
Differential Effects of Western and Mediterranean-Type Diets on Gut Microbiota: A Metagenomics and Metabolomics Approach

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Participants
2.3. Intervention
2.4. Outcomes
2.4.1. Daily Symptom Questionnaire and Bowel Habit
2.4.2. Number of Anal Gas Evacuations
2.4.3. Intestinal Gas Production
2.4.4. Response to a Comfort Meal
2.4.5. Colonic Content Measurement
2.4.6. Microbiota Composition
2.4.7. Metabolomic Analysis
2.5. Statistical Analysis
3. Results
3.1. Demographics and Study Flow
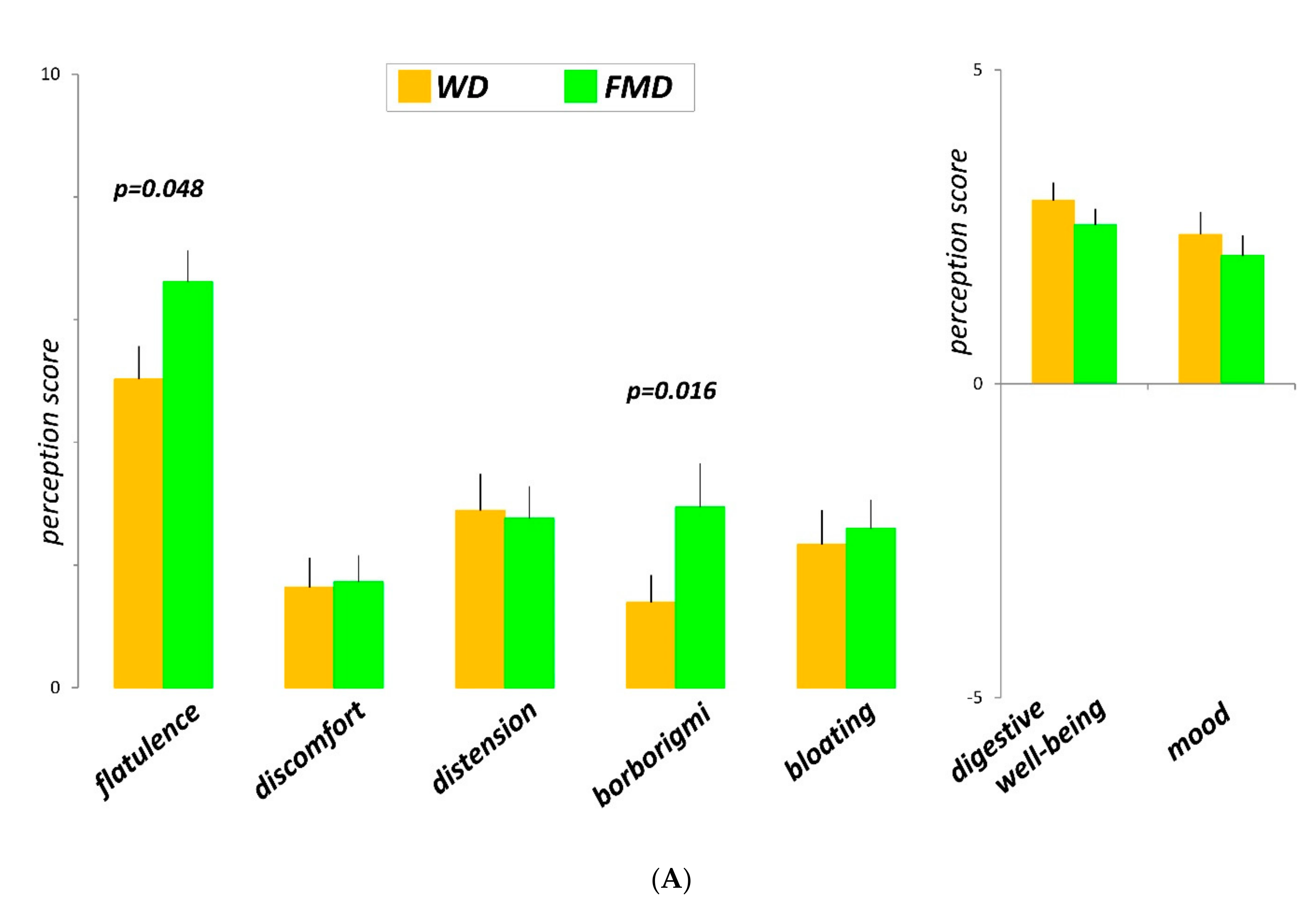
3.2. Clinical Parameters: Sensations and Bowel Habit
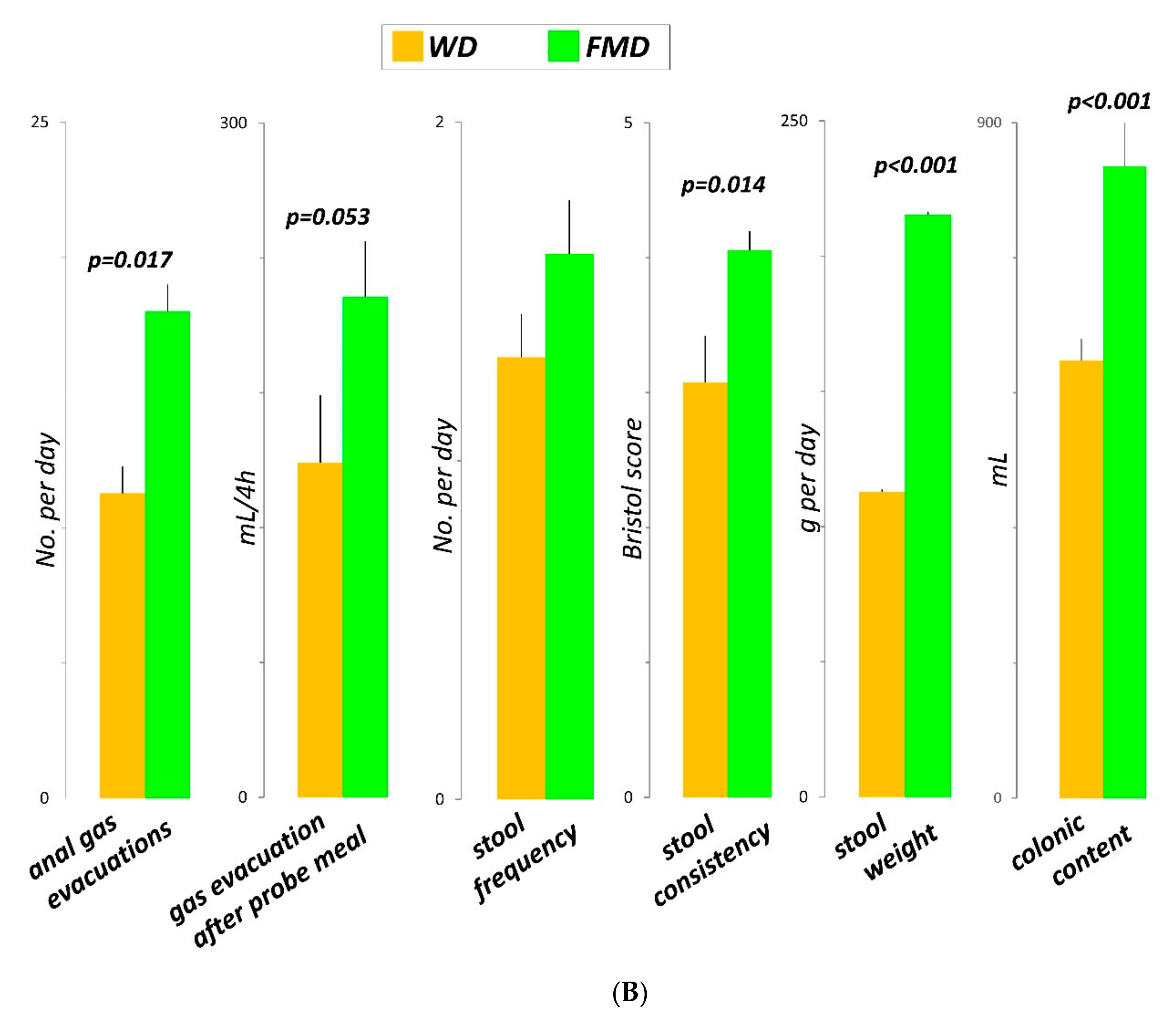
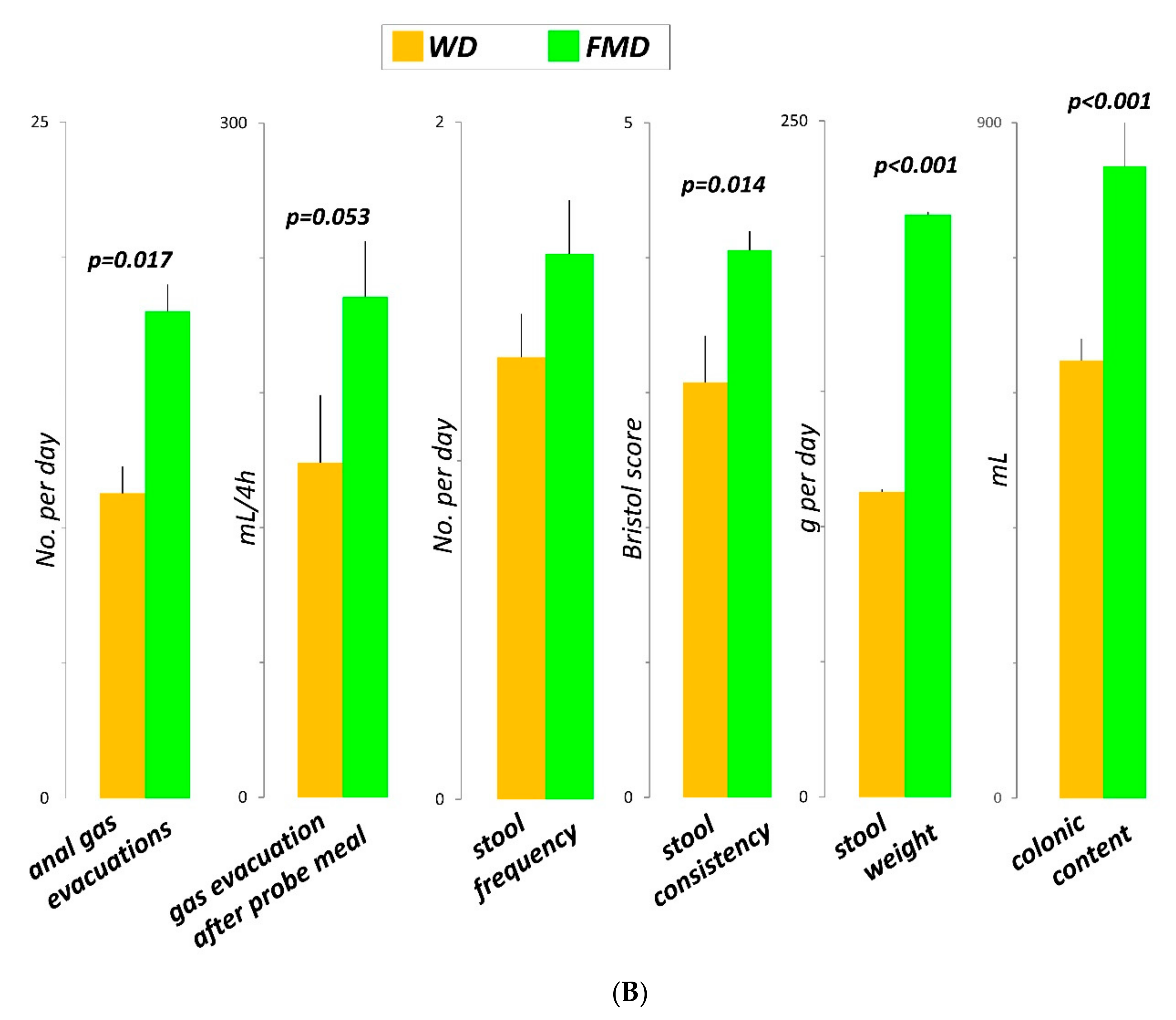
3.3. Number of Anal Gas Evacuations
3.4. Volume of Gas Evacuated after the Probe Meal
3.5. Responses to the Comfort Meal
3.6. Colonic Content
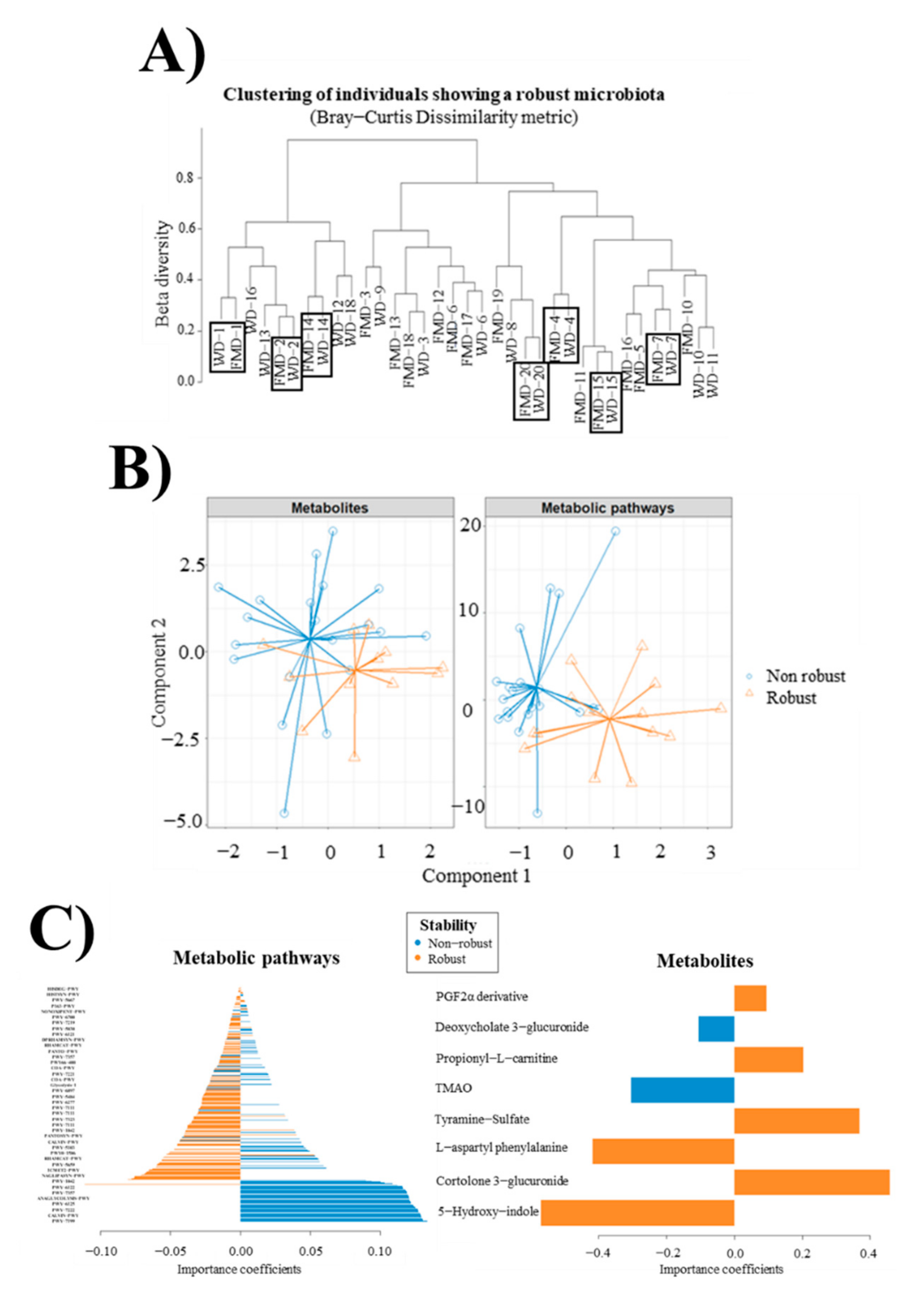
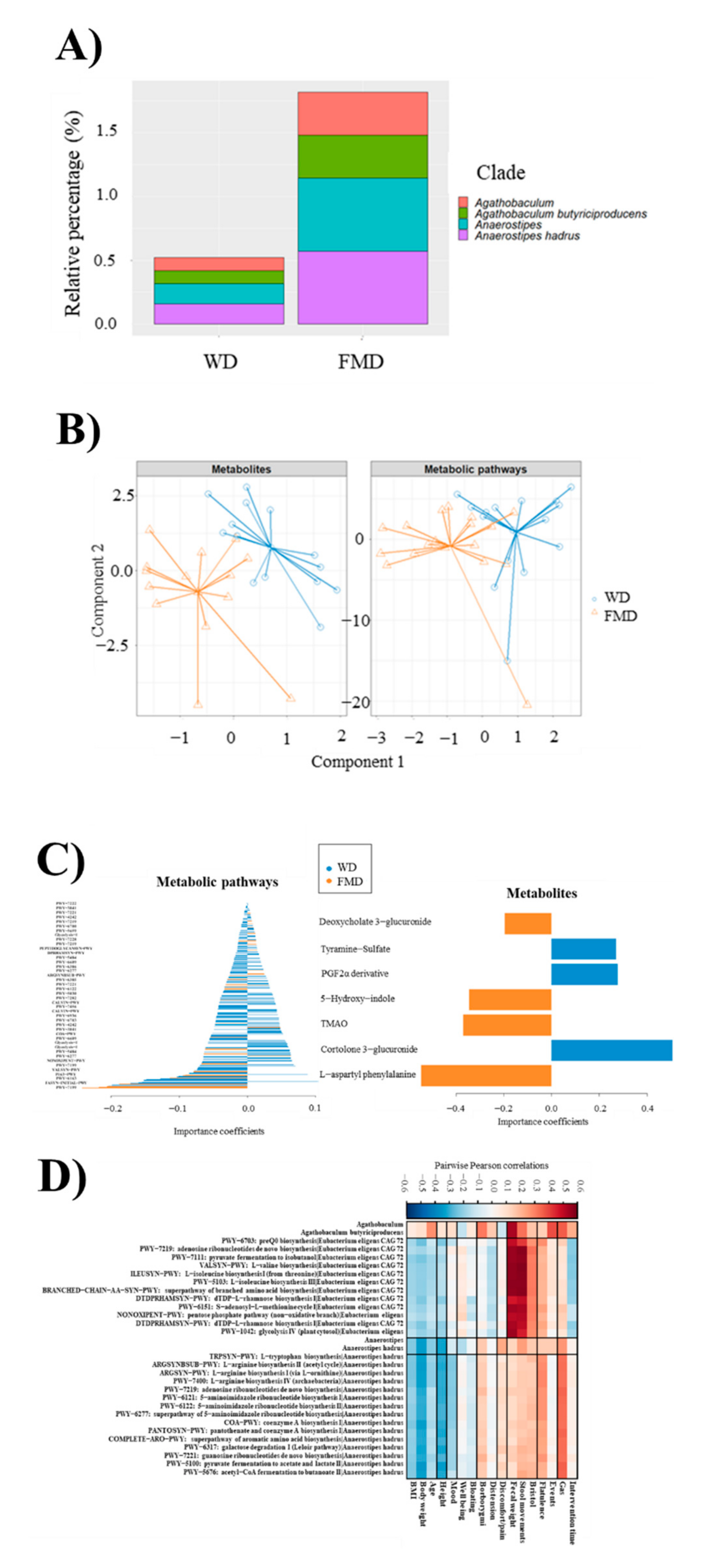
3.7. Microbiome Metagenomic Analysis
3.8. Metabolomic Analysis
