
Plasma Levels of Endocannabinoids and Their Analogues Are Related to Specific Fecal Bacterial Genera in Young Adults: Role in Gut Barrier Integrity

 , ,
, ,  ,
,  ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Design and Participants
2.2. Anthropometry and Body Composition Assessment
2.3. Determination of Plasma Levels of Endocannabinoids and Their Analogues
2.3.1. Sample Preparation
2.3.2. Liquid Chromatography-Tandem Mass Spectrometry Analysis
2.3.3. Data Pre-Processing
2.4. Fecal Microbiota Analyses
2.4.1. Stool Collection and DNA Extraction
2.4.2. Sequencing Analysis
2.4.3. Bioinformatics Analysis
2.5. Determination of Plasma Levels of Lipopolysaccharides
2.6. Statistical Analysis
3. Results
3.1. Characteristics of the Participants
3.2. The Plasma Levels of Endocannabinoids and Their Analogues Are Not Related to Fecal Microbiota Beta and Alpha Diversities
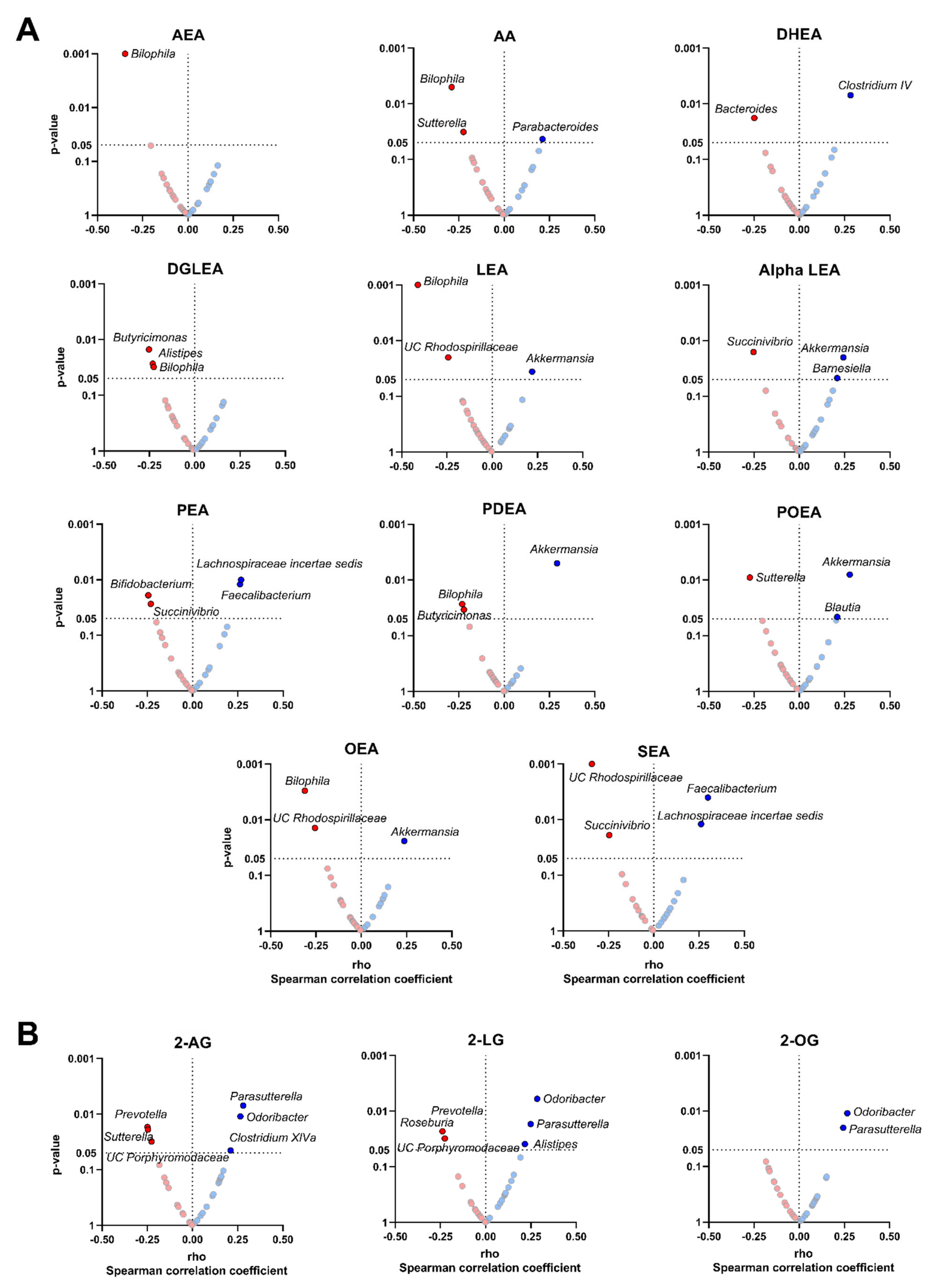
3.3. The Plasma Levels of Endocannabinoids and Their Analogues Levels Are Related to the Relative Abundance of Specific Fecal Microbiota Genera
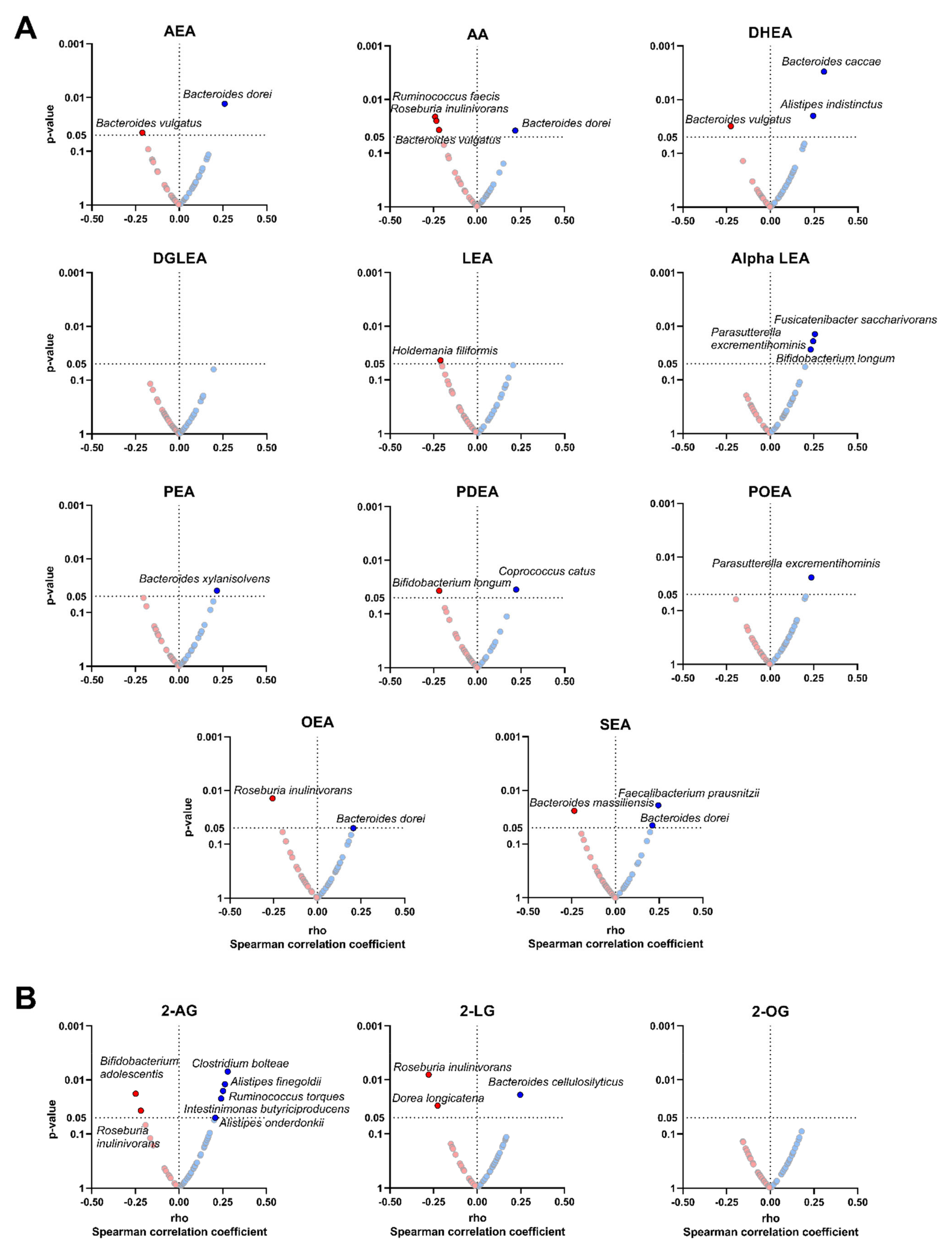
3.4. The Plasma Levels of Endocannabinoids and Their Analogues Are Related to the Relative Abundance of Specific Bacterial Species
3.5. The Plasma Levels of Endocannabinoids and Their Analogues Are Negatively Correlated to Plasma Levels of Lipopolysaccharides but Only in Those Participants with High Plasma Levels of Lipopolysaccharides
4. Discussion
4.1. Role of Endocannabinoids and Their Analogues in Gut Microbiota Diversity
4.2. Role of Endocannabinoids and Their Analogues in the Gut Barrier Integrity
4.3. Limitations and Strengths
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 2-AG | 2-arachidonoylglycerol; |
| 2-LG | 2-linoleoylglycerol; |
| 2-OG | 2-oleoylglycerol; |
| AA | arachidonic acid; |
| AEA | anandamide; |
| alpha LEA | alpha-Linolenoyl ethanolamide; |
| DGLEA | dihomo-gamma-linolenoyl ethanolamide; |
| DHEA | docosahexaenoyl ethanolamide; |
| LEA | linoleoyl ethanolamide; |
| NAEs | N-acylethanolamines; |
| OEA | oleoyl ethanolamine; |
| PDEA | pentadecanoyl ethanolamide; |
| PEA | palmitoyl ethanolamide; |
