
Iodine Content of Wild and Farmed Seafood and Its Estimated Contribution to UK Dietary Iodine Intake


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Iodine Analysis
2.3. Seafood Contribution to Recommended Intake Levels
2.4. Statistical Analysis
3. Results and Discussion
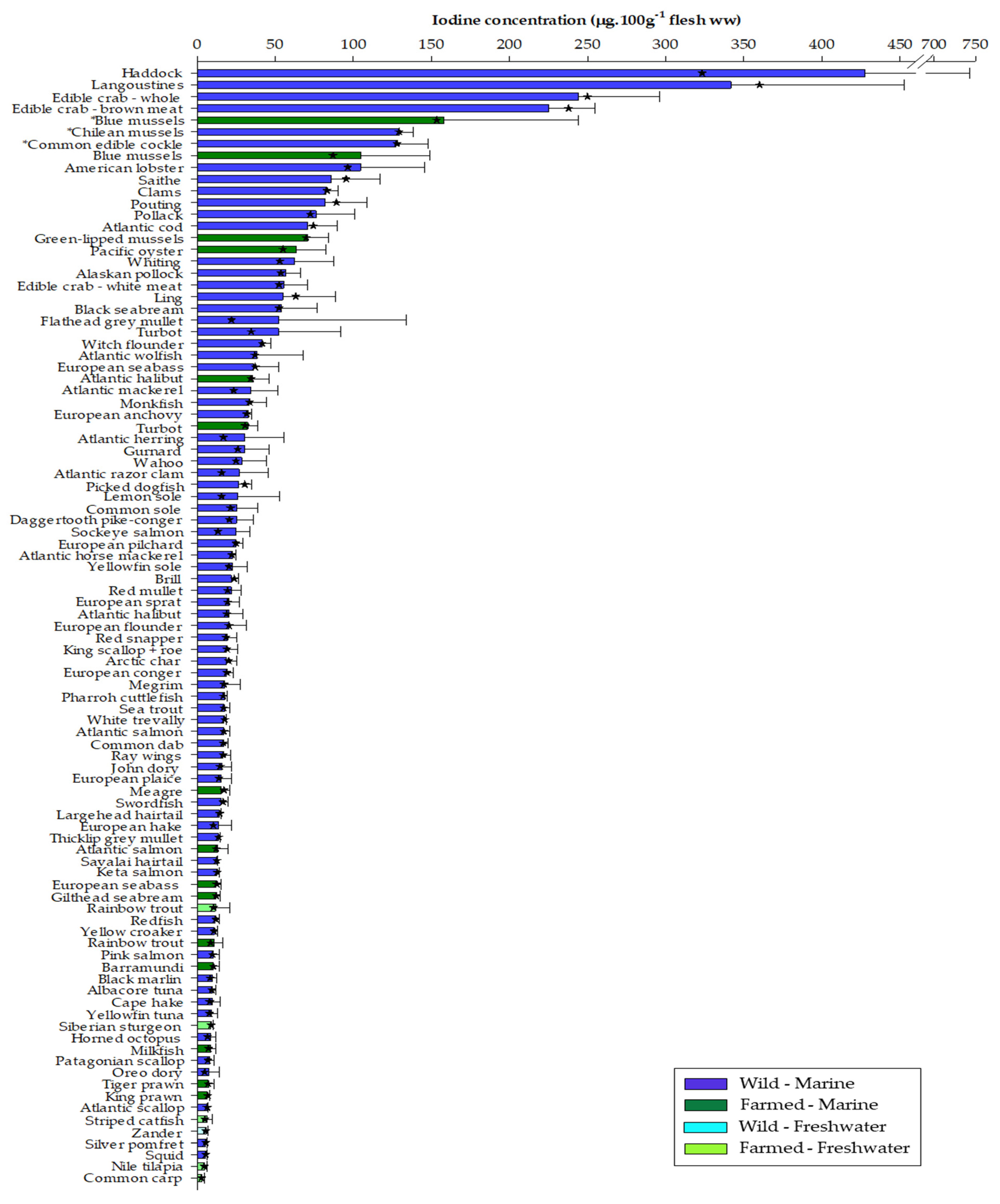
3.1. Iodine Content of Seafood
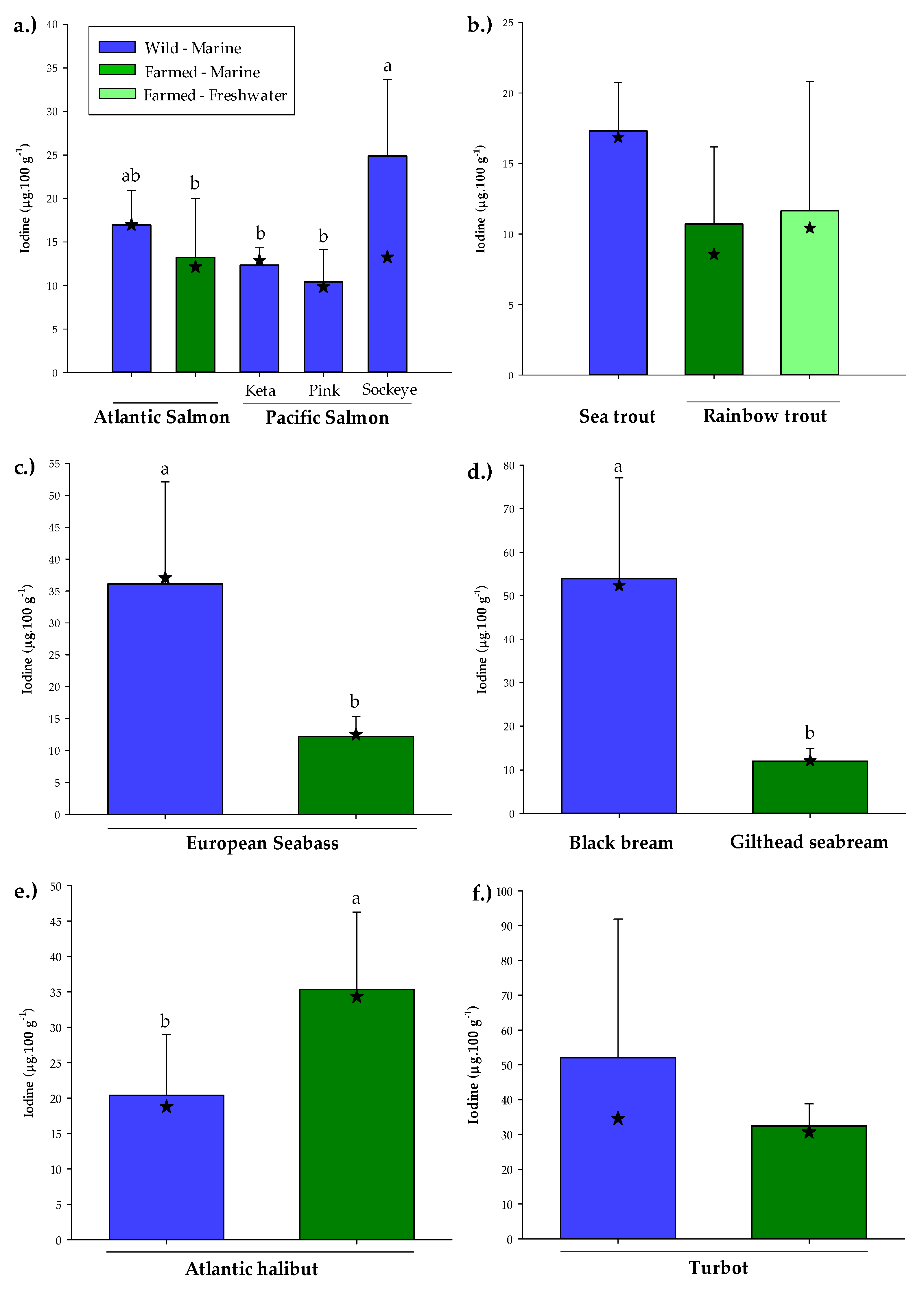
3.2. Wild Versus Farmed Seafood
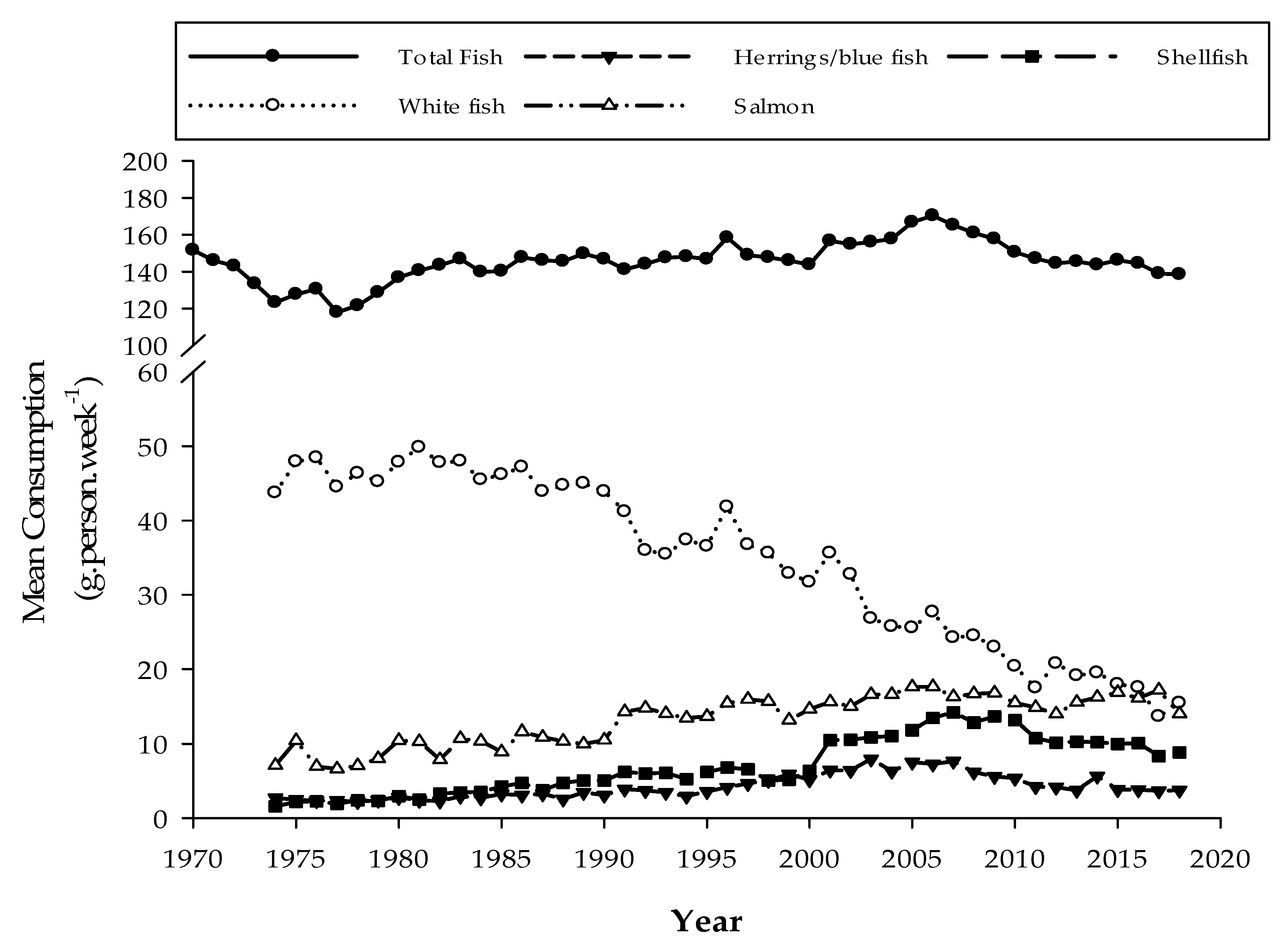
3.3. Seafood Contribution to Human Iodine Intake
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fuge, R.; Johnson, C.C. Iodine and human health, the role of environmental geochemistry and diet, a review. Appl. Geochem. 2015, 63, 282–302. [Google Scholar] [CrossRef]
- Lall, S.P. The Minerals. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 259–308. [Google Scholar]
- Lall, S.P. The Minerals. In Fish Nutrition, 4th ed.; Hardy, R.W., Kaushik, S.J., Eds.; Academic Press: London, UK, 2021; pp. 469–554. [Google Scholar]
- Bath, S.C.; Steer, C.D.; Golding, J.; Emmett, P.; Rayman, M.P. Effect of inadequate iodine status in UK pregnant women on cognitive outcomes in their children: Results from the Avon Longitudinal Study of Parents and Children (ALSPAC). Lancet 2013, 382, 331–337. [Google Scholar] [CrossRef]
- Bath, S.C.; Rayman, M.P. A review of the iodine status of UK pregnant women and its implications for the off spring. Environ. Geochem. Health 2015, 37, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Scientific Advisory Committee on Nutrition (SACN). Statement on Iodine and Health. February 2014. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/339439/SACN_Iodine_and_Health_2014.pdf (accessed on 10 March 2021).
- World Health Organization (WHO). United Nations Children’s Fund & International Council for the Control of Iodine Deficiency Disorders. In Assessment of Iodine Deficiency Disorders and Monitoring Their Elimination, 3rd ed.; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- EFSA. European Food Safety Authority (EFSA) Scientific Opinion on Dietray Reference Values for iodine: EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). EFSA J. 2014, 12, 3660. [Google Scholar] [CrossRef] [Green Version]
- National Health and Medical Research Council, Australian Government Department of Health and Ageing, New Zealand Ministry of Health. Nutrient Values for Australia and New Zealand; National Health and Medical Research Council: Canberra, Australia, 2006. Available online: https://www.nhmrc.gov.au/sites/default/files/images/nutrient-reference-dietary-intakes.pdf (accessed on 23 September 2021).
- Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academy Press: Washington, DC, USA, 2001. [Google Scholar]
- Carlsen, M.H.; Anderson, L.F.; Dahl, L.; Norberg, N.; Hjartåker, A. New iodine food composition database and updated calculations of iodine intake among Norwegians. Nutrients 2018, 10, 930. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Iodine Deficiency in Europe: A Continuing Public Health Problem; Andersson, M., de Benoist, B., Darnton-Hill, I., Delange, F., Eds.; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Woodside, J.V.; Mullan, K.R. Iodine status in UK—An accidental public health triumph gone sour. Clin. Endocrinol. 2021, 94, 692–699. [Google Scholar] [CrossRef]
- Zimmermann, M.B.; Andersson, M. Update on iodine status worldwide. Curr. Opin. Endocrinol. Diabetes Obes. 2012, 19, 382–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, L.; Johansson, L.; Julshamn, K.; Meltzer, H.M. The iodine content of Norwegian foods and diets. Public Health Nutr. 2004, 7, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Fordyce, F. Database of the Iodine Content of Food and Diets Populated with Data from Published Literature; British Geological Survey Commissioned Report; NERC Open Research Archive: Nottingham, UK, 2003. [Google Scholar]
- Rasmussen, L.B.; Andersen, S.; Ovesen, L.; Laurberg, P. Iodine intake and food choice. In Comprehensive Handbook of Iodine: Nutritional, Biochemical, Pathological and Therapeutic Aspects, 1st ed.; Preedy, V.R., Burrow, G.N., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2009; pp. 346–360. [Google Scholar]
- Roberts, C.; Steer, T.; Maplethorpe, N.; Cox, L.; Nicholson, S.; Page, P.; Swan, G. National Diet and Nutrition Survey. Results from Years 7-8 (Combined) of the Rolling Programme (2014/15 to 2015/16); PHE Publication gateway number: 2017851; Public Health England: London, UK, 2018. [Google Scholar]
- Van der Reijden, O.L.; Zimmermann, M.D.; Galetti, V. Iodine in dairy milk: Sources, concentrations and importance to human health. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Coneyworth, L.J.; Coulthard, L.C.H.A.; Bailey, E.H.; Young, S.D.; Stubberfield, J.; Parsons, L.; Saunders, N.; Watson, E.; Homer, E.M.; Welham, S.J.M. Geographical and seasonal variation in iodine content of cow’s milk in the UK and consequences for the consumer’s supply. J. Trace Elem. Med. Biol. 2020, 59, 126453. [Google Scholar] [CrossRef]
- Flachowsky, G.; Franke, K.; Meyer, U.; Leiterer, M.; Schöne, F. Influencing factors on iodine content of cow milk. Eur. J. Nutr. 2014, 53, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Flynn, A. Minerals and trace elements in milk. Adv. Food Nutr. Res. 1992, 36, 209–252. [Google Scholar] [CrossRef]
- Stevenson, M.C.; Drake, C.; Givens, D.I. Further studies on the iodine concentration of conventional, organic and UHT semi-skimmed milk at retail in the UK. Food Chem. 2018, 239, 551–555. [Google Scholar] [CrossRef]
- Hou, X. Iodine speciation in foodstuffs, tissues, and environmental samples: Iodine species and analytical methods. In Comprehensive Handbook of Iodine: Nutritional, Biochemical, Pathological and Therapeutic Aspects, 1st ed.; Preedy, V.R., Burrow, G.N., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2009; pp. 139–150. [Google Scholar]
- Ito, K.; Hirokawa, T. Iodine and iodine species in seawater: Speciation, distribution and dynamics. In Comprehensive Handbook of Iodine: Nutritional, Biochemical, Pathological and Therapeutic Aspects, 1st ed.; Preedy, V.R., Burrow, G.N., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2009; pp. 83–91. [Google Scholar]
- EFSA. European Food Safety Authority (EFSA) Opinion of the scientific panel on contaminants in the food chain on a request from the European Parliament related to the safety assessment of wild and farmed fish. EFSA J. 2005, 236, 1–118. [Google Scholar] [CrossRef]
- Rimm, E.B.; Appel, L.J.; Chiuve, S.E.; Djoussé, L.; Engler, M.B.; Kris-Etherton, P.M.; Mozaffarian, D.; Siscovick, D.S.; Lichtenstein, A.H. Seafood long-chain n-3 polyunsaturated fatty acids and cardiovascular disease: A Science Advisory form the American Heart Association. Circulation 2018, 138, e35–e47. [Google Scholar] [CrossRef]
- Scientific Advisory Committee on Nutrition (SACN) and Committee on Toxicity (COT). Advice on Fish Consumption: Benefits and Risks; The Stationary Office: Norwich, UK, 2004; p. 222. [Google Scholar]
- World Health Organization (WHO). Technical Report No. 916. Diet Nutrition and the Prevention of Chronic Diseases; WHO: Geneva, Switzerland, 2003. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture (SOFIA); Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Cai, J.; Zhou, X.; Yan, X.; Lucente, D.; Lagana, C. Top 10 Species Groups in Global Aquaculture 2017. Food and Agriculture Organization of the United Nations, FAO Fisheries and Aquaculture Department. Available online: http://www.fao.org/3/ca5224en/CA5224EN.pdf (accessed on 30 January 2020).
