
Effects of Taro (Colocasia esculenta) Water-Soluble Non-Starch Polysaccharide, Lactobacillus acidophilus, Bifidobacterium breve, Bifidobacterium infantis, and Their Synbiotic Mixtures on Pro-Inflammatory Cytokine Interleukin-8 Production



Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Extraction of Taro Water-Soluble Non-Starch Polysaccharides (Tc-WS-NSP)
2.2.2. Digestion of Tc-WS-NSP
2.2.3. Culture Media and Bacterial Culture Preparations
2.2.4. Cell Culture Preparations
2.2.5. Cytotoxicity Evaluation Using 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Assay
2.2.6. Incubation of Undigested or Digested Tc-WS-NSPs, the Probiotics, and Their Synbiotic Mixtures on IL-8 Production by TNF-α Stimulated HT-29 Cells in the Presence of NEC-Positive Associated Pathogenic Bacterium K. oxytoca and Non-Pathogenic E. coli
2.2.7. IL-8 Quantification in Cell Culture Supernatants Using Enzyme-Linked Immunosorbent Assay (ELISA)
2.2.8. Statistical Analysis
3. Results
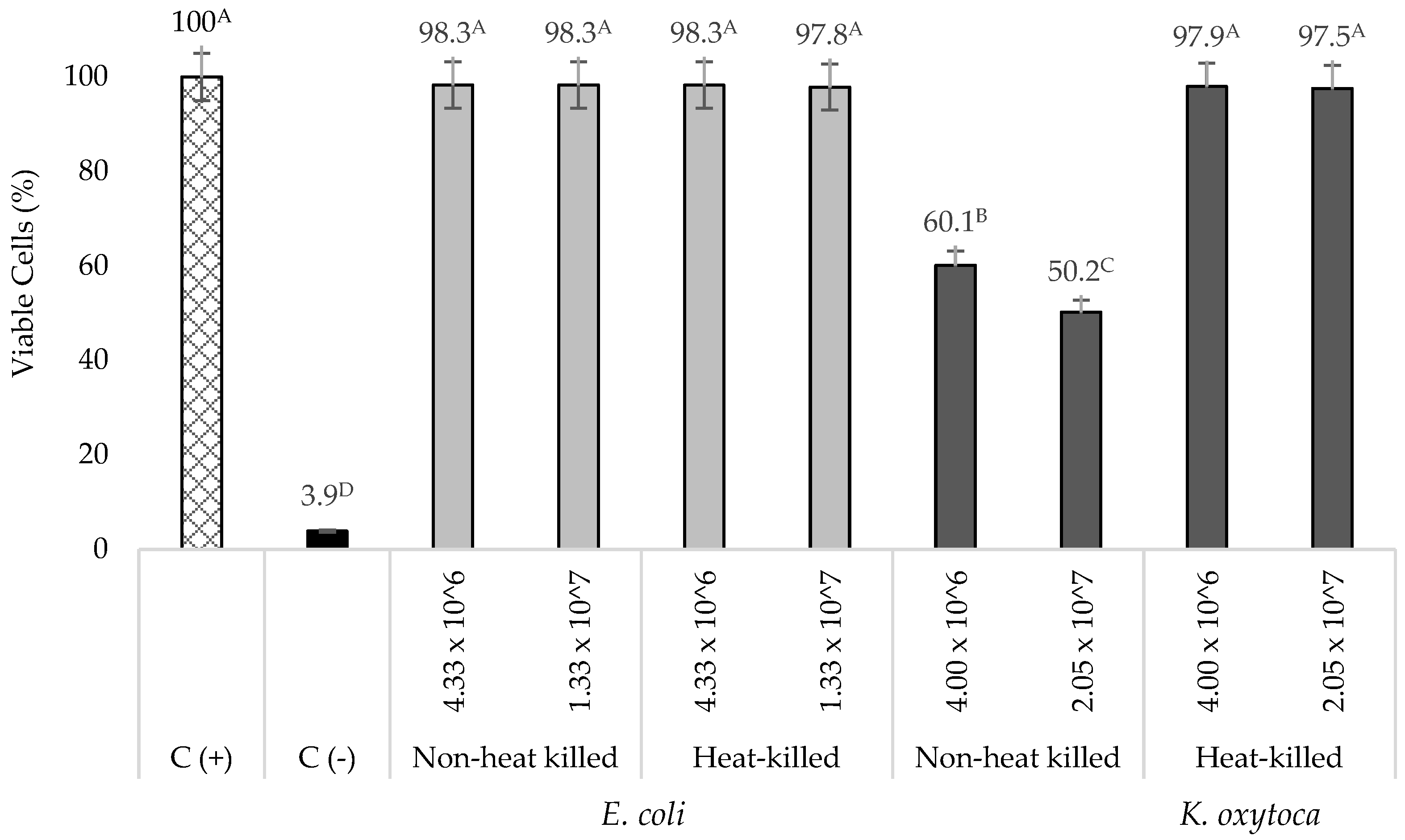
3.1. Cytotoxicity of Live K. oxytoca and Non-Pathogenic E. coli on TNF-α Stimulated HT-29 Cells
3.2. Cytotoxicity of Undigested or Digested Tc-WS-NSPs, Live Probiotics, and Their Synbiotic Mixtures with Heat-Killed K. oxytoca or E. coli on TNF-α Stimulated HT-29 Cells
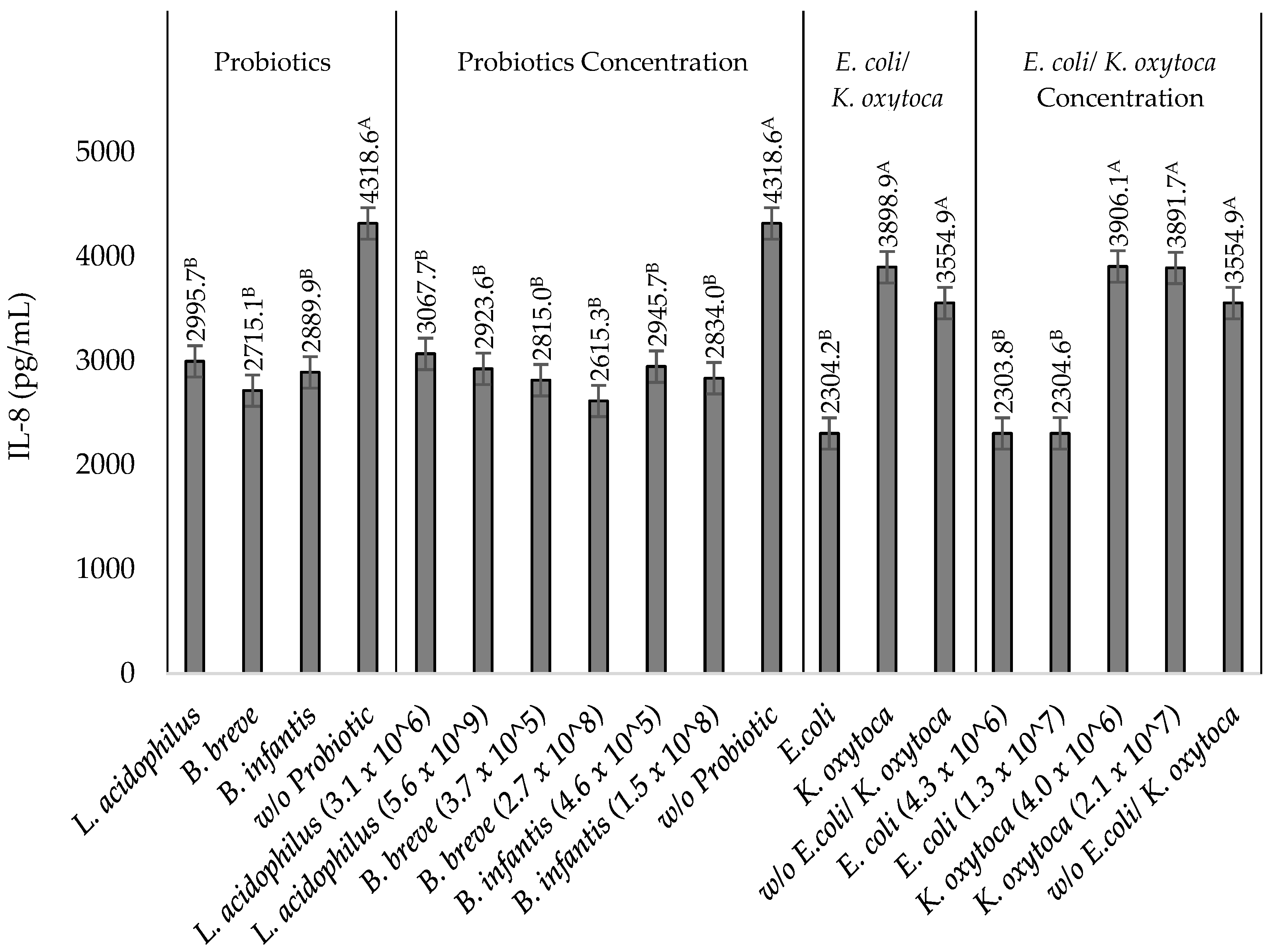
3.3. IL-8 Production by TNF-α Stimulated HT-29 Cells upon Incubation with Heat-Killed NEC-Positive Associated Bacterium K. oxytoca or Non-Pathogenic E. coli
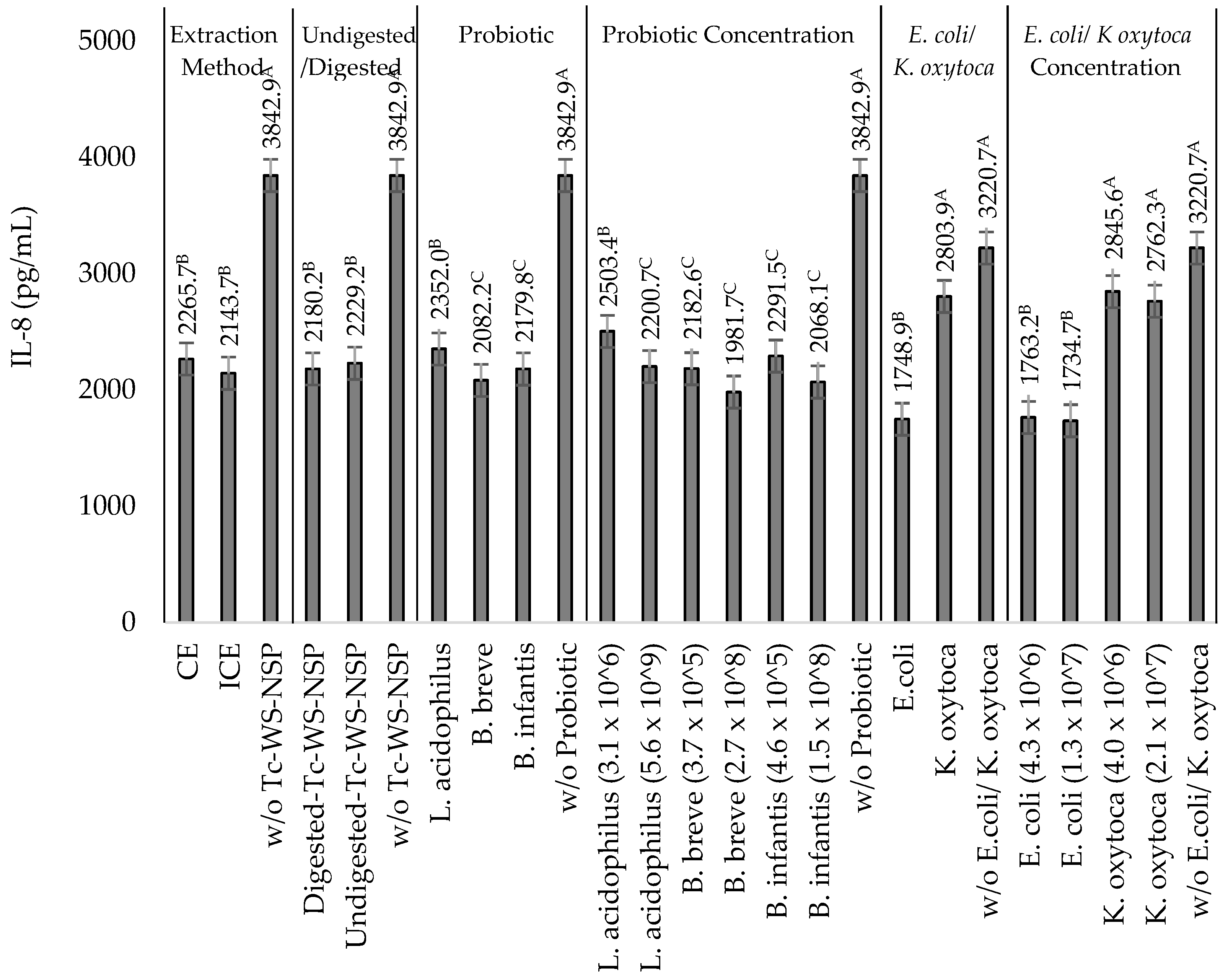
3.4. Effects of Undigested or Digested Tc-WS-NSPs on IL-8 Production by TNF-α Stimulated HT-29 Cells in the Presence of Heat-Killed NEC-Positive Associated Bacterium K. oxytoca or Non-Pathogenic E. coli
3.5. Effects of the Probiotics L. acidophilus, B. breve, and B. infantis on IL-8 Production by TNF-α Stimulated HT-29 Cells in the Presence of Heat-Killed NEC-Positive Associated Bacterium K. oxytoca or Non-Pathogenic E. coli
3.6. Effects of the Synbiotic Mixtures of the Undigested or Digested Tc-WS-NSPs and the Probiotics L. acidophilus, B. breve, and B. infantis on IL-8 Production by TNF-α Stimulated HT-29 Cells in the Presence of Heat-Killed NEC-Positive Associated Bacterium K. oxytoca or Non-Pathogenic E. coli
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Masi, A.C.; Stewart, C.J. The role of the preterm intestinal microbiome in sepsis and necrotising enterocolitis. Early Hum. Dev. 2019, 138, 104854. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.R.; Walker, W.A. Probiotics: Role in Pathophysiology and Prevention in Necrotizing Enterocolitis. Semin. Perinatol. 2008, 32, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Hintz, S.R.; Kendrick, D.E.; Stoll, B.J.; Vohr, B.R.; Fanaroff, A.A.; Donovan, E.F.; Poole, W.K.; Blakely, M.L.; Wright, L.; Higgins, R.; et al. Neurodevelopmental and Growth Outcomes of Extremely Low Birth Weight Infants After Necrotizing Enterocolitis. Pediatrics 2005, 115, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Raba, A.A.; O’Sullivan, A.; Miletin, J. Pathogenesis of necrotising enterocolitis: The impact of the altered gut microbiota and antibiotic exposure in preterm infants. Acta Paediatr. 2020, 110, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Zozaya, C.; Avila-Alvarez, A.; Argibay, S.; Oikonomopoulou, N.; Encinas, J.L.; Saenz de Pipaón, M.; Couce, M.L. Prevention, diagnosis and treatment of necrotising enterocolitis in new-borns less than 32 weeks at birth in Spain. Ann. Pediatr. 2020, 93, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Hau, E.-M.; Meyer, S.C.; Berger, S.; Goutaki, M.; Kordasz, M.; Kessler, U. Gastrointestinal sequelae after surgery for necrotising enterocolitis: A systematic review and meta-analysis. Arch. Dis. Child. Fetal Neonatal Ed. 2018, 104, F265–F273. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.S.; Goree, M.; Good, M. An Updated Overview of the Medical Management of Necrotizing Enterocolitis. In Necrotizing Enterocolitis: Pathogenesis, Diagnosis, and Treatment, 1st ed.; David, J.H., Ed.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2021; pp. 8–11. [Google Scholar] [CrossRef]
