
Four Weeks of 16/8 Time Restrictive Feeding in Endurance Trained Male Runners Decreases Fat Mass, without Affecting Exercise Performance


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Subjects
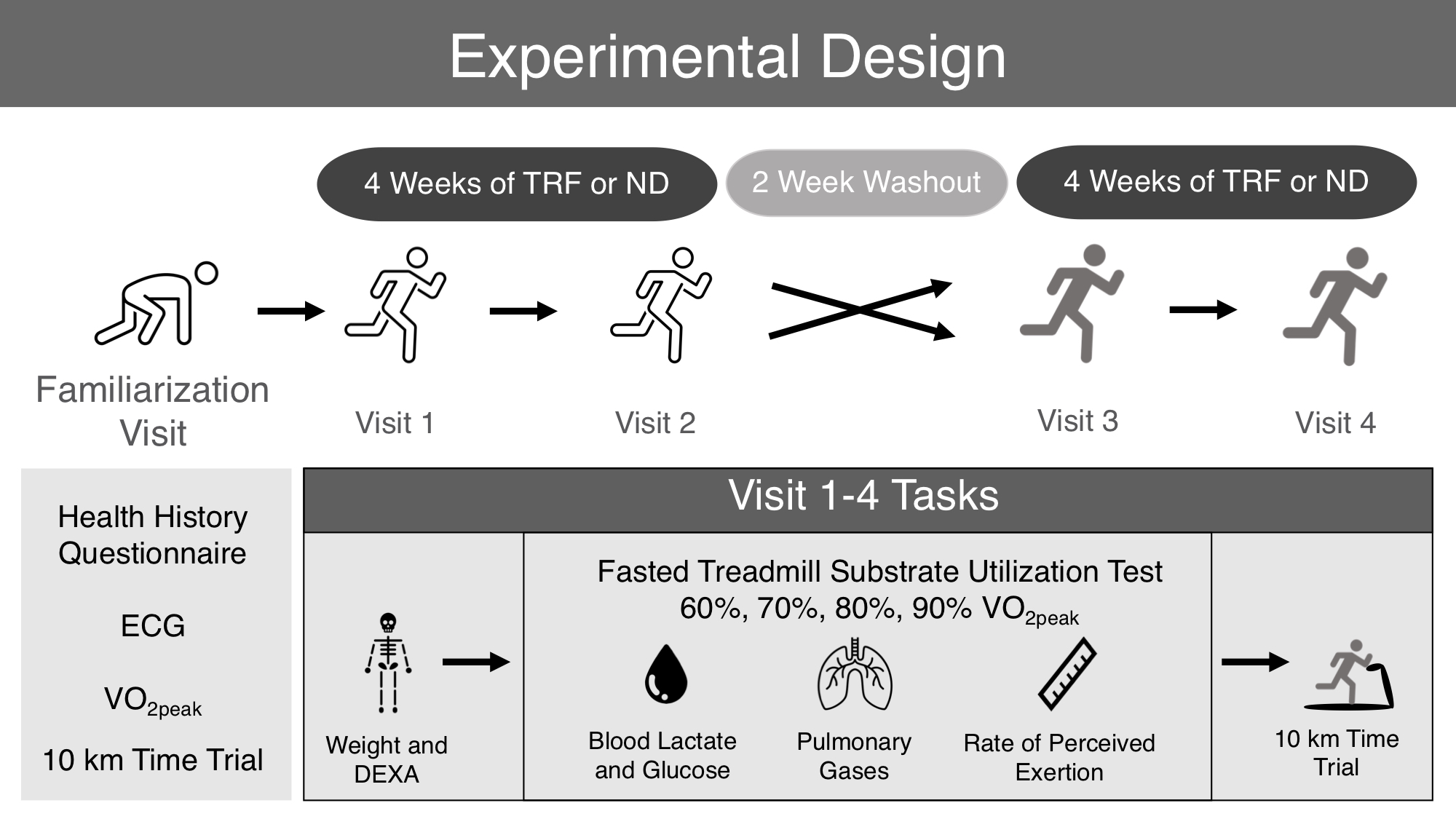
2.2. Experimental Design
2.3. Familiarization Visit
2.4. Assessment of Dietary Intake
2.5. Assessment of Training
2.6. Experimental Protocol for Test Days
2.7. Anthropometric Testing
2.8. Substrate Utilization Testing
2.9. 10 km Time Trial
2.10. Statistics
3. Results
3.1. Subject Characteristics and Dietary Patterns
3.2. Body Composition
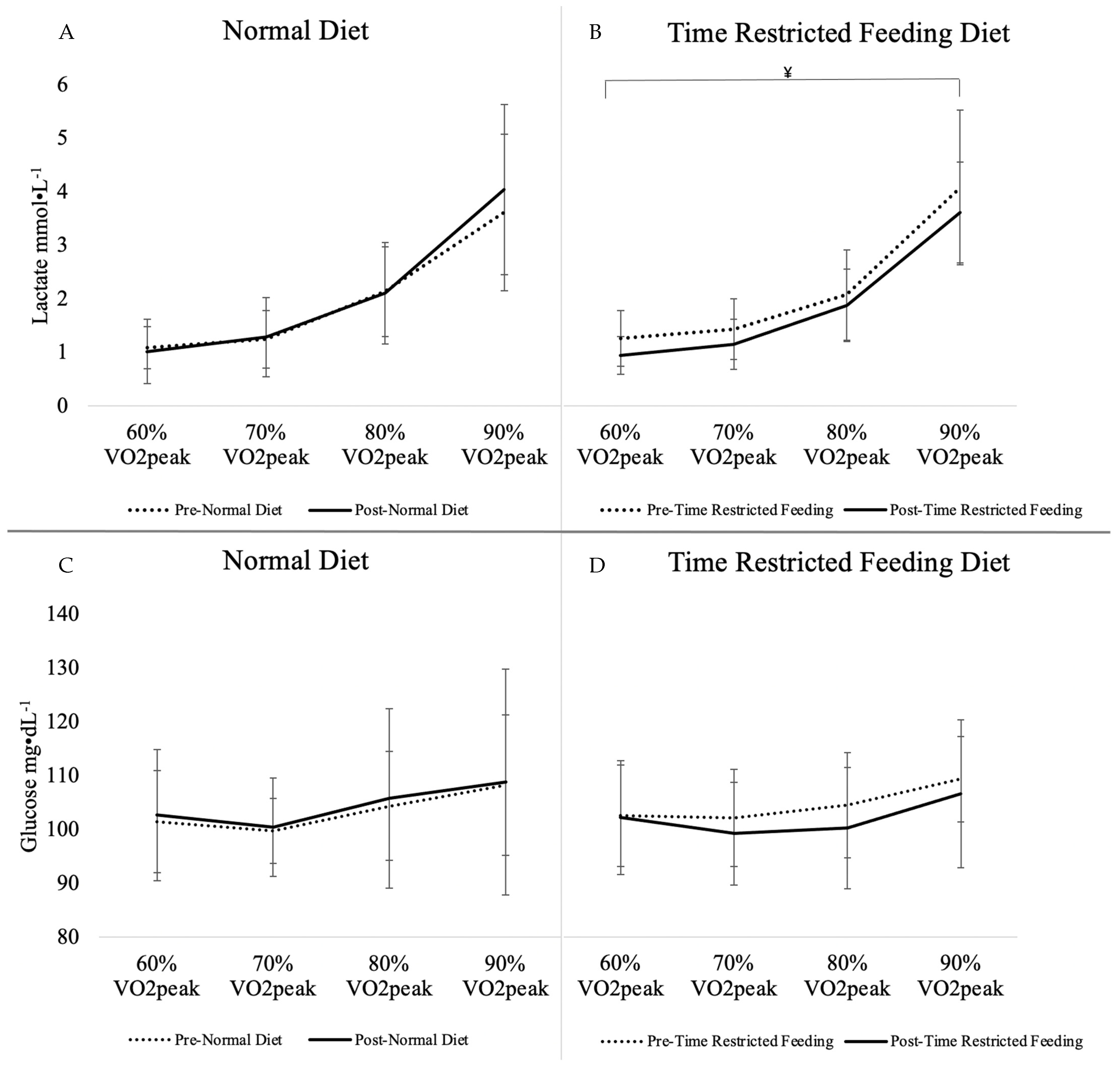
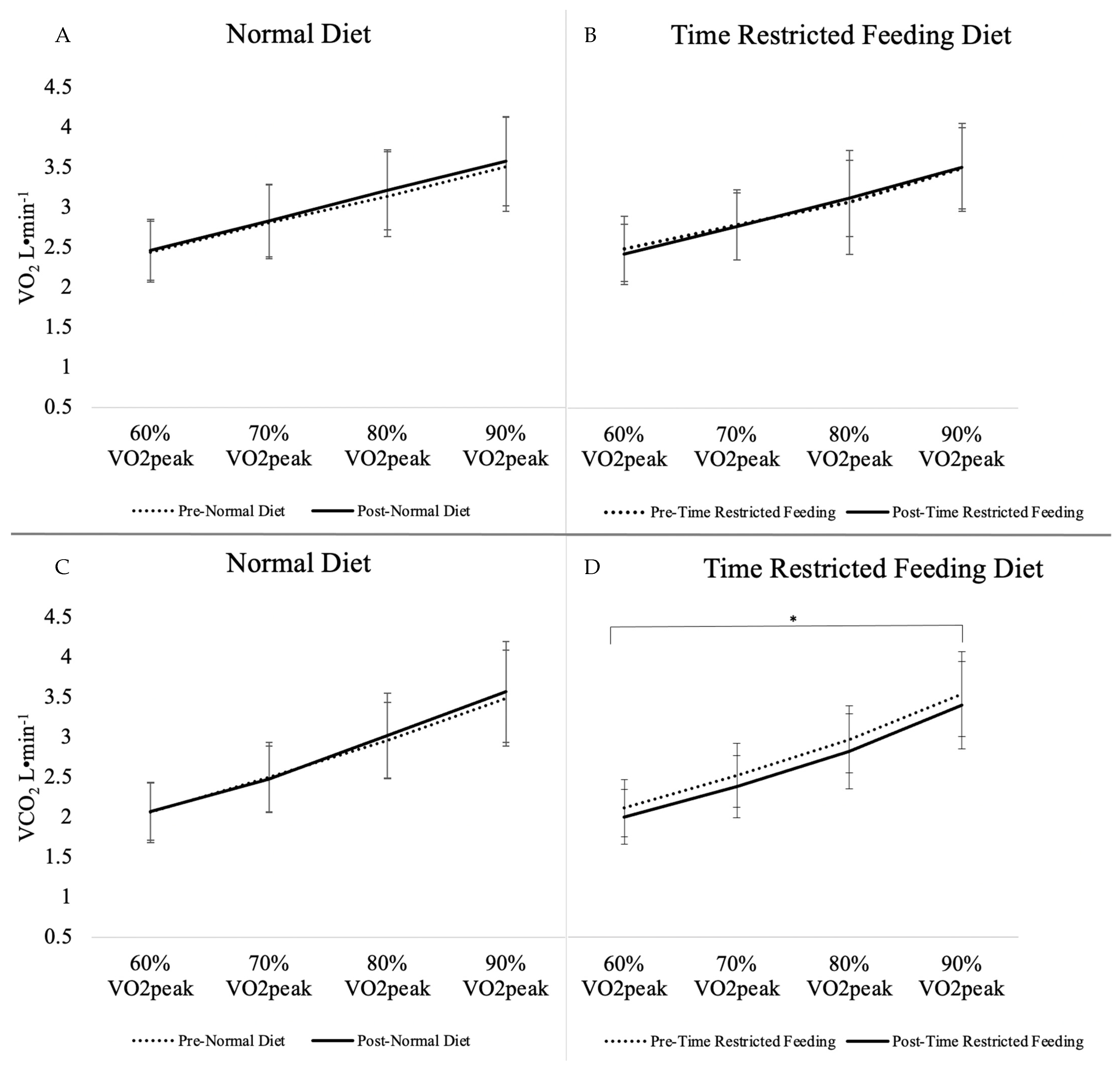
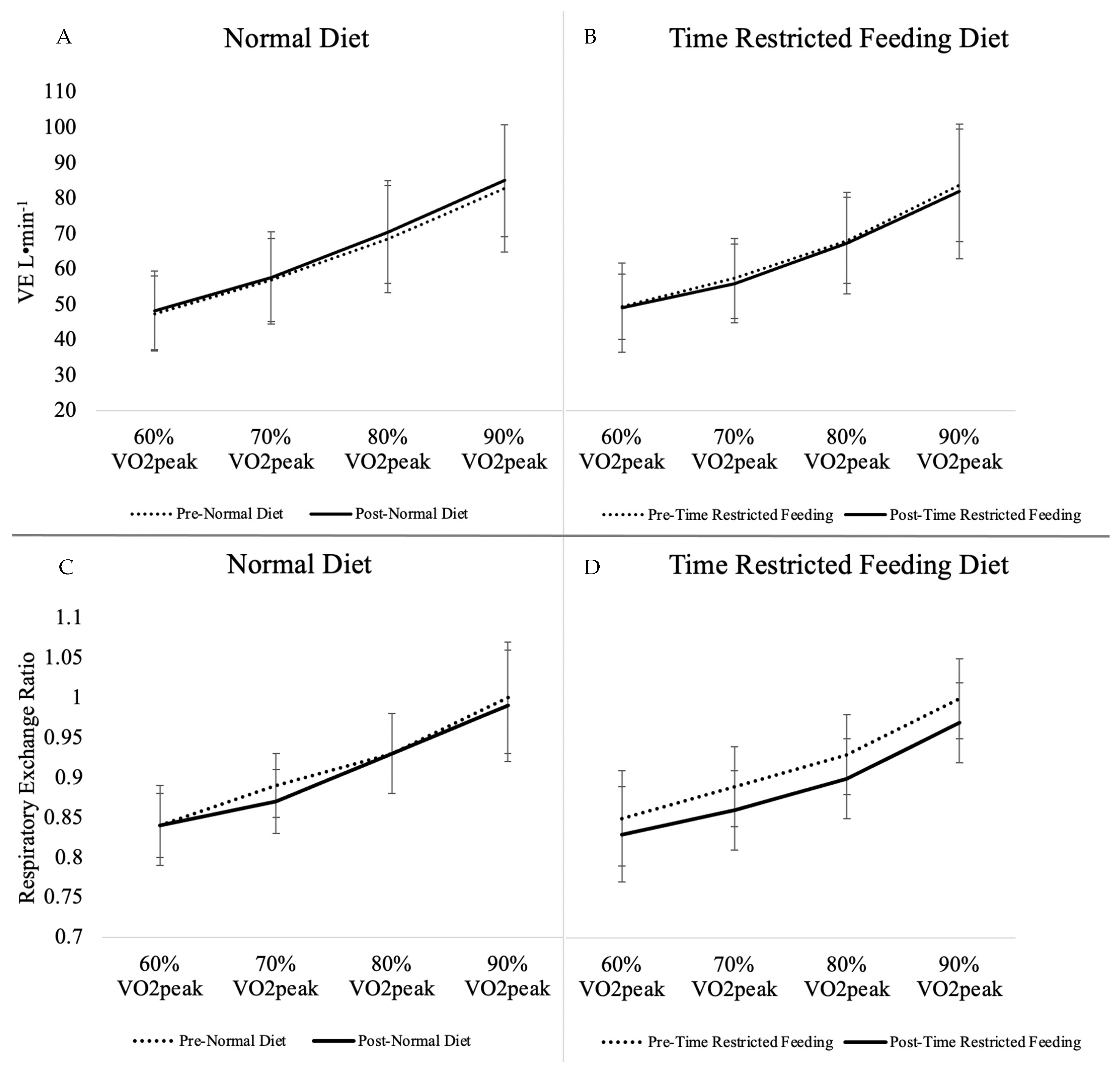
3.3. Incremental Exercise Test
3.4. Performance during 10 km Time Trial
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. American College of Sports Medicine Joint Position Statement. Nutrition and Athletic Performance. Med. Sci. Sports Exerc. 2016, 48, 543–568. [Google Scholar] [CrossRef]
- Stellingwerff, T.; Morton, J.P.; Burke, L.M. A Framework for Periodized Nutrition for Athletics. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Baar, K. Nutrition and the adaptation to endurance training. Sports Med. 2014, 44, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.M.; Ross, M.L.; Garvican-Lewis, L.A.; Welvaert, M.; Heikura, I.A.; Forbes, S.G.; Mirtschin, J.G.; Cato, L.E.; Strobel, N.; Sharma, A.P.; et al. Low carbohydrate, high fat diet impairs exercise economy and negates the performance benefit from intensified training in elite race walkers. J. Physiol. 2017, 595, 2785–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiśniewski, J.R.; Friedrich, A.; Keller, T.; Mann, M.; Koepsell, H. The impact of high-fat diet on metabolism and immune defense in small intestine mucosa. J. Proteome Res. 2015, 14, 353–365. [Google Scholar] [CrossRef]
- Paoli, A.; Tinsley, G.; Bianco, A.; Moro, T. The Influence of Meal Frequency and Timing on Health in Humans: The Role of Fasting. Nutrients 2019, 11, 719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabel, K.; Hoddy, K.K.; Haggerty, N.; Song, J.; Kroeger, C.M.; Trepanowski, J.F.; Panda, S.; Varady, K.A. Effects of 8-hour time restricted feeding on body weight and metabolic disease risk factors in obese adults: A pilot study. Nutr. Healthy Aging 2018, 4, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.; Panda, S. A Smartphone App Reveals Erratic Diurnal Eating Patterns in Humans that Can Be Modulated for Health Benefits. Cell Metab. 2015, 22, 789–798. [Google Scholar] [CrossRef] [Green Version]