- Aas, T.S.; Ytrestøyl, T.; Åsgård, T. Utilization of feed resources in the production of Atlantic salmon (Salmo salar) in Norway: An update for 2016. Aquac. Rep. 2019, 15, 100216. [Google Scholar] [CrossRef]
- Betancor, M.B.; Dam, T.M.; Walton, J.; Morken, T.; Campbell, P.J.; Tocher, D.R. Modulation of selenium tissue distribution and seleno protein expression in Atlantic salmon (Salmo salar L.) fed diets with graded levels of plant ingredients. Br. J. Nutr. 2016, 115, 1325–1338. [Google Scholar] [CrossRef] [Green Version]
- Sissener, H.; Julshamn, K.; Espe, M.; Lunestad, B.T.; Hemre, G.I.; Waagbø, R.; Måge, A. Surveillance of selected nutrients, additives and undesirables in commercial Norwegian fish feeds in the years 2000–2010. Aquac. Nutr. 2013, 19, 555–572. [Google Scholar] [CrossRef]
- Sprague, M.; Dick, J.R.; Tocher, D.R. Impact of sustainable feeds on omega-3 long-chain fatty acids in farmed Atlantic salmon, 2006–2015. Sci. Rep. 2010, 6, 21892. [Google Scholar] [CrossRef] [Green Version]
- De Roos, B.; Sneddon, A.A.; Sprague, M.; Horgan, G.W.; Brouwer, I.A. The potential impact of compositional changes in farmed fish on its health-giving properties: Is it time to reconsider current dietary recommendations? Public Health Nutr. 2017, 20, 2042–2049. [Google Scholar] [CrossRef] [Green Version]
- Dahl, L.; Meltzer, H.M. The iodine content of foods and diets: Norwegian perspectives. In Comprehensive Handbook of Iodine: Nutritional, Biochemical, Pathological and Therapeutic Aspects, 1st ed.; Preedy, V.R., Burrow, G.N., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2009; pp. 360–391. [Google Scholar]
- Delgado, I.; Ventura, M.; Gueifão, S.; Coelho, I.; Nascimento, A.C.; Silva, J.A.L.; Castanheira, I. 12th IFDC 2017 special issue—Iodine, selenium and iron contents in Portuguese key foods as consumed. J. Food Compost. Anal. 2019, 79, 39–46. [Google Scholar] [CrossRef]
- Haldimann, M.; Alt, A.; Blanc, A.; Blondeau, K. Iodine content of food groups. J. Food Compost. Anal. 2005, 18, 461–471. [Google Scholar] [CrossRef]
- Nerhus, I.; Markhus, M.W.; Nilsen, B.M.; Øyen, J.; Maage, A.; Ødegård, E.R.; Midtbø, L.K.; Frantzen, S.; Kögel, T.; Graff, I.E.; et al. Iodine content of six fish species, Norwegian dairy products and hen’s eggs. Food Nutr. Res. 2018, 62, 1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pehrsson, P.R.; Patterson, K.Y.; Spungen, J.H.; Wirtz, M.S.; Andrews, K.W.; Dwyer, J.T.; Swanson, C.A. Iodine in food- and dietary supplement—composition databases. Am. J. Clin. Nutr. 2016, 104, 868S–876S. [Google Scholar] [CrossRef] [Green Version]
- Wenlock, R.W.; Buss, D.H.; Moxon, R.E.; Bunton, N.G. Trace nutrients 4 that of farmed salmon. Iodine in British food. Br. J. Nutr. 1982, 47, 381–390. [Google Scholar] [CrossRef]
- Foods Standards Agency (FSA). McCance and Widdowson’s the Composition of Foods, 7th ed.; Royal Society of Chemistry: Cambridge, UK, 2014. [Google Scholar]
- Traka, M.H.; Plumb, J.; Berry, R.; Pinchen, H.; Finglas, P.M. Maintaining and updating food composition datasets for multiple users and novel technologies: Current challenges from a UK perspective. Nutr. Bull. 2020, 45, 230–240. [Google Scholar] [CrossRef]
- Department of Health (DH). Nutrient Analysis of Fish and Fish Products. Analytical Report. 2013; 88p. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/167923/Nutrient_analysis_of_fish_and_fish_products_-_Analytical_Report.pdf (accessed on 28 March 2020).
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; AOAC International: Gaithersburg, Maryland, MD, USA, 2000. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). Fisheries and Aquaculture Division. ASFIS List of Species for Fishery Statistics Purposes. Available online: https://www.fao.org/fishery/collection/asfis/en (accessed on 16 March 2021).
- Food and Agriculture Organization of the United Nations (FAO), Fisheries and Aquaculture Division. FAO Major Fishing Areas. Available online: https://www.fao.org/fishery/area/search/en (accessed on 16 March 2021).
- Fecher, P.; Goldmann, I.; Nagengast, A. Determination of iodine in food samples by inductively coupled plasma mass spectrometry after alkaline extraction. J. Anal. At. Spectrom. 1998, 13, 977–982. [Google Scholar] [CrossRef]
- Public Health England (PHE). National Diet and Nutrition Survey (NDNS). Available online: https://www.gov.uk/government/collections/national-diet-and-nutrition-survey#current-ndns-results (accessed on 22 February 2021).
- Julshamn, K.; Dahl, L.; Eckhoff, K. Determination of iodine in seafood by inductively coupled plasma/mass spectrometry. J. AOAC Int. 2001, 84, 1976–1983. [Google Scholar] [CrossRef] [Green Version]
- Karl, H.; Münkner, W.; Krause, S.; Bagge, I. Determination, spatial variation and distribution of iodine in fish. Dtsch. Lebensm.-Rundsch. 2001, 97, 89–96. [Google Scholar]
- Bell, J.G. Current aspects of lipid nutrition in fish farming. In Biology of Farmed Fish; Black, K.D., Pickering, A.D., Eds.; Academic Press Ltd.: Sheffield, UK, 1998; pp. 114–145. [Google Scholar]
- Karl, H.; Basak, S.; Ziebell, S.; Quast, P. Changes in the iodine content in fish during household preparation and smoking. Dtsch. Lebensm.-Rundsch. 2005, 101, 1–6. [Google Scholar]
- Bell, J.G.; McEvoy, J.; Webster, J.L.; McGhee, F.; Millar, R.M.; Sargent, J.R. Flesh lipid and carotenoid composition of Scottish farmed Atlantic salmon (Salmo salar). J. Agric. Food Chem. 1998, 46, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Katikou, P.; Hughes, S.I.; Robb, D. Lipid distribution within Atlantic salmon (Salmo salar) fillets. Aquaculture 2001, 202, 89–99. [Google Scholar] [CrossRef]
- Lall, S.P. Macro and trace elements in fish and shellfish. In Fish and Fishery Products; Ruiter, A., Ed.; CAB International: Wallingford, UK, 1995; pp. 187–213. [Google Scholar]
- VKM. Benefit-Risk Assessment of Fish and Fish Products in the Norwegian Diet—An Update; VKM Report 15; Scientific Opinion of the Scientific Steering Committee: Oslo, Norway, 2014; 293p, Available online: https://vkm.no/download/18.2994e95b15cc54507161ea1a/1498222018046/0a646edc5e.pdf (accessed on 21 February 2021).
- Bourre, J.M.; Paquotte, P. Seafood (wild and farmed) for the elderly: Contribution to the dietary intakes of selenium, DHA and Vitamins B12 and D. J. Nutr. Health Aging 2008, 12, 186–192. [Google Scholar] [CrossRef]
- World Health Organization (WHO) Regional Office for Europe. Scientific Update on the Iodine Content of Portuguese Foods. 2018. Available online: https://www.euro.who.int/en/countries/portugal/publications/scientific-update-on-the-iodine-content-of-portuguese-foods-2018 (accessed on 25 March 2020).
- National Research Council (NRC). Nutrient Requirements of Fish and Shrimp; The National Academies Press: Washington, DC, USA, 2011.
- Coulson, E.J. The Iodine Content of Oysters; Bureau of Fisheries Investigational Report, 18; U.S., Department of Commerce: Washington, DC, USA, 1934; p. 10.
