
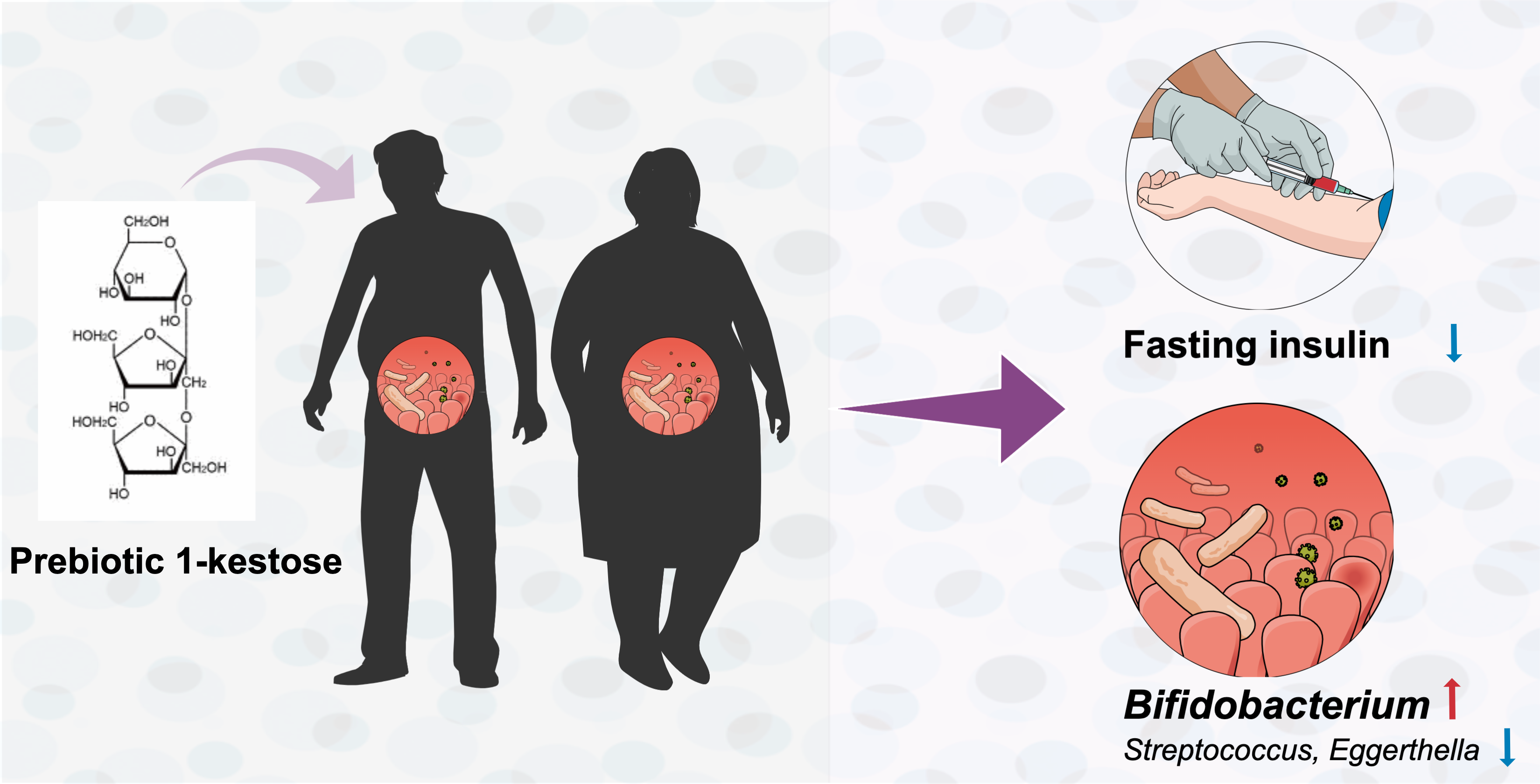
Supplementation of 1-Kestose Modulates the Gut Microbiota Composition to Ameliorate Glucose Metabolism in Obesity-Prone Hosts

 , , and
, , and
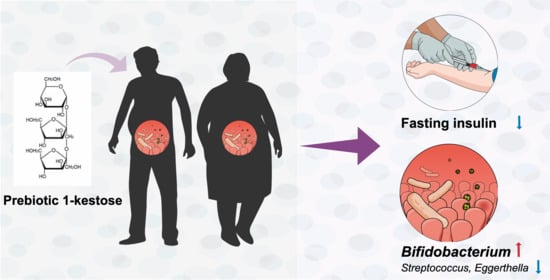
Abstract
:
1. Introduction
2. Materials and methods
2.1. Rodent Studies
2.1.1. Animals, Diets, and Experimental Design
2.1.2. Analyses of Blood Components
2.1.3. Statistical Analysis
2.2. Clinical Trial
2.2.1. Study Design
2.2.2. Participants
2.2.3. Outcomes
2.2.4. Study Protocol
2.2.5. Anthropometric Characteristics
2.2.6. OGTT
2.2.7. Serum Analysis
2.2.8. Gut Microbiota Composition
2.2.9. Statistical Analysis
3. Results
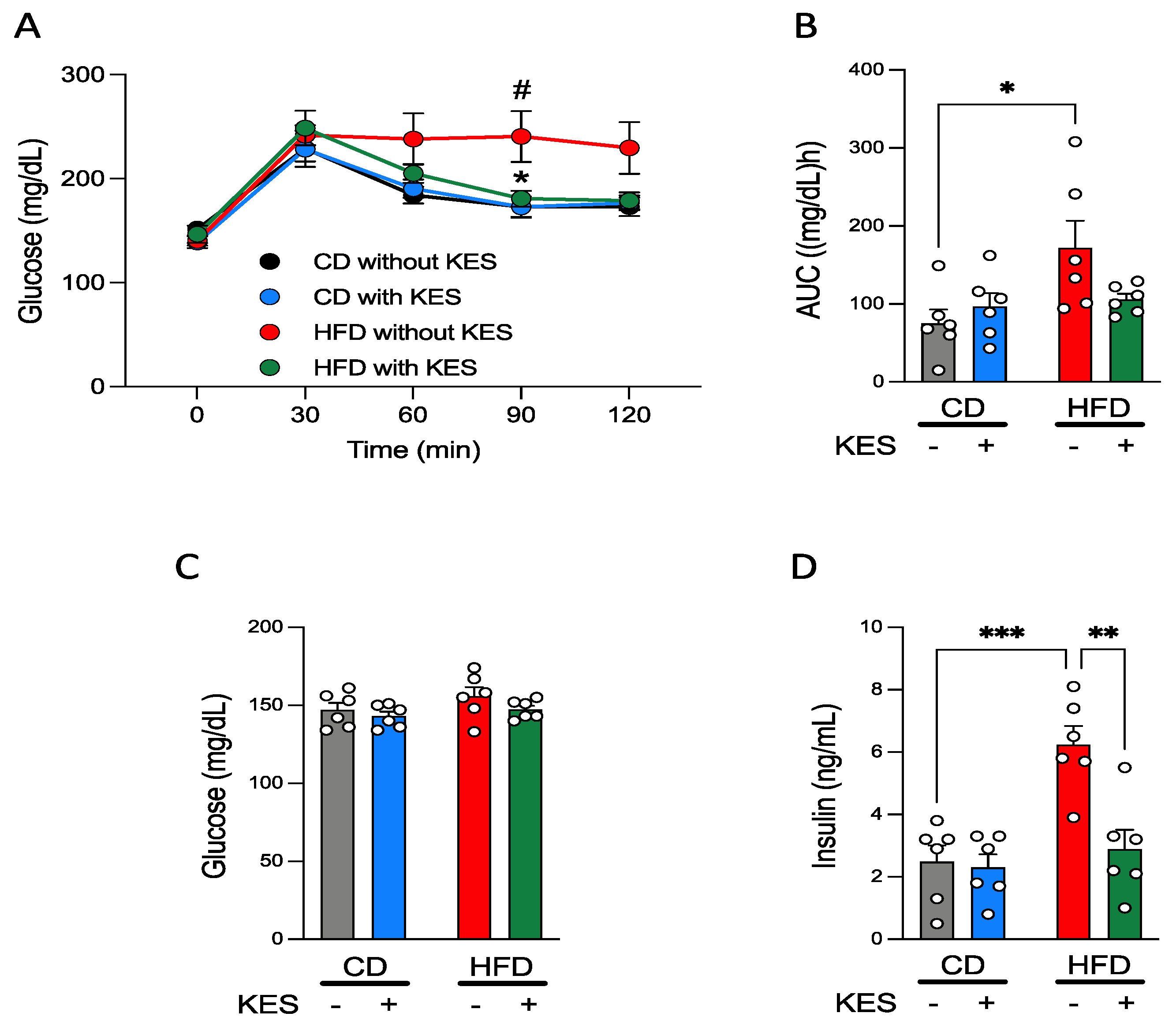
3.1. Animal Study
3.1.1. A High-Fat Diet Induces Obesity in Rats
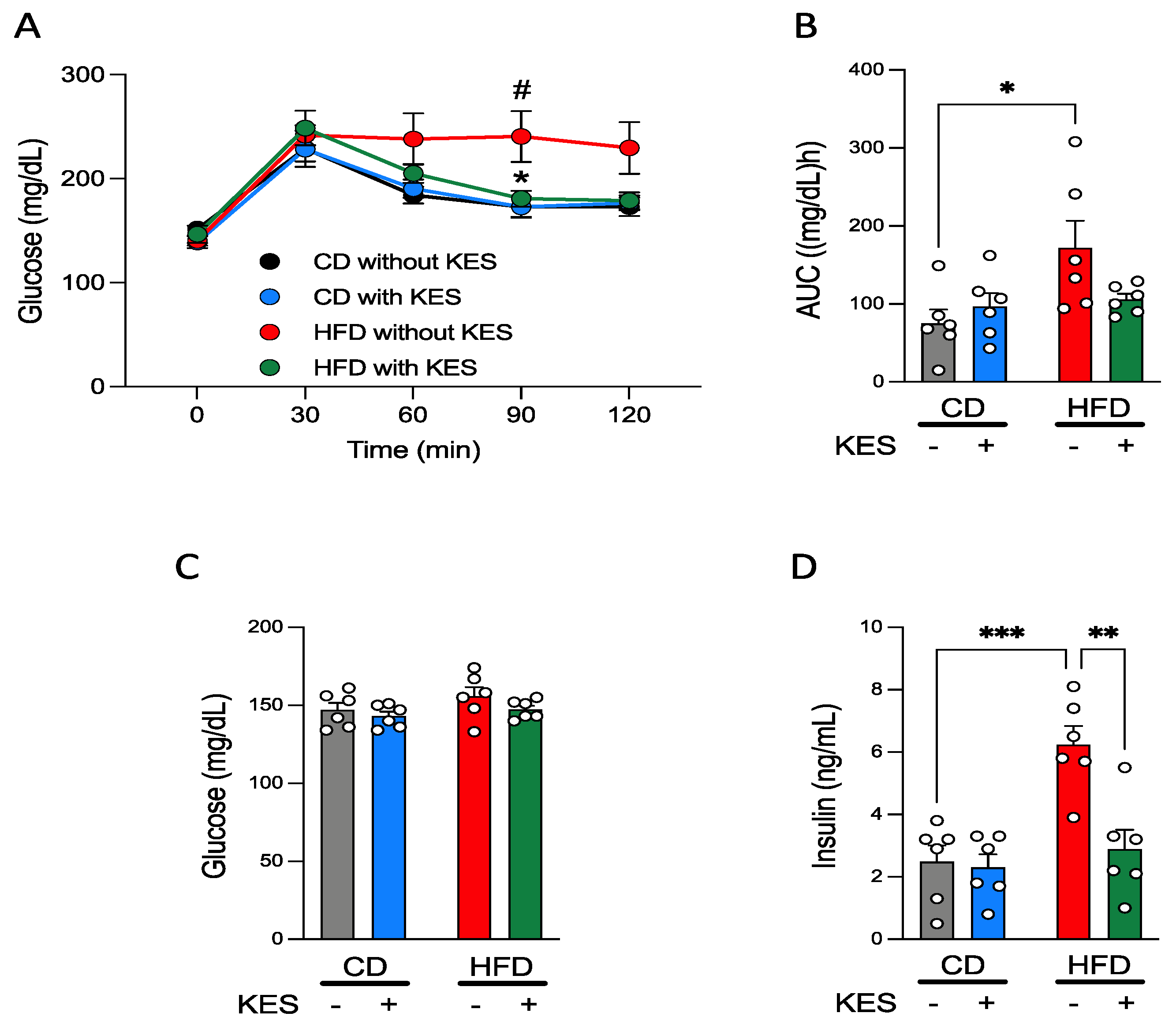
3.1.2. Supplementation of 1-Kestose Improves Glucose Tolerance in Rats
3.1.3. Concentrations of Plasma Glucose and Insulin under Fasting and Fed States in Rats
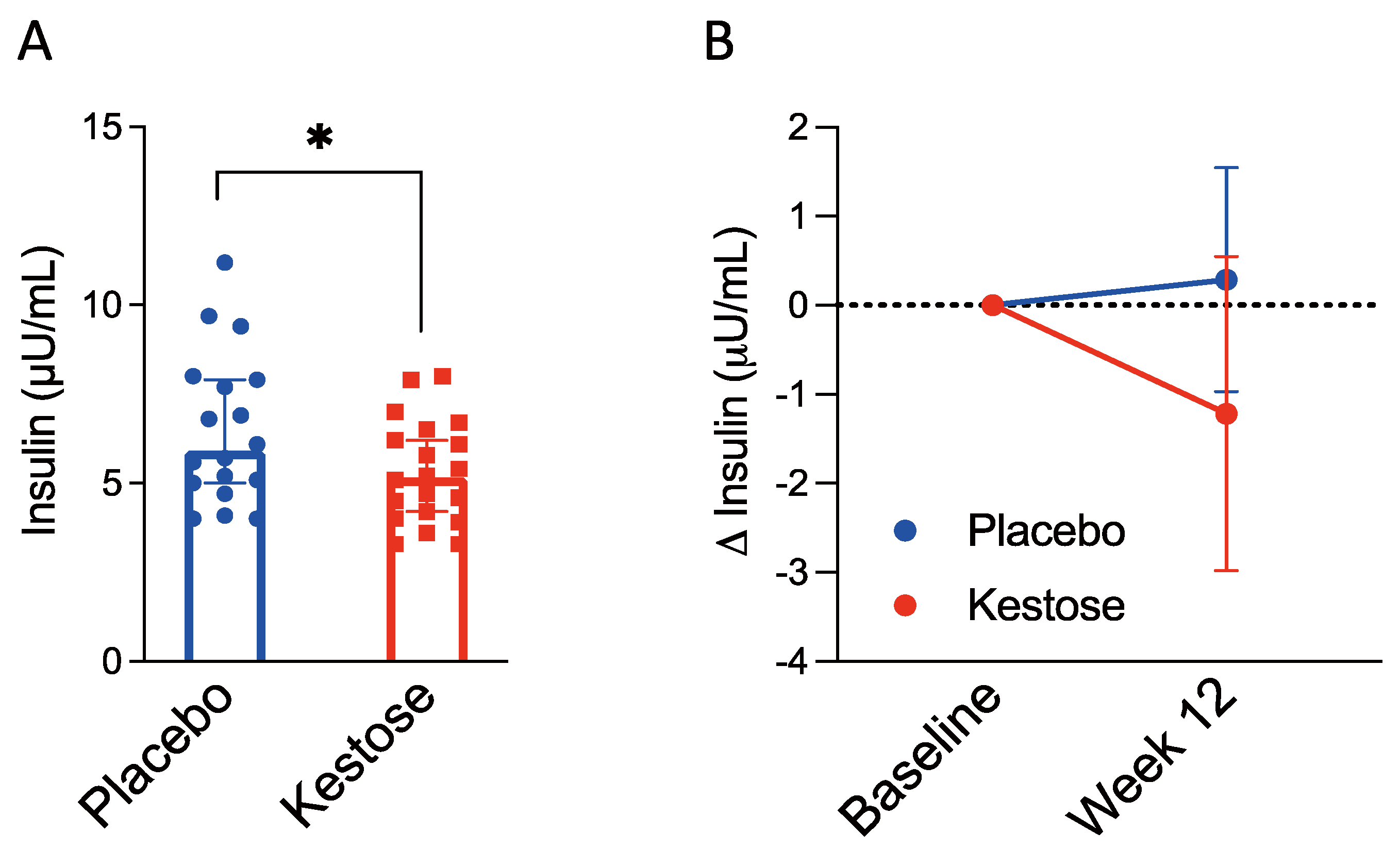
3.2. Interventional Human Study
3.2.1. Characteristics of Participants
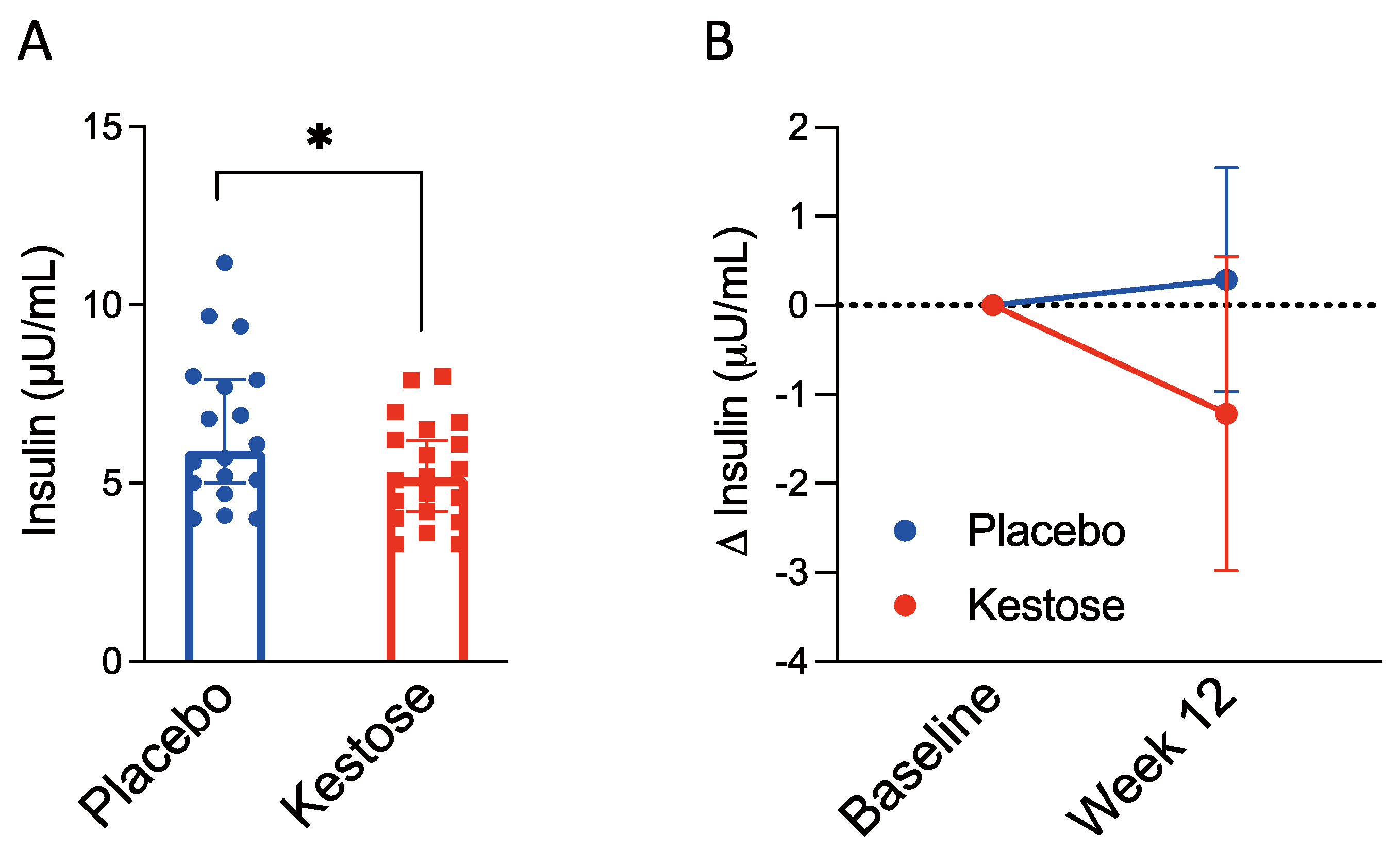
3.2.2. Serum Insulin Concentration
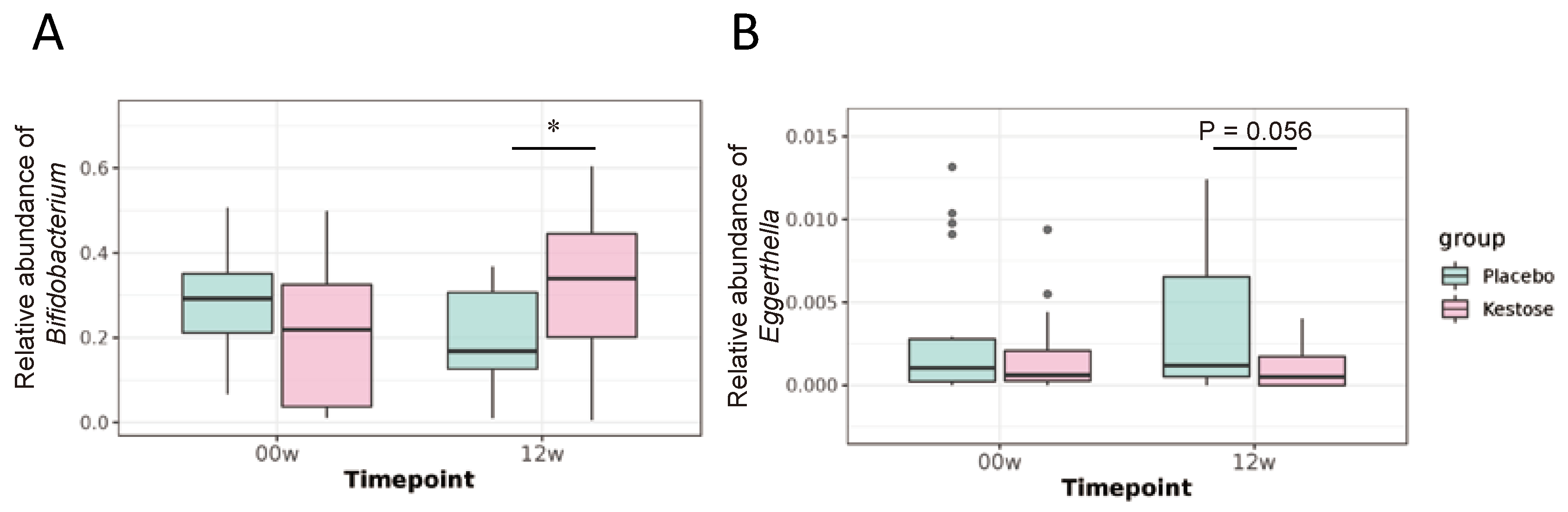
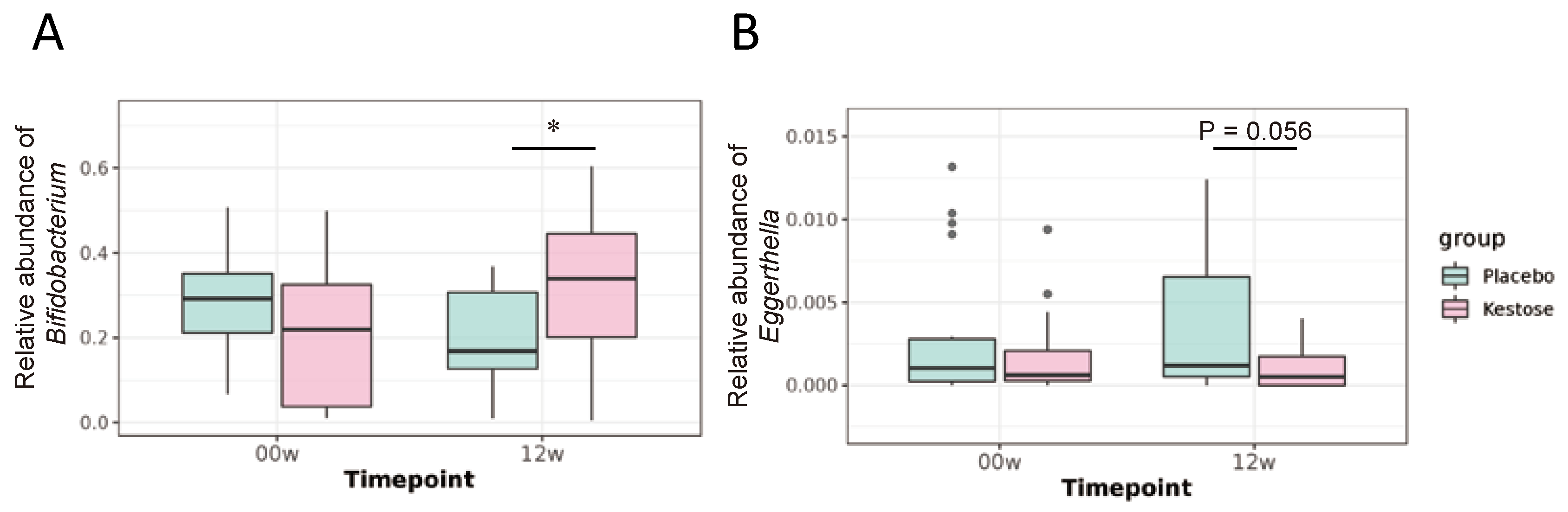
3.2.3. Supplementation of 1-Kestose Alters Gut Microbial Composition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sasaki, H.; Lyu, Y.; Nakayama, Y.; Nakamura, F.; Watanabe, A.; Miyakawa, H.; Nakao, Y.; Shibata, S. Combinatorial Effects of Soluble, Insoluble, and Organic Extracts from Jerusalem Artichokes on Gut Microbiota in Mice. Microorganisms 2020, 8, 954. [Google Scholar] [CrossRef]
- Wang, Q.; Jokelainen, J.; Auvinen, J.; Puukka, K.; Keinänen-Kiukaanniemi, S.; Järvelin, M.-R.; Kettunen, J.; Mäkinen, V.-P.; Ala-Korpela, M. Insulin resistance and systemic metabolic changes in oral glucose tolerance test in 5340 individuals: An interventional study. BMC Med. 2019, 17, 217. [Google Scholar] [CrossRef]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.R.; Delzenne, N.M. Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 2007, 50, 2374–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhang, M.; Wang, S.; Han, R.; Cao, Y.; Hua, W.; Mao, Y.; Zhang, X.; Pang, X.; Wei, C.; et al. Interactions between gut microbiota, host genetics and diet relevant to development of metabolic syndromes in mice. ISME J. 2010, 4, 232–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.M.; Olefsky, J.M. The origins and drivers of insulin resistance. Cell 2013, 152, 673–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, A.; Molinaro, A.; Ståhlman, M.; Khan, M.T.; Schmidt, C.; Mannerås-Holm, L.; Wu, H.; Carreras, A.; Jeong, H.; Olofsson, L.E.; et al. Microbially Produced Imidazole Propionate Impairs Insulin Signaling through mTORC1. Cell 2018, 175, 947–961.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menni, C.; Zhu, J.; Le Roy, C.I.; Mompeo, O.; Young, K.; Rebholz, C.M.; Selvin, E.; North, K.E.; Mohney, R.P.; Bell, J.T.; et al. Serum metabolites reflecting gut microbiome alpha diversity predict type 2 diabetes. Gut Microbes 2020, 11, 1632–1642. [Google Scholar] [CrossRef]
- Koh, A.; Mannerås-Holm, L.; Yunn, N.-O.; Nilsson, P.M.; Ryu, S.H.; Molinaro, A.; Perkins, R.; Smith, J.G.; Bäckhed, F. Microbial Imidazole Propionate Affects Responses to Metformin through p38γ-Dependent Inhibitory AMPK Phosphorylation. Cell Metab. 2020, 32, 643–653.e4. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, M.A.; Hoffmann, C.; Sherrill–Mix, S.A.; Keilbaugh, S.A.; Hamady, M.; Chen, Y.Y.; Knight, R.; Ahima, R.S.; Bushman, F.; Wu, G.D. High-Fat Diet Determines the Composition of the Murine Gut Microbiome Independently of Obesity. Gastroenterology 2009, 137, 1716–1724.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delzenne, N.M.; Neyrinck, A.M.; Bäckhed, F.; Cani, P.D. Targeting gut microbiota in obesity: Effects of prebiotics and probiotics. Nat. Rev. Endocrinol. 2011, 7, 639–646. [Google Scholar] [CrossRef]
- Delzenne, N.M.; Olivares, M.; Neyrinck, A.M.; Beaumont, M.; Kjølbæk, L.; Larsen, T.M.; Benítez-Páez, A.; Romaní-Pérez, M.; Garcia-Campayo, V.; Bosscher, D.; et al. Nutritional interest of dietary fiber and prebiotics in obesity: Lessons from the MyNewGut consortium. Clin. Nutr. 2020, 39, 414–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Q.; Chen, H.; Hu, J.; Tan, H.; Nie, S.; Xie, M. Effects of Nondigestible Oligosaccharides on Obesity. Annu. Rev. Food Sci. Technol. 2020, 11, 205–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorbani, Y.; Schwenger, K.J.P.; Allard, J.P. Manipulation of intestinal microbiome as potential treatment for insulin resistance and type 2 diabetes. Eur. J. Nutr. 2021, 60, 2361–2379. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tang, H.; Zhang, C.; Zhao, Y.; Derrien, M.; Rocher, E.; Van-Hylckama Vlieg, J.E.; Strissel, K.; Zhao, L.; Obin, M.; et al. Modulation of gut microbiota during probiotic-mediated attenuation of metabolic syndrome in high fat diet-fed mice. ISME J. 2015, 9, 1–15. [Google Scholar] [CrossRef]
- Kobyliak, N.; Falalyeyeva, T.; Mykhalchyshyn, G.; Kyriienko, D.; Komissarenko, I. Effect of alive probiotic on insulin resistance in type 2 diabetes patients: Randomized clinical trial. Diabetes Metab. Syndr. Clin. Res. Rev. 2018, 12, 617–624. [Google Scholar] [CrossRef]
- Zhang, C.; Yin, A.; Li, H.; Wang, R.; Wu, G.; Shen, J.; Zhang, M.; Wang, L.; Hou, Y.; Ouyang, H.; et al. Dietary Modulation of Gut Microbiota Contributes to Alleviation of Both Genetic and Simple Obesity in Children. EBioMedicine 2015, 2, 968–984. [Google Scholar] [CrossRef]
- Hiel, S.; Gianfrancesco, M.A.; Rodriguez, J.; Portheault, D.; Leyrolle, Q.; Bindels, L.B.; Gomes Da Silveira Cauduro, C.; Mulders, M.D.G.H.; Zamariola, G.; Azzi, A.-S.; et al. Link between gut microbiota and health outcomes in inulin-treated obese patients: Lessons from the Food4Gut multicenter randomized placebo-controlled trial. Clin. Nutr. 2020, 39, 3618–3628. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Vahedi, K.; Achour, L.; Attar, A.; Salfati, J.; Pochart, P.; Marteau, P.; Flourie, B.; Bornet, F.; Rambaud, J.C. Short-chain fructo-oligosaccharide administration dose-dependently increases fecal bifidobacteria in healthy humans. J. Nutr. 1999, 129, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Endo, A.; Nakamura, S.; Konishi, K.; Nakagawa, J.; Tochio, T. Variations in prebiotic oligosaccharide fermentation by intestinal lactic acid bacteria. Int. J. Food Sci. Nutr. 2016, 67, 125–132. [Google Scholar] [CrossRef]
- Tochio, T.; Kitaura, Y.; Nakamura, S.; Sugawa, C.; Takahashi, M.; Endo, A.; Shimomura, Y. An Alteration in the Cecal Microbiota Composition by Feeding of 1-Kestose Results in a Marked Increase in the Cecal Butyrate Content in Rats. PLoS ONE 2016, 11, e0166850. [Google Scholar] [CrossRef]
- Watanabe, A.; Kadota, Y.; Kamio, R.; Tochio, T.; Endo, A.; Shimomura, Y.; Kitaura, Y. 1-Kestose supplementation mitigates the progressive deterioration of glucose metabolism in type 2 diabetes OLETF rats. Sci. Rep. 2020, 10, 15674. [Google Scholar] [CrossRef]
