
Therapeutic Benefits and Dietary Restrictions of Fiber Intake: A State of the Art Review

 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
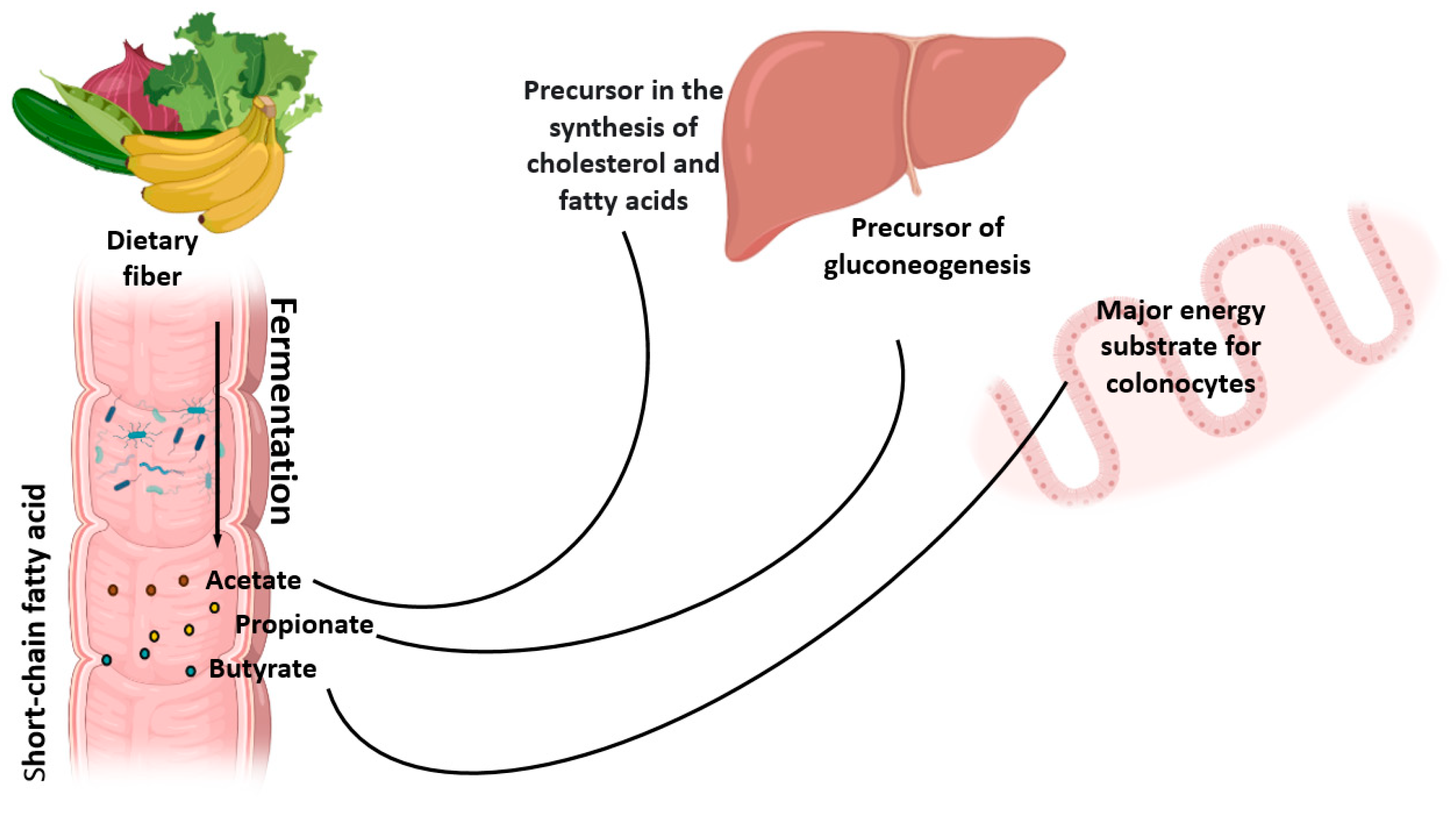
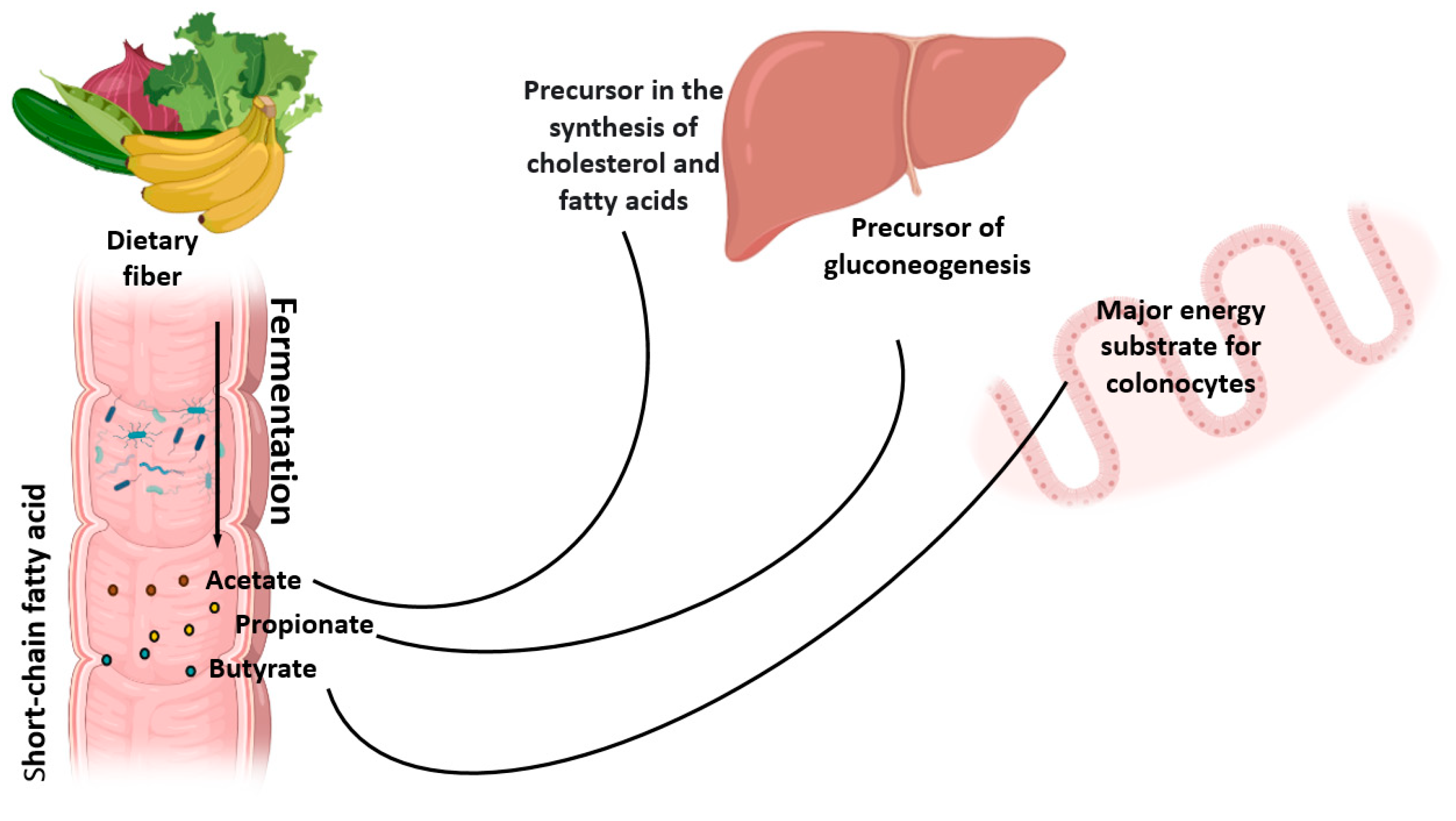
2. Metabolism of Natural Dietary Fibers (NDF)
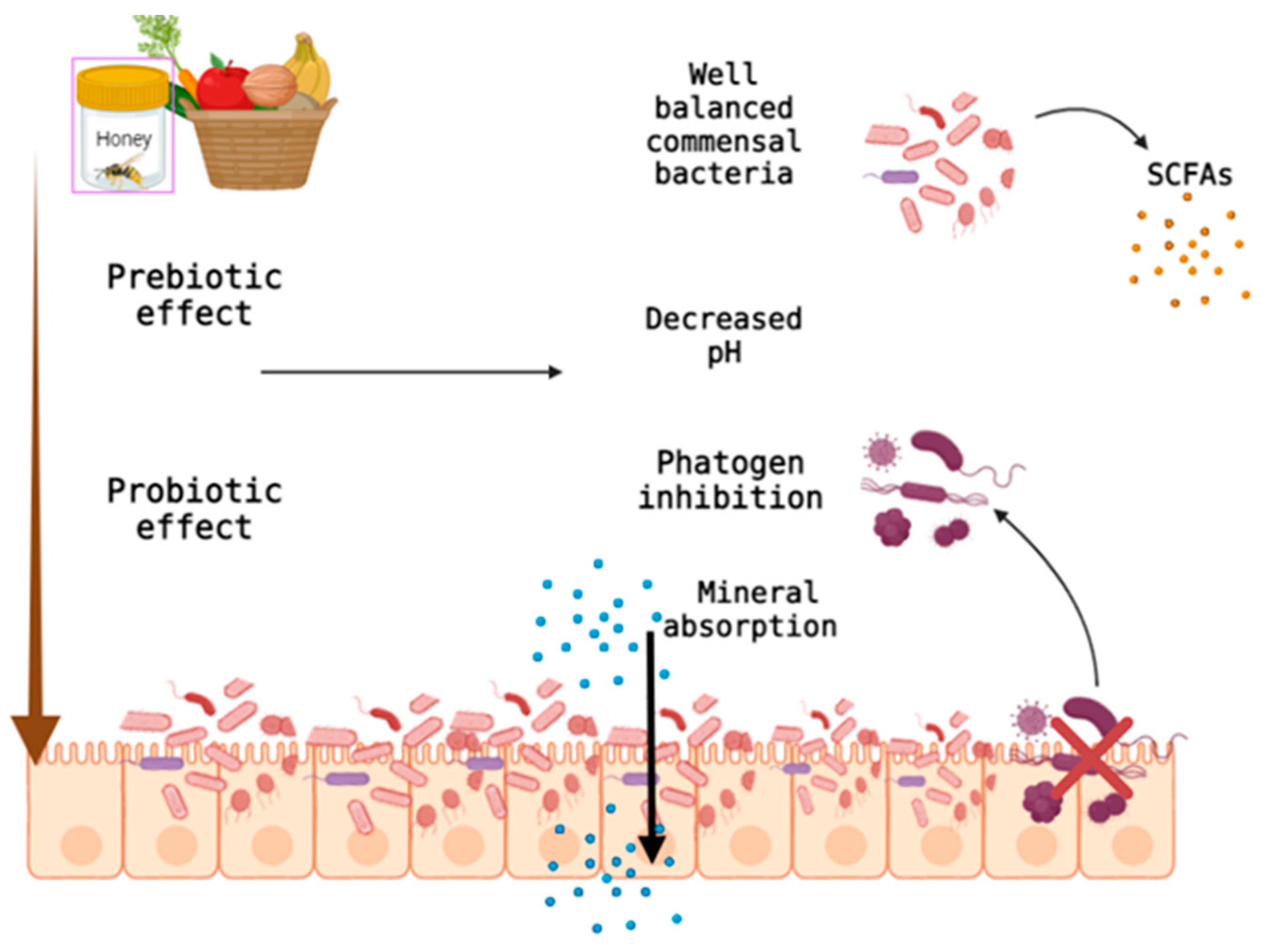
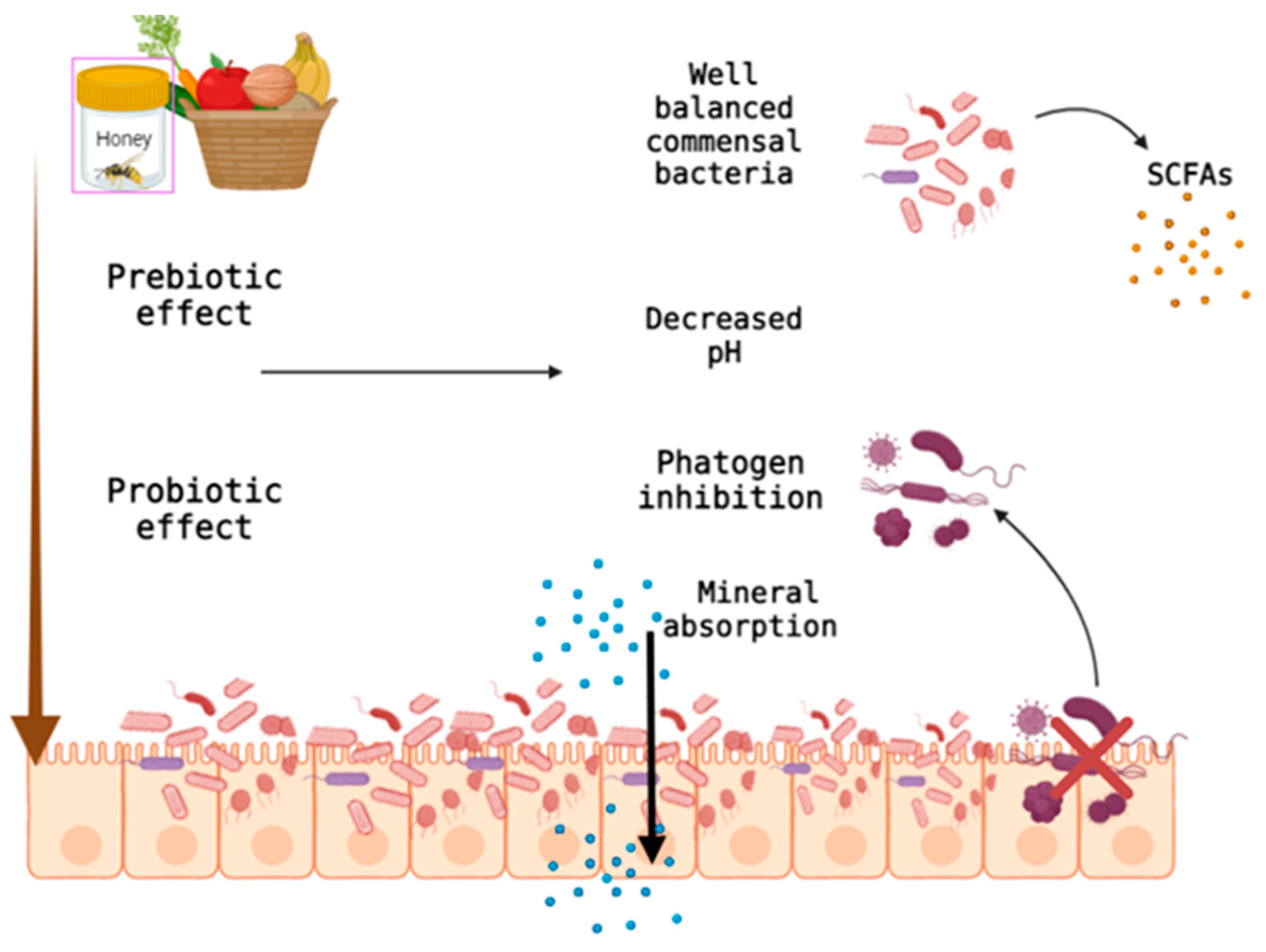
3. Natural Dietary Fibers and the Intestinal Microbiome
4. Dietary Fiber Consumption in Different Countries
5. Dietary Fibers and Metabolic Diseases
5.1. Obesity
- -
- the formation of a mechanical barrier at the intestinal level by increasing the viscosity, which causes a decrease in intestinal transit, a reduction in the absorption of glucose and fatty acids and a decrease in the percentage of adipose tissue;
- -
- lowering the rate of glucose absorption and reducing the glycemic index of food, reducing the feeling of hunger;
- -
- soluble dietary fiber influences the gut microbiome by growing saprotrophic bacterial species, having a prebiotic effect; and
- -
- by fermentative degradation in the colon of the dietary fiber, saturated fatty acids are generated with short chains that contribute to the reduction of body weight by delaying the evacuation of the stomach, followed by the increase of the feeling of satiety and the reduction of insulin sensitivity.