- Martin, C.R.; Dammann, O.; Allred, E.N.; Patel, S.; O’Shea, T.M.; Kuban, K.C.; Leviton, A. Neurodevelopment of extremely preterm infants who had necrotizing enterocolitis with or without late bacteraemia. J.Pediatr. 2010, 157, 751–756. [Google Scholar] [CrossRef]
- Blau, J.; Calo, J.M.; Dozor, D.; Sutton, M.; Alpan, G.; La Gamma, E.F. Transfusion-related acute gut injury: Necrotising enterocolitis in very low birth weight neonates after packed red blood cell transfusion. J. Pediatr. 2011, 158, 403–409. [Google Scholar] [CrossRef]
- Jin, Y.-T.; Duan, Y.; Deng, X.-K.; Lin, J. Prevention of necrotizing enterocolitis in premature infants—An updated review. World J. Clin. Pediatr. 2019, 8, 23–32. [Google Scholar] [CrossRef]
- Mayer, H. Anatomy of a disaster–overview of necrotising enterocolitis. Prof. Nurs. Today 2020, 24, 6–10. [Google Scholar]
- Qasem, W.; Alnaqi, A.; Jorgensen, S.; Friel, J. Probiotics and synbiotics in necrotizing enterocolitis: Where are we? J. Pediatr. Neonatal Care 2014, 1, 1–7. [Google Scholar] [CrossRef][Green Version]
- Nanthakumar, N.; Meng, D.; Goldstein, A.M.; Zhu, W.; Lu, L.; Uauy, R.; Llanos, A.; Claud, E.C.; Walker, W.A. The Mechanism of Excessive Intestinal Inflammation in Necrotizing Enterocolitis: An Immature Innate Immune Response. PLoS ONE 2011, 6, e17776. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Henry, K.C.; Abrahamsson, T.R.; Wu, R.Y.; Sherman, P.M. Probiotics, Prebiotics, and Synbiotics for the Prevention of Necrotizing Enterocolitis. Adv. Nutr. Int. Rev. J. 2016, 7, 928–937. [Google Scholar] [CrossRef]
- Masi, A.C.; Embleton, N.D.; A Lamb, C.; Young, G.; Granger, C.L.; Najera, J.; Smith, D.P.; Hoffman, K.L.; Petrosino, J.F.; Bode, L.; et al. Human milk oligosaccharide DSLNT and gut microbiome in preterm infants predicts necrotising enterocolitis. Gut 2020, 70, 2273–2282. [Google Scholar] [CrossRef] [PubMed]
- Sharif, S.; Meader, N.; Oddie, S.J.; Rojas-Reyes, M.X.; McGuire, W. Probiotics to prevent necrotising enterocolitis in very preterm or very low birth weight infants. Cochrane Database Syst. Rev. 2020, 2020, CD005496. [Google Scholar] [CrossRef]
- Duffield, S.D.; Clarke, P. Current use of probiotics to prevent necrotising enterocolitis. Arch. Dis. Child. Fetal Neonatal Ed. 2018, 104, F228. [Google Scholar] [CrossRef]
- Wong, C.B.; Iwabuchi, N.; Xiao, J.-Z. Exploring the Science behind Bifidobacterium breve M-16V in Infant Health. Nutrients 2019, 11, 1724. [Google Scholar] [CrossRef]
- Pehlevan, O.S.; Benzer, D.; Gursoy, T.; Karatekin, G.; Ovali, F. Synbiotics use for preventing sepsis and necrotizing enterocolitis in very low birth weight neonates: A randomized controlled trial. Clin. Exp. Pediatr. 2020, 63, 226–231. [Google Scholar] [CrossRef]
- Dilli, D.; Aydin, B.; Fettah, N.D.; Özyazıcı, E.; Beken, S.; Zenciroğlu, A.; Turan, Ö. The propre-save study: Effects of probiotics and prebiotics alone or combined on necrotising enterocolitis in very low birth weight infants. J.Pediatr. 2015, 166, 545–551. [Google Scholar] [CrossRef]
- Armanian, A.-M.; Sadeghnia, A.; Hoseinzadeh, M.; Mirlohi, M.; Feizi, A.; Salehimehr, N.; Saee, N.; Nazari, J. The Effect of Neutral Oligosaccharides on Reducing the Incidence of Necrotizing Enterocolitis in Preterm infants: A Randomized Clinical Trial. Int. J. Prev. Med. 2014, 5, 1387–1395. [Google Scholar]