3.9. Multi-Omic Analysis
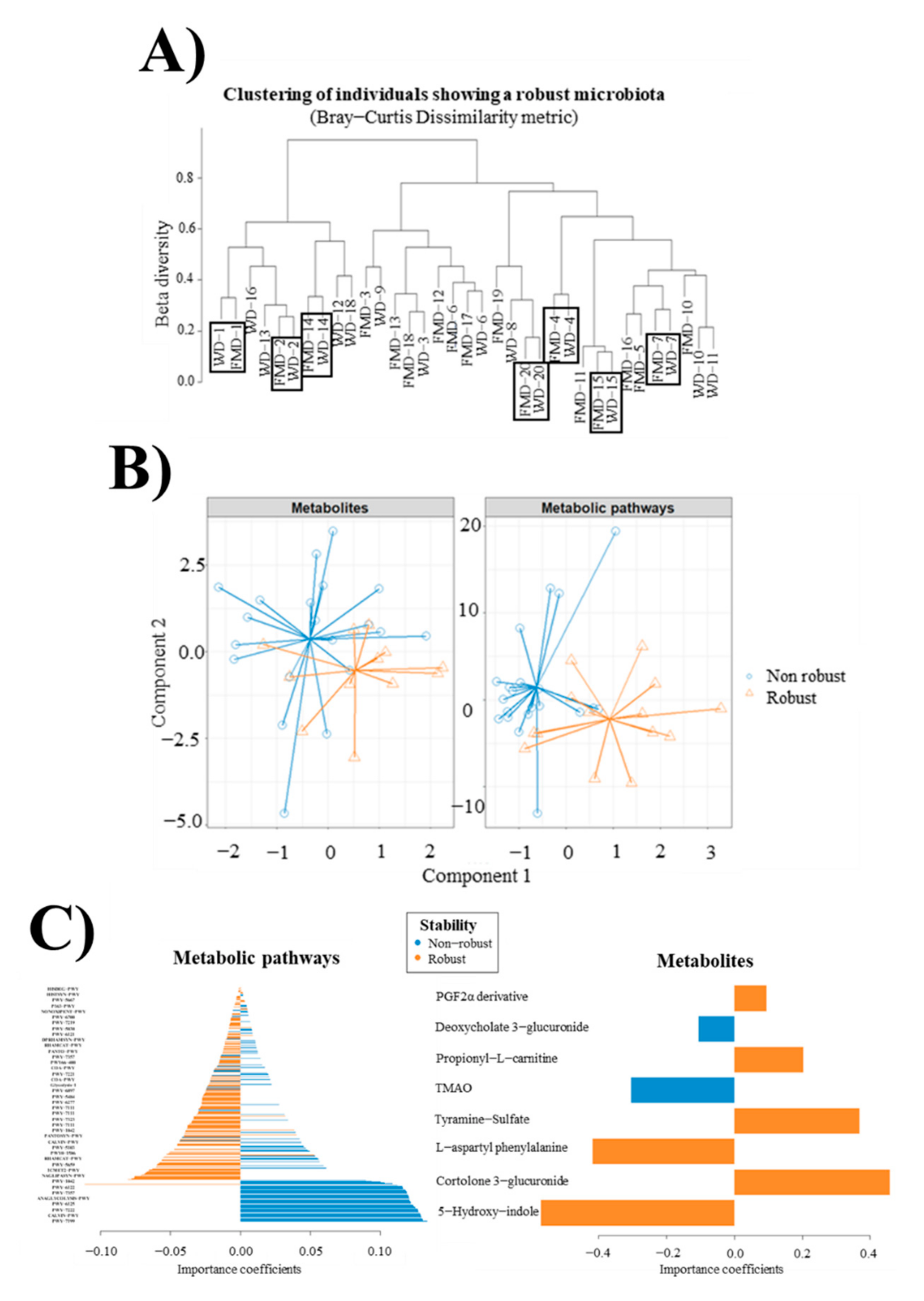
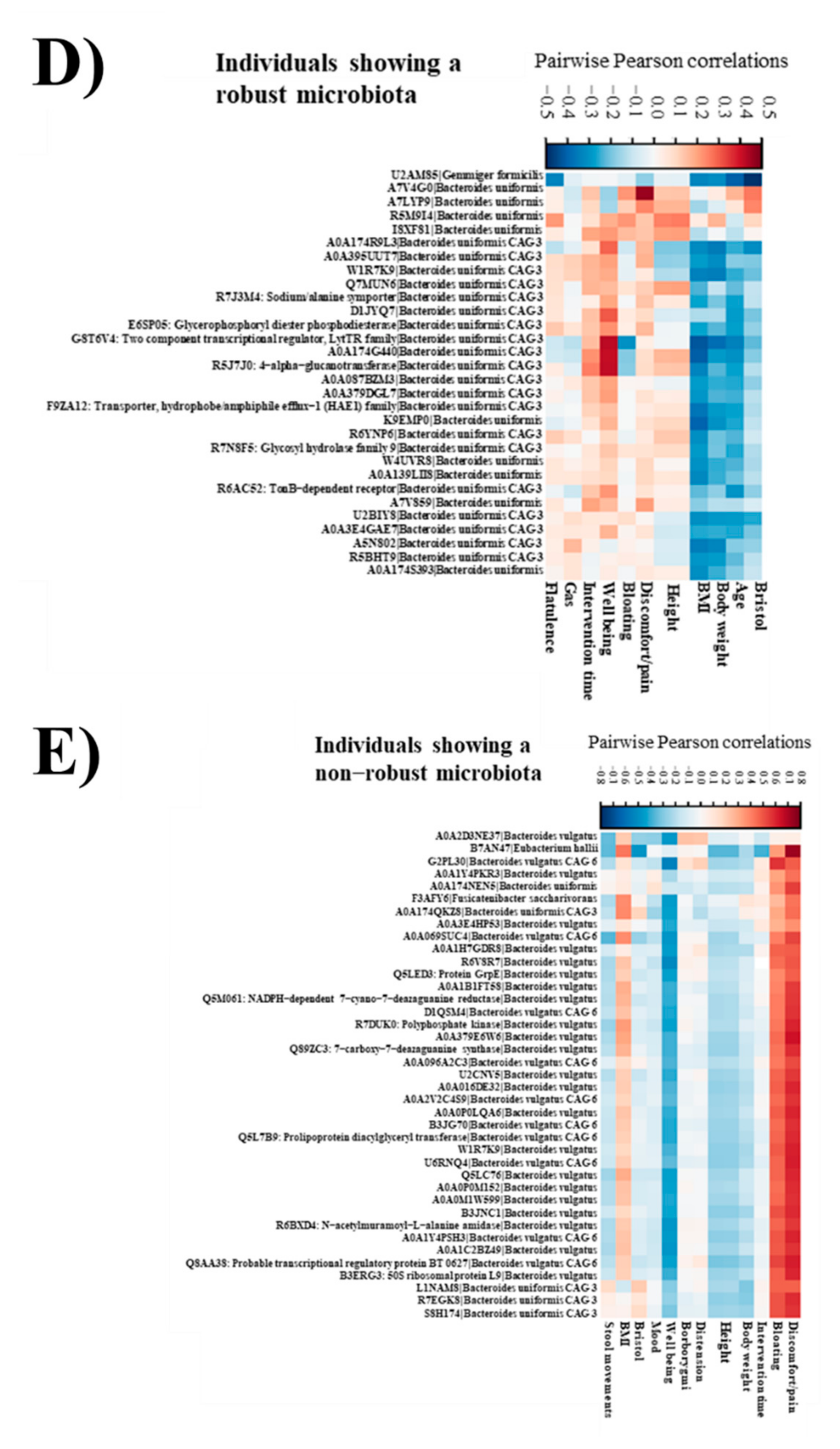
3.10. Robust and Non-Robust Microbiotas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manichanh, C.; Eck, A.; Varela, E.; Roca, J.; Clemente, J.C.; Gonzalez, A.; Knights, D.; Knight, R.; Estrella, S.; Hernandez, S.; et al. Anal gas evacuation and colonic microbiota in patients with flatulence: Effect of diet. Gut 2014, 63, 401–408. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Azpiroz, F.; Hernandez, C.; Guyonnet, D.; Accarino, A.; Santos, J.; Malagelada, J.R.; Guarner, F. Effect of a low-flatulogenic diet in patients with flatulence and functional digestive symptoms. Neurogastroenterol. Motil. 2014, 26, 779–785. [Google Scholar] [CrossRef]
- Barba, E.; Burri, E.; Accarino, A.; Cisternas, D.; Quiroga, S.; Monclus, E.; Navazo, I.; Malagelada, J.