- Kassab, S.E.; Abdul-Ghaffar, T.; Nagalla, D.S.; Sachdeva, U.; Nayar, U. Serum leptin and insulin levels during chronic diurnal fasting. Asia Pac. J. Clin. Nutr. 2003, 12, 483–487. [Google Scholar]
- Moro, T.; Tinsley, G.; Bianco, A.; Marcolin, G.; Pacelli, Q.F.; Battaglia, G.; Palma, A.; Gentil, P.; Neri, M.; Paoli, A. Effects of eight weeks of time-restricted feeding (16/8) on basal metabolism, maximal strength, body composition, inflammation, and cardiovascular risk factors in resistance-trained males. J. Transl. Med. 2016, 14, 290. [Google Scholar] [CrossRef]
- Tinsley, G.M.; Moore, M.L.; Graybeal, A.J.; Paoli, A.; Kim, Y.; Gonzales, J.U.; Harry, J.R.; VanDusseldorp, T.A.; Kennedy, D.N.; Cruz, M.R. Time-restricted feeding plus resistance training in active females: A randomized trial. Am. J. Clin. Nutr. 2019, 110, 628–640. [Google Scholar] [CrossRef] [Green Version]
- Alvero-Cruz, J.R.; Parent Mathias, V.; Garcia Romero, J.; Carrillo de Albornoz-Gil, M.; Benítez-Porres, J.; Ordoñez, F.J.; Rosemann, T.; Nikolaidis, P.T.; Knechtle, B. Prediction of Performance in a Short Trail Running Race: The Role of Body Composition. Front. Physiol. 2019, 10, 1306. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The "crossover" concept. J. Appl. Physiol. 1994, 76, 2253–2261. [Google Scholar] [CrossRef] [Green Version]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.; et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [Green Version]
- Brady, A.J.; Langton, H.M.; Mulligan, M.; Egan, B. Effects of Eight Weeks of 16: 8 Time-restricted Eating in Male Middle- and Long-Distance Runners. Med. Sci. Sports Exerc. 2020, 53, 633–642. [Google Scholar] [CrossRef]
- Mountjoy, M.; Sundgot-Borgen, J.; Burke, L.; Carter, S.; Constantini, N.; Lebrun, C.; Meyer, N.; Sherman, R.; Steffen, K.; Budgett, R.; et al. The IOC consensus statement: Beyond the Female Athlete Triad--Relative Energy Deficiency in Sport (RED-S). Br. J. Sports Med. 2014, 48, 491–497. [Google Scholar] [CrossRef]
- Munoz, I.; Seiler, S.; Alcocer, A.; Carr, N.; Esteve-Lanao, J. Specific Intensity for Peaking: Is Race Pace the Best Option? Asian J. Sports Med. 2015, 6, e24900. [Google Scholar] [CrossRef] [Green Version]