- Sprague, M.; Fawcett, S.; Betancor, M.B.; Struthers, W.; Tocher, D.R. Variation in the nutrient composition of farmed Atlantic salmon (Salmo salar L.) fillets available to UK consumers, with emphasis on EPA and DHA contents. J. Food Compost. Anal. 2020, 94, 103618. [Google Scholar] [CrossRef]
- Fordyce, F. Selenium deficiency and toxicity in the environment. In Essentials of Medical Geology: Impacts of the Natural Environment on Public Health, 1st ed.; Selinus, O., Ed.; Elsevier: New York, NY, USA, 2005; pp. 373–415. [Google Scholar]
- Whitehead, D.C. The distribution and transformations of iodine in the environment. Environ. Int. 1984, 10, 321–339. [Google Scholar] [CrossRef]
- Watanabe, T.; Kiron, V.; Satoh, S. Trace minerals in fish nutrition. Aquaculture 1997, 151, 185–207. [Google Scholar] [CrossRef]
- Crow, G.L. Goiter in elasmobranchs. In The Elasmobranch Husbandry Manual: Captive Care of Sharks, Rays and Their Relatives; Smith, M., Warmolts, D., Thoney, D., Hueter, R., Eds.; Ohio Biological Survey Inc.: Ohio, OH, USA, 2004; pp. 441–446. [Google Scholar]
- Julshamn, J.; Maage, A.; Waagbø, R.; Lundeby, A.-K. A preliminary study on tailoring of fillet iodine concentrations in adult Atlantic salmon (Salmo salar L.) through dietary supplementation. Aquac. Nutr. 2006, 12, 45–51. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Rema, P.; Ferraro, V.; Pintado, M.; Sousa-Pinto, I.; Cunha, L.M.; Oliveira, M.B.; Araújo, M. Iodine enrichment of rainbow trout flesh by dietary supplementation with the red seaweed Gracilaria Vermiculophylla. Aquaculture 2015, 446, 132–139. [Google Scholar] [CrossRef]
- Seafish. Aquaculture Profiles. Available online: https://www.seafish.org/responsible-sourcing/aquaculture-farming-seafood/species-farmed-in-aquaculture/aquaculture-profiles/ (accessed on 5 November 2021).
- EUMOFA The EU Market Overview, Species Profiles. Available online: https://www.eumofa.eu/the-eu-market (accessed on 5 November 2021).
- Seafish. SR694 Aquaculture in England, Wales and Northern Ireland: An Analysis of the Economic Contribution and Value of the Major Sub-Sectors and the Most Important Farmed Species. Available online: https://www.seafish.org/document?id=4382b7aa-ffce-448b-850d-46a8f7959115 (accessed on 5 November 2021).
- Calder, P. Very long-chain n-3 fatty acids and human health: Fact, Fiction and the future. Proc. Nutr. Soc. USA 2018, 77, 52–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Department for Environment, Food & Rural Affairs (DEFRA). Defra Family Food Datasets. Available online: https://www.gov.uk/government/statistical-data-sets/family-food-datasets (accessed on 22 April 2020).
- EUMOFA. The EU Fish Market (2020 Edition). Available online: https://www.eumofa.eu/documents/20178/415635/EN_The+EU+fish+market_2020.pdf/fe6285bb-5bb-5446-ac1a-e213-6fd6f64d0d85?t=1604671147068 (accessed on 5 November 2021).
- Seafish. Chilled Seafood in Multiple Retail (2020). A Market Insight Analysis. Available online: https://www.seafish.org/document/?id=19b3d61f-04ef-481e-affb-2abcda67dff0 (accessed on 5 November 2021).
- Bouga, M.; Lean, M.E.J.; Combet, E. Fish and seafood products as a source of iodine in the UK and reasons for consumers’ low intake. Proc. Nutr. Soc. USA 2018, 77, E178. [Google Scholar] [CrossRef]
- Govzman, S.; Looby, S.; Wang, X.; Butler, F.; Gibney, E.R.; Timon, C.M. A systematic review of the determinants of seafood consumption. Br. J. Nutr. 2021, 126, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Public Health England. Official Statistics—NDNS: Time Trend and Income Analyses for Years 1 to 9. Available online: https://www.gov.uk/government/statistics/ndns-time-trend-and-income-analyses-for-years-1-to-9 (accessed on 27 April 2021).
- Department of Health (DH). Dietary Reference Values, a Guide. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/743790/Dietary_Reference_Values_-_A_Guide__1991_.pdf (accessed on 6 April 2021).
- Gunnarsdottir, I.; Gunnarsdottir, B.E.; Steingrimsdottir, L.; Maage, A.; Johannesson, A.J.; Thorsdottir, I. Iodine status of adolescent girls in a population changing from high to lower fish consumption. Eur. J. Clin. Nutr. 2010, 64, 958–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, M.B.; Andersson, M. Assessment of iodine nutrition in populations: Past, present, and future. Nutr. Rev. 2012, 70, 553–570. [Google Scholar] [CrossRef]
- Øyen, J.; Aadland, E.K.; Liaset, B.; Fjære, E.; Dahl, L.; Madsen, L. Lean-seafood intake increases urinary iodine concentrations and plasma selenium levels: A randomized controlled trial with crossover design. Eur. J. Nutr. 2021, 60, 1679–1689. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Common Name 1 | Scientific Name | Farmed/Wild | Location 2 | N |
|---|---|---|---|---|
| FRESHWATER FISH | ||||