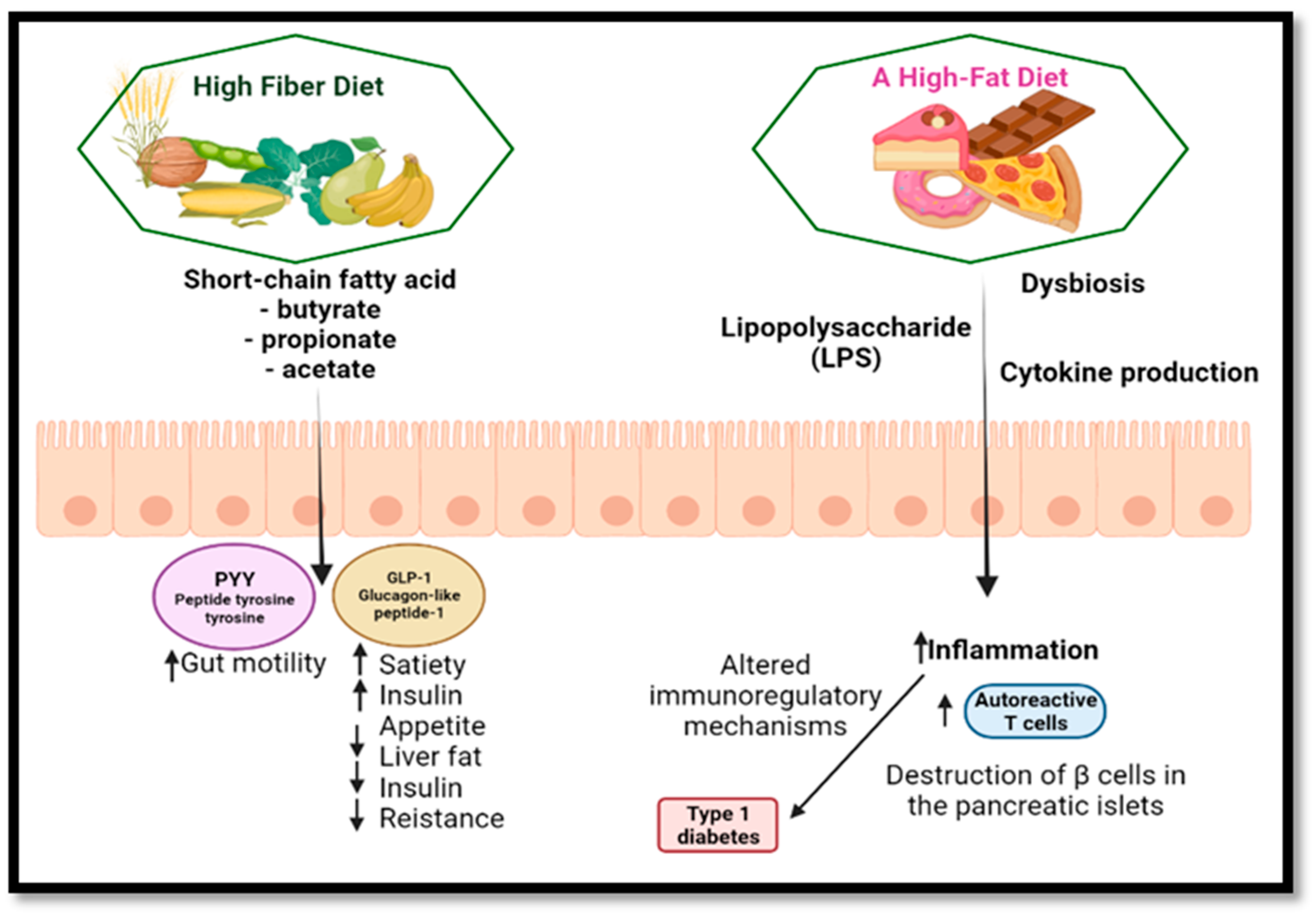
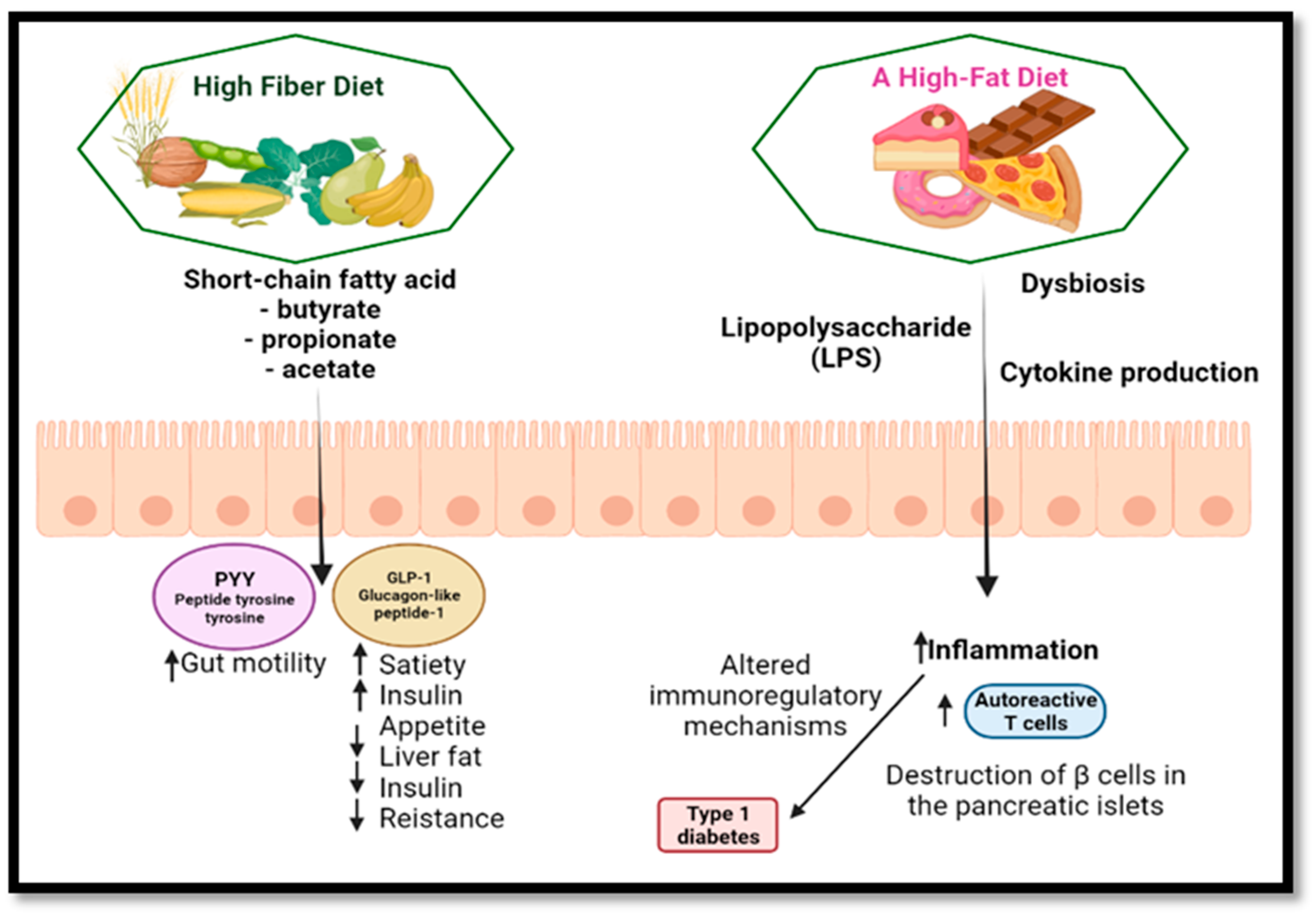
5.2. Diabetes
6. Dietary Fibers and Neurological Impact
7. Dietary Fiber in the Diet and the Risk of Cardiovascular Disease
8. Autoimmune Diseases
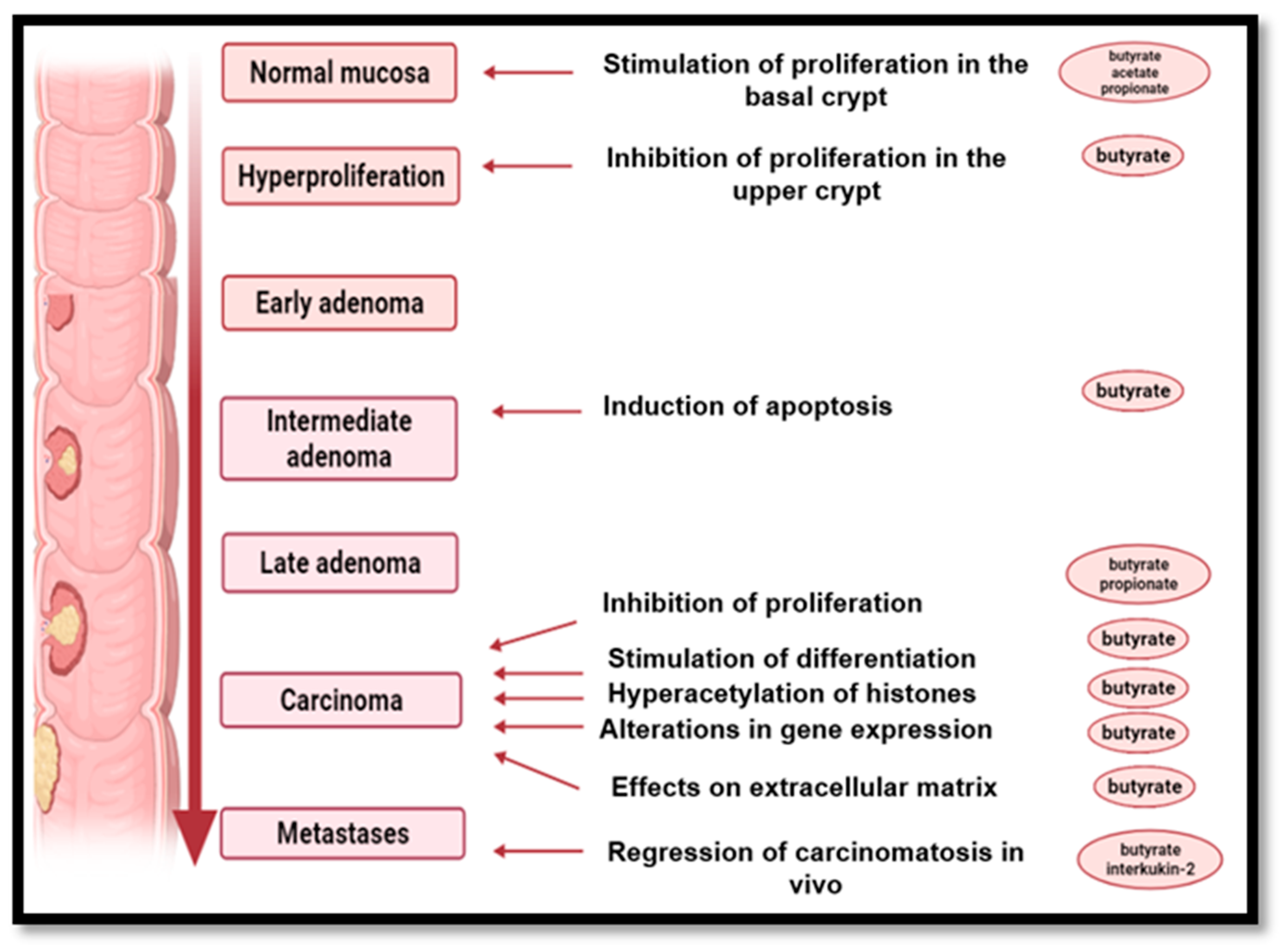
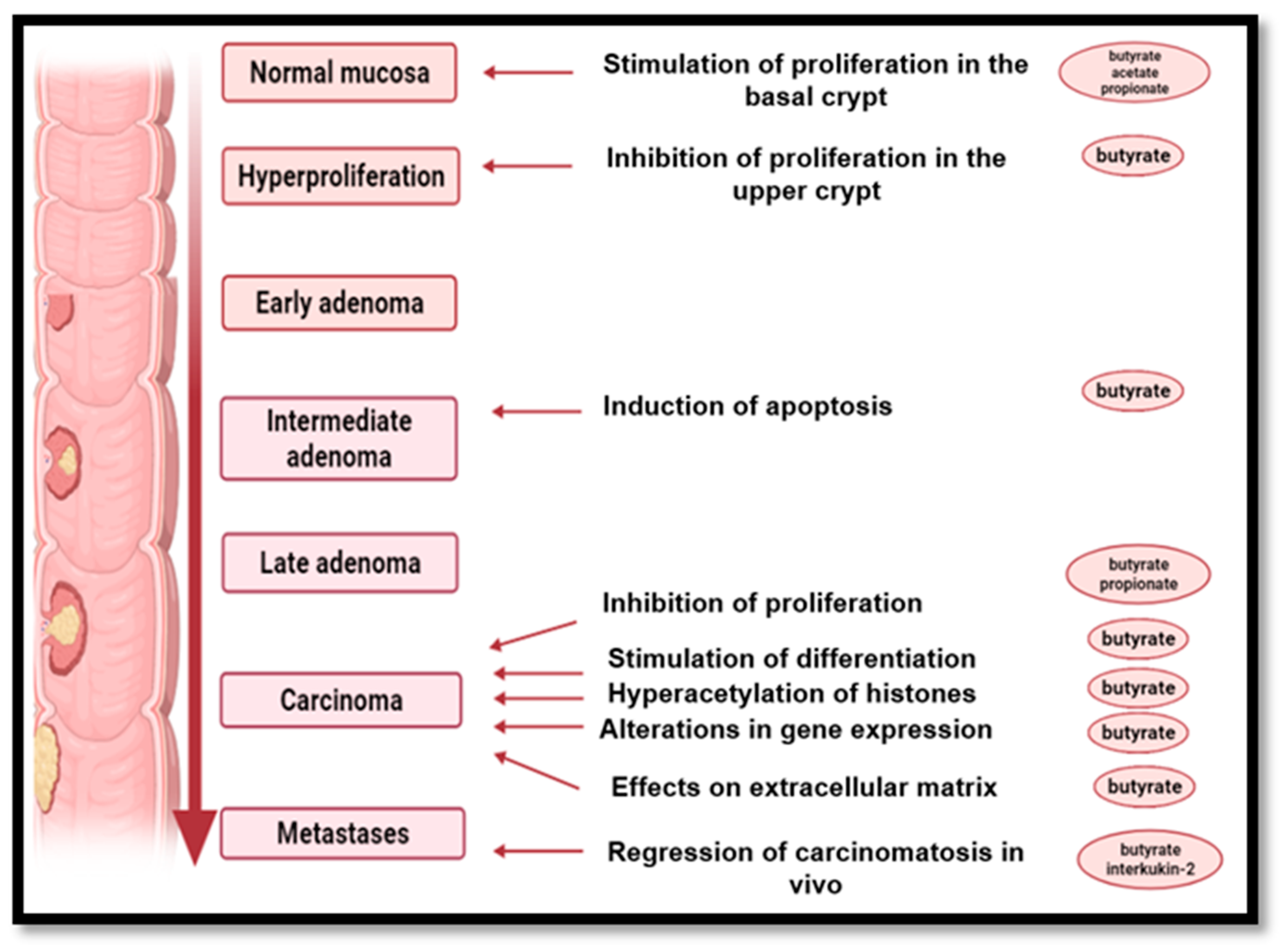
9. The Role of High-Fiber Diets in Cancer Prevention
- -
- insoluble fiber increases the mass of the fecal bowl, increase intestinal peristalsis and reduce the contact time of toxic compounds with the intestinal mucosa;
- -
- soluble fiber in particular, but also resistant starch, modify the fecal microorganisms and increase the number of saprotrophic bacteria, which exert a beneficial effect on the intestinal microbiome by SCFAs generated by the fermentation process while causing a decrease in pH, thus inhibiting pH-sensitive, potentially pathogenic bacterial species that could lead to potentially carcinogenic compounds;
- -
- production of butyrate by fermentation of soluble fiber as well as resistant starch delays the proliferation of malignant cells, reduces inflammatory processes and promotes DNA regeneration;
- -
- reducing the percentage of body fat, contributes to the decrease of estrogen secretion due to decreasing in the number of secretory adipocytes. The consumption of NDF protects against breast and endometrial cancer by binding estrogen to the colon and increasing its fecal elimination. Along with fiber and other constituents of vegetables, fruits and whole grains protect against cancer, especially compounds with an antioxidant action (flavones, polyphenolic compounds, anthocyanins, carotenoid pigments, etc.) [182].
10. Dietary Fiber Consumption Restrictions
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hansel, B. Que Conseiller Aux Patients Diabétiques de Type 2: Deux Ou Six Prises Alimentaires Quotidiennes? Médecine Des Mal. Métaboliques 2015, 9, 33–34. [Google Scholar] [CrossRef]
- Bureau of Nutritional Sciences Food Directorate, Health Products and Food Branch, Health Canada. Policy for Labelling and Advertising of Dietary Fibre-Containing Food. Available online: http://www.hc-sc.gc.ca (accessed on 19 February 2022).
- Institute of Medicine (IOM), U.S. National Academy of Sciences. Dietary Reference Intakes; National Academies Press: Washington, DC, USA, 2001; ISBN 978-0-309-07564-0. [Google Scholar]
- Cheong, T. Why Is Dietary Fibre So Important? Singapore Health Promotion Board. Available online: http://www.healthxchange.com.sg/healthyliving/DietandNutrition/Pages/Why-Is-Dietary-Fibre-So-Important.aspx (accessed on 21 February 2022).
- Crawford, M.; Bell, G.; Bueno, A.; Cunnane, S.; Clough, P.; Emmett, P.; Galli, C.; Golding, J.; Harbige, L.; Hibbeln, J.R.; et al. Outcome of the Public Consultation on the Draft Opinion of the Scientific Panel on Dietetic Products, Nutrition, and Allergies (NDA) on Dietary Reference Values for Carbohydrates and Dietary Fibre. EFSA J. 2010, 8, 1507. [Google Scholar] [CrossRef] [Green Version]
