
Anti-Obesity and Hypocholesterolemic Actions of Protamine-Derived Peptide RPR (Arg-Pro-Arg) and Protamine in High-Fat Diet-Induced C57BL/6J Mice



Abstract
1. Introduction
2. Materials and Methods
2.1. In Silico Digestion
2.2. Animals and Diets
2.3. Biochemical Analyses
2.4. RNA Preparation from Mouse Tissues and Real-Time PCR
2.5. Statistical Analyses
3. Results
3.1. In Silico Analysis
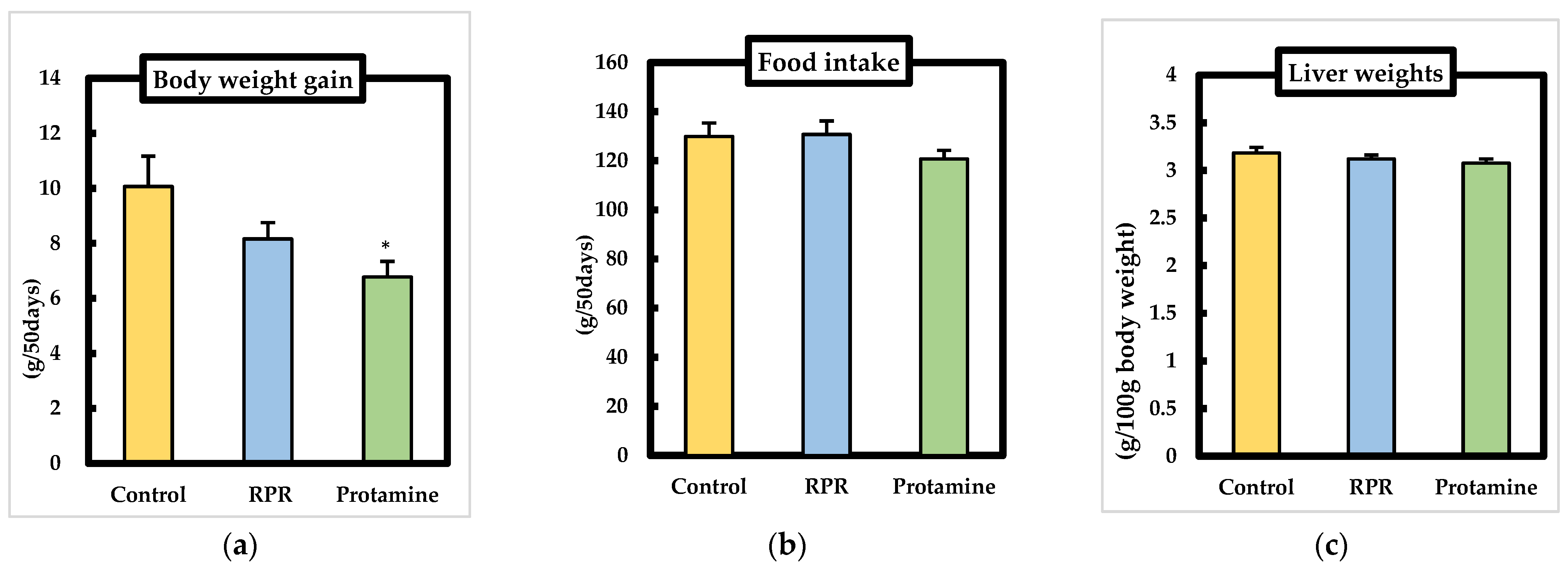