| POEA | palmitoleoyl ethanolamide; |
| SEA | stearoyl ethanolamide; |
| UC | unclassified. |
References
- Muccioli, G.G. Endocannabinoid Biosynthesis and Inactivation, from Simple to Complex. Drug Discov. Today 2010, 15, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Devane, W.A.; Hanuš, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and Structure of a Brain Constituent That Binds to the Cannabinoid Receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Cristino, L.; Becker, T.; Di Marzo, V. Endocannabinoids and Energy Homeostasis: An Update. BioFactors 2014, 40, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Chiurchiù, V.; Leuti, A.; Maccarrone, M. Bioactive Lipids and Chronic Inflammation: Managing the Fire Within. Front. Immunol. 2018, 9, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meletis, C. The Important Role of the Endocannabinoid System and the Endocannabinoidome in Gut Health. Altern. Ther. Health Med. 2019, 25, 24–28. [Google Scholar]
- Rosenbaum, M.; Knight, R.; Leibel, R.L. The Gut Microbiota in Human Energy Homeostasis and Obesity. Trends Endocrinol. Metab. 2015, 26, 493–501. [Google Scholar] [CrossRef] [Green Version]
- Everard, A.; Cani, P.D. Diabetes, Obesity and Gut Microbiota. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Iebba, V.; Totino, V.; Gagliardi, A.; Santangelo, F.; Cacciotti, F.; Trancassini, M.; Mancini, C.; Cicerone, C.; Corazziari, E.; Pantanella, F.; et al. Eubiosis and Dysbiosis: The Two Sides of the Microbiota. New Microbiol. 2016, 39, 1–12. [Google Scholar]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive Impact of Non-Antibiotic Drugs on Human Gut Bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef]
- Forslund, S.K.; Chakaroun, R.; Zimmermann-Kogadeeva, M.; Markó, L.; Aron-Wisnewsky, J.; Nielsen, T.; Moitinho-Silva, L.B.; Schmidt, T.S.; Falony, G.; Vieira-Silva, S.; et al. Combinatorial, Additive and Dose-Dependent Drug–Microbiome Associations. Nataliya Sokolovska 2021, 600, 500. [Google Scholar] [CrossRef] [PubMed]