- Banauch, D.; Brümmer, W.; Ebeling, W.; Metz, H.; Rindfrey, H.; Lang, H.; Leybold, K.; Rick, W.; Staudinger, H.J. A glucose dehydrogenase for the determination of glucose concentrations in body fluids (author’s transl). Z. Klin. Chem. Klin. Biochem. 1975, 13, 101–107. [Google Scholar] [PubMed]
- Sugo, S.; Matsumoto, Y.; Yamaoka, T.; Sakurabayashi, I. Improved enzymatic method for determining free fatty acids in serum, with use of 3-octenoic acid. Clin. Chem. 1990, 36, 163. [Google Scholar] [CrossRef] [PubMed]
- Shiwaku, K.; Anuurad, E.; Enkhmaa, B.; Nogi, A.; Kitajima, K.; Shimono, K.; Yamane, Y.; Oyunsuren, T. Overweight Japanese with body mass indexes of 23.0–24.9 have higher risks for obesity-associated disorders: A comparison of Japanese and Mongolians. Int. J. Obes. 2004, 28, 152–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, S.; Murakami, K.; Sasaki, S.; Okubo, H.; Hirota, N.; Notsu, A.; Fukui, M.; Date, C. Comparison of relative validity of food group intakes estimated by comprehensive and brief-type self-administered diet history questionnaires against 16 d dietary records in Japanese adults. Public Health Nutr. 2011, 14, 1200–1211. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- McGough, J.J.; Faraone, S.V. Estimating the size of treatment effects: Moving beyond p values. Psychiatry 2009, 6, 21–29. [Google Scholar]
- Borestein, M. Effect size of continuous data. In The Handbook of Research Synthesis and Meta—Analysis, 2nd ed.; Cooper, H., Hedges, L.V., Valentine, J.C., Eds.; Russell Sage Foundation: New York, NY, USA, 2009; pp. 221–235. [Google Scholar]
- Criter, T.E.C.O. New Criteria for `Obesity Disease’ in Japan. Circ. J. 2002, 66, 987–992. [Google Scholar]
- Cani, P.D.; Knauf, C.; Iglesias, M.A.; Drucker, D.J.; Delzenne, N.M.; Burcelin, R. Improvement of Glucose Tolerance and Hepatic Insulin Sensitivity by Oligofructose Requires a Functional Glucagon-Like Peptide 1 Receptor. Diabetes 2006, 55, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Boll, E.V.; Ekström, L.M.; Courtin, C.M.; Delcour, J.A.; Nilsson, A.C.; Björck, I.M.; Östman, E.M. Effects of wheat bran extract rich in arabinoxylan oligosaccharides and resistant starch on overnight glucose tolerance and markers of gut fermentation in healthy young adults. Eur. J. Nutr. 2016, 55, 1661–1670. [Google Scholar] [CrossRef]