3.2. Body Weights, Food Intake, Liver Weights
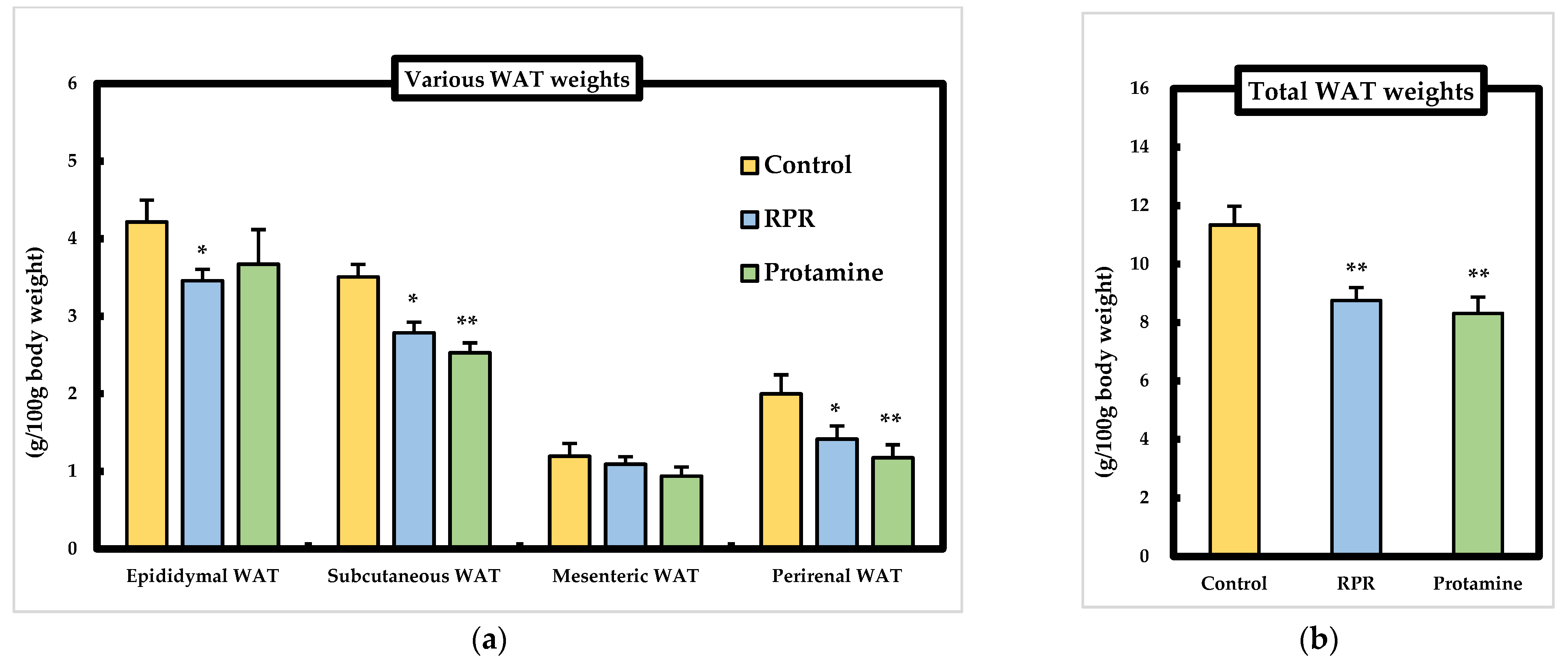
3.3. WAT Weights
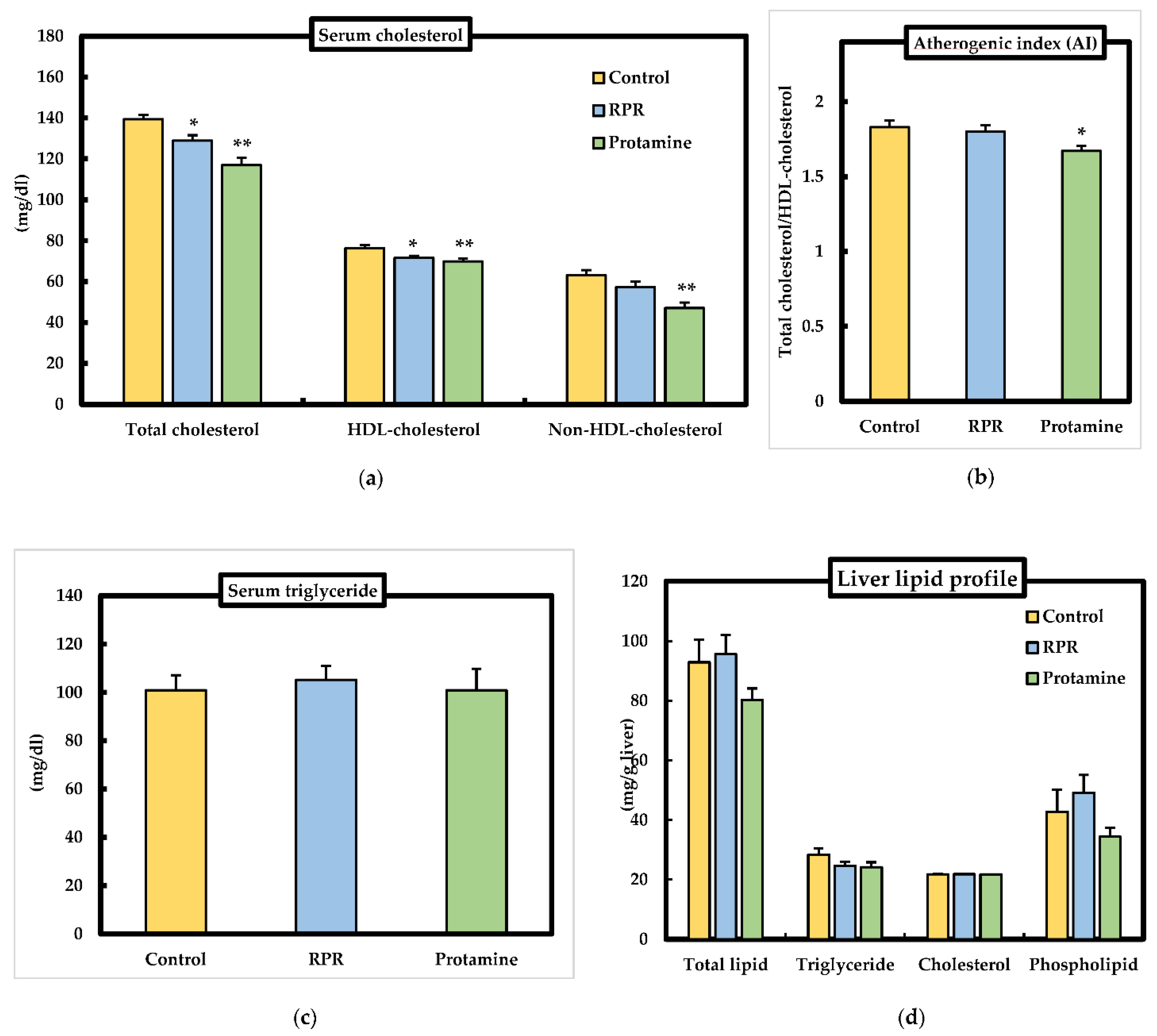
3.4. Serum and Liver Lipid Parameters
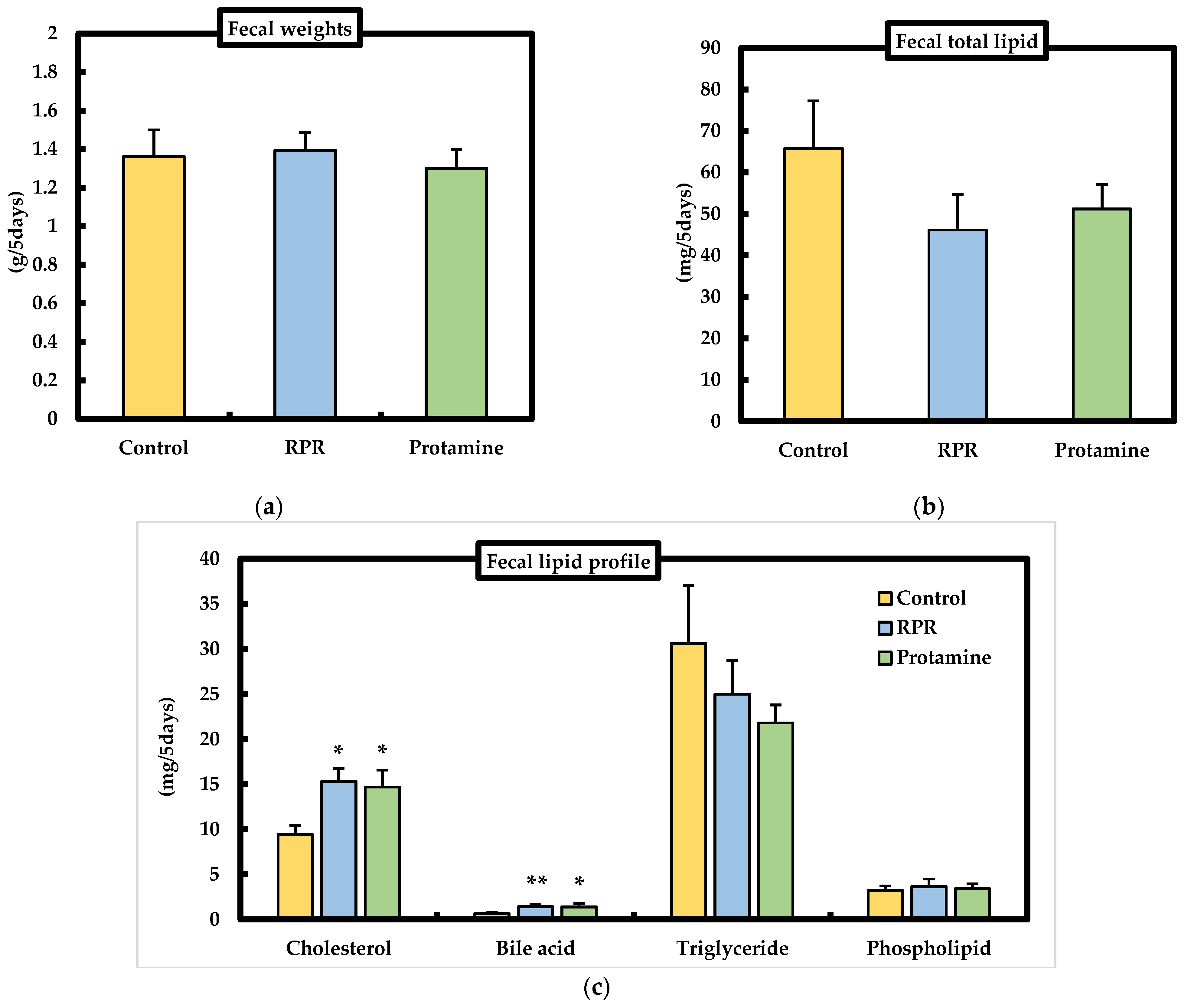