- Gérard, P. Gut Microbiota and Obesity. Cell. Mol. Life Sci. 2016, 73, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in Gut Microbiota Control Metabolic Endotoxemia-Induced Inflammation in High-Fat Diet–Induced Obesity and Diabetes in Mice. Diabetes 2008, 57, 1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in Gut Microbiota Control Inflammation in Obese Mice through a Mechanism Involving GLP-2-Driven Improvement of Gut Permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; Everard, A.; Duparc, T. Gut Microbiota, Enteroendocrine Functions and Metabolism. Curr. Opin. Pharmacol. 2013, 13, 935–940. [Google Scholar] [CrossRef]
- Cani, P.D.; Plovier, H.; Van Hul, M.; Geurts, L.; Delzenne, N.M.; Druart, C.; Everard, A. Endocannabinoids—At the Crossroads between the Gut Microbiota and Host Metabolism. Nat. Rev. Endocrinol. 2016, 12, 133–143. [Google Scholar] [CrossRef]
- Geurts, L.; Everard, A.; Van Hul, M.; Essaghir, A.; Duparc, T.; Matamoros, S.; Plovier, H.; Castel, J.; Denis, R.G.P.; Bergiers, M.; et al. Adipose Tissue NAPE-PLD Controls Fat Mass Development by Altering the Browning Process and Gut Microbiota. Nat. Commun. 2015, 6, 6495. [Google Scholar] [CrossRef] [Green Version]
- Tagliamonte, S.; Laiola, M.; Ferracane, R.; Vitale, M.; Gallo, M.A.; Meslier, V.; Pons, N.; Ercolini, D.; Vitaglione, P. Mediterranean Diet Consumption Affects the Endocannabinoid System in Overweight and Obese Subjects: Possible Links with Gut Microbiome, Insulin Resistance and Inflammation. Eur. J. Nutr. 2021, 60, 3703–3716. [Google Scholar] [CrossRef]
- Oláh, A.; Szekanecz, Z.; Bíró, T. Targeting Cannabinoid Signaling in the Immune System: “High”-Ly Exciting Questions, Possibilities, and Challenges. Front. Immunol. 2017, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Muccioli, G.G.; Naslain, D.; Bäckhed, F.; Reigstad, C.S.; Lambert, D.M.; Delzenne, N.M.; Cani, P.D. The Endocannabinoid System Links Gut Microbiota to Adipogenesis. Mol. Syst. Biol. 2010, 6, 392. [Google Scholar] [CrossRef]