- Lin, H.-C.; Hsu, C.-H.; Chen, H.-L.; Chung, M.-Y.; Hsu, J.-F.; Lien, R.-I.; Tsao, L.-Y.; Chen, C.-H.; Su, B.-H. Oral Probiotics Prevent Necrotizing Enterocolitis in Very Low Birth Weight Preterm Infants: A Multicenter, Randomized, Controlled Trial. Pediatrics 2008, 122, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Härtel, C.; Pagel, J.; Rupp, J.; Bendiks, M.; Guthmann, F.; Rieger-Fackeldey, E.; Heckmann, M.; Franz, A.; Schiffmann, J.-H.; Zimmermann, B.; et al. Prophylactic Use of Lactobacillus acidophilus/Bifidobacterium infantis Probiotics and Outcome in Very Low Birth Weight Infants. J. Pediatr. 2014, 165, 285–289.e1. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.A.; Salzman, N.H.; Bennett, S.H.; Barman, M.; Mills, D.; Marcobal, A.; Tancredi, D.J.; Bevins, C.L.; Sherman, M.P. A randomized placebo-controlled comparison of two prebiotic/probiotic combinations in preterm infants: Impact on weight gain, intestinal microbiota, and fecal short chain fatty acids. J. Pediatr. Gastroenterol. Nutr. 2009, 48, 216. [Google Scholar] [CrossRef] [PubMed]
- Nandhini, L.P.; Biswal, N.; Adhisivam, B.; Mandal, J.; Bhat, B.V.; Mathai, B. Synbiotics for decreasing incidence of necrotizing enterocolitis among preterm neonates—A randomized controlled trial. J. Maternal-Fetal Neonatal Med. 2016, 29, 821–825. [Google Scholar] [CrossRef]
- Weichert, S.; Schroten, H.; Adam, R. The Role of Prebiotics and Probiotics in Prevention and Treatment of Childhood Infectious Diseases. Pediatr. Infect. Dis. J. 2012, 31, 859–862. [Google Scholar] [CrossRef]
- Seo, Y.-M.; Lin, Y.-K.; Im, S.-A.; Sung, I.-K.; Youn, Y.-A. Interleukin 8 may predict surgical necrotizing enterocolitis in infants born less than 1500 g. Cytokine 2020, 137, 155343. [Google Scholar] [CrossRef]
- Akker, C.H.V.D.; van Goudoever, J.B.; Shamir, R.; Domellöf, M.; Embleton, N.D.; Hojsak, I.; Lapillonne, A.; Mihatsch, W.A.; Canani, R.B.; Bronsky, J.; et al. Probiotics and Preterm Infants: A Position Paper by the European Society for Paediatric Gastroenterology Hepatology and Nutrition Committee on Nutrition and the European Society for Paediatric Gastroenterology Hepatology and Nutrition Working Group for Probiotics and Prebiotics. J. Pediatr. Gastroenterol. Nutr. 2020, 70, 664–680. [Google Scholar] [CrossRef]
- Anwar, M.; Mros, S.; McConnell, M.; Bekhit, A.E.-D.A. Effects of extraction methods on the digestibility, cytotoxicity, prebiotic potential and immunomodulatory activity of taro (Colocasia esculenta) water-soluble non-starch polysaccharide. Food Hydrocoll. 2021, 121, 107068. [Google Scholar] [CrossRef]
- Manhivi, V.E.; Venter, S.; Amonsou, E.; Kudanga, T. Composition, thermal and rheological properties of polysaccharides from amadumbe (Colocasia esculenta) and cactus (Opuntia spp.). Carbohydr. Polym. 2018, 195, 163–169. [Google Scholar] [CrossRef]
- Nguimbou, R.M.; Boudjeko, T.; Njintang, N.Y.; Himeda, M.; Scher, J.; Mbofung, C.M.F. Mucilage chemical profile and antioxidant properties of giant swamp taro tubers. J. Food Sci. Technol. 2012, 51, 3559–3567. [Google Scholar] [CrossRef]
- Zhu, F. Structure, properties, and applications of aroid starch. Food Hydrocoll. 2016, 52, 378–392. [Google Scholar] [CrossRef]
- Nagata, C.L.P.; Andrade, L.A.; Pereira, J. Optimization of taro mucilage and fat levels in sliced breads. J. Food Sci. Technol. 2014, 52, 5890–5897. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Ramsden, L. Characterisation and yield of the arabinogalactan-protein mucilage of taro corms. J. Sci. Food Agric. 1999, 79, 671–674. [Google Scholar] [CrossRef]