- Wehrens, S.M.T.; Christou, S.; Isherwood, C.; Middleton, B.; Gibbs, M.A.; Archer, S.N.; Skene, D.J.; Johnston, J.D. Meal Timing Regulates the Human Circadian System. Curr. Biol. 2017, 27, 1768–1775.e1763. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, A.T.; Regmi, P.; Manoogian, E.N.C.; Fleischer, J.G.; Wittert, G.A.; Panda, S.; Heilbronn, L.K. Time-Restricted Feeding Improves Glucose Tolerance in Men at Risk for Type 2 Diabetes: A Randomized Crossover Trial. Obesity (Silver Spring) 2019, 27, 724–732. [Google Scholar] [CrossRef]
- Sciarrillo, C.M.; Keirns, B.H.; Elliott, D.C.; Emerson, S.R. The effect of black coffee on fasting metabolic markers and an abbreviated fat tolerance test. Clin. Nutr. ESPEN 2021, 41, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Noble, B.J.; Borg, G.A.; Jacobs, I.; Ceci, R.; Kaiser, P. A category-ratio perceived exertion scale: Relationship to blood and muscle lactates and heart rate. Med. Sci. Sports Exerc. 1983, 15, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Beltz, N.M.; Gibson, A.L.; Janot, J.M.; Kravitz, L.; Mermier, C.M.; Dalleck, L.C. Graded Exercise Testing Protocols for the Determination of VO. J. Sports Med. 2016, 2016, 3968393. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.; Doust, J.H. A 1% treadmill grade most accurately reflects the energetic cost of outdoor running. J. Sports Sci. 1996, 14, 321–327. [Google Scholar] [CrossRef]
- Kasch, F.W.; Wallace, J.P.; Huhn, R.R.; Krogh, L.A.; Hurl, P.M. VO2max during horizontal and inclined treadmill running. J. Appl. Physiol. 1976, 40, 982–983. [Google Scholar] [CrossRef]
- Poole, D.C.; Jones, A.M. Measurement of the maximum oxygen uptake V̇o2max: V̇o2peak is no longer acceptable. J. Appl. Physiol. 2017, 122, 997–1002. [Google Scholar] [CrossRef]
- Howley, E.T.; Bassett, D.R.; Welch, H.G. Criteria for maximal oxygen uptake: Review and commentary. Med. Sci. Sports Exerc. 1995, 27, 1292–1301. [Google Scholar] [CrossRef]
- Jospe, M.R.; Fairbairn, K.A.; Green, P.; Perry, T.L. Diet app use by sports dietitians: A survey in five countries. JMIR Mhealth Uhealth 2015, 3, e7. [Google Scholar] [CrossRef]
- Beasley, J.M.; Riley, W.T.; Davis, A.; Singh, J. Evaluation of a PDA-based dietary assessment and intervention program: A randomized controlled trial. J. Am. Coll. Nutr. 2008, 27, 280–286. [Google Scholar] [CrossRef]