- van der Beek, C.M.; Canfora, E.E.; Kip, A.M.; Gorissen, S.H.M.; Olde Damink, S.W.M.; van Eijk, H.M.; Holst, J.J.; Blaak, E.E.; Dejong, C.H.C.; Lenaerts, K. The prebiotic inulin improves substrate metabolism and promotes short-chain fatty acid production in overweight to obese men. Metabolism 2018, 87, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Odamaki, T.; Mitsuyama, E.; Sugahara, H.; Xiao, J.-Z.; Osawa, R. Age-Related Changes in the Composition of Gut Bifidobacterium Species. Curr. Microbiol. 2017, 74, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Naito, Y.; Inoue, R.; Kashiwagi, S.; Uchiyama, K.; Mizushima, K.; Tsuchiya, S.; Dohi, O.; Yoshida, N.; Kamada, K.; et al. Differences in gut microbiota associated with age, sex, and stool consistency in healthy Japanese subjects. J. Gastroenterol. 2019, 54, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Parnell, J.A.; Reimer, R.A. Weight loss during oligofructose supplementation is associated with decreased ghrelin and increased peptide YY in overweight and obese adults. Am. J. Clin. Nutr. 2009, 89, 1751–1759. [Google Scholar] [CrossRef] [Green Version]
- Nogacka, A.M.; Salazar, N.; Arboleya, S.; Ruas-Madiedo, P.; Mancabelli, L.; Suarez, A.; Martinez-Faedo, C.; Ventura, M.; Tochio, T.; Hirano, K.; et al. In Vitro Evaluation of Different Prebiotics on the Modulation of Gut Microbiota Composition and Function in Morbid Obese and Normal-Weight Subjects. Int. J. Mol. Sci. 2020, 21, 906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ose, R.; Hirano, K.; Maeno, S.; Nakagawa, J.; Salminen, S.; Tochio, T.; Endo, A. The ability of human intestinal anaerobes to metabolize different oligosaccharides: Novel means for microbiota modulation? Anaerobe 2018, 51, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Tanno, H.; Fujii, T.; Ose, R.; Hirano, K.; Tochio, T.; Endo, A. Characterization of fructooligosaccharide-degrading enzymes in human commensal Bifidobacterium longum and Anaerostipes caccae. Biochem. Biophys. Res. Commun. 