- Nishida, C.; Uauy, R.; Kumanyika, S.; Shetty, P. The Joint WHO/FAO Expert Consultation on Diet, Nutrition and the Prevention of Chronic Diseases: Process, Product and Policy Implications. Public Health Nutr. 2004, 7, 245–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keefe, S.J. The Association between Dietary Fibre Deficiency and High-Income Lifestyle-Associated Diseases: Burkitt’s Hypothesis Revisited. Lancet Gastroenterol. Hepatol. 2019, 4, 984–996. [Google Scholar] [CrossRef]
- Slavin, J.L. Dietary Fiber and Body Weight. Nutrition 2005, 21, 411–418. [Google Scholar] [CrossRef]
- Hipsley, E.H. Dietary “Fibre” and Pregnancy Toxaemia. BMJ 1953, 2, 420–422. [Google Scholar] [CrossRef]
- Sharma, S.K.; Bansal, S.; Mangal, M.; Dixit, A.K.; Gupta, R.K.; Mangal, A.K. Utilization of Food Processing By-Products as Dietary, Functional, and Novel Fiber: A Review. Crit. Rev. Food Sci. Nutr. 2016, 56, 1647–1661. [Google Scholar] [CrossRef]
- Jones, J.M. New AACC Dietary Fiber Definition Sets the Stage for Annual Meeting Technical Round Tables. Cereal Foods World 2000, 45, 404. [Google Scholar]
- Jones, J. Update on Defining Dietary Fiber. Cereal Foods World 2000, 45, 219–220. [Google Scholar]
- DeVries, J.W.; Camire, M.E.; Cho, S.; Craig, S.; Gordon, D.; Jones, J.M.; Li, B.; Lineback, D.; Prosky, L.; Tungland, B.C. The Definition of Dietary Fiber. Cereal Foods World 2001, 46, 112–129. [Google Scholar]
- Hussain, S.; Jõudu, I.; Bhat, R. Dietary Fiber from Underutilized Plant Resources-A Positive Approach for Valorization of Fruit and Vegetable Wastes. Sustainability 2020, 12, 5401. [Google Scholar] [CrossRef]
- Kochen, M. Dietary Fiber. West. J. Med. 1979, 130, 375–376. [Google Scholar] [CrossRef] [PubMed]
- Stephen, A.M.; Champ, M.M.-J.; Cloran, S.J.; Fleith, M.; van Lieshout, L.; Mejborn, H.; Burley, V.J. Dietary Fibre in Europe: Current State of Knowledge on Definitions, Sources, Recommendations, Intakes and Relationships to Health. Nutr. Res. Rev. 2017, 30, 149–190. [Google Scholar] [CrossRef] [PubMed]
- “10 Dietary Fiber.” National Research Council. Diet and Health; National Academies Press: Washington, DC, USA, 1989; ISBN 978-0-309-03994-9. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, P.D.; Pawar, H.A. Recently Investigated Natural Gums and Mucilages as Pharmaceutical Excipients: An Overview. J. Pharm. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Linus Pauling Institute, Micronutrient Information Center, Oregon State University. Fiber. Available online: https://lpi.oregonstate.edu/mic/other-nutrients/fiber#authors-reviewers (accessed on 25 February 2022).