- Maringer, M.; Wisse-Voorwinden, N.; Veer, P.V.; Geelen, A. Food identification by barcode scanning in the Netherlands: A quality assessment of labelled food product databases underlying popular nutrition applications. Public Health Nutr. 2019, 22, 1215–1222. [Google Scholar] [CrossRef] [PubMed]
- Fallaize, R.; Zenun Franco, R.; Pasang, J.; Hwang, F.; Lovegrove, J.A. Popular Nutrition-Related Mobile Apps: An Agreement Assessment Against a UK Reference Method. JMIR Mhealth Uhealth 2019, 7, e9838. [Google Scholar] [CrossRef]
- Griffiths, C.; Harnack, L.; Pereira, M.A. Assessment of the accuracy of nutrient calculations of five popular nutrition tracking applications. Public Health Nutr. 2018, 21, 1495–1502. [Google Scholar] [CrossRef] [Green Version]
- Frayn, K.N. Calculation of substrate oxidation rates in vivo from gaseous exchange. J. Appl Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef] [Green Version]
- Russell, R.D.; Redmann, S.M.; Ravussin, E.; Hunter, G.R.; Larson-Meyer, D.E. Reproducibility of endurance performance on a treadmill using a preloaded time trial. Med. Sci. Sports Exerc. 2004, 36, 717–724. [Google Scholar] [CrossRef] [Green Version]
- San-Millán, I.; Brooks, G.A. Assessment of Metabolic Flexibility by Means of Measuring Blood Lactate, Fat, and Carbohydrate Oxidation Responses to Exercise in Professional Endurance Athletes and Less-Fit Individuals. Sports Med. 2018, 48, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Koubi, H.E.; Desplanches, D.; Gabrielle, C.; Cottet-Emard, J.M.; Sempore, B.; Favier, R.J. Exercise endurance and fuel utilization: A reevaluation of the effects of fasting. J. Appl. Physiol. 1991, 70, 1337–1343. [Google Scholar] [CrossRef]
- De Bock, K.; Derave, W.; Eijnde, B.O.; Hesselink, M.K.; Koninckx, E.; Rose, A.J.; Schrauwen, P.; Bonen, A.; Richter, E.A.; Hespel, P. Effect of training in the fasted state on metabolic responses during exercise with carbohydrate intake. J. Appl. Physiol. 2008, 104, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Van Proeyen, K.; Szlufcik, K.; Nielens, H.; Ramaekers, M.; Hespel, P. Beneficial metabolic adaptations due to endurance exercise training in the fasted state. J. Appl. Physiol. 2011, 110, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Anton, S.D.; Moehl, K.; Donahoo, W.T.; Marosi, K.; Lee, S.A.; Mainous, A.G.; Leeuwenburgh, C.; Mattson, M.P. Flipping the Metabolic Switch: Understanding and Applying the Health Benefits of Fasting. Obesity (Silver Spring) 2018, 26, 254–268. [Google Scholar] [CrossRef]