- Sarkar, G.; Saha, N.R.; Roy, I.; Bhattacharyya, A.; Bose, M.; Mishra, R.; Rana, D.; Bhattacharjee, D.; Chattopadhyay, D. Taro corms mucilage/HPMC based transdermal patch: An efficient device for delivery of diltiazem hydrochloride. Int. J. Biol. Macromol. 2014, 66, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Chukwuma, C.I.; Islam, M.S.; Amonsou, E.O. A comparative study on the physicochemical, anti-oxidative, anti-hyperglycaemic and anti-lipidemic properties of amadumbe (Colocasia esculenta) and okra (Abelmoschus esculentus) mucilage. J. Food Biochem. 2018, 42, 1–12. [Google Scholar] [CrossRef]
- Park, H.-R.; Lee, H.-S.; Cho, S.Y.; Kim, Y.-S.; Shin, K.-S. Anti-metastatic effect of polysaccharide isolated from Colocasia esculenta is exerted through immunostimulation. Int. J. Mol. Med. 2012, 31, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro Pereira, P.; Bertozzi de Aquino Mattos, E.; Nitzsche Teixeira Fernandes Correa, A.C.; Afonso Vericimo, M.; Margaret Flosi Paschoalin, V. Anticancer and Immunomodulatory Benefits of Taro (Colocasia esculenta) Corms, an underexploited tuber crop. Int. J. Mol. Sci. 2020, 22, 265. [Google Scholar] [CrossRef]
- Li, H.; Dong, Z.; Liu, X.; Chen, H.; Lai, F.; Zhang, M. Structure characterization of two novel polysaccharides from Colocasia esculenta (taro) and a comparative study of their immunomodulatory activities. J. Funct. Foods 2018, 42, 47–57. [Google Scholar] [CrossRef]
- Anwar, M.; Birch, E.J.; Ding, Y.; Bekhit, A.E.-D. Water-soluble non-starch polysaccharides of root and tuber crops: Extraction, characteristics, properties, bioactivities, and applications. Crit. Rev. Food Sci. Nutr. 2020, 62, 2309–2341. [Google Scholar] [CrossRef]
- Anwar, M.; McConnell, M.; Bekhit, A.E.-D. New freeze-thaw method for improved extraction of water-soluble non-starch polysaccharide from taro (Colocasia esculenta): Optimization and comprehensive characterization of physico-chemical and structural properties. Food Chem. 2021, 349, 129210. [Google Scholar] [CrossRef]
- Lin, H.; Huang, A.S. Chemical composition and some physical properties of a water-soluble gum in taro (Colocasia esculenta). Food Chem. 1993, 48, 403–409. [Google Scholar] [CrossRef]
- Schneider, K.P.; Gewessler, U.; Flock, T.; Heinzle, A.; Schenk, V.; Kaufmann, F.; Sigl, E.; Guebitz, G.M. Signal enhancement in polysaccharide based sensors for infections by incorporation of chemically modified laccase. New Biotechnol. 2012, 29, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xie, M.; Wan, P.; Chen, D.; Ye, H.; Chen, L.; Zeng, X.; Liu, Z. Digestion under saliva, simulated gastric and small intestinal conditions and fermentation in vitro by human intestinal microbiota of polysaccharides from Fuzhuan brick tea. Food Chem. 2018, 244, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, C.; Huang, Q.; Fu, X.; Liu, R.H. In vitro digestibility and prebiotic potential of a novel polysaccharide from Rosa roxburghii Tratt fruit. J. Funct. Foods 2018, 52, 408–417. [Google Scholar] [CrossRef]
- Anwar, M.A. Taro (Colocasia esculenta) Water-Soluble Non-Starch Polysaccharide: Extraction, Characteristics, Properties, and Biological Activities. Ph.D. Thesis, Department of Food Science, University of Otago, Dunedin, New Zealand, 2020. Available online: https://ourarchive.otago.ac.nz/handle/10523/10470 (accessed on 12 February 2021).
- Arulselvan, P.; Gothai, S.; Muniandy, K.; Esa, N.M.; Subbiah, S.K. Anticancer potential of Alternanthera sessilis extract on HT-29 human colon cancer cells. Asian Pac. J. Trop. Biomed. 2018, 8, 394. [Google Scholar] [CrossRef]
- Darby, A.; Lertpiriyapong, K.; Sarkar, U.; Seneviratne, U.; Park, D.S.; Gamazon, E.; Batchelder, C.; Cheung, C.; Buckley, E.M.; Taylor, N.S.; et al. Cytotoxic and Pathogenic Properties of Klebsiella oxytoca Isolated from Laboratory Animals. PLoS ONE 2014, 9, e100542. [Google Scholar] [CrossRef] [PubMed]