- Lan, Y. Waxes. In Encyclopedia of Food Chemistry; Elsevier: Amsterdam, The Netherlands, 2019; pp. 312–316. [Google Scholar]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant Food Anti-Nutritional Factors and Their Reduction Strategies: An Overview. Food Prod. Process. Nutr. 2020, 2, 6. [Google Scholar] [CrossRef]
- Williams, B.A.; Mikkelsen, D.; Flanagan, B.M.; Gidley, M.J. “Dietary Fibre”: Moving beyond the “Soluble/Insoluble” Classification for Monogastric Nutrition, with an Emphasis on Humans and Pigs. J. Anim. Sci. Biotechnol. 2019, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Cui, L.; Qi, J.; Ojo, O.; Du, X.; Liu, Y.; Wang, X. The Effect of Dietary Fiber (Oat Bran) Supplement on Blood Pressure in Patients with Essential Hypertension: A Randomized Controlled Trial. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2458–2470. [Google Scholar] [CrossRef]
- Topping, D.L.; Fukushima, M.; Bird, A.R. Resistant Starch as a Prebiotic and Synbiotic: State of the Art. Proc. Nutr. Soc. 2003, 62, 171–176. [Google Scholar] [CrossRef] [Green Version]
- US Department of Agriculture, National Agricultural Library and National Academy of Sciences, Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; National Academies Press: Washington, DC, USA, 2005; ISBN 978-0-309-08525-0. [Google Scholar] [CrossRef]
- Yang, J. Effect of Dietary Fiber on Constipation: A Meta Analysis. World J. Gastroenterol. 2012, 18, 7378. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health Benefits of Fruits and Vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Martin, A. The “Apports Nutritionnels Conseillés (ANC)” for the French Population. Reprod. Nutr. Dev. 2001, 41, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Boclé, J.-C.; Champ, M.; Berta, J.-L. Les Fibres Alimentaires: Déterminants Physico-Chimiques, Définition, Aspects Analytiques et Physiologiques. Cah. Nutr. Diététique 2005, 40, 15–21. [Google Scholar] [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of Dietary Fiber and Its Components on Metabolic Health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holscher, H.D. Dietary Fiber and Prebiotics and the Gastrointestinal Microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The Short-Chain Fatty Acid Acetate Reduces Appetite via a Central Homeostatic Mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [Green Version]
- Mcrorie, J.W.; Fahey, G.C. A Review of Gastrointestinal Physiology and the Mechanisms Underlying the Health Benefits of Dietary Fiber: Matching an Effective Fiber with Specific Patient Needs. Clin. Nurs. Stud. 2013, 1, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.-W.; Cephas, K.D.; Holscher, H.D.; Kerr, K.R.; Mangian, H.F.; Tappenden, K.A.; Swanson, K.S. Nondigestible Fructans Alter Gastrointestinal Barrier Function, Gene Expression, Histomorphology, and the Microbiota Profiles of Diet-Induced Obese C57BL/6J Mice. J. Nutr. 2016, 146, 949–956. [Google Scholar] [CrossRef]
- McRorie, J.W. Psyllium Is Not Fermented in the Human Gut. Neurogastroenterol. Motil. 2015, 27, 1681–1682. [Google Scholar] [CrossRef]
- Holscher, H.D.; Doligale, J.L.; Bauer, L.L.; Gourineni, V.; Pelkman, C.L.; Fahey, G.C.; Swanson, K.S. Gastrointestinal Tolerance and Utilization of Agave Inulin by Healthy Adults. Food Funct. 2014, 5, 1142. [Google Scholar] [CrossRef]
- Rendón-Huerta, J.A.; Juárez-Flores, B.; Pinos-Rodríguez, J.M.; Aguirre-Rivera, J.R.; Delgado-Portales, R.E. Effects of Different Sources of Fructans on Body Weight, Blood Metabolites and Fecal Bacteria in Normal and Obese Non-Diabetic and Diabetic Rats. Plant Foods Hum. Nutr. 2012, 67, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, M.; Kotlo, K.U.; Dudeja, P.K.; Layden, B.T. Role of Short Chain Fatty Acid Receptors in Intestinal Physiology and Pathophysiology. In Comprehensive Physiology; Wiley: Hoboken, NJ, USA, 2018; pp. 1091–1115. [Google Scholar]
- Layden, B.T.; Angueira, A.R.; Brodsky, M.; Durai, V.; Lowe, W.L. Short Chain Fatty Acids and Their Receptors: New Metabolic Targets. Transl. Res. 2013, 161, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Ohira, H.; Tsutsui, W.; Mamoto, R.; Yamaguchi, S.; Nishida, M.; Ito, M.; Fujioka, Y. Butyrate Attenuates Lipolysis in Adipocytes Co-Cultured with Macrophages through Non-Prostaglandin E2–Mediated and Prostaglandin E2–Mediated Pathways. Lipids Health Dis. 2016, 15, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Curto, E.; Milligan, G. Metabolism Meets Immunity: The Role of Free Fatty Acid Receptors in the Immune System. Biochem. Pharmacol. 2016, 114, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Mulders, R.J.; de Git, K.C.G.; Schéle, E.; Dickson, S.L.; Sanz, Y.; Adan, R.A.H. Microbiota in Obesity: Interactions with Enteroendocrine, Immune and Central Nervous Systems. Obes. Rev. 2018, 19, 435–451. [Google Scholar] [CrossRef]
- Borgeraas, H.; Johnson, L.K.; Skattebu, J.; Hertel, J.K.; Hjelmesaeth, J. Effects of Probiotics on Body Weight, Body Mass Index, Fat Mass and Fat Percentage in Subjects with Overweight or Obesity: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Obes. Rev. 2018, 19, 219–232. [Google Scholar] [CrossRef] [Green Version]
- Lazar, V.; Ditu, L.-M.; Pircalabioru, G.G.; Picu, A.; Petcu, L.; Cucu, N.; Chifiriuc, M.C. Gut Microbiota, Host Organism, and Diet Trialogue in Diabetes and Obesity. Front. Nutr. 2019, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Aziz, Q.; Doré, J.; Emmanuel, A.; Guarner, F.; Quigley, E.M.M. Gut Microbiota and Gastrointestinal Health: Current Concepts and Future Directions. Neurogastroenterol. Motil. 2013, 25, 4–15. [Google Scholar] [CrossRef]
- Pulikkan, J.; Mazumder, A.; Grace, T. Role of the Gut Microbiome in Autism Spectrum Disorders. In Reviews on Biomarker Studies in Psychiatric and Neurodegenerative Disorders; Springer: Berlin/Heidelberg, Germany, 2019; pp. 253–269. [Google Scholar]
- Mosoni, P. Dégradation Des Fibres Alimentaires Par Le Microbiote Colique de l’ Homme. Innov. Agron. INRAE 2014, 36, 83–96. [Google Scholar]
- Ioniță, A.C. In Vitro Effects of Some Synthesized Aminoacetanilide N’-Substituted on Human Leukocytes Separated From Peripheral Blood. Farmacia 2019, 67, 684–690. [Google Scholar] [CrossRef]
- Kieffer, D.A.; Martin, R.J.; Adams, S.H. Impact of Dietary Fibers on Nutrient Management and Detoxification Organs: Gut, Liver, and Kidneys. Adv. Nutr. An Int. Rev. J. 2016, 7, 1111–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howlett, J.; Betteridge, V.; Champ, M.; Craig, S.S.; Meheust, A.; Jones, J.M. The Definition of Dietary Fiber—Discussions at the Ninth Vahouny Fiber Symposium: Building Scientific Agreement. Food Nutr. Res. 2010, 54, 5750. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic Analysis of the Human Distal Gut Microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Donohoe, D.R.; Collins, L.B.; Wali, A.; Bigler, R.; Sun, W.; Bultman, S.J. The Warburg Effect Dictates the Mechanism of Butyrate-Mediated Histone Acetylation and Cell Proliferation. Mol. Cell 2012, 48, 612–626. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; Leroy, F. Cross-Feeding between Bifidobacteria and Butyrate-Producing Colon Bacteria Explains Bifdobacterial Competitiveness, Butyrate Production, and Gas Production. Int. J. Food Microbiol. 2011, 149, 73–80. [Google Scholar] [CrossRef]
- Goldsmith, J.R.; Sartor, R.B. The Role of Diet on Intestinal Microbiota Metabolism: Downstream Impacts on Host Immune Function and Health, and Therapeutic Implications. J. Gastroenterol. 2014, 49, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Aponte, M.; Murru, N.; Shoukat, M. Therapeutic, Prophylactic, and Functional Use of Probiotics: A Current Perspective. Front. Microbiol. 2020, 11, 562048. [Google Scholar] [CrossRef]
- Ilie, C.-I.; Oprea, E.; Geana, E.-I.; Spoiala, A.; Buleandra, M.; Gradisteanu Pircalabioru, G.; Badea, I.A.; Ficai, D.; Andronescu, E.; Ficai, A.; et al. Bee Pollen Extracts: Chemical Composition, Antioxidant Properties, and Effect on the Growth of Selected Probiotic and Pathogenic Bacteria. Antioxidants 2022, 11, 959. [Google Scholar] [CrossRef]
- Bakour, M.; Laaroussi, H.; Ousaaid, D.; El Ghouizi, A.; Es-Safi, I.; Mechchate, H.; Lyoussi, B. Bee Bread as a Promising Source of Bioactive Molecules and Functional Properties: An Up-To-Date Review. Antibiotics 2022, 11, 203. [Google Scholar] [CrossRef]
- Miryan, M.; Alavinejad, P.; Abbaspour, M.; Soleimani, D.; Ostadrahimi, A. Does Propolis Affect the Quality of Life and Complications in Subjects with Irritable Bowel Syndrome (Diagnosed with Rome IV Criteria)? A Study Protocol of the Randomized, Double-Blinded, Placebo-Controlled Clinical Trial. Trials 2020, 21, 698. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, G.E.G.; Mamdouh, A.M.; Eman, M.G.; Jihan, S.H. The Effect of Honey Supplementation on Plasma Levels of Short Chain Fatty Acids in Healthy Infants. Med. J. Cairo Univ. 2021, 89, 499–504. [Google Scholar] [CrossRef]
- de Melo, F.H.C.; Menezes, F.N.D.D.; de Sousa, J.M.B.; dos Santos Lima, M.; da Silva Campelo Borges, G.; de Souza, E.L.; Magnani, M. Prebiotic Activity of Monofloral Honeys Produced by Stingless Bees in the Semi-Arid Region of Brazilian Northeastern toward Lactobacillus Acidophilus LA-05 and Bifidobacterium Lactis BB-12. Food Res. Int. 2020, 128, 108809. [Google Scholar] [CrossRef] [PubMed]
- Kajiwara, S.; Gandhi, H.; Ustunol, Z. Effect of Honey on the Growth of and Acid Production by Human Intestinal Bifidobacterium Spp.: An In Vitro Comparison with Commercial Oligosaccharides and Inulin. J. Food Prot. 2002, 65, 214–218. [Google Scholar] [CrossRef]
- Afrin, S.; Haneefa, S.M.; Fernandez-Cabezudo, M.J.; Giampieri, F.; Al-Ramadi, B.K.; Battino, M. Therapeutic and Preventive Properties of Honey and Its Bioactive Compounds in Cancer: An Evidence-Based Review. Nutr. Res. Rev. 2020, 33, 50–76. [Google Scholar] [CrossRef] [PubMed]
- Erejuwa, O.; Sulaiman, S.; Wahab, M. Effects of Honey and Its Mechanisms of Action on the Development and Progression of Cancer. Molecules 2014, 19, 2497–2522. [Google Scholar] [CrossRef] [Green Version]
- Dreher, M. Whole Fruits and Fruit Fiber Emerging Health Effects. Nutrients 2018, 10, 1833. [Google Scholar] [CrossRef] [Green Version]
- USDA. National Nutrient Database for Standard Reference. 2018. Available online: https://fdc.nal.usda.gov/ (accessed on 25 February 2022).