- Dubois, V.; Eeckhoute, J.; Lefebvre, P.; Staels, B. Distinct but complementary contributions of PPAR isotypes to energy homeostasis. J. Clin. Investig. 2017, 127, 1202–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinsley, G.M.; Forsse, J.S.; Butler, N.K.; Paoli, A.; Bane, A.A.; La Bounty, P.M.; Morgan, G.B.; Grandjean, P.W. Time-restricted feeding in young men performing resistance training: A randomized controlled trial. Eur. J. Sport Sci. 2017, 17, 200–207. [Google Scholar] [CrossRef]
- Wilkinson, D.J.; Brook, M.S.; Smith, K.; Atherton, P.J. Stable isotope tracers and exercise physiology: Past, present and future. J. Physiol. 2017, 595, 2873–2882. [Google Scholar] [CrossRef] [Green Version]
- Barnes, K.R.; Kilding, A.E. Running economy: Measurement, norms, and determining factors. Sports Med. Open 2015, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Burke, L.M.; Jeukendrup, A.E.; Jones, A.M.; Mooses, M. Contemporary Nutrition Strategies to Optimize Performance in Distance Runners and Race Walkers. Int. J. Sport Nutr. Exerc. Metab. 2019, 29, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Noakes, T.D. Physiological models to understand exercise fatigue and the adaptations that predict or enhance athletic performance. Scand. J. Med. Sci. Sports 2000, 10, 123–145. [Google Scholar] [CrossRef] [Green Version]
- Philp, A.; Hargreaves, M.; Baar, K. More than a store: Regulatory roles for glycogen in skeletal muscle adaptation to exercise. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1343–E1351. [Google Scholar] [CrossRef] [Green Version]
- Uusitalo, A.L.; Huttunen, P.; Hanin, Y.; Uusitalo, A.J.; Rusko, H.K. Hormonal responses to endurance training and overtraining in female athletes. Clin. J. Sport Med. 1998, 8, 178–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaal, K.; Van Loan, M.D.; Casazza, G.A. Reduced catecholamine response to exercise in amenorrheic athletes. Med. Sci. Sports Exerc. 2011, 43, 34–43. [Google Scholar] [CrossRef]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.; Wightman, S.; Basevitch, I.; Johnstone, J.; Espejo-Sanchez, C.; Beckford, C.; Boal, M.; Scruton, A.; Ferrandino, M.; Merzbach, V. Physiological and training characteristics of recreational marathon runners. Open Access J. Sports Med. 2017, 8, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Tarnopolsky, M.A. Gender differences in substrate metabolism during endurance exercise. Can. J. Appl. Physiol. 2000, 25, 312–327. [Google Scholar] [CrossRef] [PubMed]