2019, 518, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Treuren, W.V.; Knight, R.; Bell, T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Ross, M.C.; Musky, D.M.; Mccormick, J.B.; Gibbs, R.A.; Fisher-Hoch, S.P.; Petrosino, J.F. 16S gut community of the Cameron County Hispanic Cohort. Microbiome 2015, 3, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duvallet, C.; Gibbons, S.M.; Gurry, T.; Irizarry, R.A.; Alm, E.J. Meta-analysis of gut microbiome studies identifies disease-specific and shared responses. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, A.; Hirano, K.; Ose, R.; Maeno, S.; Tochio, T. Impact of kestose supplementation on the healthy adult microbiota in in vitro fecal batch cultures. Anaerobe 2020, 61, 7. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Placebo | Kestose | p Value | ||
|---|---|---|---|---|---|
| (n = 18) | (n = 20) | ||||
| Sex (male/female) | 10/8 | 14/6 | |||
| Age (years) | 43.4 | (11.7) | 45.2 | (9.5) | 0.76 |
| Weight (kg) | 72.6 | (9.4) | 75.5 | (9.5) | 0.33 |
| Hight (cm) | 166.8 | (8.5) | 170.1 | (7.2) | 0.21 |
| BMI (kg/m2) * | 26.1 | (2.3) | 26.1 | (2.8) | 0.86 |
| HbA1c (%) | 5.4 | (0.3) | 5.4 | (0.3) | 0.77 |
| HbA1c (mmol/mol) | 35.1 | (2.9) | 35.0 | (3.3) | |
| Glucose (mg/dL) | 85.9 | (5.5) | 87.2 | (11.4) | 0.59 |
| Insulin (µU/mL) | 6.2 | (1.7) | 6.5 | (4.0) | 0.49 |
| HOMA-IR † | 1.3 | (0.4) | 1.5 | (1.2) | 0.35 |
| Total chol (mg/dL) | 191.3 | (18.1) | 201.1 | (27.4) | 0.16 |
| LDL chol (mg/dL) | 110.7 | (15.5) | 120.9 | (21.4) | 0.11 |
| HDL chol (mg/dL) | 55.6 | (10.2) | 55.5 | (12.8) | 0.97 |
| TG (mg/dL) | 89.6 | (40.5) | 88.4 | (31.3) | 0.91 |
| AST (U/L) | 18.8 | (5.4) | 20.2 | (7.3) | 0.97 |
| ALT (U/L) | 18.3 | (9.5) | 26.7 | (22.9) | 0.22 |
| γGT (U/L) | 39.9 | (44.8) | 38.8 | (21.5) | 0.36 |
| ALP (U/L) | 210.3 | (57.9) | 212.2 | (57.7) | 0.98 |
| BUN (mg/dL) | 12.9 | (2.9) | 13.8 | (2.7) | 0.40 |
| Creatinine (mg/dL) | 0.7 | (0.1) | 0.8 | (0.1) | 0.22 |
| Uric acid (mg/dL) | 5.0 | (1.5) | 5.7 | (1.2) | 0.20 |
| Albumin (g/dL) | 4.4 | (0.3) | 4.4 | (0.3) | 0.75 |
| Total protein (g/dL) | 7.5 | (0.4) | 7.4 | (0.4) | 0.47 |
| LDH (U/L) | 180.0 | (28.8) | 175.6 | (29.8) | 0.60 |
| Sodium (mEq/L) | 142.7 | (2.1) | 143.0 | (1.9) | 0.82 |
| Chloride (mEq/L) | 102.0 | (2.0) | 102.2 | (2.4) | 0.84 |
| Potassium (mEq/L) | 4.2 | (0.3) | 4.0 | (0.4) | 0.48 |
| Calcium (mg/dL) | 9.4 | (0.4) | 9.4 | (0.3) | 0.89 |
| I. Ph (mg/dL) | 4.1 | (1.1) | 4.0 | (0.9) | 0.87 |
| Mg (mg/dL) | 2.2 | (0.1) | 2.2 | (0.2) | 0.92 |