- Joainig, M.M.; Gorkiewicz, G.; Leitner, E.; Weberhofer, P.; Zollner-Schwetz, I.; Lippe, I.; Zechner, E.L. Cytotoxic effects of Klebsiella oxytoca strains isolated from patients with antibiotic-associated haemorrhagic colitis or other diseases caused by infections and from healthy subjects. J. Clin. Microbiol. 2010, 48, 817–824. [Google Scholar] [CrossRef]
- Martínez, F.; Gómez, S.; Rodriguez, P.; Páez, J.; Urbina, G.; Ortiz, B. Klebsiella pneumoniae y K. oxytoca aisladas de niños con diarrheal: Adherencia y citotoxicidad en líneas celulares. Revista de la Sociedad Venezolana de Microbiología 2005, 25, 236–243. [Google Scholar]
- Hering, N.A.; Fromm, A.; Bücker, R.; Gorkiewicz, G.; Zechner, E.; Högenauer, C.; Schulzke, J.-D.; Troeger, H. Tilivalline- and Tilimycin-Independent Effects of Klebsiella oxytoca on Tight Junction-Mediated Intestinal Barrier Impairment. Int. J. Mol. Sci. 2019, 20, 5595. [Google Scholar] [CrossRef]
- Shi, L. Bioactivities, isolation and purification methods of polysaccharides from natural products: A review. Int. J. Biol. Macromol. 2016, 92, 37–48. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Najarian, A.; Sharif, S.; Griffiths, M. Evaluation of protective effect of Lactobacillus acidophilus La-5 on toxicity and colonization of Clostridium difficile in human epithelial cells in vitro. Anaerobe 2018, 55, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Liang, N.; Luo, X.; Zhang, T.C. Lactobacillus acidophilus strain suppresses the transcription of proinflammatory-related factors in human HT-29 cells. J. Microbiol. Biotechnol. 2013, 23, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Candela, M.; Perna, F.; Carnevali, P.; Vitali, B.; Ciati, R.; Gionchetti, P.; Rizzello, F.; Campieri, M.; Brigidi, P. Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells: Adhesion properties, competition against enteropathogens and modulation of IL-8 production. Int. J. Food Microbiol. 2008, 125, 286–292. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, A.M.; O’Regan, P.; Fanning, A.; O’Mahony, C.; MacSharry, J.; Lyons, A.; Bienenstock, J.; O’Mahony, L.; Shanahan, F. Functional modulation of human intestinal epithelial cell responses by Bifidobacterium infantis and Lactobacillus salivarius. Immunology 2006, 118, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Doron, S.; Snydman, D.R. Risk and Safety of Probiotics. Clin. Infect. Dis. 2015, 60, S129–S134. [Google Scholar] [CrossRef]
- Whelan, K. Probiotics and prebiotics in the management of irritable bowel syndrome: A review of recent clinical trials and systematic reviews. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 581–587. [Google Scholar] [CrossRef]
- Hedin, C.; Whelan, K.; Lindsay, J.O. Evidence for the use of probiotics and prebiotics in inflammatory bowel disease: A review of clinical trials. Proc. Nutr. Soc. 2007, 66, 307–315. [Google Scholar] [CrossRef]
- Wildt, S.; Munck, L.K.; Vinter-Jensen, L.; Hanse, B.F.; Nordgaard-Lassen, I.; Christensen, S.; Avnstroem, S.; Rasmussen, S.N.; Rumessen, J.J. Probiotic treatment of collagenous colitis: A randomized, double-blind, placebo-controlled trial with Lactobacillus acidophilus and Bifidobacterium animalis subsp. Lactis. Inflamm. Bowel Dis. 2006, 12, 395–401. [Google Scholar] [CrossRef]
- MacDonald, T.T.; Monteleone, G. Immunity, Inflammation, and Allergy in the Gut. Science 2005, 307, 1920–1925. [Google Scholar] [CrossRef]
- Thomson, K.L.J. The Immunological Role of Enterocytes and Probiotics in Necrotizing Enterocolitis. Master Thesis, Department of Microbiology and Immunology, University of Otago, Dunedin, New Zealand, 2014. Available online: https://ourarchive-otago-ac-nz.ezproxy.otago.ac.nz/bitstream/handle/10523/4970/ThomsonKristianLJ2014MSc.pdf?sequence=1&isAllowed= (accessed on 16 September 2019).