- Hector, A.J.; Phillips, S.M. Protein Recommendations for Weight Loss in Elite Athletes: A Focus on Body Composition and Performance. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Jesus, F.; Castela, I.; Silva, A.M.; Branco, P.A.; Sousa, M. Risk of Low Energy Availability among Female and Male Elite Runners Competing at the 26th European Cross-Country Championships. Nutrients 2021, 13, 873. [Google Scholar] [CrossRef]
- Misra, M. Neuroendocrine mechanisms in athletes. Handb. Clin. Neurol. 2014, 124, 373–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamshed, H.; Beyl, R.A.; Della Manna, D.L.; Yang, E.S.; Ravussin, E.; Peterson, C.M. Early Time-Restricted Feeding Improves 24-Hour Glucose Levels and Affects Markers of the Circadian Clock, Aging, and Autophagy in Humans. Nutrients 2019, 11, 1234. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, F.; Epstein, D.A.; Thomaz, E.; Bales, E.; Jagannathan, A.K.; Abowd, G.D.; Fogarty, J. Barriers and Negative Nudges: Exploring Challenges in Food Journaling. In Proceedings of the 33rd Annual ACM Conference on Human Factors in Computing Systems, Seoul, Korea, 18–23 April 2015; pp. 1159–1162. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (years) | 28.7 ± 5.2 |
| Height (cm) | 177.7 ± 6.6 |
| Weight (kg) | 73.5 ± 8.6 |
| Fat Free Mass (kg) | 57.6 ± 7.6 |
| Fat mass (kg) | 12.0 ± 4.5 |
| Body Fat (%) | 16.5 ± 5.6 |
| VO2peak (mL·kg·min−1) | 55.5 ± 5.7 |
| VO2peak speed (km·h−1) | 16.3 ± 1.8 |
| Average Running Distance (km·week−1) | 53.0 ± 24.1 |
| Years of Training | 7.8 ± 6.0 |
| Normal Diet | TRF Diet | p-Value | |
|---|---|---|---|
| Total caloric intake (kcal·day−1) | 2513 ± 367 | 2421 ± 478 | 0.41 |
| Total carbohydrate intake (g·day−1) | 284.8 ± 79.3 | 269.4 ± 68.4 | 0.27 |
| Total carbohydrate intake (g·kg−1·day−1) | 3.9 ± 1.2 | 3.7 ± 1.2 | 0.19 |
| Total protein intake (g·day−1) | 112.5 ± 27.1 | 113.1 ± 24.4 | 0.42 |
| Total protein intake (g·kg−1·day−1) | 1.6 ± 0.4 | 1.6 ± 0.4 | 0.72 |
| Total fat intake (g·day−1) | 97.5 ± 24.5 | 96.8 ± 33.0 | 0.91 |
| Caloric intake-HIT days (kcal·day−1) | 2626 ± 524 | 2493 ± 495 | 0.20 |
| Carbohydrate intake-HIT days (g·day−1) | 307.5 ± 99.9 | 271.6 ± 70.0 | 0.07 |
| Protein intake-HIT days (g·day−1) | 120.1 ± 42.1 | 117.0 ± 27.9 | 0.64 |
| Fat intake-HIT days (g·day−1) | 97.9 ± 38.2 | 98.7 ± 35.9 | 0.94 |