- Sanchez-Delgado, G.; Martinez-Tellez, B.; Olza, J.; Aguilera, C.M.; Labayen, I.; Ortega, F.B.; Chillon, P.; Fernandez-Reguera, C.; Alcantara, J.M.A.; Martinez-Avila, W.D.; et al. Activating Brown Adipose Tissue through Exercise (ACTIBATE) in Young Adults: Rationale, Design and Methodology. Contemp. Clin. Trials 2015, 45, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shabat, S.; Fride, E.; Sheskin, T.; Tamiri, T.; Rhee, M.-H.; Vogel, Z.; Bisogno, T.; De Petrocellis, L.; Di Marzo, V.; Mechoulam, R. An Entourage Effect: Inactive Endogenous Fatty Acid Glycerol Esters Enhance 2-Arachidonoyl-Glycerol Cannabinoid Activity. Eur. J. Pharmacol. 1998, 353, 23–31. [Google Scholar] [CrossRef]
- Lambert, D.M.; Di Marzo, V. The Palmitoylethanolamide and Oleamide Enigmas: Are These Two Fatty Acid Amides Cannabimimetic? Curr. Med. Chem. 1999, 6, 757–773. [Google Scholar] [CrossRef] [PubMed]
- FDA. Bioanalytical Method Validation Guidance; FDA: Silver Spring, MD, USA, 2018; Volume 1043, p. 25. [Google Scholar]
- Van der Kloet, F.M.; Bobeldijk, I.; Verheij, E.R.; Jellema, R.H. Analytical Error Reduction Using Single Point Calibration for Accurate and Precise Metabolomic Phenotyping. J. Proteome Res. 2009, 8, 5132–5141. [Google Scholar] [CrossRef]
- Martin, J.B. The Equilibrium between Symmetrical and Unsymmetrical Monoglycerides and Determination of Total Monoglycerides. J. Am. Chem. Soc. 1953, 75, 5483–5486. [Google Scholar] [CrossRef]
- Pr Herlemann, D.; Labrenz, M.; Jü Rgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in Bacterial Communities along the 2000 Km Salinity Gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 1 March 2021).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of RRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and Tools for High Throughput RRNA Analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [Green Version]
- Schulz, C.; Schütte, K.; Koch, N.; Vilchez-Vargas, R.; Wos-Oxley, M.L.; Oxley, A.P.A.; Vital, M.; Malfertheiner, P.; Pieper, D.H. The Active Bacterial Assemblages of the Upper GI Tract in Individuals with and without Helicobacter Infection. Gut 2018, 67, 216–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.A.; Knight, R. Species Divergence and the Measurement of Microbial Diversity. FEMS Microbiol. Rev. 2008, 32, 557–578. [Google Scholar] [CrossRef] [PubMed]