| Clinical Test Item | Placebo (n = 18) | Kestose (n = 20) | ||||||
|---|---|---|---|---|---|---|---|---|
| Baseline | Week 12 | Baseline | Week 12 | |||||
| Glucose (mg/dL) | ||||||||
| 0 min | 85.9 | (5.5) | 86.7 | (7.5) | 87.2 | (11.4) | 88.6 | (8.3) |
| 30 min | 136.0 | (21.3) | 132.1 | (19.7) | 145.1 | (23.4) | 142.4 | (28.6) |
| 60 min | 137.7 | (35.8) | 132.4 | (46.7) | 140.5 | (46.7) | 151.3 | (45.1) |
| 90 min | 122.3 | (28.3) | 118.5 | (32.9) | 126.6 | (38.5) | 121.4 | (35.9) |
| 120 min | 109.4 | (19.4) | 105.5 | (20.6) | 115.8 | (29.3) | 100.4 | (21.3) * |
| AUC [(mg/dL)h] | 75.1 | (37.2) | 67.7 | (43.0) | 84.3 | (48.0) | 78.1 | (48.2) |
| Insulin (µU/mL) | ||||||||
| 0 min | 6.2 | (1.7) | 6.5 | (2.1) | 6.5 | (4.0) | 5.3 | (1.4) |
| 30 min | 46.5 | (35.2) | 45.9 | (23.8) | 43.9 | (33.7) | 29.2 | (13.2) |
| 60 min | 45.5 | (21.0) | 49.2 | (24.2) | 39.7 | (21.7) | 42.7 | (19.9) |
| 90 min | 40.3 | (17.6) | 38.1 | (19.9) | 35.6 | (21.3) | 35.9 | (21.0) |
| 120 min | 35.5 | (18.1) | 36.6 | (13.5) | 34.8 | (22.0) | 36.5 | (27.9) |
| AUC [(µU/mL)min] | 128.3 | (58.9) | 128.7 | (53.3) | 113.9 | (55.7) | 107.5 | (52.0) |
| Placebo (n = 18) | Kestose (n = 20) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Baseline | Week 12 | Baseline | Week 12 | ||||||||||||
| Relative abundance | |||||||||||||||
| Blautia | 0.1665 | ± | 0.0777 | 0.1984 | ± | 0.0800 | 0.1530 | ± | 0.0777 | 0.1281 | ± | 0.0656 | †† | ||
| Bifidobacterium | 0.2723 | ± | 0.1201 | 0.1971 | ± | 0.1158 | * | 0.2209 | ± | 0.1762 | 0.3244 | ± | 0.1526 | * | †† |
| Sellimonas | 0.0031 | ± | 0.0072 | 0.0044 | ± | 0.0097 | 0.0009 | ± | 0.0022 | 0.0006 | ± | 0.0012 | † | ||
| Erysipelatoclostridium | 0.0022 | ± | 0.0030 | 0.0028 | ± | 0.0034 | 0.0007 | ± | 0.0010 | 0.0008 | ± | 0.0014 | † | ||
| Megasphaera | 0.0019 | ± | 0.0051 | 0.0025 | ± | 0.0081 | 0.0098 | ± | 0.0281 | 0.0143 | ± | 0.0364 | * | ||
| Streptococcus | 0.0414 | ± | 0.0478 | 0.0451 | ± | 0.0654 | 0.0379 | ± | 0.0429 | 0.0183 | ± | 0.0179 | * | ||
| Ruminiclostridium 5 | 0.0028 | ± | 0.0044 | 0.0085 | ± | 0.0137 | ** | 0.0091 | ± | 0.0204 | 0.0054 | ± | 0.0119 | ||
| Ruminococcaceae UCG-013 | 0.0052 | ± | 0.0088 | 0.0081 | ± | 0.0125 | * | 0.0039 | ± | 0.0046 | 0.0054 | ± | 0.0081 | ||
| Lachnospiraceae NK4A136 group | 0.0006 | ± | 0.0014 | 0.0029 | ± | 0.0044 | ** | 0.0040 | ± | 0.0078 | 0.0014 | ± | 0.0023 | ||