| Caloric intake-MIT days (kcal·day−1) | 2421 ± 360 | 2360 ± 475 | 0.58 |
| Carbohydrate intake-MIT days (g·day−1) | 253.1 ± 71.7 | 271.2 ± 69.7 | 0.41 |
| Protein intake-MIT days (g·day−1) | 109.0 ± 29.0 | 107.6 ± 26.3 | 0.91 |
| Fat intake-MIT days (g·day−1) | 101.5 ± 27.5 | 92.3 ± 36.1 | 0.23 |
| Caloric intake-rest days (kcal·day−1) | 2489 ± 475 | 2401 ± 559 | 0.62 |
| Carbohydrate intake-rest days (g·day−1) | 293.9 ± 104.9 | 265.3 ± 86.3 | 0.18 |
| Protein intake-rest days (g·day−1) | 108.4 ± 29.4 | 115.4 ± 27.7 | 0.17 |
| Fat intake-rest days (g·day−1) | 93.1 ± 18.6 | 99.3 ± 34.5 | 0.22 |
| Exercise (running km·week−1) | 39.3 ± 14.2 | 43.0 ± 20.4 | 0.18 |
| Feeding window (hours) | 11.8 ± 0.6 | 7.6 ± 0.4 | <0.01 * |
| Pre-Normal Diet | Post-Normal Diet | Δ Normal Diet | Pre-Time Restricted Feeding | Post-Time Restricted Feeding | Δ Time Restricted Feeding | Mixed Linear Model Diet p-Value | |
|---|---|---|---|---|---|---|---|
| Body Mass (kg) | 73.0 ± 8.6 | 73.3 ± 8.7 | +0.3 ± 1.1 | 73.8 ± 8.6 | 73.0 ± 9.0 | −0.8 ± 1.9 | 0.09 |
| Lean Mass (kg) | 57.6 ± 7.2 | 58.3 ±7.8 | +0.7 ± 2.4 | 57.7 ± 7.3 | 57.8 ± 7.2 | +0.1 ± 1.7 | 0.45 |
| Fat Mass (kg) | 11.7 ± 4.8 | 11.8 ± 4.3 | +0.1 ± 4.3 | 12.3 ± 4.3 | 11.5 ± 4.4 | −0.8 ± 1.3 | 0.05 * |
| Body Fat % | 16.1 ± 5.7 | 16.2 ± 5.3 | +0.1 ± 1.3 | 16.8 ± 5.3 | 15.8 ± 5.2 | −1.0 ± 1.5 | 0.04 * |
| ND Pre | ND Post | ΔND | TRF Pre | TRF Post | ΔTRF | Mixed Linear Model Diet p-Value | Mixed Linear Model Diet * Intensity p-Value | |
|---|---|---|---|---|---|---|---|---|
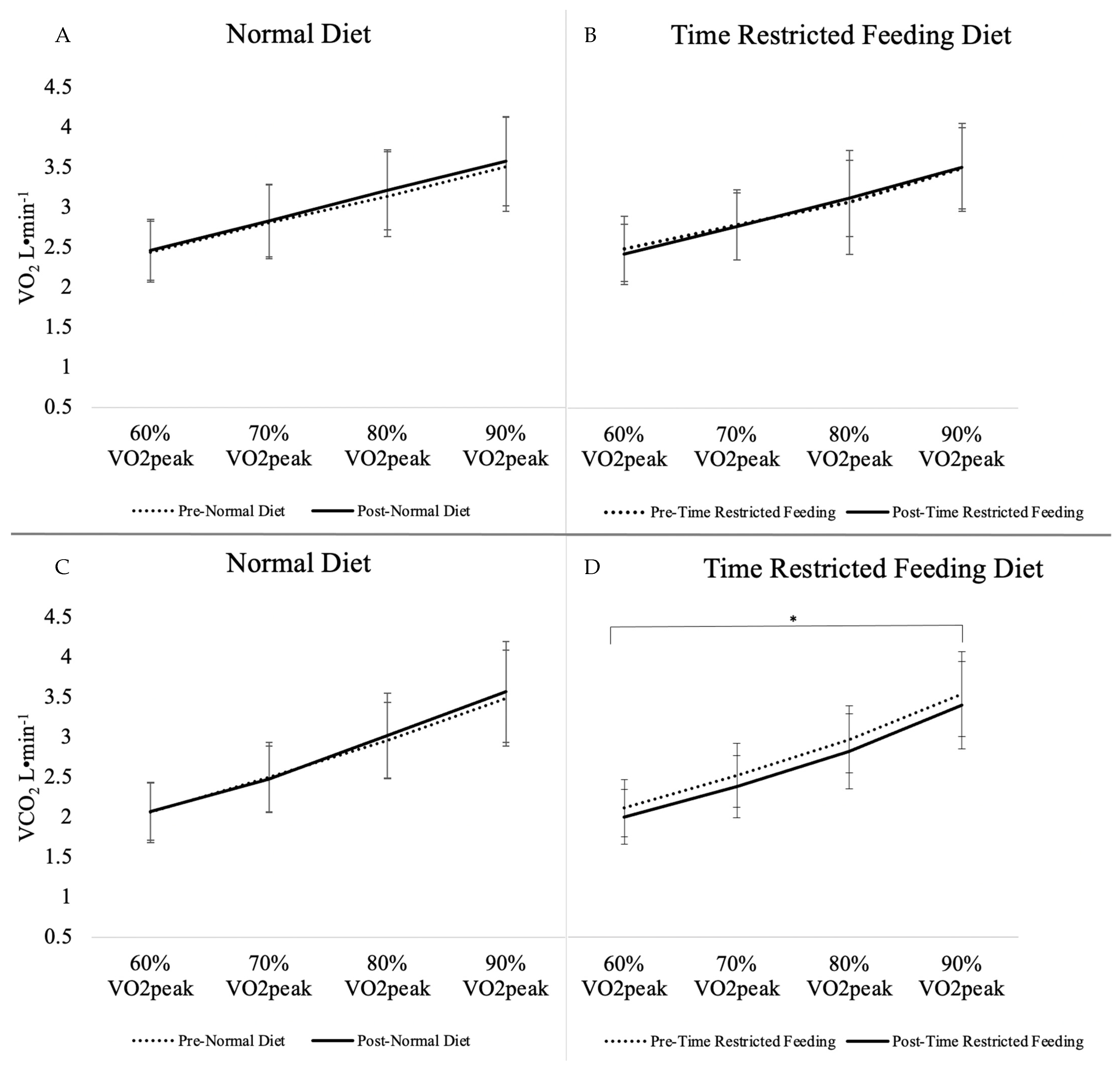
| VO2 (L·min−1) | 2.97 ± 0.65 | 3.02 ± 0.63 | +0.05 ± 0.14 | 2.97 ± 0.62 | 2.97 ± 0.61 | 0.00 ± 0.18 | 0.09 | 0.88 |
| VO2 (ml·kg·min−1) | 39.98 ± 8.02 | 40.43 ± 7.92 | +0.45 ± 1.50 | 39.95 ± 7.13 | 40.04 ± 7.72 | +0.09 ± 1.81 | 0.42 | 0.98 |
| VCO2 (L·min−1) | 2.75 ± 0.71 | 2.78 ± 0.74 | +0.03 ± 0.22 | 2.79 ± 0.68 | 2.66 ± 0.68 | −0.13 ± 0.12 | <0.01 * | 0.23 |
| VE STDP (L·min−1) | 63.79 ± 18.76 | 65.37 ± 19.61 | +1.58 ± 5.49 | 64.69 ± 17.72 | 63.66 ± 18.96 | −1.03 ± 4.99 | 0.08 | 0.64 |
| RPE (0-10) | 3.91 ± 2.43 | 3.73 ± 2.25 | −0.18 ± 0.91 | 3.80 ± 2.21 | 3.90 ± 2.29 | +0.10 ± 1.11 | 0.36 | 0.14 |
| HR (BPM) | 159.77 ± 18.13 | 158. 28 ± 18.39 | −1.49 ± 4.66 | 158.98 ± 18.48 | 157.74 ± 18.33 | −1.24 ± 6.38 | 0.86 | 0.28 |
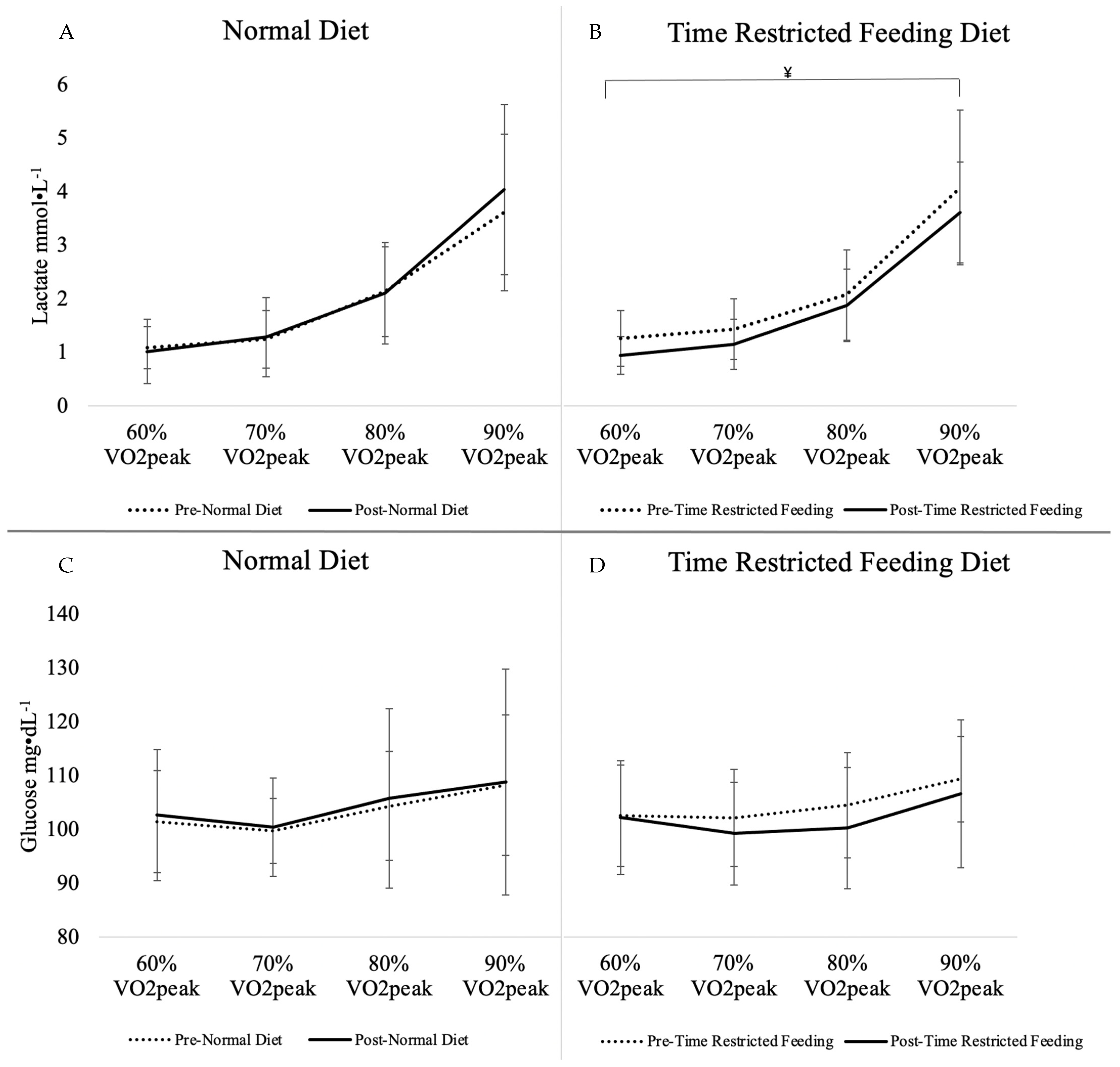