- Finotello, F.; Mastrorilli, E.; Di Camillo, B. Measuring the Diversity of the Human Microbiota with Targeted Next-Generation Sequencing. Brief. Bioinform. 2018, 19, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Leo, L.; Shetty, S. Microbiome R Package. 2019. Available online: http://microbiome.github.io (accessed on 1 March 2021).
- Gotelli, N.J.; Colwell, R.K. Estimating Species Richness. Biol. Divers. Front. Meas. Assess. 2011, 12, 39–54. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell: Oxford, UK, 2004; ISBN 0-632-05633-9. [Google Scholar]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Camargo, J.A. New Diversity Index for Assessing Structural Alterations in Aquatic Communities. Bull. Environ. Contam. Toxicol. 1992, 48, 428–434. [Google Scholar] [CrossRef]
- Clemente-Postigo, M.; Oliva-Olivera, W.; Coin-Aragüez, L.; Ramos-Molina, B.; Giraldez-Perez, R.M.; Lhamyani, S.; Alcaide-Torres, J.; Perez-Martinez, P.; El Bekay, R.; Cardona, F.; et al. Metabolic Endotoxemia Promotes Adipose Dysfunction and Inflammation in Human Obesity. Am. J. Physiol. Metab. 2019, 316, E319–E332. [Google Scholar] [CrossRef]
- Castonguay-Paradis, S.; Lacroix, S.; Rochefort, G.; Parent, L.; Perron, J.; Martin, C.; Lamarche, B.; Raymond, F.; Flamand, N.; Di Marzo, V.; et al. Dietary Fatty Acid Intake and Gut Microbiota Determine Circulating Endocannabinoidome Signaling beyond the Effect of Body Fat. Sci. Rep. 2020, 10, 15975. [Google Scholar] [CrossRef]
- Fanelli, F.; Mezzullo, M.; Repaci, A.; Belluomo, I.; Ibarra Gasparini, D.; Di Dalmazi, G.; Mastroroberto, M.; Vicennati, V.; Gambineri, A.; Morselli-Labate, A.M.; et al. Profiling Plasma N-Acylethanolamine Levels and Their Ratios as a Biomarker of Obesity and Dysmetabolism. Mol. Metab. 2018, 14, 82–94. [Google Scholar] [CrossRef]
- Revelle, W.R. Psych: Procedures for Personality and Psychological Research. R Package Version 1.9.12. 2017. Available online: http://personality-project.org/r (accessed on 1 March 2021).
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. R Package “Corrplot”: Visualization of a Correlation Matrix. 2017. Available online: https://github.com/taiyun/corrplot (accessed on 1 March 2021).