- De Vadder, F.; Mithieux, G. Les Fibres Alimentaires Induisent Des Bénéfices Métaboliques via l’activation de La Néoglucogenèse Intestinale. Obésité 2014, 9, 280–285. [Google Scholar] [CrossRef]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between Diet, Gut Microbiota Composition and Gut Metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Iljazovic, A.; Roy, U.; Gálvez, E.J.C.; Lesker, T.R.; Zhao, B.; Gronow, A.; Amend, L.; Will, S.E.; Hofmann, J.D.; Pils, M.C.; et al. Perturbation of the Gut Microbiome by Prevotella Spp. Enhances Host Susceptibility to Mucosal Inflammation. Mucosal Immunol. 2021, 14, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Franke, T.; Deppenmeier, U. Physiology and Central Carbon Metabolism of the Gut Bacterium Prevotella Copri. Mol. Microbiol. 2018, 109, 528–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frame, L.A.; Costa, E.; Jackson, S.A. Current Explorations of Nutrition and the Gut Microbiome: A Comprehensive Evaluation of the Review Literature. Nutr. Rev. 2020, 78, 798–812. [Google Scholar] [CrossRef]
- Gerasimidis, K.; Nichols, B.; McGowan, M.; Svolos, V.; Papadopoulou, R.; Kokkorou, M.; Rebull, M.; Bello Gonzalez, T.; Hansen, R.; Russell, R.K.; et al. P705 The Effects of Commonly Consumed Dietary Fibres on the Gut Microbiome and Its Fibre Fermentative Capacity in Adults with Inflammatory Bowel Disease in Remission. J. Crohn’s Colitis 2022, 16, i602. [Google Scholar] [CrossRef]
- Nenciu, D.S. The Structure Of Romanian’s Food Consumption and Its Implications on Health Condition and Quality Of Life. Rev. Română Stat. 2014, 61, 41–51. [Google Scholar]
- Cînpeanu, O.-C.; Tarcea, M.; Cojan, P.; Iorga, D.; Olah, P.; Guiné, R.P.F. Perception of Healthy Eating among Romanian Adults. J. Interdiscip. Med. 2019, 4, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Mititelu, M. The Impact of COVID-19 Lockdown on the Lifestyle and Dietary Patterns Among Romanian Population. Farmacia 2021, 69, 1–11. [Google Scholar] [CrossRef]
- Năstăsescu, V.; Mititelu, M.; Stanciu, T.I.; Drăgănescu, D.; Grigore, N.D.; Udeanu, D.I.; Stanciu, G.; Neacșu, S.M.; Dinu-Pîrvu, C.E.; Oprea, E.; et al. Food Habits and Lifestyle of Romanians in the Context of the COVID-19 Pandemic. Nutrients 2022, 14, 504. [Google Scholar] [CrossRef]
- La Santé Vient en Mangeant. Document d’accompagnement du Guide Alimentaire Pour Tous Destiné aux Professionnels de Santé. Programme National Nutrition-Santé. September 2002. Available online: https://solidarites-sante.gouv.fr/IMG/pdf/guide_alimentairetous.pdf (accessed on 25 February 2022).
- World Health Organization. A Guide for Population-Based Approaches to Increasing Levels of Physical Activity: Implementation of the WHO Global Strategy on Diet, Physical Activity and Health; World Health Organization: Geneva, Switzerland, 2007. Available online: https://apps.who.int/iris/handle/10665/43612 (accessed on 25 February 2022).
- Quagliani, D.; Felt-Gunderson, P. Closing America’s Fiber Intake Gap. Am. J. Lifestyle Med. 2017, 11, 80–85. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture, Agricultural Research Service. Nutrient Intakes from Food: Mean Amounts Consumed per Individual, by Gender and Age, What We Eat in America, NHANES 2009–2010. 2012. Available online: www.ars.usda.gov/ba/bhnrc/fsrg (accessed on 25 February 2022).
- Jin, J. Dietary Guidelines for Americans. JAMA-J. Am. Med. Assoc. 2016, 315, 528. [Google Scholar] [CrossRef] [Green Version]
- Lăcătușu, C.-M.; Grigorescu, E.-D.; Floria, M.; Onofriescu, A.; Mihai, B.-M. The Mediterranean Diet: From an Environment-Driven Food Culture to an Emerging Medical Prescription. Int. J. Environ. Res. Public Health 2019, 16, 942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Wang, Z.; Fei, Y.; Zhou, B.; Zheng, S.; Wang, L.; Huang, L.; Jiang, S.; Liu, Z.; Jiang, J.; et al. The Difference in Nutrient Intakes between Chinese and Mediterranean, Japanese and American Diets. Nutrients 2015, 7, 4661–4688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trichopoulou, A.; Costacou, T.; Bamia, C.; Trichopoulos, D. Adherence to a Mediterranean Diet and Survival in a Greek Population. N. Engl. J. Med. 2003, 348, 2599–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ros, E.; Martínez-González, M.A.; Estruch, R.; Salas-Salvadó, J.; Fitó, M.; Martínez, J.A.; Corella, D. Mediterranean Diet and Cardiovascular Health: Teachings of the PREDIMED Study. Adv. Nutr. 2014, 5, 330S–336S. [Google Scholar] [CrossRef] [Green Version]
- Kopel, E.; Sidi, Y.; Kivity, S. Mediterranean Diet for Primary Prevention of Cardiovascular Disease. N. Engl. J. Med. 2013, 369, 672–677. [Google Scholar] [CrossRef]
- Belahsen, R.; Rguibi, M. Population Health and Mediterranean Diet in Southern Mediterranean Countries. Public Health Nutr. 2006, 9, 1130–1135. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.B.; Dubnov, G.; Niaz, M.A.; Ghosh, S.; Singh, R.; Rastogi, S.S.; Manor, O.; Pella, D.; Berry, E.M. Effect of an Indo-Mediterranean Diet on Progression of Coronary Artery Disease in High Risk Patients (Indo-Mediterranean Diet Heart Study): A Randomised Single-Blind Trial. Lancet 2002, 360, 1455–1461. [Google Scholar] [CrossRef]
- Indian Dietetic Association Working Committee on Position Paper on Dietary Fibre. Position of the Indian Dietetic Association: Dietary Fibre and Health. December 2018, pp. 1–14. Available online: http://idaindia.com/wp-content/uploads/2018/12/IDA-position-paper-fibre-24.12.18.pdf (accessed on 25 February 2022).
- Saidane, O.; Courties, A.; Sellam, J. Les Fibres Alimentaires Contre l’arthrose: Quels Éléments de Preuve? Rev. Rhum. 2020, 87, 4–7. [Google Scholar] [CrossRef]
- Queenan, K.M.; Stewart, M.L.; Smith, K.N.; Thomas, W.; Fulcher, R.G.; Slavin, J.L. Concentrated Oat β-Glucan, a Fermentable Fiber, Lowers Serum Cholesterol in Hypercholesterolemic Adults in a Randomized Controlled Trial. Nutr. J. 2007, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.L.; Steiniger, J.; Reich, S.C.; Weickert, M.O.; Harsch, I.; Machowetz, A.; Mohlig, M.; Spranger, J.; Rudovich, N.N.; Meuser, F.; et al. Arabinoxylan Fibre Consumption Improved Glucose Metabolism, but Did Not Affect Serum Adipokines in Subjects with Impaired Glucose Tolerance. Horm. Metab. Res. 2006, 38, 761–766. [Google Scholar] [CrossRef]
- Kapoor, M.P.; Ishihara, N.; Okubo, T. Soluble Dietary Fibre Partially Hydrolysed Guar Gum Markedly Impacts on Postprandial Hyperglycaemia, Hyperlipidaemia and Incretins Metabolic Hormones over Time in Healthy and Glucose Intolerant Subjects. J. Funct. Foods 2016, 24, 207–220. [Google Scholar] [CrossRef]
- Ganji, V.; Kuo, J. Serum Lipid Responses to Psyllium Fiber: Differences between Pre- and Post-Menopausal, Hypercholesterolemic Women. Nutr. J. 2008, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.S.; Johnson, S.K.; Baxter, A.L.; Ball, M.J. Lupin Kernel Fibre-Enriched Foods Beneficially Modify Serum Lipids in Men. Eur. J. Clin. Nutr. 2005, 59, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasir, O.; Babiker, S.; Salim, A.-M.M. Protective Effect of Gum Arabic Supplementation for Type-2 Diabetes Mellitus and Its Complications. Int. J. Multidiscip. Curr. Res. 2016, 4, 2321–3121. [Google Scholar]
- Jmal, L.; Jmal, A.; Abdennebi, M.; Feki, M.; Boukthir, S. Prevalence of Metabolic Syndrome in Tunisian Overweight and Obese Children. Tunis. Med. 2019, 97, 133–139. [Google Scholar]
- Mohamed, A.B.; Rémond, D.; Della-valle, G.; Chiron, H.; Cohade, B.; Béchet, D.; Coudy-Gandilhon, C.; David, J.; Dardevet, D.; Dore, J.; et al. Les Fibres Alimentaires Limitent Le Stockage de Lipides Hépatiques En Situation de Surnutrition: Quels Mécanismes et Quels Médiateurs? Nutr. Clin. Métabolisme 2018, 32, 280. [Google Scholar] [CrossRef]
- Pauline, K.-B.; Rimm, E.B. Whole Grain Consumption and Weight Gain: A Review of the Epidemiological Evidence, Potential Mechanisms and Opportunities for Future Research. Proc. Nutr. Soc. 2003, 62, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Kromhout, D.; Bloemberg, B.; Seidell, J.; Nissinen, A.; Menotti, A. Physical Activity and Dietary Fiber Determine Population Body Fat Levels: The Seven Countries Study. Int. J. Obes. 2001, 25, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Bajzer, M.; Seeley, R.J. Obesity and Gut Flora. Nature 2006, 444, 1009–1010. [Google Scholar] [CrossRef]
- Heiss, C.N.; Olofsson, L.E. Gut Microbiota-Dependent Modulation of Energy Metabolism. J. Innate Immun. 2018, 10, 163–171. [Google Scholar] [CrossRef]
- Cédrin-Durnerin, I.; Sonigo, C.; Seroka, A.; Comtet, M.; Vinolas, C.; Grynberg, M.; Sermondade, N.; Herbemont, C.; Sifer, C. Nutrition et Reproduction: Les Aspects Féminins. Obésité 2017, 12, 179–185. [Google Scholar] [CrossRef]
- Silvestris, E.; Lovero, D.; Palmirotta, R. Nutrition and Female Fertility: An Interdependent Correlation. Front. Endocrinol. 2019, 10, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinu, M.; Pagliai, G.; Casini, A.; Sofi, F. Mediterranean Diet and Multiple Health Outcomes: An Umbrella Review of Meta-Analyses of Observational Studies and Randomised Trials. Eur. J. Clin. Nutr. 2018, 72, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, R.A.; Palmer, D.J. High-Fiber Diet during Pregnancy Characterized by More Fruit and Vegetable Consumption. Nutrients 2021, 13, 35. [Google Scholar] [CrossRef]
- Toh, H.; Thomson, J.A.; Jiang, P. Maternal High-Fiber Diet Protects Offspring against Type 2 Diabetes. Nutrients 2020, 13, 94. [Google Scholar] [CrossRef]
- Cluny, N.L.; Eller, L.K.; Keenan, C.M.; Reimer, R.A.; Sharkey, K.A. Interactive Effects of Oligofructose and Obesity Predisposition on Gut Hormones and Microbiota in Diet-Induced Obese Rats. Obesity 2015, 23, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Prabhakar, M.; Ju, J.; Long, H.; Zhou, H.-W. Effect of Inulin-Type Fructans on Blood Lipid Profile and Glucose Level: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Eur. J. Clin. Nutr. 2017, 71, 9–20. [Google Scholar] [CrossRef]