| Lactate (mmol·L−1) | 2.02 ± 1.34 | 2.11 ± 1.57 | +0.09 ± 0.75 | 2.23 ± 1.45 | 1.91 ± 1.23 | −0.32 ± 0.55 | 0.07 | 0.02 * |
| Glucose (mg·L−1) | 103.50 ± 10.43 | 104.52 ± 15.61 | +1.02 ± 13.50 | 104.89 ± 9.38 | 102.22 ± 11.56 | −2.67 ± 9.87 | 0.19 | 0.58 |
| RER | 0.92 ± 0.07 | 0.91 ± 0.08 | −0.01 ± 0.05 | 0.92 ± 0.08 | 0.89 ± 0.07 | −0.03 ± 0.03 | 0.15 | 0.70 |
| % Fat Oxidation | 27.55 ± 26.36 | 30.30 ± 27.26 | −2.75 ± 17.89 | 28.06 ± 25.79 | 37.48 ± 24.93 | +9.42 ± 10.60 | 0.11 | 0.81 |
| % Carbohydrate Oxidation | 72.45 ± 26.36 | 69.70 ± 27.26 | −2.75 ± 17.89 | 71.94 ± 25.79 | 62.52 ± 24.93 | −9.42 ± 10.60 | 0.11 | 0.81 |
| ND Pre | ND Post | ΔND | TRF Pre | TRF Post | ΔTRF | Mixed Linear Model Diet | |
|---|---|---|---|---|---|---|---|
| Time (min:sec) | 50:02 ± 10:33 | 49:26 ± 10:04 | −00:36 ± 2:57 | 48:42 ± 8:39 | 48:22 ± 9:24 | −00:20 ± 3:34 | 0.53 |
| Average HR (bpm) | 167.8 + 11.9 | 169.5 ± 10.6 | +1.7 ± 4.7 | 170.5 ± 8.5 | 167.6 ± 12.3 | −2.9 ± 13.9 | 0.20 |
| Maximal HR (bpm) | 179.0 ± 13.3 | 181.8 ± 6.9 | +2.8 ± 8.3 | 181.4 ± 7.5 | 180.8 ± 12.3 | −0.6 ± 11.8 | 0.40 |
| Peak RPE (0–10) | 7.6 ± 2.3 | 7.3 ± 2.2 | −0.4 ± 1.3 | 7.3 ± 1.9 | 7.3 ± 1.8 | 0.0 ± 1.3 | 0.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tovar, A.P.; Richardson, C.E.; Keim, N.L.; Van Loan, M.D.; Davis, B.A.; Casazza, G.A. Four Weeks of 16/8 Time Restrictive Feeding in Endurance Trained Male Runners Decreases Fat Mass, without Affecting Exercise Performance. Nutrients 2021, 13, 2941. https://doi.org/10.3390/nu13092941
Tovar AP, Richardson CE, Keim NL, Van Loan MD, Davis BA, Casazza GA. Four Weeks of 16/8 Time Restrictive Feeding in Endurance Trained Male Runners Decreases Fat Mass, without Affecting Exercise Performance. Nutrients. 2021; 13(9):2941. https://doi.org/10.3390/nu13092941
Chicago/Turabian StyleTovar, Ashley P., Christine E. Richardson, Nancy L. Keim, Marta D. Van Loan, Brian A. Davis, and Gretchen A. Casazza. 2021. "Four Weeks of 16/8 Time Restrictive Feeding in Endurance Trained Male Runners Decreases Fat Mass, without Affecting Exercise Performance" Nutrients 13, no. 9: 2941. https://doi.org/10.3390/nu13092941
APA StyleTovar, A. P., Richardson, C. E., Keim, N. L., Van Loan, M. D., Davis, B. A., & Casazza, G. A. (2021). Four Weeks of 16/8 Time Restrictive Feeding in Endurance Trained Male Runners Decreases Fat Mass, without Affecting Exercise Performance. Nutrients, 13(9), 2941. https://doi.org/10.3390/nu13092941