- Massier, L.; Blüher, M.; Kovacs, P.; Chakaroun, R.M. Impaired Intestinal Barrier and Tissue Bacteria: Pathomechanisms for Metabolic Diseases. Front. Endocrinol. 2021, 12, 1. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human Gut Microbiome Viewed across Age and Geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia Muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef] [PubMed]
- Payahoo, L.; Khajebishak, Y.; Alivand, M.R.; Soleimanzade, H.; Alipour, S.; Barzegari, A.; Ostadrahimi, A. Investigation the Effect of Oleoylethanolamide Supplementation on the Abundance of Akkermansia Muciniphila Bacterium and the Dietary Intakes in People with Obesity: A Randomized Clinical Trial. Appetite 2019, 141, 104301. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Halder, C.V.; Faria, A.V.d.S.; Andrade, S.S. Action and Function of Faecalibacterium Prausnitzii in Health and Disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Burger-van Paassen, N.; Vincent, A.; Puiman, P.J.; van der Sluis, M.; Bouma, J.; Boehm, G.; van Goudoever, J.B.; van Seuningen, I.; Renes, I.B. The Regulation of Intestinal Mucin MUC2 Expression by Short-Chain Fatty Acids: Implications for Epithelial Protection. Biochem. J. 2009, 420, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain Fatty Acids Stimulate Glucagon-Like Peptide-1 Secretion via the G-Protein–Coupled Receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, S.; Pechereau, F.; Leblanc, N.; Boubertakh, B.; Houde, A.; Martin, C.; Flamand, N.; Silvestri, C.; Raymond, F.; Di Marzo, V.; et al. Rapid and Concomitant Gut Microbiota and Endocannabinoidome Response to Diet-Induced Obesity in Mice. mSystems 2019, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ju, T.; Kong, J.Y.; Stothard, P.; Willing, B.P. Defining the Role of Parasutterella, a Previously Uncharacterized Member of the Core Gut Microbiota. ISME J. 2019, 13, 1520–1534. [Google Scholar] [CrossRef]
- Lee, J.S.; Wang, R.X.; Alexeev, E.E.; Lanis, J.M.; Battista, K.D.; Glover, L.E.; Colgan, S.P. Hypoxanthine Is a Checkpoint Stress Metabolite in Colonic Epithelial Energy Modulation and Barrier Function. J. Biol. Chem. 2018, 293, 6039–6051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etxeberria, U.; Hijona, E.; Aguirre, L.; Milagro, F.I.; Bujanda, L.; Rimando, A.M.; Martínez, J.A.; Portillo, M.P. Pterostilbene-Induced Changes in Gut Microbiota Composition in Relation to Obesity. Mol. Nutr. Food Res. 2017, 61, 1500906. [Google Scholar] [CrossRef] [PubMed]
- Vijay, A.; Kouraki, A.; Gohir, S.; Turnbull, J.; Kelly, A.; Chapman, V.; Barrett, D.A.; Bulsiewicz, W.J.; Valdes, A.M. The Anti-Inflammatory Effect of Bacterial Short Chain Fatty Acids Is Partially Mediated by Endocannabinoids. Gut Microbes 2021, 13, 1997559. [Google Scholar] [CrossRef] [PubMed]