| Arctic char | Salvelinus alpinus | Farmed | UK | 4 |
| Common carp | Cyprinus carpio | Farmed | Poland | 5 |
| Milkfish | Chanos chanos | Farmed | Indonesia | 4 |
| Nile tilapia | Oreochromis niloticus | Farmed | China | 4 |
| Rainbow trout | Oncorhynchus mykiss | Farmed | UK | 5 |
| Siberian sturgeon | Acipenser baerii | Farmed | France | 4 |
| Striped catfish (Basa Pangasius) | Pangasius hypophthalmus | Farmed | Vietnam | 5 |
| Zander (pike-perch) | Sander lucioperca | Wild | Kazakhstan | 4 |
| MARINE FISH | ||||
| Anguilliformes | ||||
| Daggertooth pike-conger | Muraenesx cinerus | Wild | NE Atlantic (FAO 27) | 4 |
| European conger | Conger conger | Wild | NE Atlantic (FAO 27) | 4 |
| Clupeiformes | ||||
| Atlantic herring | Clupea harengus | Wild | NE Atlantic (FAO 27 IV, VII) | 7 |
| European anchovy | Engraulis encrasicolus | Wild | W. Mediterranean (FAO 37.1) | 3 |
| European pilchard (sardine) | Sardina pilchardus | Wild | NE Atlantic (FAO 27 IV, VII) | 5 |
| European sprat | Sprattus sprattus | Wild | NE Atlantic (FAO 27 VII) | 6 |
| Gadiformes | ||||
| Atlantic cod | Gadus morhua | Wild | NE Atlantic (FAO 27 I, II, IV, V) | 6 |
| Alaskan pollock | Theragra chalcogramma | Wild | NE Pacific (FAO 67) NW Pacific (FAO 61) | 3 |
| Cape hake | Merluccius capensis | Wild | SE Atlantic (FAO 47) | 4 |
| European hake | Merluccius merluccius | Wild | NE Atlantic (FAO 27 IV, VII) | 4 |
| Haddock | Melanogrammus aeglefinus | Wild | NE Atlantic (FAO 27 I, II, IV, V, VII) | 5 |
| Ling | Molva molva | Wild | NE Atlantic (FAO 27) | 5 |
| Pollack (Atlantic pollock) | Pollachius pollachius | Wild | NE Atlantic (FAO 27 IV) | 3 |
| Pouting | Trisopterus luscus | Wild | NE Atlantic (FAO 27 IV, VII) | 3 |
| Saithe (Coley) | Pollachius virens | Wild | NE Atlantic (FAO 27 IV, VII) | 4 |
| Whiting | Merlangius merlangus | Wild | NE Atlantic (FAO 27 IV, VII) | 5 |
| Lophiformes | ||||
| Monkfish | Lophius piscatorius | Wild | N.E. Atlantic (FAO 27) | 4 |
| Mugiliformes | ||||
| Flathead grey mullet | Mugil cephalus | Wild | N.E. Atlantic (FAO 27) | 5 |
| Thicklip grey mullet | Chelon labrosus | Wild | N.E. Atlantic (FAO 27) | 4 |
| Percoideri | ||||
| Atlantic horse mackerel | Trachurus trachurus | Wild | NE Atlantic (FAO 27 IX) | 3 |
| Barramundi (Asian seabass) | Lates calcarifer | Farmed | Vietnam | 6 |
| Black seabream | Spondyliosoma cantharus | Wild | NE Atlantic (FAO 27) | 6 |
| European seabass | Dicentrachus labrax | Wild | NE Atlantic (FAO 27) | 4 |
| Farmed | Greece, Turkey | 6 | ||
| Gilthead seabream | Sparus aurata | Farmed | Greece, Turkey | 6 |
| Meagre | Argyrosomus regius | Farmed | Greece, Turkey | 4 |
| Red mullet (Indian goatfish) | Parupeneus indicus, P. heptacanthus | Wild | W Indian (FAO 51), E Indian (FAO 57) | 8 |
| Red snapper | Lutjanus malabaricus, L. sebae, Pinjalo pinjalo | Wild | W Central Pacific (FAO 71) | 4 |
| White trevally (Trevally jack) | Pseudocaranx dentex | Wild | SW Pacific (FAO 81) | 3 |
| Yellow croaker | Larimichthys polyactis | Wild | NW Pacific (FAO 61) | 3 |
| Pleuronectiformes | ||||
| Atlantic halibut | Hippoglossus hipposglossus | Wild | NE Atlantic (FAO 27) | 6 |
| Farmed | Norway | 6 | ||
| Brill | Scophthalmus rhombus | Wild | NE Atlantic (FAO 27) | 5 |
| Common dab | Limanda limanda | Wild | NE Atlantic (FAO 27 IV, VII) | 4 |
| Common sole (Dover sole) | Solea solea | Wild | NE Atlantic (FAO 27 IV, VII) | 6 |
| European flounder | Platichys flesus | Wild | NE Atlantic (FAO 27) | 4 |
| European plaice | Pleuronectes platessa | Wild | NE Atlantic (FAO 27 IV, V, VII) | 6 |
| Lemon sole | Microstomus kitt | Wild | NE Atlantic (FAO 27 IV, V, VII) | 7 |
| Megrim | Lepidorhombus whiffiagonis | Wild | NE Atlantic (FAO 27 IV, VII) | 4 |
| Turbot | Psetta maxima (Scophthalmus maximus) | Wild | NE Atlantic (FAO 27) | 5 |
| Witch flounder (Witch sole) | Glyptocephalus cynoglossus | Wild | NE Atlantic (FAO 27) | 5 |
| Yellowfin sole | Limanda aspera | Wild | NE Pacific Ocean (FAO 67) | 3 |
| MARINE FISH | ||||
| Rajiformes | ||||
| Ray wings | Leucoraja naevus, Raja montagui, R. clavata | Wild | NE Atlantic (FAO 27 II, IV, VI, VII) | 4 |
| Salmoniformes | ||||
| Atlantic salmon | Salmo salar | Wild | UK, Norway | 6 |
| Farmed | UK, Norway | 32 | ||
| Keta salmon (Chum) | Oncorhynchus keta | Wild | NE Pacific (FAO 67) | 5 |
| Pink salmon (Humpback) | Oncorhynchus gorbuscha | Wild | NE Pacific (FAO 67) | 4 |
| Rainbow trout | Oncorhynchus mykiss | Farmed | UK | 5 |
| Sea trout | Salmo trutta | Wild | NE Atlantic (FAO 27 IV) | 4 |
| Sockeye salmon (Red) | Oncorhynchus nerka | Wild | NE Pacific (FAO 67), NW Pacific (FAO 61) | 6 |
| Scombroidei | ||||
| Albacore tuna | Thunnus alalunga | Wild | W Indian (FAO 51) E Indian (FAO 57), W Central Pacific (FAO 71) E Central Pacific (FAO 77) W Central Atlantic (FAO 31) | 5 |