- Brooks, H.J.; McConnell, M.A.; Corbett, J.; Buchan, G.S.; Fitzpatrick, C.E.; Broadbent, R.S. Potential prophylactic value of bovine colostrum in necrotizing enterocolitis in neonates: An in vitro study on bacterial attachment, antibody levels and cytokine production, FEMS Immunol. Med. Microbiol. 2006, 48, 347–354. [Google Scholar]
- Wallace, T.D.; Bradley, S.; Buckley, N.D.; Green-Johnson, J.M. Interactions of Lactic Acid Bacteria with Human Intestinal Epithelial Cells: Effects on Cytokine Production. J. Food Prot. 2003, 66, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Espinal-Ruiz, M.; Parada-Alfonso, F.; Restrepo-Sánchez, L.-P.; Narváez-Cuenca, C.-E. Inhibition of digestive enzyme activities by pectic polysaccharides in model solutions. Bioact. Carbohydr. Diet. Fibre 2014, 4, 27–38. [Google Scholar] [CrossRef]
- Liu, J.; Willför, S.; Xu, C. A review of bioactive plant polysaccharides: Biological activities, functionalization, and biomedical applications. Bioact. Carbohydr. Diet. Fibre 2015, 5, 31–61. [Google Scholar] [CrossRef]
- Brück, W.M.; Kelleher, S.L.; Gibson, G.R.; Nielsen, K.E.; Chatterton, D.E.W.; Lönnerdal, B. rRNA Probes Used to Quantify the Effects of Glycomacropeptide and α-Lactalbumin Supplementation on the Predominant Groups of Intestinal Bacteria of Infant Rhesus Monkeys Challenged with Enteropathogenic Escherichia coli. J. Pediatr. Gastroenterol. Nutr. 2003, 37, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; McCartney, A.L.; Rastall, R.A. Prebiotics and resistance to gastrointestinal infections. Br. J. Nutr. 2005, 93, S31–S34. [Google Scholar] [CrossRef]
- Asahara, T.; Nomoto, K.; Shimizu, K.; Watanuki, M.; Tanaka, R. Increased resistance of mice to Salmonella enterica serovar Typhimurium infection by synbiotic administration of Bifidobacteria and transgalactosylated oligosaccharides. J. Appl. Microbiol. 2001, 91, 985–996. [Google Scholar] [CrossRef]
- Cummings, J.H.; Macfarlane, G.T. Gastrointestinal effects of prebiotics. Br. J. Nutr. 2002, 87, S145–S151. [Google Scholar] [CrossRef]
- Meyer, P.; Tungland, B.; Causey, J.; Slavin, J. The immune effects of inulin in vitro and in vivo. Agro Food Ind. Hi-Tech 2000, 11, 18–20. [Google Scholar]
- Messing, J.; Niehues, M.; Shevtsova, A.; Borén, T.; Hensel, A. Antiadhesive Properties of Arabinogalactan Protein from Ribes nigrum Seeds against Bacterial Adhesion of Helicobacter pylori. Molecules 2014, 19, 3696. [Google Scholar] [CrossRef]
- Quintero, M.; Maldonado, M.; Perez-Munoz, M.; Jimenez, R.; Fangman, T.; Rupnow, J.; Wittke, A.; Russell, M.; Hutkins, R. Adherence Inhibition of Cronobacter sakazakii to Intestinal Epithelial Cells by Prebiotic Oligosaccharides. Curr. Microbiol. 2011, 62, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed]
- Gourbeyre, P.; Denery, S.; Bodinier, M. Probiotics, prebiotics, and synbiotics: Impact on the gut immune system and allergic reactions. J. Leukoc. Biol. 2011, 89, 685–695. [Google Scholar] [CrossRef]
- Mukai, T.; Kaneko, S.; Matsumoto, M.; Ohori, H. Binding of Bifidobacterium bifidum and Lactobacillus reuteri to the carbohydrate moieties of intestinal glycolipids recognized by peanut agglutinin. Int. J. Food Microbiol. 2003, 90, 357–362. [Google Scholar] [CrossRef]
- Motherway, M.O.; Zomer, A.; Leahy, S.C.; Reunanen, J.; Bottacini, F.; Claesson, M.J.; O’Brien, F.; Flynn, K.; Casey, P.G.; Munoz, J.A.M.; et al. Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl. Acad. Sci. USA 2011, 108, 11217–11222. [Google Scholar] [CrossRef]
- Huberts, D.H.; van der Klei, I.J. Moonlighting proteins: An intriguing mode of multitasking. Biochim. Biophys. Acta-Mol. Cell Res. 2010, 1803, 520–525. [Google Scholar] [CrossRef]