- Lassenius, M.I.; Pietiläinen, K.H.; Kaartinen, K.; Pussinen, P.J.; Syrjänen, J.; Forsblom, C.; Pörsti, I.; Rissanen, A.; Kaprio, J.; Mustonen, J.; et al. Bacterial Endotoxin Activity in Human Serum Is Associated with Dyslipidemia, Insulin Resistance, Obesity, and Chronic Inflammation. Diabetes Care 2011, 34, 1809–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bariani, M.V.; Domínguez Rubio, A.P.; Cella, M.; Burdet, J.; Franchi, A.M.; Aisemberg, J. Role of the Endocannabinoid System in the Mechanisms Involved in the LPS-Induced Preterm Labor. Reproduction 2015, 150, 463–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| N | Mean | ± | SD | |
|---|---|---|---|---|
| Sex (women, %) | 92 | 71% | ||
| Age (years) | 92 | 22 | ± | 2 |
| Anthropometry and body composition assessment | ||||
| Body mass index (kg/m2) | 90 | 24.87 | ± | 4.72 |
| Lean mass (kg) | 83 | 41.13 | ± | 8.98 |
| Fat mass (kg) | 83 | 24.91 | ± | 9.09 |
| Lean mass index (kg/m2) | 83 | 14.43 | ± | 2.24 |
| Fat mass index (kg/m2) | 83 | 8.84 | ± | 3.14 |
| Fat mass percentage (%) | 83 | 35.94 | ± | 7.88 |
| Visceral adipose tissue (g) | 83 | 321.05 | ± | 177.19 |
| Waist circumference (cm) | 90 | 80.60 | ± | 13.97 |
| Plasma levels of endocannabinoids and their analogues (peak area ratio) | ||||
| AEA and related N-acylethanolamines | ||||
| AEA | 88 | 0.14 | ± | 0.06 |
| AA | 88 | 65.18 | ± | 21.75 |
| DHEA * | 88 | 0.08 | ± | 0.16 |
| DGLEA * | 87 | 0.02 | ± | 0.01 |
| LEA | 88 | 0.22 | ± | 0.09 |
| alpha LEA | 88 | 0.008 | ± | 0.004 |
| PEA | 87 | 1.78 | ± | 0.28 |
| PDEA * | 88 | 0.03 | ± | 0.01 |
| POEA | 88 | 0.28 | ± | 0.22 |
| OEA | 88 | 0.70 | ± | 0.21 |
| SEA | 88 | 1.33 | ± | 0.23 |
| 2-AG and related acylglycerols | ||||
| 2-AG * | 88 | 0.02 | ± | 0.09 |
| 2-LG * | 88 | 0.22 | ± | 1.41 |
| 2-OG * | 88 | 0.01 | ± | 0.06 |
| Fecal microbiota parameters | ||||
| Alpha diversity indexes | ||||
| Richness Chao | 92 | 430.68 | ± | 150.99 |
| Shannon diversity | 92 | 4.21 | ± | 0.37 |
| Inverse Simpson diversity | 92 | 34.20 | ± | 14.16 |
| Evenness Camargo | 92 | 0.24 | ± | 0.05 |
| Composition (phylum) | ||||
| Actinobacteria (%) | 92 | 1.59 | ± | 1.55 |
| Bacteroidetes (%) | 92 | 39.80 | ± | 9.06 |
| Firmicutes (%) | 92 | 48.65 | ± | 10.10 |
| Proteobacteria (%) | 92 | 6.81 | ± | 5.29 |
| Verrucomicrobia (%) | 92 | 2.09 | ± | 4.12 |
| Plasma levels of lipopolysaccharide (EU/mL) | 85 | 0.98 | ± | 1.10 |
| Tertiles of the Plasma Levels of Endocannabinoids and Their Analogues (Peak Area Ratio) | Phylum | Genus | ||
|---|---|---|---|---|
| Pseudo-F | p-Value | Pseudo-F | p-Value | |
| AEA and related N-acylethanolamines | ||||
| AEA | −6.626 | 0.383 | −6.789 | 0.336 |
| AA | −5.918 | 0.348 | −6.275 | 0.397 |
| DHEA | −6.581 | 0.548 | −6.620 | 0.684 |
| DGLEA | −6.339 | 0.657 | −6.146 | 0.579 |
| LEA | −6.469 | 0.405 | −6.670 | 0.418 |
| alpha LEA | −6.211 | 0.361 | −6.854 | 0.746 |
| PEA | −6.032 | 0.591 | −5.996 | 0.723 |
| PDEA | −5.818 | 0.312 | −6.370 | 0.485 |
| POEA | −6.495 | 0.423 | −6.516 | 0.336 |
| OEA | −6.410 | 0.372 | −6.645 | 0.369 |
