; Ambrosio, G.; Milei, J. Functional and Morphological Changes in Endocrine Pancreas Following Cola Drink Consumption in Rats. PLoS ONE 2015, 10, e0118700. [Google Scholar] [CrossRef]
- Boakes, R.A.; Martire, S.I.; Rooney, K.B.; Kendig, M.D. Individual Differences in Saccharin Acceptance Predict Rats’ Food Intake. Physiol. Behav. 2016, 164, 151–156. [Google Scholar] [CrossRef]
- Dess, N.K. Responses to Basic Taste Qualities in Rats Selectively Bred for High versus Low Saccharin Intake. Physiol. Behav. 2000, 69, 247–257. [Google Scholar] [CrossRef]
- Kendig, M.D.; Rooney, K.B.; Corbit, L.H.; Boakes, R.A. Persisting Adiposity Following Chronic Consumption of 10% Sucrose Solution: Strain Differences and Behavioural Effects. Physiol. Behav. 2014, 130, 54–65. [Google Scholar] [CrossRef]
- Chusyd, D.E.; Wang, D.; Huffman, D.M.; Nagy, T.R. Relationships between Rodent White Adipose Fat Pads and Human White Adipose Fat Depots. Front. Nutr. 2016, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Vital, P.; Larrieta, E.; Hiriart, M. Sexual Dimorphism in Insulin Sensitivity and Susceptibility to Develop Diabetes in Rats. J. Endocrinol. 2006, 190, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. The Artificial Sweetener Acesulfame Potassium Affects the Gut Microbiome and Body Weight Gain in CD-1 Mice. PLoS ONE 2017, 12, e0178426. [Google Scholar] [CrossRef]
- Uebanso, T.; Ohnishi, A.; Kitayama, R.; Yoshimoto, A.; Nakahashi, M.; Shimohata, T.; Mawatari, K.; Takahashi, A. Effects of Low-Dose Non-Caloric Sweetener Consumption on Gut Microbiota in Mice. Nutrients 2017, 9, 560. [Google Scholar] [CrossRef] [Green Version]
- Baker, M. 1,500 Scientists Lift the Lid on Reproducibility. Nature 2016, 533, 452–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyhan, A.A. Lost in Translation: The Valley of Death across Preclinical and Clinical Divide—Identification of Problems and Overcoming Obstacles. Transl. Med. Commun. 2019, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, G.S.; Veening-Griffioen, D.H.; Boon, W.P.C.; Moors, E.H.M.; van Meer, P.J.K. Levelling the Translational Gap for Animal to Human Efficacy Data. Animals 2020, 10, 1199. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.; Hambly, C.; Mitchell, S.; Król, E. Animal Models of Obesity. Obes. Rev. 2007, 8, 55–61. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Coke™ | Sprite™ | Cottee’s Cordial Raspberry (No Added Sugar)™ | Remedy Kombucha Ginger and Lemon™ | |
|---|---|---|---|---|
| Sweetener(s) * | aspartame acesulfame K | sucrose stevia | cyclamate acesulfame K sucralose | stevia erythritol |
| Other ingredients * | water, colour (caramel IV), caffeine, phosphoric acid, citric acid | water, citric acid, sodium citrate, flavour, potassium sorbate | water, citric acid, flavour, sodium carbomethylcellulose, sodium benzoate, sodium metabisulphites, colour (carmoisine) | water, kombucha culture, black tea, green tea, ginger, lemon |
| Nutritional information (per 100 mL) | ||||
| Energy (kCal) | 0.5 | 36 | 0.25 | 7 |
| Energy (kJ) | 1.9 | 151 | 2 | 29 |
| Fat (g) | 0 | 0 | 0 | < 0.1 |
| Protein (g) | 0.05 | 0 | 0 | < 0.1 |
| Carbohydrates (g) | 0.1 | 8.6 | 0 | 1.5 |
| including sugars | 0 | 8.6 | 0 | 0 |
| Sodium (mg) | 1.0 | 6.7 | 12 | < 5 |
| Phase 1 Sucrose Exposure | Control (n = 8) | Diet Coke (n = 8) | Sprite (n = 8) | Cordial (n = 8) | Kombucha (n = 8) |
|---|---|---|---|---|---|
| Baseline BW (g) | 142.6 ± 4.4 | 141.9 ± 4.1 | 141.7 ± 4.1 | 142.1.1 ± 3.7 | 142.4 ± 3.6 |
| Chow intake (g/rat/day) | 11.4 ± 0.4 | 11.5 ± 0.3 | 11.3 ± 0.4 | 12.8 ± 0.5 | 12.2 ± 0.4 |
| Sucrose (mL/rat/day) | 78.6 ± 1.4 | 81.5 ± 1.3 | 89.9 ± 3.0 | 75.6 ± 4.0 | 72.4 ± 2.4 |
| Water (mL/rat/day) | 0.7 ± 0.2 | 0.5 ± 0.1 | 0.7 ± 0.01 | 0.7 ± 0.01 | 0.7 ± 0.01 |