- Adam, C.L.; Gratz, S.W.; Peinado, D.I.; Thomson, L.M.; Garden, K.E.; Williams, P.A.; Richardson, A.J.; Ross, A.W. Effects of Dietary Fibre (Pectin) and/or Increased Protein (Casein or Pea) on Satiety, Body Weight, Adiposity and Caecal Fermentation in High Fat Diet-Induced Obese Rats. PLoS ONE 2016, 11, e0155871. [Google Scholar] [CrossRef]
- Adam, C.L.; Thomson, L.M.; Williams, P.A.; Ross, A.W. Soluble Fermentable Dietary Fibre (Pectin) Decreases Caloric Intake, Adiposity and Lipidaemia in High-Fat Diet-Induced Obese Rats. PLoS ONE 2015, 10, e0140392. [Google Scholar] [CrossRef] [Green Version]
- Bodinham, C.L.; Smith, L.; Thomas, E.L.; Bell, J.D.; Swann, J.R.; Costabile, A.; Russell-Jones, D.; Umpleby, A.M.; Robertson, M.D. Efficacy of Increased Resistant Starch Consumption in Human Type 2 Diabetes. Endocr. Connect. 2014, 3, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.A.; Raine, L.B.; Drollette, E.S.; Scudder, M.R.; Kramer, A.F.; Hillman, C.H. Dietary Fiber Is Positively Associated with Cognitive Control among Prepubertal Children. World Rev. Nutr. Diet. 2016, 114, 88–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bretin, A.; Chassaing, B. Inuline: La Belle et La Bête. Médecine/Sci. 2019, 35, 100–102. [Google Scholar] [CrossRef] [PubMed]
- Schulze, M.B. Fiber and Magnesium Intake and Incidence of Type 2 Diabetes. Arch. Intern. Med. 2007, 167, 956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slavin, J.; Green, H. Dietary Fibre and Satiety. Nutr. Bull. 2007, 32, 32–42. [Google Scholar] [CrossRef]
- Chaudhri, O.B.; Salem, V.; Murphy, K.G.; Bloom, S.R. Gastrointestinal Satiety Signals. Annu. Rev. Physiol. 2008, 70, 239–255. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Hoffmann, G.; Lampousi, A.-M.; Knüppel, S.; Iqbal, K.; Schwedhelm, C.; Bechthold, A.; Schlesinger, S.; Boeing, H. Food Groups and Risk of Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Prospective Studies. Eur. J. Epidemiol. 2017, 32, 363–375. [Google Scholar] [CrossRef] [Green Version]
- Widmer, R.J.; Flammer, A.J.; Lerman, L.O.; Lerman, A. The Mediterranean Diet, Its Components, and Cardiovascular Disease. Am. J. Med. 2015, 128, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Kelly, T.; Unwin, D.; Finucane, F. Low-Carbohydrate Diets in the Management of Obesity and Type 2 Diabetes: A Review from Clinicians Using the Approach in Practice. Int. J. Environ. Res. Public Health 2020, 17, 2557. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Cortegana, C.; Iglesias, P.; Ribalta, J.; Vilariño-García, T.; Montañez, L.; Arrieta, F.; Aguilar, M.; Durán, S.; Obaya, J.C.; Becerra, A.; et al. Nutrients and Dietary Approaches in Patients with Type 2 Diabetes Mellitus and Cardiovascular Disease: A Narrative Review. Nutrients 2021, 13, 4150. [Google Scholar] [CrossRef]
- Schlienger, J.-L. Prise En Charge Nutritionnelle Dans La Prévention et Le Traitement Des Maladies Chroniques. Médecine Des Mal. Métaboliques 2017, 11, 258–265. [Google Scholar] [CrossRef]
- Mithieux, G. Dialogue Intestin-Cerveau via La Néoglucogenèse Intestinale. Médecine Des Mal. Métaboliques 2018, 12, 650–656. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Generated Metabolites Promote Metabolic Benefits via Gut-Brain Neural Circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinet, F.; Taki, A.; Raffin, M.; De Vadder, F.; Mithieux, G.; Gautier-Stein, A. Bénéfices Des Fibres Alimentaires Sur Le Comportement Émotionnel. Ann. Endocrinol. 2015, 76, 375. [Google Scholar] [CrossRef]
- Parks, B.W.; Nam, E.; Org, E.; Kostem, E.; Norheim, F.; Hui, S.T.; Pan, C.; Civelek, M.; Rau, C.D.; Bennett, B.J.; et al. Genetic Control of Obesity and Gut Microbiota Composition in Response to High-Fat, High-Sucrose Diet in Mice. Cell Metab. 2013, 17, 141–152. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut Metagenome in European Women with Normal, Impaired and Diabetic Glucose Control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef]
- Yao, B.; Fang, H.; Xu, W.; Yan, Y.; Xu, H.; Liu, Y.; Mo, M.; Zhang, H.; Zhao, Y. Dietary Fiber Intake and Risk of Type 2 Diabetes: A Dose–Response Analysis of Prospective Studies. Eur. J. Epidemiol. 2014, 29, 79–88. [Google Scholar] [CrossRef]
- Silva, F.M.; Kramer, C.K.; de Almeida, J.C.; Steemburgo, T.; Gross, J.L.; Azevedo, M.J. Fiber Intake and Glycemic Control in Patients with Type 2 Diabetes Mellitus: A Systematic Review with Meta-Analysis of Randomized Controlled Trials. Nutr. Rev. 2013, 71, 790–801. [Google Scholar] [CrossRef]
- Thompson, S.V.; Hannon, B.A.; An, R.; Holscher, H.D. Effects of Isolated Soluble Fiber Supplementation on Body Weight, Glycemia, and Insulinemia in Adults with Overweight and Obesity: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Am. J. Clin. Nutr. 2017, 106, 1514–1528. [Google Scholar] [CrossRef] [Green Version]
- McRae, M.P. Dietary Fiber Intake and Type 2 Diabetes Mellitus: An Umbrella Review of Meta-Analyses. J. Chiropr. Med. 2018, 17, 44–53. [Google Scholar] [CrossRef]
- Sandberg, J.C.; Björck, I.M.E.; Nilsson, A.C. Effects of Whole Grain Rye, with and without Resistant Starch Type 2 Supplementation, on Glucose Tolerance, Gut Hormones, Inflammation and Appetite Regulation in an 11–14.5 Hour Perspective; a Randomized Controlled Study in Healthy Subjects. Nutr. J. 2017, 16, 25. [Google Scholar] [CrossRef]
- Wu, X.; Chen, D.; Yu, B.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; He, J. Effect of Different Dietary Non-Starch Fiber Fractions on Growth Performance, Nutrient Digestibility, and Intestinal Development in Weaned Pigs. Nutrition 2018, 51–52, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Ding, M.; Sampson, L.; Willett, W.C.; Manson, J.E.; Wang, M.; Rosner, B.; Hu, F.B.; Sun, Q. Intake of Whole Grain Foods and Risk of Type 2 Diabetes: Results from Three Prospective Cohort Studies. BMJ 2020, 370, m2206. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The Role of Short-Chain Fatty Acids in Microbiota–Gut–Brain Communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, M.W.; Alim, I.; Bultman, S.J.; Ratan, R.R. Butyrate, Neuroepigenetics and the Gut Microbiome: Can a High Fiber Diet Improve Brain Health? Neurosci. Lett. 2016, 625, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanko, D.P.; Barrett, R.M.; Ly, A.R.; Reolon, G.K.; Wood, M.A. Modulation of Long-Term Memory for Object Recognition via HDAC Inhibition. Proc. Natl. Acad. Sci. USA 2009, 106, 9447–9452. [Google Scholar] [CrossRef] [Green Version]
- Coudron, O. Rôle de La Nutrition et de La Micronutrition Dans La Lutte Contre Les Troubles de l’humeur. Actual. Pharm. 2019, 58, 33–38. [Google Scholar] [CrossRef]
- Tengeler, A.C.; Kozicz, T.; Kiliaan, A.J. Relationship between Diet, the Gut Microbiota, and Brain Function. Nutr. Rev. 2018, 76, 603–617. [Google Scholar] [CrossRef]
- Selkrig, J.; Wong, P.; Zhang, X.; Pettersson, S. Metabolic Tinkering by the Gut Microbiome. Gut Microbes 2014, 5, 369–380. [Google Scholar] [CrossRef]
- Shi, H.; Ge, X.; Ma, X.; Zheng, M.; Cui, X.; Pan, W.; Zheng, P.; Yang, X.; Zhang, P.; Hu, M.; et al. A Fiber-Deprived Diet Causes Cognitive Impairment and Hippocampal Microglia-Mediated Synaptic Loss through the Gut Microbiota and Metabolites. Microbiome 2021, 9, 223. [Google Scholar] [CrossRef]
- Smith, A.; Sutherland, D.; Hewlett, P. An Investigation of the Acute Effects of Oligofructose-Enriched Inulin on Subjective Wellbeing, Mood and Cognitive Performance. Nutrients 2015, 7, 8887–8896. [Google Scholar] [CrossRef]
- Subash, S.; Essa, M.; Braidy, N.; Awlad-Thani, K.; Vaishnav, R.; Al-Asmi, A.; Guillemin, G.; Al-Adawi, S.; Subash, S. Diet Rich in Date Palm Fruits Improves Memory, Learning and Reduces Beta Amyloid in Transgenic Mouse Model of Alzheimer′s Disease. J. Ayurveda Integr. Med. 2015, 6, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, L.L.; Quigley, E.M.M.; Pfeiffer, R.F. Gastrointestinal Dysfunction in Parkinson’s Disease: Frequency and Pathophysiology. Neurology 1992, 42, 726. [Google Scholar] [CrossRef] [PubMed]
- Cantu-Jungles, T.M.; Rasmussen, H.E.; Hamaker, B.R. Potential of Prebiotic Butyrogenic Fibers in Parkinson’s Disease. Front. Neurol. 2019, 10, 663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Li, B.; Lou, P.; Dai, T.; Chen, Y.; Zhuge, A.; Yuan, Y.; Li, L. The Relationship Between the Gut Microbiome and Neurodegenerative Diseases. Neurosci. Bull. 2021, 37, 1510–1522. [Google Scholar] [CrossRef]
- Lorente-Picón, M.; Laguna, A. New Avenues for Parkinson’s Disease Therapeutics: Disease-Modifying Strategies Based on the Gut Microbiota. Biomolecules 2021, 11, 433. [Google Scholar] [CrossRef]
- Appiakannan, H.S.; Rasimowicz, M.L.; Harrison, C.B.; Weber, E.T. Differential Effects of High-Fat Diet on Glucose Tolerance, Food Intake, and Glucocorticoid Regulation in Male C57BL/6J and BALB/CJ Mice. Physiol. Behav. 2020, 215, 112773. [Google Scholar] [CrossRef]
- Forte, N.; Fernández-Rilo, A.C.; Palomba, L.; Di Marzo, V.; Cristino, L. Obesity Affects the Microbiota–Gut–Brain Axis and the Regulation Thereof by Endocannabinoids and Related Mediators. Int. J. Mol. Sci. 2020, 21, 1554. [Google Scholar] [CrossRef] [Green Version]
- Holzer, P. Neuropeptides, Microbiota, and Behavior. Int. Rev. Neurobiol. 2016, 131, 67–89. [Google Scholar]