| Atlantic mackerel | Scomber scombrus | Wild | NE Atlantic (FAO 27 IV, VI, VII, VIII) | 7 |
| Black marlin | Makaira indica | Wild | W Central Pacific (FAO 71) | 5 |
| Largehead hairtail (Beltfish) | Trichiurus lepturus | Wild | W Central Pacific (FAO 71) | 3 |
| Savalai hairtail (Ribbonfish) | Lepturacanthus savala | Wild | W Indian (FAO 51) | 3 |
| Swordfish | Xiphias gladius | Wild | E Indian (FAO 57) W Indian (FAO 51) E Central Pacific (FAO 77) W Central Pacific (FAO 71) | 5 |
| Wahoo (Kingfish) | Acanthocybium solandri | Wild | E Indian (FAO 57) W Indian (FAO 51) | 4 |
| Yellowfin tuna | Thunnus albacares | Wild | E Indian (FAO 57) W Indian (FAO 51) W Central Pacific (FAO 71) E Central Pacific (FAO 77) SE Pacific (FAO 87) SW Pacific (FAO 81) | 6 |
| Scorpaeniformes | ||||
| Gurnard | Chelidonichthys lucerna, C. spinosus, Eutrigla gurnadus | Wild | NE Atlantic (FAO 27 IV, VII) | 8 |
| Redfish (Norway Redfish) | Sebastes spp. | Wild | NE Atlantic (FAO 27 V) | 4 |
| Stromateoidei, Anabantoidei | ||||
| Silver pomfret | Pampus argentus | Wild | W Indian (FAO 51) | 4 |
| Squaliformes | ||||
| Picked dogfish (Huss) | Squalus acanthias | Wild | NW Atlantic (FAO 21) | 3 |
| Zeiformes | ||||
| John dory | Zeus faber | Wild | NE Atlantic (FAO 27) | 7 |
| Oreo dory (Smooth oreo) | Pseudocyttus maculatus | Wild | SW Pacific (FAO 81) | 5 |
| Zoarcoidei | ||||
| Atlantic wolfish | Anarhichas lupus | Wild | NE Atlantic (FAO 27) | 4 |
| Scombroidei | ||||
| Albacore tuna | Thunnus alalunga | Wild | W Indian (FAO 51) E Indian (FAO 57), W Central Pacific (FAO 71) E Central Pacific (FAO 77) W Central Atlantic (FAO 31) | 5 |
| Atlantic mackerel | Scomber scombrus | Wild | NE Atlantic (FAO 27 IV, VI, VII, VIII) | 7 |
| Black marlin | Makaira indica | Wild | W Central Pacific (FAO 71) | 5 |
| SHELLFISH | ||||
| CRUSTACEANS | ||||
| American lobster | Homarus americanus | Wild | NW Atlantic (FAO 21) | 4 |
| * Edible crab—whole | Cancer pagrus | Wild | NE Atlantic (FAO 27 VII) | 3 |
| * Edible crab—brown meat | Wild | NE Atlantic (FAO 27 IV, VII) | 4 | |
| * Edible crab—white meat | Wild | NE Atlantic (FAO 27 VII) | 3 | |
| King prawn | Litopenaeus vannamei | Farmed | Vietnam, Ecuador, Honduras | 4 |
| Langoustines (Norway lobster) | Nephrops norvegicus | Wild | NE Atlantic (FAO 27) | 4 |
| Tiger Prawn | Penaeus monodon | Farmed | Vietnam | 4 |
| MOLLUSCS | ||||
| Bivalves | ||||
| Atlantic (deep-sea) scallop | Placopecten magellanicus | Wild | NW Atlantic (FAO 21) | 3 |
| Atlantic razor clam | Ensis directus | Wild | NE Atlantic (FAO 27 IV??) | 5 |
| * Blue mussels | Mytilus edulis | Wild | NE Atlantic (FAO 27) | 4 |
| Blue mussels | Farmed | UK | 4 | |
| * Chilean mussels | Mytilus chilensis | Farmed | Chile | 4 |
| SHELLFISH | ||||
| MOLLUSCS | ||||
| Bivalves. | ||||
| * Common edible cockle | Cerastoderma edule | Wild | NE Atlantic (FAO 27 IV) | 4 |
| Green-lipped mussels | Perna canaliculus | Farmed | New Zealand | 4 |
| King scallop + roe | Pecten maximus | Wild | NE Atlantic (FAO 27) | 3 |
| Pacific oyster | Crassostrea gigas | Farmed | UK, Ireland | 3 |
| Patagonian scallop | Zygochlamys patagonica | Wild | SW Atlantic (FAO 41) | 3 |
| Cephalopods | ||||
| Pharaoh cuttlefish | Sepia pharaonis | Wild | W Indian (FAO 51) | 4 |
| Horned octopus (curled) | Eledone cirrhosa | Wild | NE Atlantic (FAO 27 IV, VII) | 5 |
| Squid | Loligo vulgaris, L. forbesi | Wild | W Central Pacific (FAO 71) E Indian (FAO 57) W Indian (Area 51) | 4 |
| Common Name 1 | n | Iodine Content (µg·100 g−1 Flesh ww) | ||||
|---|---|---|---|---|---|---|
| Mean | Geo-Mean | Median | Min. | Max. | ||
| FRESHWATER FISH | 35 | 8.27 | 6.52 | 6.40 | 1.13 | 26.46 |
| MARINE FISH | 342 | 31.17 | 19.97 | 17.31 | 3.49 | 909.77 |
| Anguilliformes | 8 | 21.90 | 20.79 | 20.44 | 13.60 | 41.53 |
| Clupeiformes | 21 | 26.55 | 23.59 | 23.72 | 10.83 | 69.70 |
| Gadiformes | 42 | 100.72 | 54.69 | 65.14 | 5.97 | 909.77 |
| Lophiformes | 4 | 33.35 | 31.90 | 33.56 | 21.95 | 41.44 |
| Mugiliformes | 9 | 35.08 | 18.29 | 13.81 | 7.18 | 197.26 |
| Percoideri | 53 | 21.48 | 17.70 | 16.11 | 5.94 | 95.78 |
| Pleuronectiformes | 66 | 27.06 | 22.91 | 23.48 | 7.07 | 103.01 |
| Rajiformes | 4 | 16.63 | 16.11 | 16.62 | 10.88 | 22.42 |
| Salmoniformes | 62 | 14.51 | 13.14 | 13.64 | 4.54 | 34.39 |
| Scombroidei | 38 | 17.48 | 14.43 | 13.82 | 6.04 | 58.50 |
| Scorpaeniformes | 12 | 24.07 | 20.52 | 19.41 | 8.42 | 66.43 |
| Stromateoidei, Anabantoidei | 4 | 5.35 | 5.28 | 5.40 | 4.12 | 6.48 |
| Squaliformes | 3 | 26.29 | 25.25 | 30.45 | 16.65 | 31.76 |
| Zeiformes | 12 | 12.52 | 10.43 | 13.61 | 3.49 | 27.81 |
| Zoarcoidei | 4 | 38.34 | 28.78 | 36.95 | 12.75 | 66.71 |
| SHELLFISH | 79 | 86.09 | 39.33 | 58.41 | 3.81 | 440.18 |
| Crustaceans | 26 | 139.98 | 60.26 | 96.36 | 3.81 | 440.18 |