-R.; Azpiroz, F. Abdomino-thoracic mechanisms of functional abdominal distension and correction by biofeedback. Gastroenterology 2015, 148, 732–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burri, E.; Barba, E.; Huaman, J.W.; Cisternas, D.; Accarino, A.; Soldevilla, A.; Malagelada, J.-R.; Azpiroz, F. Mechanisms of postprandial abdominal bloating and distension in functional dyspepsia. Gut 2014, 63, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Malagelada, C.; Drozdzal, M.; Segui, S.; Mendez, S.; Vitria, J.; Radeva, P.; Santos, J.; Accarino, A.; Malagelada, J.R.; Azpiroz, F. Classification of functional bowel disorders by objective physiological criteria based on endoluminal image analysis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G413–G419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malagelada, C.; Barba, I.; Accarino, A.; Molne, L.; Mendez, S.; Campos, E.; Gonzalez, A.; Alonso-Cotoner, C.; Santos, J.; Malagelada, J.-R.; et al. Cognitive and hedonic responses to meal ingestion correlate with changes in circulating metabolites. Neurogastroenterol. Motil. 2016, 28, 1806–1814. [Google Scholar] [CrossRef] [PubMed]
- Biesiekierski, J.R.; Rosella, O.; Rose, R.; Liels, K.; Barrett, J.S.; Shepherd, S.J.; Gibson, P.R.; Muir, J.G. Quantification of fructans, galacto-oligosacharides and other short-chain carbohydrates in processed grains and cereals. J. Hum. Nutr. Diet. 2011, 24, 154–176. [Google Scholar] [CrossRef]
- Hospital Clínico Universitario de Santiago de Compostela. Odimet, Organizador Dietético Metabólico; Hospital Clínico Universitario de Santiago de Compostela: Santiago de Compostela, Spain, 2014. [Google Scholar]
- Malagelada, C.; Accarino, A.; Molne, L.; Mendez, S.; Campos, E.; Gonzalez, A.; Malagelada, J.-R.; Azpiroz, F. Digestive, cognitive and hedonic responses to a meal. Neurogastroenterol. Motil. 2015, 27, 389–396. [Google Scholar] [CrossRef]
- Muir, J.G.; Rose, R.; Rosella, O.; Liels, K.; Barrett, J.S.; Shepherd, S.J.; Gibson, P.R. Measurement of short-chain carbohydrates in common Australian vegetables and fruits by high-performance liquid chromatography (HPLC). J. Agric. Food Chem. 2009, 57, 554–565. [Google Scholar] [CrossRef]