- Preising, J.; Philippe, D.; Gleinser, M.; Wei, H.; Blum, S.; Eikmanns, B.J.; Niess, J.-H.; Riedel, C.U. Selection of Bifidobacteria Based on Adhesion and Anti-Inflammatory Capacity In Vitro for Amelioration of Murine Colitis. Appl. Environ. Microbiol. 2010, 76, 3048–3051. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Sánchez, B.; Milani, C.; Ventura, M.; Margolles, A.; Ruas-Madiedo, P. Genomic Overview and Biological Functions of Exopolysaccharide Biosynthesis in Bifidobacterium spp. Appl. Environ. Microbiol. 2014, 80, 9–18. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Isolate | Bacterial Isolate Concentration (CFU/mL) | Tc-WS-NSP | IL-8 Reduction (%) | |
|---|---|---|---|---|
| K. oxytoca | 4.0 × 106 | CE | Undigested | 17.2 ± 3.0 BC,M |
| Digested | 21.3 ± 2.4 AB,LM | |||
| ICE | Undigested | 19.2 ± 0.8 AB,LM | ||
| Digested | 24.5 ± 3.4 A,L | |||
| 2.1 × 107 | CE | Undigested | 18.1 ± 1.2 ABC,M | |
| Digested | 21.8 ± 1.0 AB,LM | |||
| ICE | Undigested | 16.8 ± 2.0 BC,M | ||
| Digested | 20.5 ± 2.3 AB,LM | |||
| E. coli | 4.3 × 106 | CE | Undigested | 8.4 ± 3.0 DE,MN |
| Digested | 10.0 ± 1.3 DE,LM | |||
| ICE | Undigested | 5.2 ± 2.0 E,N | ||
| Digested | 12.2 ± 0.8 CD,MN | |||
| 1.3 × 107 | CE | Undigested | 8.1 ± 2.4 DE,MN | |
| Digested | 16.6 ± 2.6 BC,L | |||
| ICE | Undigested | 8.1 ± 0.7 DE,MN | ||
| Digested | 17.1 ± 2.7 BC,L | |||
| Bacterial Isolate | L. acidophilus | IL-8 Reduction (%) | B. breve | IL-8 Reduction (%) | B. infantis | IL-8 Reduction (%) | |
|---|---|---|---|---|---|---|---|
| K. oxytoca | 4.0 × 106 | 3.1 × 106 | 29.1±2.2 H,NO,X | 3.73 × 105 | 31.7 ± 1.0 EFGH,N,Y | 1.53 × 105 | 29.5 ± 1.0 GH,N,X |
| 2.1 × 107 | 28.5 ± 2.5 H,O,X | 31.7 ± 1.3 EFGH,N,Y | 30.2 ± 1.0 FGH,MN,X | ||||
| 4.0 × 106 | 5.6 × 109 | 32.3 ± 1.5 EFGH,MNO,X | 2.70 × 108 | 36.3 ± 2.7 CDEFGH,N,XY | 4.56 × 108 | 32.0 ± 2.7 EFGH,LMN,X | |
| 2.1 × 107 | 31.2 ± 2.9 EFGH,MNO,X | 37.8 ± 1.9 BCDEFGH,MN,X | 33.1 ± 2.9 DEFGH,LMN,X | ||||
| E. coli | 4.3 × 106 | 3.1 × 106 | 37.5 ± 2.5 BCDEFGH,LMN,X | 3.73 × 105 | 45.9 ± 4.1 ABC,LM,X | 1.53 × 105 | 41.2 ± 3.9 ABCDEF,LMN,X |
| 1.3 × 107 | 35.9 ± 5.1 CDEFGH,LMNO,X | 46.7 ± 5.2 ABC,LM,X | 42.5 ± 4.8 ABCDE,LM,X | ||||
| 4.3 × 106 | 5.6 × 109 | 39.6 ± 2.8 ABCDEFGH,LM,X | 2.70 × 108 | 48.4 ± 4.0 AB,L,X | 4.56 × 108 | 44.1 ± 7.5 ABCD,L,X | |
| 1.3 × 107 | 40.9 ± 3.3 ABCDEFG,L,X | 50.2 ± 3.3 A,L,X | 44.3 ± 7.3 ABCD,L,X | ||||
| T WS-NSP | L. acidophilus (CFU/mL) | E. coli (CFU/mL) | IL-8 Reduction (%) | K. oxytoca (CFU/mL) | IL-8 Reduction (%) | |
|---|---|---|---|---|---|---|
| CE | Undigested | 3.08 × 106 | 4.33 × 106 | 43.0 ± 1.8 ABCDE,L,Q,X | 4.00 × 106 | 32.5 ± 3.8 E,L,Q,X |
| Digested | 43.8 ± 1.4 ABCDE,L,Q,X | 33.4 ± 5.4 CDE,L,Q,X | ||||
| ICE | Undigested | 41.8 ± 4.4 ABCDE,L,Q,X | 32.5 ± 4.1 E,L,Q,X | |||
| Digested | 43.6 ± 2.4 ABCDE,L,Q,X | 33.2 ± 4.1 CDE,L,Q,X | ||||
| CE | Undigested | 5.57 × 109 | 44.4 ± 2.2 ABCD,L,Q,X | 38.5 ± 3.8 ABCDE,L,Q,X | ||
| Digested | 45.0 ± 3.7 AB,L,Q,X | 40.3 ± 2.8 ABCDE,L,Q,X | ||||
| ICE | Undigested | 44.5 ± 3.5 ABC,L,Q,X | 40.8 ± 1.9 ABCDE,L,Q,X | |||
| Digested | 46.3 ± 4.3 A,L,Q,X | 41.3 ± 2.5 ABCDE,L,Q,X | ||||
| CE | Undigested | 3.08 × 106 | 1.33 × 107 | 40.5 ± 1.7 ABCDE,L,Q,X | 2.05 × 107 | 33.0 ± 4.7 E,L,Q,X |
| Digested | 43.7 ± 1.2 ABCDE,L,Q,X | 34.8 ± 6.4 BCDE,L,Q,X | ||||
| ICE | Undigested | 42.8 ± 4.2 ABCDE,L,Q,X | 33.1 ± 4.4 DE,L,Q,X | |||
| Digested | 43.7 ± 3.1 ABCDE,L,Q,X | 34.9 ± 5.3 BCDE,L,Q,X | ||||
| CE | Undigested | 5.57 × 109 | 43.2 ± 4.0 ABCDE,L,Q,X | 40.4 ± 2.7 ABCDE,L,Q,X | ||
| Digested | 45.7 ± 2.5 AB,L,Q,X | 40.8 ± 2.1 ABCDE,L,Q,X | ||||
| ICE | Undigested | 45.6 ± 2.4 AB,L,Q,X | 41.2 ± 3.3 ABCDE,L,Q,X | |||
| Digested | 46.8 ± 2.9 A,L,Q,X | 42.0 ± 2.5 ABCDE,L,Q,X | ||||
| T WS-NSP | B. breve (CFU/mL) | E. coli (CFU/mL) | IL-8 Reduction (%) | K. oxytoca (CFU/mL) | IL-8 Reduction (%) | |
|---|---|---|---|---|---|---|
| CE | Undigested | 3.7 × 105 | 4.3 × 106 | 51.7 ± 3.7 ABC,L,Q,X | 4.0 × 106 | 38.2 ± 2.7 E,L,Q,X |