- Schmidt, K.; Cowen, P.J.; Harmer, C.J.; Tzortzis, G.; Errington, S.; Burnet, P.W.J. Prebiotic Intake Reduces the Waking Cortisol Response and Alters Emotional Bias in Healthy Volunteers. Psychopharmacology 2015, 232, 1793–1801. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.W.; Allgood, L.D.; Lawrence, A.; Altringer, L.A.; Jerdack, G.R.; Hengehold, D.A.; Morel, J.G. Cholesterol-Lowering Effects of Psyllium Intake Adjunctive to Diet Therapy in Men and Women with Hypercholesterolemia: Meta-Analysis of 8 Controlled Trials. Am. J. Clin. Nutr. 2000, 71, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, D.J.; Kendall, C.W.; Vuksan, V.; Vidgen, E.; Parker, T.; Faulkner, D.; Mehling, C.C.; Garsetti, M.; Testolin, G.; Cunnane, S.C.; et al. Soluble Fiber Intake at a Dose Approved by the US Food and Drug Administration for a Claim of Health Benefits: Serum Lipid Risk Factors for Cardiovascular Disease Assessed in a Randomized Controlled Crossover Trial. Am. J. Clin. Nutr. 2002, 75, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Ros, E.; Hu, F.B. Consumption of Plant Seeds and Cardiovascular Health. Circulation 2013, 128, 553–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellen, P.B.; Walsh, T.F.; Herrington, D.M. Whole Grain Intake and Cardiovascular Disease: A Meta-Analysis. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Griffith, J.A.; Chasan-Taber, L.; Olendzki, B.C.; Jackson, E.; Stanek, E.J.; Li, W.; Pagoto, S.L.; Hafner, A.R.; Ockene, I.S. Association between Dietary Fiber and Serum C-Reactive Protein. Am. J. Clin. Nutr. 2006, 83, 760–766. [Google Scholar] [CrossRef]
- Chu, A.; Foster, M.; Samman, S. Zinc Status and Risk of Cardiovascular Diseases and Type 2 Diabetes Mellitus—A Systematic Review of Prospective Cohort Studies. Nutrients 2016, 8, 707. [Google Scholar] [CrossRef]
- Rodriguez-Leyva, D.; Weighell, W.; Edel, A.L.; LaVallee, R.; Dibrov, E.; Pinneker, R.; Maddaford, T.G.; Ramjiawan, B.; Aliani, M.; Guzman, R.; et al. Potent Antihypertensive Action of Dietary Flaxseed in Hypertensive Patients. Hypertension 2013, 62, 1081–1089. [Google Scholar] [CrossRef] [Green Version]
- Grooms, K.N.; Ommerborn, M.J.; Pham, D.Q.; Djoussé, L.; Clark, C.R. Dietary Fiber Intake and Cardiometabolic Risks among US Adults, NHANES 1999-2010. Am. J. Med. 2013, 126, 1059–1067.e4. [Google Scholar] [CrossRef] [Green Version]
- Rodondi, P.Y.; Degoumois, F.; Marques-Vidal, P.; Rodondi, N. Peut-on Abaisser Son Taux de Cholestérol Avec Des Compléments Alimentaires? Rev. Med. Suisse 2016, 12, 451–453. [Google Scholar]
- Cicero, A.F.G.; Fogacci, F.; Veronesi, M.; Strocchi, E.; Grandi, E.; Rizzoli, E.; Poli, A.; Marangoni, F.; Borghi, C. A Randomized Placebo-Controlled Clinical Trial to Evaluate the Medium-Term Effects of Oat Fibers on Human Health: The Beta-Glucan Effects on Lipid Profile, Glycemia and InTestinal Health (BELT) Study. Nutrients 2020, 12, 686. [Google Scholar] [CrossRef] [Green Version]
- Cicero, A.F.G.; Fogacci, F.; Stoian, A.P.; Vrablik, M.; Al Rasadi, K.; Banach, M.; Toth, P.P.; Rizzo, M. Nutraceuticals in the Management of Dyslipidemia: Which, When, and for Whom? Could Nutraceuticals Help Low-Risk Individuals with Non-Optimal Lipid Levels? Curr. Atheroscler. Rep. 2021, 23, 57. [Google Scholar] [CrossRef]
- Hartley, L.; May, M.D.; Rees, K. Dietary Fibre for the Primary Prevention of Cardiovascular Disease. In Cochrane Database of Systematic Reviews; Rees, K., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2015. [Google Scholar]
- Hajishafiee, M.; Saneei, P.; Benisi-Kohansal, S.; Esmaillzadeh, A. Cereal Fibre Intake and Risk of Mortality from All Causes, CVD, Cancer and Inflammatory Diseases: A Systematic Review and Meta-Analysis of Prospective Cohort Studies. Br. J. Nutr. 2016, 116, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Schlienger, J.L. Modifications Thérapeutiques Du Mode de Vie et Prévention Cardiovasculaire Chez Les Sujets Diabétiques de Type 2. Médecine Des Mal. Métaboliques 2019, 13, 27–35. [Google Scholar] [CrossRef]
- Mariño, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J.; et al. Gut Microbial Metabolites Limit the Frequency of Autoimmune T Cells and Protect against Type 1 Diabetes. Nat. Immunol. 2017, 18, 552–562. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Farias, A.; de Melo, A.C.; de Sales Ferreira, J.C. A Importância Da Alimentação Saudável Para Os Portadores de Doença Autoimune. Res. Soc. Dev. 2021, 10, e233101220452. [Google Scholar] [CrossRef]
- Savi, V.; Desmouliere, A.; Lepage, P.; Halioua, B.; Pham, T.; Sokol, H.; Pourcel, G.; Brun, M.; Mary, K.; Pain, E.; et al. Alimentation Et Maladies Auto-Immunes Existe-t-Il Des Nutriments Dont Les Effets Sur La Modulation de l’ Inflammation Ou l’ Immunité Auraient Été Démontrés Sur Plusieurs Maladies Auto-Immunes Différentes? Travail de Bachelor. Nutr. Clin. Métabolisme 2017, 56, 70–71. [Google Scholar]
- Sabaté, J.-M. Syndrome de l’intestin Irritable. Actualités Concernant Les Mécanismes et La Prise En Charge. Douleur Analgésie 2017, 30, 205–216. [Google Scholar] [CrossRef]
- Luciano, L. Est-Il Encore Raisonnable de Donner Des Conseils Diététiques à Un Constipé Chronique? Côlon Rectum 2018, 12, 14–16. [Google Scholar] [CrossRef]
- Sabaté, J. Régimes et Syndrome de l’ Intestin Irritable. POST’U 2015, 213–220. Available online: https://www.fmcgastro.org/wp-content/uploads/file/pdf-2015/213_220_Sabate.pdf (accessed on 25 February 2022).
- Menadi, N.; Meziani, S.; Zairi, M.; Bouazza, S.; Demmouche, A.; Djadel, T.; Benabdeli, K.; Belbraouet, S. Effet Du Son de Blé Dur Dans Le Traitement Du Syndrome de l’intestin Irritable. Phytothérapie 2021, 19, 348–354. [Google Scholar] [CrossRef]
- Fischer, M.H.; Yu, N.; Gray, G.R.; Ralph, J.; Anderson, L.; Marlett, J.A. The Gel-Forming Polysaccharide of Psyllium Husk (Plantago Ovata Forsk). Carbohydr. Res. 2004, 339, 2009–2017. [Google Scholar] [CrossRef]
- Marlett, J.A.; Kajs, T.M.; Fischer, M.H. An Unfermented Gel Component of Psyllium Seed Husk Promotes Laxation as a Lubricant in Humans. Am. J. Clin. Nutr. 2000, 72, 784–789. [Google Scholar] [CrossRef] [Green Version]
- El-Salhy, M.; Ystad, S.O.; Mazzawi, T.; Gundersen, D. Dietary Fiber in Irritable Bowel Syndrome (Review). Int. J. Mol. Med. 2017, 40, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternini, C.; Anselmi, L.; Rozengurt, E. Enteroendocrine Cells: A Site of ‘Taste’ in Gastrointestinal Chemosensing. Curr. Opin. Endocrinol. Diabetes Obes. 2008, 15, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seim, I.; El-Salhy, M.; Hausken, T.; Gundersen, D.; Chopin, L. Ghrelin and the Brain-Gut Axis as a Pharmacological Target for Appetite Control. Curr. Pharm. Des. 2012, 18, 768–775. [Google Scholar] [CrossRef]
- Arora, T.; Loo, R.L.; Anastasovska, J.; Gibson, G.R.; Tuohy, K.M.; Sharma, R.K.; Swann, J.R.; Deaville, E.R.; Sleeth, M.L.; Thomas, E.L.; et al. Differential Effects of Two Fermentable Carbohydrates on Central Appetite Regulation and Body Composition. PLoS ONE 2012, 7, e43263. [Google Scholar] [CrossRef] [PubMed]
- Moayyedi, P.; Quigley, E.M.M.; Lacy, B.E.; Lembo, A.J.; Saito, Y.A.; Schiller, L.R.; Soffer, E.E.; Spiegel, B.M.R.; Ford, A.C. The Effect of Fiber Supplementation on Irritable Bowel Syndrome: A Systematic Review and Meta-Analysis. Am. J. Gastroenterol. 2014, 109, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, M.S. Nutrition and Cancer: A Review of the Evidence for an Anti-Cancer Diet. Nutr. J. 2004, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Bingham, S.A.; Day, N.E.; Luben, R.; Ferrari, P.; Slimani, N.; Norat, T.; Clavel-Chapelon, F.; Kesse, E.; Nieters, A.; Boeing, H.; et al. Dietary Fibre in Food and Protection against Colorectal Cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC): An Observational Study. Lancet 2003, 361, 1496–1501. [Google Scholar] [CrossRef]
- Zeng, H. Mechanisms Linking Dietary Fiber, Gut Microbiota and Colon Cancer Prevention. World J. Gastrointest. Oncol. 2014, 6, 41. [Google Scholar] [CrossRef]
- Veettil, S.K.; Wong, T.Y.; Loo, Y.S.; Playdon, M.C.; Lai, N.M.; Giovannucci, E.L.; Chaiyakunapruk, N. Role of Diet in Colorectal Cancer Incidence. JAMA Netw. Open 2021, 4, e2037341. [Google Scholar] [CrossRef]
- Farvid, M.S.; Spence, N.D.; Holmes, M.D.; Barnett, J.B. Fiber Consumption and Breast Cancer Incidence: A Systematic Review and Meta-analysis of Prospective Studies. Cancer 2020, 126, 3061–3075. [Google Scholar] [CrossRef]
- Lasserre, A.; Gaillot, J.; Deutsch, A.; Chauvet, C.; Bessette, D.; Ancellin, R. Prévention Des Cancers En France: Quel Rôle Pour Les Professionnels de Santé? Bull. Cancer 2017, 104, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Syam, A.F. The Effect of Fiber Diet on Colonic Cancer Formation: The Role of Butyrate. Med. J. Indones. 2003, 12, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Scheppach, W.; Bartram, H.; Richter, F. Role of Short-Chain Fatty Acids in the Prevention of Colorectal Cancer. Eur. J. Cancer 1995, 31, 1077–1080. [Google Scholar] [CrossRef]
- Shang, F.-M.; Liu, H.-L. Fusobacterium Nucleatum and Colorectal Cancer: A Review. World J. Gastrointest. Oncol. 2018, 10, 71–81. [Google Scholar] [CrossRef]
- Han, Y.W. Fusobacterium Nucleatum: A Commensal-Turned Pathogen. Curr. Opin. Microbiol. 2015, 23, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Brennan, C.A.; Garrett, W.S. Fusobacterium Nucleatum—Symbiont, Opportunist and Oncobacterium. Nat. Rev. Microbiol. 2019, 17, 156–166. [Google Scholar] [CrossRef]
- Mehta, R.S.; Nishihara, R.; Cao, Y.; Song, M.; Mima, K.; Qian, Z.R.; Nowak, J.A.; Kosumi, K.; Hamada, T.; Masugi, Y.; et al. Association of Dietary Patterns with Risk of Colorectal Cancer Subtypes Classified by Fusobacterium Nucleatum in Tumor Tissue. JAMA Oncol. 2017, 3, 921. [Google Scholar] [CrossRef] [Green Version]
- Chhim, A.-S.; Fassier, P.; Latino-Martel, P.; Druesne-Pecollo, N.; Zelek, L.; Duverger, L.; Hercberg, S.; Galan, P.; Deschasaux, M.; Touvier, M. O22 Étude prospective des liens entre consommation d’alcool et risque de cancers hormono-dépendants: Modulation par l’apport en fibres alimentaires. Nutr. Clin. Métabolisme 2014, 28, S40–S41. [Google Scholar] [CrossRef]