| Total energy intake (kcal/rat/day) | 70.3 ± 1.3 | 70.6 ± 1.1 | 74.6 ± 0.9 | 73.6 ± 1.1 | 70.5 ± 1.2 |
| Fasting blood glucose (mM) | 4.7 ± 0.1 | 4.7 ± 0.1 | 4.8 ± 0.1 | 5.0 ± 0.1 | 4.7 ± 0.1 |
| Phase 2 NNS exposure | |||||
| Chow intake (g/rat/day) | 14.0 ± 0.7 b | 14.7 ± 0.2 | 11.4 ± 0.4 * | 15.3 ± 0.4 | 15.8 ± 0.5 a |
| Test drink (mL/rat/day) | N/A | 17.5 ± 1.5 | 50.9 ± 1.6 | 6.8 ± 0.6 | 9.6 ± 0.4 |
| Water (mL/rat/day) | 21.4 ± 0.4 | 10.5 ± 0.7 | 2.6 ± 0.3 | 18.8 ± 0.6 | 20.3 ± 0.8 |
| Total fluid intake (mL/rat/day) | 21.4 ± 0.4 | 28.0 ± 0.9 | 53.5 ± 1.7 | 25.7 ± 0.4 | 30.0 ± 0.9 |
| Total energy intake (kcal/rat/day) | 47.3 ± 0.9 | 49.9 ± 0.7 | 56.7 ± 0.7 | 51.3 ± 0.7 | 53.8 ± 0.8 |
| Fasting blood glucose (mM) | 4.6 ± 0.1 | 4.8 ± 0.1 | 5.1 ± 0.1 * | 4.7 ± 0.1 | 4.7 ± 0.1 |
| Fasting plasma insulin (ng/mL) | NR | NR | NR | NR | NR |
| Retroperitoneal fat mass (g) | 13.0 ± 0.9 | 17.6 ± 2.2 | 16.0 ± 1.7 | 17.8 ± 2.5 | 18.2 ± 2.3 |
| Phase 1 Sucrose Exposure | Control (n = 8) | Diet Coke (n = 8) | Sprite (n = 8) | Cordial (n = 8) | Kombucha (n = 8) |
|---|---|---|---|---|---|
| Baseline BW (g) | 247.6 ± 5.9 | 243.9 ± 5.0 | 248.4 ± 3.4 | 248.1 ± 5.2 | 244.6 ± 4.6 |
| Chow intake (g/rat/day) | 22.5 ± 0.6 | 22.0 ± 0.5 | 20.6 ± 0.7 | 21.9 ± 0.6 | 21.9 ± 0.5 |
| Sucrose (mL/rat/day) | 101.7 ± 4.7 | 85.2 ± 1.5 | 90.1 ± 4.1 | 77.0 ± 1.6 | 119.9 ± 7.4 |
| Water (mL/rat/day) | 0.8 ± 0.1 | 0.8 ± 0.03 | 1.0 ± 0.1 | 1.0 ± 0.2 | 1.1 ± 0.2 |
| Total energy intake (kcal/rat/day) | 115.9 ± 1.4 | 108.1 ± 1.8 | 105.2 ± 2.0 | 104.6 ± 2.3 | 122.6 ± 1.9 |
| Fasting blood glucose (mM) | 4.8 ± 0.1 | 5.1 ± 0.1 | 4.9 ± 0.1 | 5.2 ± 0.2 | 5.2 ± 0.1 |
| Fasting plasma insulin (ng/mL) | 0.7 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.1 | 1.1 ± 0.1 | 1.0 ± 0.1 |
| Phase 2 NNS exposure | |||||
| Chow intake (g/rat/d) | 25.9 ± 0.2 | 24.6 ± 0.2 | 20.9 ± 0.2 * | 24.7 ± 0.3 | 24.8 ± 0.2 |
| Test drink (mL/rat/day) | N/A | 26.0 ± 1.8 | 77.5 ± 1.8 | 9.0 ± 0.7 | 12.6 ± 1.2 |
| Water (mL/rat/day) | 33.5 ± 0.4 | 13.5 ± 0.7 | 3.6 ± 0.3 | 24.1 ± 0.6 | 31.9 ± 0.6 |
| Total fluid intake (mL/rat/day) | 33.5 ± 0.4 | 39.5 ± 1.3 | 81.1 ± 2.0 | 33.1 ± 0.4 | 44.5 ± 1.2 |
| Total energy intake (kcal/rat/day) | 88.1 ± 0.8 | 83.7 ± 0.7 | 99.0 ± 1.0 * | 84.0 ± 1.0 | 85.1 ± 0.9 |
| Fasting BGL (mM) | 5.5 ± 0.1 | 5.3 ± 0.1 | 5.2 ± 0.1 | 5.2 ± 0.1 | 5.2 ± 0.2 |
| Fasting plasma insulin (ng/mL) | 0.5 ± 0.1 b | 0.8 ± 0.1 | 1.0 ± 0.1 a | 0.9 ± 0.1 | 0.8 ± 0.1 |
| Retroperitoneal fat mass (g) | 25.1 ± 1.8 | 24.1 ± 1.8 | 23.9 ± 3.1 | 19.3 ± 1.5 | 21.4 ± 2.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morahan, H.; Rooney, K. An Ecological Validity Model for the Prevention of Obesity: Non-Nutritive Sweetener Consumption in Rats and the Effects of Switching from Sugar-Sweetened to Diet Beverages. Nutrients 2022, 14, 2758. https://doi.org/10.3390/nu14132758
Morahan H, Rooney K. An Ecological Validity Model for the Prevention of Obesity: Non-Nutritive Sweetener Consumption in Rats and the Effects of Switching from Sugar-Sweetened to Diet Beverages. Nutrients. 2022; 14(13):2758. https://doi.org/10.3390/nu14132758
Chicago/Turabian StyleMorahan, Heidi, and Kieron Rooney. 2022. "An Ecological Validity Model for the Prevention of Obesity: Non-Nutritive Sweetener Consumption in Rats and the Effects of Switching from Sugar-Sweetened to Diet Beverages" Nutrients 14, no. 13: 2758. https://doi.org/10.3390/nu14132758
APA StyleMorahan, H., & Rooney, K. (2022). An Ecological Validity Model for the Prevention of Obesity: Non-Nutritive Sweetener Consumption in Rats and the Effects of Switching from Sugar-Sweetened to Diet Beverages. Nutrients, 14(13), 2758. https://doi.org/10.3390/nu14132758