| Digested | 51.5 ± 1.9 ABCD,L,Q,X | 39.3 ± 2.6 E,L,Q,X | ||||
| ICE | Undigested | 51.4 ± 2.6 ABCD,L,Q,X | 38.6 ± 4.1 E,L,Q,X | |||
| Digested | 52.3 ± 3.1 AB,L,Q,X | 39.2 ± 4.1 E,L,Q,X | ||||
| CE | Undigested | 2.7 × 108 | 54.4 ± 2.6 A,L,Q,X | 40.3 ± 3.3 E,L,Q,X | ||
| Digested | 55.2 ± 3.0 A,L,Q,X | 41.5 ± 2.3 E,L,Q,X | ||||
| ICE | Undigested | 54.5 ± 2.1 A,L,Q,X | 42.0 ± 1.9 E,L,Q,X | |||
| Digested | 55.2 ± 2.3 A,L,Q,X | 43.2 ± 2.5 DE,L,Q,X | ||||
| CE | Undigested | 3.7 × 105 | 1.3 × 107 | 54.0 ± 2.1 A,L,Q,X | 2.1 × 107 | 41.4 ± 4.1 E,L,Q,X |
| Digested | 54.4 ± 2.4 A,L,Q,X | 41.9 ± 5.9 E,L,Q,X | ||||
| ICE | Undigested | 54.3 ± 2.4 A,L,Q,X | 41.4 ± 3.8 E,L,Q,X | |||
| Digested | 55.0 ± 1.9 A,L,Q,X | 42.0 ± 4.7 E,L,Q,X | ||||
| CE | Undigested | 2.7 × 108 | 56.0 ± 2.1 A,L,Q,X | 43.3 ± 2.1 CDE,L,Q,X | ||
| Digested | 57.1 ± 2.4 A,L,Q,X | 44.2 ± 1.6 BCDE,L,Q,X | ||||
| ICE | Undigested | 56.1 ± 2.6 A,L,Q,X | 43.7 ± 3.3 CDE,L,Q,X | |||
| Digested | 58.1 ± 2.3 A,L,Q,X | 45.0 ± 2.5 BCDE,L,Q,X | ||||
| T WS-NSP | B. infantis (CFU/mL) | E. coli (CFU/mL) | IL-8 Reduction (%) | K. oxytoca (CFU/mL) | IL-8 Reduction (%) | |
|---|---|---|---|---|---|---|
| CE | Undigested | 4.56 × 105 | 4.33 × 106 | 49.2 ± 3.1 ABCDEF,L,Q,X | 4.00 × 106 | 34.1 ± 2.3 G,L,S,X |
| Digested | 50.6 ± 2.9 ABCD,L,Q,X | 35.4 ± 1.3 G,L,RS,X | ||||
| ICE | Undigested | 49.3 ± 2.2 ABCDEF,L,Q,X | 35.2 ± 1.5 G,L,RS,X | |||
| Digested | 51.4 ± 3.4 ABC,L,Q,X | 35.5 ± 1.7 G,L,RS,X | ||||
| CE | Undigested | 1.53 × 108 | 52.0 ± 4.1 ABC,L,Q,X | 38.3 ± 1.4 G,L,QRS,X | ||
| Digested | 52.4 ± 3.0 AB,L,Q,X | 38.9 ± 1.1 G,L,QR,X | ||||
| ICE | Undigested | 52.4 ± 4.5 AB,L,Q,X | 39.4 ± 1.1 FG,L,QR,X | |||
| Digested | 54.2 ± 3.6 A,L,Q,X | 40.9 ± 2.1 DEFG,L,Q,X | ||||
| CE | Undigested | 4.56 × 105 | 1.33 × 107 | 50.7 ± 1.3 ABCD,L,Q,X | 2.05 × 107 | 37.8 ± 1.9 G,L,Q,X |
| Digested | 52.5 ± 2.9 AB,L,Q,X | 39.7 ± 5.7 FG,L,Q,X | ||||
| ICE | Undigested | 50.4 ± 2.1 ABCDE,L,Q,X | 39.7 ± 5.1 FG,L,Q,X | |||
| Digested | 52.1 ± 0.7 ABC,L,Q,X | 40.3 ± 4.8 EFG,L,Q,X | ||||
| CE | Undigested | 1.53 × 108 | 52.4 ± 2.8 AB,L,Q,X | 41.1 ± 3.4 DEFG,L,Q,X | ||
| Digested | 53.1 ± 3.1 AB,L,Q,X | 42.2 ± 4.6 CDEFG,L,Q,X | ||||
| ICE | Undigested | 52.1 ± 1.2 ABC,L,Q,X | 42.1 ± 4.5 CDEFG,L,Q,X | |||
| Digested | 53.7 ± 2.3 A,L,Q,X | 43.1 ± 5.1 BCDEFG,L,Q,X | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anwar, M.; Mros, S.; McConnell, M.; Bekhit, A.E.-D.A. Effects of Taro (Colocasia esculenta) Water-Soluble Non-Starch Polysaccharide, Lactobacillus acidophilus, Bifidobacterium breve, Bifidobacterium infantis, and Their Synbiotic Mixtures on Pro-Inflammatory Cytokine Interleukin-8 Production. Nutrients 2022, 14, 2128. https://doi.org/10.3390/nu14102128
Anwar M, Mros S, McConnell M, Bekhit AE-DA. Effects of Taro (Colocasia esculenta) Water-Soluble Non-Starch Polysaccharide, Lactobacillus acidophilus, Bifidobacterium breve, Bifidobacterium infantis, and Their Synbiotic Mixtures on Pro-Inflammatory Cytokine Interleukin-8 Production. Nutrients. 2022; 14(10):2128. https://doi.org/10.3390/nu14102128
Chicago/Turabian StyleAnwar, Mylene, Sonya Mros, Michelle McConnell, and Alaa El-Din A. Bekhit. 2022. "Effects of Taro (Colocasia esculenta) Water-Soluble Non-Starch Polysaccharide, Lactobacillus acidophilus, Bifidobacterium breve, Bifidobacterium infantis, and Their Synbiotic Mixtures on Pro-Inflammatory Cytokine Interleukin-8 Production" Nutrients 14, no. 10: 2128. https://doi.org/10.3390/nu14102128
APA StyleAnwar, M., Mros, S., McConnell, M., & Bekhit, A. E.-D. A. (2022). Effects of Taro (Colocasia esculenta) Water-Soluble Non-Starch Polysaccharide, Lactobacillus acidophilus, Bifidobacterium breve, Bifidobacterium infantis, and Their Synbiotic Mixtures on Pro-Inflammatory Cytokine Interleukin-8 Production. Nutrients, 14(10), 2128. https://doi.org/10.3390/nu14102128