- Serra, J.; Azpiroz, F.; Malagelada, J.R. Gastric distension and duodenal lipid infusion modulate intestinal gas transit and tolerance in humans. Am. J. Gastroenterol. 2002, 97, 2225–2230. [Google Scholar] [CrossRef]
- Serra, J.; Azpiroz, F.; Malagelada, J.R. Mechanisms of intestinal gas retention in humans:impaired propulsion versus obstructed evacuation. Am. J. Physiol. 2001, 281, G138–G143. [Google Scholar]
- Serra, J.; Azpiroz, F.; Malagelada, J.R. Intestinal gas dynamics and tolerance in humans. Gastroenterology 1998, 115, 542–550. [Google Scholar] [CrossRef]
- Cardona, S.; Eck, A.; Cassellas, M.; Gallart, M.; Alastrue, C.; Dore, J.; Azpiroz, F.; Roca, J.; Guarner, F.; Manichanh, C. Storage conditions of intestinal microbiota matter in metagenomic analysis. BMC Microbiol. 2012, 12, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, J.; Salvioli, B.; Azpiroz, F.; Malagelada, J.R. Lipid-induced intestinal gas retention in the irritable bowel syndrome. Gastroenterology 2002, 123, 700–706. [Google Scholar] [CrossRef]
- Hernando-Harder, A.C.; Serra, J.; Azpiroz, F.; Mila, M.; Aguade, S.; Malagelada, C.; Tremolaterra, F.; Villoria, A.; Malagelada, J.-R. Colonic responses to gas loads in subgroups of patients with abdominal bloating. Am. J. Gastroenterol. 2010, 105, 876–882. [Google Scholar] [CrossRef]
- Pribic, T.; Nieto, A.; Hernandez, L.; Malagelada, C.; Accarino, A.; Azpiroz, F. Appetite influences the responses to meal ingestion. Neurogastroenterol. Motil. 2017, 29, e13072. [Google Scholar] [CrossRef] [PubMed]
- Pribic, T.; Kilpatrick, L.; Ciccantelli, B.; Malagelada, C.; Accarino, A.; Rovira, A.; Pareto, D.; Mayer, E.; Azpiroz, F. Brain networks associated with cognitive and hedonic responses to a meal. Neurogastroenterol. Motil. 2017, 29, e13031. [Google Scholar] [CrossRef] [PubMed]
- Halmos, E.P.; Power, V.A.; Shepherd, S.J.; Gibson, P.R.; Muir, J.G. A diet low in FODMAPs reduces symptoms of irritable bowel syndrome. Gastroenterology 2014, 146, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Bendezu, R.A.; Mego, M.; Monclus, E.; Merino, X.; Accarino, A.; Malagelada, J.R.; Navazo, I.; Azpiroz, F. Colonic content: Effect of diet, meals, and defecation. Neurogastroenterol. Motil. 2017, 29, e12930. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzosa, E.A.; McIver, L.J.; Rahnavard, G.; Thompson, L.R.; Schirmer, M.; Weingart, G.; Schwarzberg, K.; Knight, R.; Caporaso, J.G.; Segata, N. Species-level functional profiling of metagenomes and metatranscriptomes. Nat. Methods 2018, 15, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Truong, D.T.; Tett, A.; Pasolli, E.; Huttenhower, C.; Segata, N. Microbial strain-level population structure and genetic diversity from metagenomes. Genome Res. 2017, 27, 626–638. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Lahti, L.; Shetty, S. Tools for Microbiome Analysis in R. Version 2.1.24. 2017. Available online: http://microbiome.github.com/microbiome (accessed on 20 May 2021).