| Lactobacillus | 0.0034 | ± | 0.0065 | 0.0040 | ± | 0.0088 | 0.0020 | ± | 0.0076 | 0.0142 | ± | 0.0533 | ** | ||
| Turicibacter | 0.0035 | ± | 0.0083 | 0.0045 | ± | 0.0122 | 0.0034 | ± | 0.0064 | 0.0017 | ± | 0.0047 | * | ||
| Bacillus | 0.0004 | ± | 0.0009 | 0.0095 | ± | 0.0153 | * | 0.0051 | ± | 0.0092 | 0.0038 | ± | 0.0050 | ||
| [Ruminococcus] gnavus group | 0.0065 | ± | 0.0132 | 0.0142 | ± | 0.0349 | 0.0149 | ± | 0.0238 | 0.0064 | ± | 0.0084 | ** | ||
| Collinsella | 0.0953 | ± | 0.0530 | 0.0796 | ± | 0.0485 | * | 0.0676 | ± | 0.0414 | 0.0792 | ± | 0.0571 | ||
| Christensenellaceae R-7 group | 0.0020 | ± | 0.0046 | 0.0045 | ± | 0.0144 | 0.0050 | ± | 0.0130 | 0.0033 | ± | 0.0099 | * | ||
| Eggerthella | 0.0031 | ± | 0.0043 | 0.0037 | ± | 0.0043 | 0.0017 | ± | 0.0024 | 0.0011 | ± | 0.0015 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watanabe, A.; Tochio, T.; Kadota, Y.; Takahashi, M.; Kitaura, Y.; Ishikawa, H.; Yasutake, T.; Nakano, M.; Shinohara, H.; Kudo, T.; et al. Supplementation of 1-Kestose Modulates the Gut Microbiota Composition to Ameliorate Glucose Metabolism in Obesity-Prone Hosts. Nutrients 2021, 13, 2983. https://doi.org/10.3390/nu13092983
Watanabe A, Tochio T, Kadota Y, Takahashi M, Kitaura Y, Ishikawa H, Yasutake T, Nakano M, Shinohara H, Kudo T, et al. Supplementation of 1-Kestose Modulates the Gut Microbiota Composition to Ameliorate Glucose Metabolism in Obesity-Prone Hosts. Nutrients. 2021; 13(9):2983. https://doi.org/10.3390/nu13092983
Chicago/Turabian StyleWatanabe, Ayako, Takumi Tochio, Yoshihiro Kadota, Motoki Takahashi, Yasuyuki Kitaura, Hirohito Ishikawa, Takanori Yasutake, Masahiro Nakano, Hiroe Shinohara, Toru Kudo, and et al. 2021. "Supplementation of 1-Kestose Modulates the Gut Microbiota Composition to Ameliorate Glucose Metabolism in Obesity-Prone Hosts" Nutrients 13, no. 9: 2983. https://doi.org/10.3390/nu13092983
APA StyleWatanabe, A., Tochio, T., Kadota, Y., Takahashi, M., Kitaura, Y., Ishikawa, H., Yasutake, T., Nakano, M., Shinohara, H., Kudo, T., Nishimoto, Y., Mizuguchi, Y., Endo, A., & Shimomura, Y. (2021). Supplementation of 1-Kestose Modulates the Gut Microbiota Composition to Ameliorate Glucose Metabolism in Obesity-Prone Hosts. Nutrients, 13(9), 2983. https://doi.org/10.3390/nu13092983