- McGuire, S. World Cancer Report 2014. Geneva, Switzerland: World Health Organization, International Agency for Research on Cancer, WHO Press, 2015. Adv. Nutr. 2016, 7, 418–419. [Google Scholar] [CrossRef] [Green Version]
- Sapienza, C.; Issa, J.-P. Diet, Nutrition, and Cancer Epigenetics. Annu. Rev. Nutr. 2016, 36, 665–681. [Google Scholar] [CrossRef]
- Dobbins, M.; Decorby, K.; Choi, B.C.K. The Association between Obesity and Cancer Risk: A Meta-Analysis of Observational Studies from 1985 to 2011. ISRN Prev. Med. 2013, 2013, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aune, D.; Giovannucci, E.; Boffetta, P.; Fadnes, L.T.; Keum, N.; Norat, T.; Greenwood, D.C.; Riboli, E.; Vatten, L.J.; Tonstad, S. Fruit and Vegetable Intake and the Risk of Cardiovascular Disease, Total Cancer and All-Cause Mortality—A Systematic Review and Dose-Response Meta-Analysis of Prospective Studies. Int. J. Epidemiol. 2017, 46, 1029–1056. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-C.; Tong, X.; Xu, J.-Y.; Han, S.-F.; Wan, Z.-X.; Qin, J.-B.; Qin, L.-Q. Whole-Grain Intake and Total, Cardiovascular, and Cancer Mortality: A Systematic Review and Meta-Analysis of Prospective Studies. Am. J. Clin. Nutr. 2016, 104, 164–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Je, Y. Dietary Fibre Intake and Mortality from Cardiovascular Disease and All Cancers: A Meta-Analysis of Prospective Cohort Studies. Arch. Cardiovasc. Dis. 2016, 109, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Veronese, N.; Solmi, M.; Caruso, M.G.; Giannelli, G.; Osella, A.R.; Evangelou, E.; Maggi, S.; Fontana, L.; Stubbs, B.; Tzoulaki, I. Dietary Fiber and Health Outcomes: An Umbrella Review of Systematic Reviews and Meta-Analyses. Am. J. Clin. Nutr. 2018, 107, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Wu, K.; Meyerhardt, J.A.; Ogino, S.; Wang, M.; Fuchs, C.S.; Giovannucci, E.L.; Chan, A.T. Fiber Intake and Survival After Colorectal Cancer Diagnosis. JAMA Oncol. 2018, 4, 71. [Google Scholar] [CrossRef]
- Wang, C.-H.; Qiao, C.; Wang, R.-C.; Zhou, W.-P. Dietary Fiber Intake and Pancreatic Cancer Risk: A Meta-Analysis of Epidemiologic Studies. Sci. Rep. 2015, 5, 10834. [Google Scholar] [CrossRef] [Green Version]
- Poutier, A.; Ung, C.; Delhumeau, S.; Hamidi, Y. Le Rôle Du Pharmacien Dans La Prévention de l’obésité. Actual. Pharm. 2017, 56, 25–29. [Google Scholar] [CrossRef]
- Zou, J.; Chassaing, B.; Singh, V.; Pellizzon, M.; Ricci, M.; Fythe, M.D.; Kumar, M.V.; Gewirtz, A.T. Fiber-Mediated Nourishment of Gut Microbiota Protects against Diet-Induced Obesity by Restoring IL-22-Mediated Colonic Health. Cell Host Microbe 2018, 23, 41–53.e4. [Google Scholar] [CrossRef] [Green Version]
- Akrami, M.; Sasani, M.R. Dietary Habits Affect Quality of Life: Bowel Obstruction Caused by Phytobezoar. Iran. J. Public Health 2016, 45, 1080–1082. [Google Scholar]
- Fritsch, J.; Garces, L.; Quintero, M.A.; Pignac-Kobinger, J.; Santander, A.M.; Fernández, I.; Ban, Y.J.; Kwon, D.; Phillips, M.C.; Knight, K.; et al. Low-Fat, High-Fiber Diet Reduces Markers of Inflammation and Dysbiosis and Improves Quality of Life in Patients with Ulcerative Colitis. Clin. Gastroenterol. Hepatol. 2021, 19, 1189–1199.e30. [Google Scholar] [CrossRef] [PubMed]
- Healey, G.R.; Celiberto, L.S.; Lee, S.M.; Jacobson, K. Fiber and Prebiotic Interventions in Pediatric Inflammatory Bowel Disease: What Role Does the Gut Microbiome Play? Nutrients 2020, 12, 3204. [Google Scholar] [CrossRef] [PubMed]
- de Castro, M.M.; Pascoal, L.B.; Steigleder, K.M.; Siqueira, B.P.; Corona, L.P.; de Lourdes Setsuko, M.; Milanski, M.; Leal, R.F. Role of Diet and Nutrition in Inflammatory Bowel Disease. World J. Exp. Med. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Pop, A.L.; Henteș, P.; Pali, M.-A.; Oșanu, L.; Ciobanu, A.; Nasui, B.A.; Mititelu, M.; Crișan, S.; Peneș, O.N. Study regarding a new extended-release calcium ascorbate and hesperidin solid oral formulation. Farmacia 2022, 70, 151–157. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
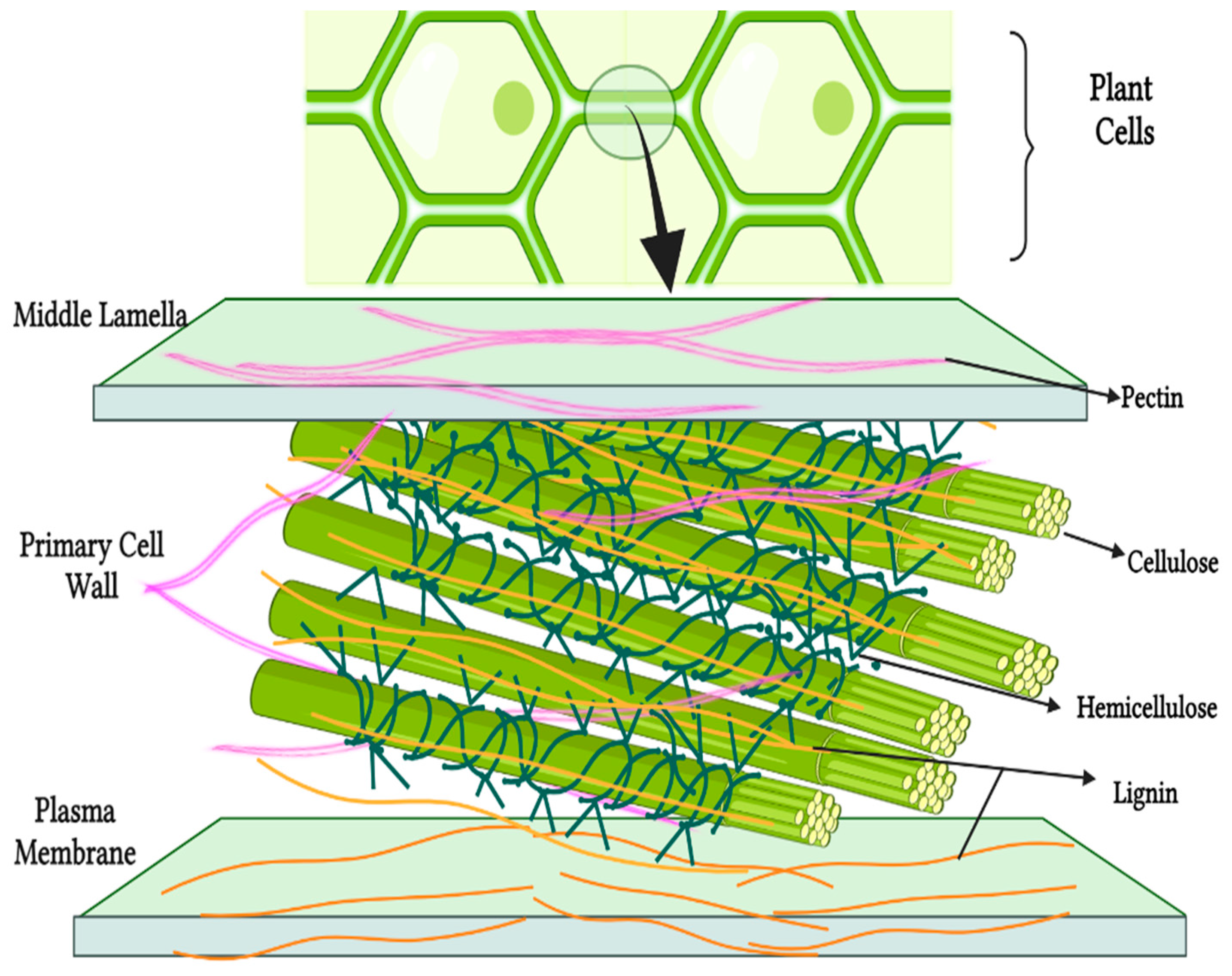
| Poly-/Oligo-Saccharides Class | Sources | Main Units | References |
|---|---|---|---|
| Non-Starch Polysaccharides, MU ≥ 10 | |||
| Cellulose * | Cereals, pulses—outer layers, root and leafy vegetables, legumes, pears and apples | Glucose monomers | [17] |
| Hemicellulose ** | Cereal bran and whole grains (starchy endosperm and aleurone layer), vegetable and fruit cell walls | D-xylose, D-mannose, D-galactose and L-arabinose | [14] |
| Mannans and heteromannans | Date, green coffee bean seeds and aloe vera Grain legumes (endosperm) Iris seeds and lily bulbs Norway spruce wood pulp | Mannans *, galactomannans #, glucomannans # and galactoglucomannans | [16] |
| Pectins # | Apple and citrus peel (and other fruits), cabbage, whole grains, beetroot and grain legumes | Arabinose, rhamnose, galactose sugars and galacturonic acids | [14] |
| Gums | Xanthan gum * Alginates ** Agar-agar ** Carrageenan ** | Pentose and hexoze monomers | [18] |
| Mucilages # | Aloe vera, Cactus, Okra, Hibiscus | Main glycoproteins | [16] |
| Inulin # and fructans | Jerusalem artichoke, Chicory root, onion and cereal grains | fructofuranosyl residues | [15] |
| Non-Digestible (resistant) oligosaccharides, MU = 3–9 | |||
| α-galactosides ** | Chickpea, bean, lentil, etc. | Raffinose, stachyose, verbascose | [15] |
| β-fructo-oligosaccharides ** | Polymers resulted from polysaccharides hydrolysis (inulin and lactose hydrolysis produce FOS and, respectively, GOS). | β-Fructo- (FOS), α-galacto- (GOS), β-galacto- (TOS), xylo- (XOS), arabino-xylo- (AXOS) oligosaccharides | [15] |
| Resistant dextrins ** | cereal-based vegetable milk, baked goods, dairy products and granola bars | Poly-D-glucose | [15] |
| Polydextrose ** | Cakes, candies, mixes and frozen desserts and beverages | Poly-D-glucose | [15] |
| Resistant Starches * (RS), MU ≥ 10 | |||
| RS type 1 | Grains and legumes (whole or partially milled) | physically inaccessible starch | [15] |
| RS type 2 | High-amylose starches, green bananas | granular starches | [15] |
| RS type 3 | Cooled starches in cooked starchy foods and enzyme-debranched starches | gelatinized and retrograded starches | [19] |
| RS type 4 | chemically modified (mainly cross-linked starches) | [15] | |
| Associated Substances Non-carbohydrates | |||
| Lignin * | Fruits, particularly strawberries and peaches | Coumaryl, coniferyl and sinapyl alcohols (aromatic alcohols) | [17] |
| Waxes * | Wax is present in rice bran, seed and seed hulls of sunflower | Long alkyl chains | [20] |
| Chitins * | Fungus’ cell walls, lobster, crab and shrimp exoskeletons and insects | N-acetylglucosamine | [15] |
| Phytates/Phytic acid # | Plant seeds, mainly in legumes, peanuts, cereals and oilseeds and generally found in almost all plant-based foods | - | [21] |
| Study Type | Nature of Participants | Duration Administered | Type and Dose of Fiber Administered | Main Findings | References |
|---|---|---|---|---|---|
| Randomized, double-blind parallel-group design controlled trial | Hypercholesterolemic adults | 6 weeks | 6 g concentrated β-glucan/day | Reduced total and LDL cholesterol | [93] |
| Randomized, single-blind, controlled, crossover intervention trial | Impaired glucose tolerance participants | 18 weeks | 15 g arabinoxylan/day | Improved fasting serum glucose | [94] |
| Randomized, single-blind, controlled, crossover intervention trial | Healthy and glucose intolerant subjects | one year | 6 g of fiber partially hydrolyzed guar gum with each meal | Reduced postprandial plasma glucose, postprandial insulin, triacylglycerol levels | [95] |
| Randomized, single-blind, controlled, crossover intervention trial | Pre and post-menopausal, hypercholesterolemic women | 6 weeks | 5 g psyllium/day | Reduced total cholesterol for post-menopausal women but not in pre-menopausal women | [96] |
| Randomized, crossover, a single-blind, dietary intervention | Free-living subjects | 28 days | Control diet: 25 g dietary fiber/day; LKFibre * diet: 55 g dietary fiber/day | Reduced total and LDL cholesterol | [97] |
| Clinical trial study | Healthy subjects; type-2-diabetes and pre-diabetics subjects. | 16 weeks | 10 g/day gum Arabic | Decrease in glycosylated hemoglobin (HbAc1), decrease fasting blood glucose, health improvement | [98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ioniță-Mîndrican, C.-B.; Ziani, K.; Mititelu, M.; Oprea, E.; Neacșu, S.M.; Moroșan, E.; Dumitrescu, D.-E.; Roșca, A.C.; Drăgănescu, D.; Negrei, C. Therapeutic Benefits and Dietary Restrictions of Fiber Intake: A State of the Art Review. Nutrients 2022, 14, 2641. https://doi.org/10.3390/nu14132641
Ioniță-Mîndrican C-B, Ziani K, Mititelu M, Oprea E, Neacșu SM, Moroșan E, Dumitrescu D-E, Roșca AC, Drăgănescu D, Negrei C. Therapeutic Benefits and Dietary Restrictions of Fiber Intake: A State of the Art Review. Nutrients. 2022; 14(13):2641. https://doi.org/10.3390/nu14132641
Chicago/Turabian StyleIoniță-Mîndrican, Corina-Bianca, Khaled Ziani, Magdalena Mititelu, Eliza Oprea, Sorinel Marius Neacșu, Elena Moroșan, Denisa-Elena Dumitrescu, Adrian Cosmin Roșca, Doina Drăgănescu, and Carolina Negrei. 2022. "Therapeutic Benefits and Dietary Restrictions of Fiber Intake: A State of the Art Review" Nutrients 14, no. 13: 2641. https://doi.org/10.3390/nu14132641
APA StyleIoniță-Mîndrican, C.-B., Ziani, K., Mititelu, M., Oprea, E., Neacșu, S. M., Moroșan, E., Dumitrescu, D.-E., Roșca, A. C., Drăgănescu, D., & Negrei, C. (2022). Therapeutic Benefits and Dietary Restrictions of Fiber Intake: A State of the Art Review. Nutrients, 14(13), 2641. https://doi.org/10.3390/nu14132641