- Avila-Galvez, M.A.; Garcia-Villalba, R.; Martinez-Diaz, F.; Ocana-Castillo, B.; Monedero-Saiz, T.; Torrecillas-Sanchez, A.; Abellan, B.; Gonzales-Sarrias, A.; Espin, J.C. Metabolic profiling of dietary polyphenols and methylxanthines in normal and malignant mammary tissues from breast cancer patients. Mol. Nutr. Food Res. 2019, 63, e1801239. [Google Scholar] [CrossRef]
- Vallejo, F.; Yuste, J.E.; Teruel-Montoya, R.; Luengo-Gil, G.; Bohdan, N.; Espin, S.; Garcia-Barbera, N.; Martinez, V.V.; Espin, J.C.; Martinez-Martinez, I. First exploratory study on the metabolome from plasma exosomes in patients with paroxysmal nocturnal hemoglobinuria. Thromb. Res. 2019, 183, 80–85. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.A. mixOmics: An R package for ’omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azpiroz, F. Intestinal gas. In Sleisenger and Fordtran’s Gastrointestinal and Liver Disease: Pathophysiology, Diagnosis, Management; Feldman, M.F.L., Brand, L.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; p. 224-e31. [Google Scholar]
- Litvak, Y.; Byndloss, M.X.; Baumler, A.J. Colonocyte metabolism shapes the gut microbiota. Science 2018, 362, eaat9076. [Google Scholar] [CrossRef] [Green Version]
- Lundman, P.; Boquist, S.; Samnegard, A.; Bennermo, M.; Held, C.; Ericsson, C.G.; Silveira, A.; Hamsten, A.; Tornvall, P. A high-fat meal is accompanied by increased plasma interleukin-6 concentrations. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Laugerette, F.; Vors, C.; Geloen, A.; Chauvin, M.A.; Soulage, C.; Lambert-Porcheron, S.; Peretti, N.; Alligier, M.; Burcelin, R.; Laville, M.; et al. Emulsified lipids increase endotoxemia: Possible role in early postprandial low-grade inflammation. J. Nutr. Biochem. 2011, 22, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Zeisel SCholine In: Ross, A.C.C.B.; Cousins, R.J.; Tucker, K.L.; Ziegler, T.R. (Eds.) Modern Nutrition in Health and Disease, 11th ed.; Lippincott Williams & Wilkins: Baltimore, MD, USA, 2014; pp. 416–426. [Google Scholar]
- Trottier, J.; Perreault, M.; Rudkowska, I.; Levy, C.; Dallaire-Theroux, A.; Verreault, M.; Caron, P.; Staels, B.; Vohl, M.-C.; Straka, R.J.; et al. Profiling serum bile acid glucuronides in humans: Gender divergences, genetic determinants, and response to fenofibrate. Clin. Pharmacol. Ther. 2013, 94, 533–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandeputte, D.; Kathagen, G.; D’Hoe, K.; Vieira-Silva, S.; Valles-Colomer, M.; Sabino, J.; Wang, J.; Tito, R.Y.; De Commer, L.; Sabino, J.; et al. Quantitative microbiome profiling links gut community variation to microbial load. Nature 2017, 551, 507–511. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barber, C.; Mego, M.; Sabater, C.; Vallejo, F.; Bendezu, R.A.; Masihy, M.; Guarner, F.; Espín, J.C.; Margolles, A.; Azpiroz, F. Differential Effects of Western and Mediterranean-Type Diets on Gut Microbiota: A Metagenomics and Metabolomics Approach. Nutrients 2021, 13, 2638. https://doi.org/10.3390/nu13082638
Barber C, Mego M, Sabater C, Vallejo F, Bendezu RA, Masihy M, Guarner F, Espín JC, Margolles A, Azpiroz F. Differential Effects of Western and Mediterranean-Type Diets on Gut Microbiota: A Metagenomics and Metabolomics Approach. Nutrients. 2021; 13(8):2638. https://doi.org/10.3390/nu13082638
Chicago/Turabian StyleBarber, Claudia, Marianela Mego, Carlos Sabater, Fernando Vallejo, Rogger Alvaro Bendezu, Marcela Masihy, Francisco Guarner, Juan Carlos Espín, Abelardo Margolles, and Fernando Azpiroz. 2021. "Differential Effects of Western and Mediterranean-Type Diets on Gut Microbiota: A Metagenomics and Metabolomics Approach" Nutrients 13, no. 8: 2638. https://doi.org/10.3390/nu13082638
APA StyleBarber, C., Mego, M., Sabater, C., Vallejo, F., Bendezu, R. A., Masihy, M., Guarner, F., Espín, J. C., Margolles, A., & Azpiroz, F. (2021). Differential Effects of Western and Mediterranean-Type Diets on Gut Microbiota: A Metagenomics and Metabolomics Approach. Nutrients, 13(8), 2638. https://doi.org/10.3390/nu13082638