| Molluscs | 53 | 59.66 | 31.90 | 49.37 | 3.92 | 241.45 |
| Bivalves | 40 | 75.71 | 48.29 | 77.77 | 5.59 | 241.45 |
| Cephalopods | 13 | 10.28 | 8.90 | 6.62 | 3.92 | 19.97 |
| Common Name 1 | n | Iodine Content (µg·100 g−1 Flesh ww) | ||||
|---|---|---|---|---|---|---|
| Mean | Geo-Mean | Median | Min. | Max. | ||
| WILD (All) | 332 | 44.89 | 24.07 | 19.55 | 3.49 | 909.77 |
| FARMED (All) | 124 | 22.97 | 13.61 | 11.72 | 1.13 | 169.64 |
| FRESHWATER FISH | ||||||
| Wild | 4 | 5.77 | 5.70 | 5.43 | 5.03 | 7.18 |
| Farmed | 31 | 8.59 | 6.63 | 6.84 | 1.13 | 26.46 |
| MARINE | ||||||
| Wild (All) | 328 | 45.36 | 24.49 | 19.70 | 3.49 | 909.77 |
| Farmed (All) | 93 | 27.77 | 17.30 | 14.19 | 3.81 | 169.64 |
| MARINE FISH | ||||||
| Wild | 272 | 35.08 | 22.03 | 18.86 | 3.49 | 909.77 |
| Farmed | 70 | 15.97 | 13.65 | 13.20 | 4.54 | 49.61 |
| SHELLFISH | ||||||
| Wild (All) | 56 | 95.31 | 40.99 | 51.49 | 3.92 | 440.18 |
| Farmed (All) | 23 | 63.66 | 35.55 | 59.13 | 3.81 | 169.64 |
| Crustaceans | ||||||
| Wild | 18 | 199.13 | 162.66 | 197.92 | 41.87 | 440.18 |
| Farmed | 8 | 6.89 | 6.51 | 6.73 | 3.81 | 11.13 |
| Molluscs | ||||||
| Wild (All) | 38 | 46.13 | 21.37 | 16.05 | 3.92 | 241.45 |
| Farmed (All) | 15 | 93.94 | 87.95 | 84.12 | 49.37 | 169.64 |
| Bivalves | ||||||
| Wild | 25 | 64.77 | 33.70 | 43.94 | 5.59 | 241.45 |
| Farmed | 15 | 93.94 | 87.94 | 84.12 | 49.37 | 169.64 |
| Cephalopods * | 13 | 10.28 | 8.90 | 6.62 | 3.92 | 19.97 |
| Fish Intake (g·Day−1) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Age Range (Years) | Males and Females | Males | Females | |||||||||
| 1–5 | 6–10 | 11–18 | 19–64 | 1–5 | 6–10 | 11–18 | 19–64 | 1–5 | 6–10 | 11–18 | 19–64 | |
| Seafood type | ||||||||||||
| Salmon | 2.0 | 2.6 | 4.4 | 4.2 | 1.9 | 2.8 | 4.4 | 4.3 | 2.0 | 2.3 | 4.4 | 4.2 |
| Other Oily | 1.0 | 1.4 | 1.5 | 3.5 | 1.1 | 1.5 | 1.5 | 3.9 | 0.8 | 1.3 | 1.5 | 3.2 |
| Total Oily | 3.0 | 4.0 | 5.9 | 7.7 | 3.0 | 4.3 | 5.9 | 8.2 | 2.8 | 3.6 | 5.9 | 7.4 |
| Other | 12.0 | 10.5 | 6.3 | 3.3 | 12.9 | 11.3 | 6.9 | 3.5 | 10.9 | 9.5 | 5.7 | 3.1 |
| Tuna | 3.1 | 5.1 | 9.5 | 4.8 | 2.7 | 4.9 | 9.4 | 5.4 | 3.6 | 5.3 | 9.6 | 4.3 |
| Whitefish | 4.9 | 8.2 | 11.2 | 8.7 | 4.7 | 9.2 | 13.0 | 10.6 | 5.0 | 7.2 | 9.6 | 7.4 |
| Total Non-Oily | 20.0 | 23.8 | 27.0 | 16.8 | 20.3 | 25.4 | 29.3 | 19.5 | 19.5 | 22.0 | 24.9 | 14.8 |
| Shellfish | 0.7 | 1.4 | 3.3 | 3.2 | 0.8 | 1.8 | 3.2 | 3.2 | 0.6 | 0.9 | 3.4 | 3.3 |
| TOTAL | 23.7 | 29.2 | 36.2 | 27.7 | 24.1 | 31.5 | 38.4 | 30.9 | 22.9 | 26.5 | 34.2 | 25.5 |
| %RWI * | 59.3 | 73.0 | 90.5 | 69.3 | 60.3 | 78.8 | 96.0 | 77.3 | 57.3 | 66.3 | 85.5 | 63.8 |
| Estimated Iodine Intake (g·Day−1) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Age Range (Years) | Males and Females | Males | Females | |||||||||
| 1–5 | 6–10 | 11–18 | 19–64 | 1–5 | 6–10 | 11–18 | 19–64 | 1–5 | 6–10 | 11–18 | 19–64 | |
| Seafood type | ||||||||||||
| Salmon | 0.30 | 0.40 | 0.64 | 0.60 | 0.31 | 0.42 | 0.66 | 0.60 | 0.30 | 0.37 | 0.63 | 0.59 |
| Other Oily | 0.22 | 0.29 | 0.30 | 0.73 | 0.24 | 0.35 | 0.31 | 0.85 | 0.21 | 0.21 | 0.28 | 0.64 |
| Total Oily | 0.52 | 0.69 | 0.94 | 1.33 | 0.55 | 0.77 | 0.97 | 1.45 | 0.51 | 0.58 | 0.91 | 1.23 |
| Other | 5.67 | 4.88 | 2.76 | 1.27 | 6.11 | 5.25 | 3.23 | 1.36 | 5.18 | 4.46 | 2.30 | 1.21 |
| Tuna | 0.28 | 0.46 | 0.85 | 0.43 | 0.25 | 0.44 | 0.86 | 0.49 | 0.32 | 0.49 | 0.84 | 0.40 |
| Whitefish | 5.27 | 9.33 | 13.38 | 10.72 | 5.05 | 10.74 | 15.23 | 13.00 | 5.52 | 7.75 | 11.62 | 9.13 |
| Total Non-Oily | 11.22 | 14.67 | 16.99 | 12.42 | 11.41 | 16.43 | 19.32 | 14.85 | 11.02 | 12.70 | 14.76 | 10.74 |
| Shellfish | 0.48 | 0.56 | 1.23 | 1.18 | 0.61 | 0.65 | 1.18 | 1.21 | 0.33 | 0.44 | 1.28 | 1.16 |
| TOTAL | 12.2 | 15.9 | 19.2 | 14.9 | 12.6 | 17.8 | 21.5 | 17.5 | 11.8 | 13.7 | 16.9 | 13.1 |
| %RNI * | 12.2–17.4 | 14.5–15.9 | 13.7–14.8 | 10.6 | 12.6–18.0 | 16.2–17.8 | 15.4–16.5 | 12.5 | 11.8–16.9 | 12.5–13.7 | 12.1–13.0 | 9.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sprague, M.; Chau, T.C.; Givens, D.I. Iodine Content of Wild and Farmed Seafood and Its Estimated Contribution to UK Dietary Iodine Intake. Nutrients 2022, 14, 195. https://doi.org/10.3390/nu14010195
Sprague M, Chau TC, Givens DI. Iodine Content of Wild and Farmed Seafood and Its Estimated Contribution to UK Dietary Iodine Intake. Nutrients. 2022; 14(1):195. https://doi.org/10.3390/nu14010195
Chicago/Turabian StyleSprague, Matthew, Tsz Chong Chau, and David I. Givens. 2022. "Iodine Content of Wild and Farmed Seafood and Its Estimated Contribution to UK Dietary Iodine Intake" Nutrients 14, no. 1: 195. https://doi.org/10.3390/nu14010195
APA StyleSprague, M., Chau, T. C., & Givens, D. I. (2022). Iodine Content of Wild and Farmed Seafood and Its Estimated Contribution to UK Dietary Iodine Intake. Nutrients, 14(1), 195. https://doi.org/10.3390/nu14010195