| SEA | −6.460 | 0.373 | −6.797 | 0.442 |
| 2-AG and related acylglycerols | ||||
| 2-AG | −6.281 | 0.640 | −5.808 | 0.473 |
| 2-LG | −6.449 | 0.570 | −6.326 | 0.595 |
| 2-OG | −7.041 | 0.578 | −6.927 | 0.629 |
| AEA and Related N-acylethanolamines (Peak Area Ratio) | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AEA | AA | DHEA | DGLEA | LEA | Alpha LEA | PEA | PDEA | POEA | OEA | SEA | ||||||||||||
| rho | p | rho | p | rho | p | rho | p | rho | p | rho | p | rho | p | rho | p | rho | p | rho | p | rho | p | |
| Q1: Low LPS (N = 19) | 0.262 | 0.012 | 0.228 | 0.030 | 0.123 | 0.246 | 0.221 | 0.035 | 0.404 | <0.001 | 0.428 | <0.001 | 0.451 | <0.001 | 0.234 | 0.026 | 0.467 | <0.001 | 0.440 | <0.001 | 0.428 | <0.001 |
| Q4: High LPS (N = 21) | −0.464 | <0.001 | −0.389 | <0.001 | −0.001 | 0.992 | −0.472 | <0.001 | −0.300 | 0.004 | 0.107 | 0.315 | −0.304 | 0.003 | −0.281 | 0.007 | −0.504 | <0.001 | −0.408 | <0.001 | −0.156 | 0.139 |
| 2-AG and Related Acylglycerols (Peak Area Ratio) | ||||||
|---|---|---|---|---|---|---|
| 2-AG | 2-LG | 2-OG | ||||
| rho | p | rho | p | rho | p | |
| Q1: Low LPS (N = 19) | 0.374 | <0.001 | 0.111 | 0.295 | 0.171 | 0.106 |
| Q4: High LPS (N = 21) | −0.243 | 0.020 | 0.312 | 0.003 | 0.053 | 0.615 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz-Alvarez, L.; Xu, H.; Di, X.; Kohler, I.; Osuna-Prieto, F.J.; Acosta, F.M.; Vilchez-Vargas, R.; Link, A.; Plaza-Díaz, J.; van der Stelt, M.; et al. Plasma Levels of Endocannabinoids and Their Analogues Are Related to Specific Fecal Bacterial Genera in Young Adults: Role in Gut Barrier Integrity. Nutrients 2022, 14, 2143. https://doi.org/10.3390/nu14102143
Ortiz-Alvarez L, Xu H, Di X, Kohler I, Osuna-Prieto FJ, Acosta FM, Vilchez-Vargas R, Link A, Plaza-Díaz J, van der Stelt M, et al. Plasma Levels of Endocannabinoids and Their Analogues Are Related to Specific Fecal Bacterial Genera in Young Adults: Role in Gut Barrier Integrity. Nutrients. 2022; 14(10):2143. https://doi.org/10.3390/nu14102143
Chicago/Turabian StyleOrtiz-Alvarez, Lourdes, Huiwen Xu, Xinyu Di, Isabelle Kohler, Francisco J. Osuna-Prieto, Francisco M. Acosta, Ramiro Vilchez-Vargas, Alexander Link, Julio Plaza-Díaz, Mario van der Stelt, and et al. 2022. "Plasma Levels of Endocannabinoids and Their Analogues Are Related to Specific Fecal Bacterial Genera in Young Adults: Role in Gut Barrier Integrity" Nutrients 14, no. 10: 2143. https://doi.org/10.3390/nu14102143
APA StyleOrtiz-Alvarez, L., Xu, H., Di, X., Kohler, I., Osuna-Prieto, F. J., Acosta, F. M., Vilchez-Vargas, R., Link, A., Plaza-Díaz, J., van der Stelt, M., Hankemeier, T., Clemente-Postigo, M., Tinahones, F. J., Gil, A., Rensen, P. C. N., Ruiz, J. R., & Martinez-Tellez, B. (2022). Plasma Levels of Endocannabinoids and Their Analogues Are Related to Specific Fecal Bacterial Genera in Young Adults: Role in Gut Barrier Integrity. Nutrients, 14(10), 2143. https://doi.org/10.3390/nu14102143