3.5. Fecal Weights and Fecal Lipid Parameters
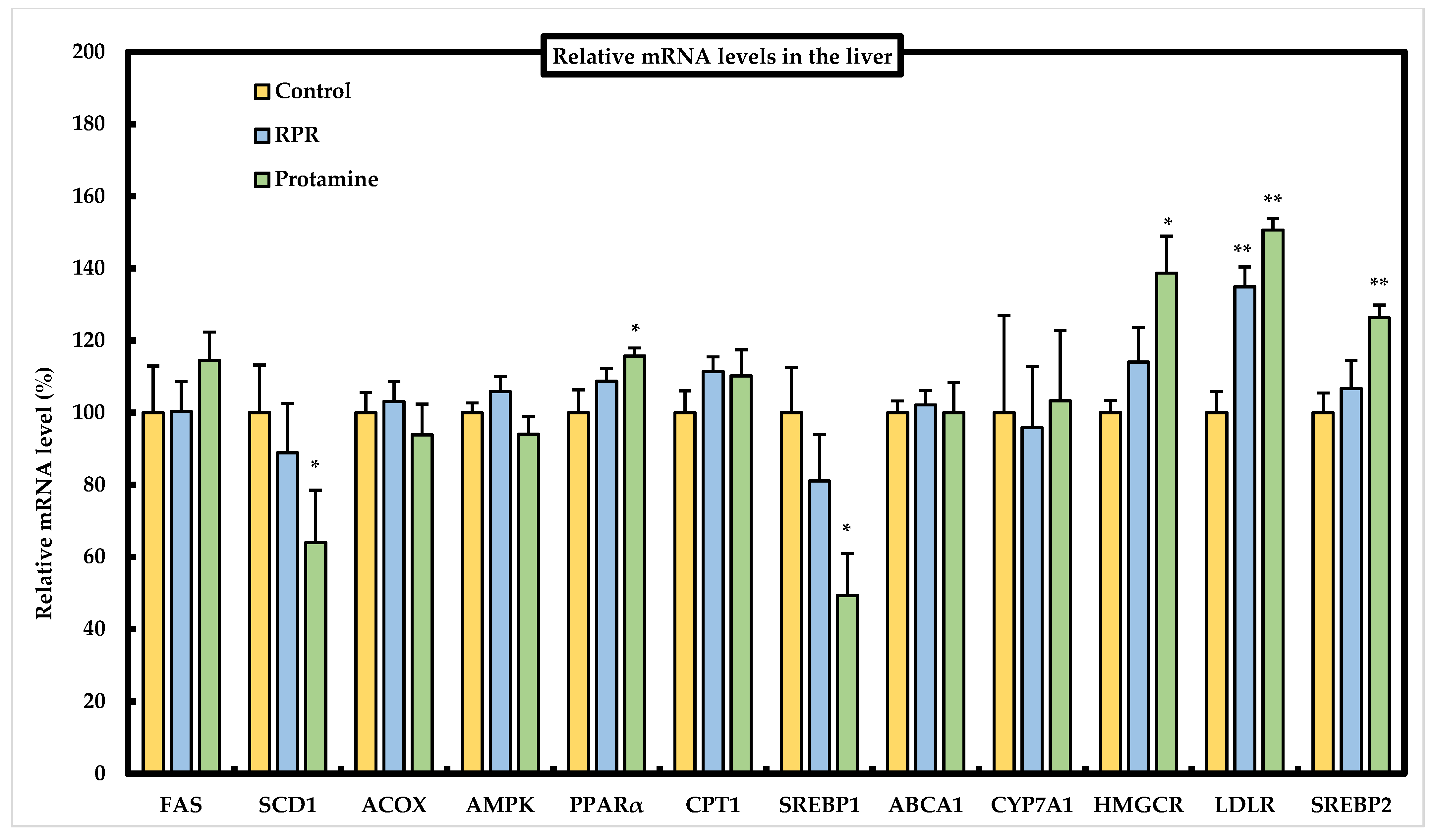
3.6. Effect of RPR or Protamine on the mRNA Levels of Lipid Metabolism-Related Genes in the Liver of Mice
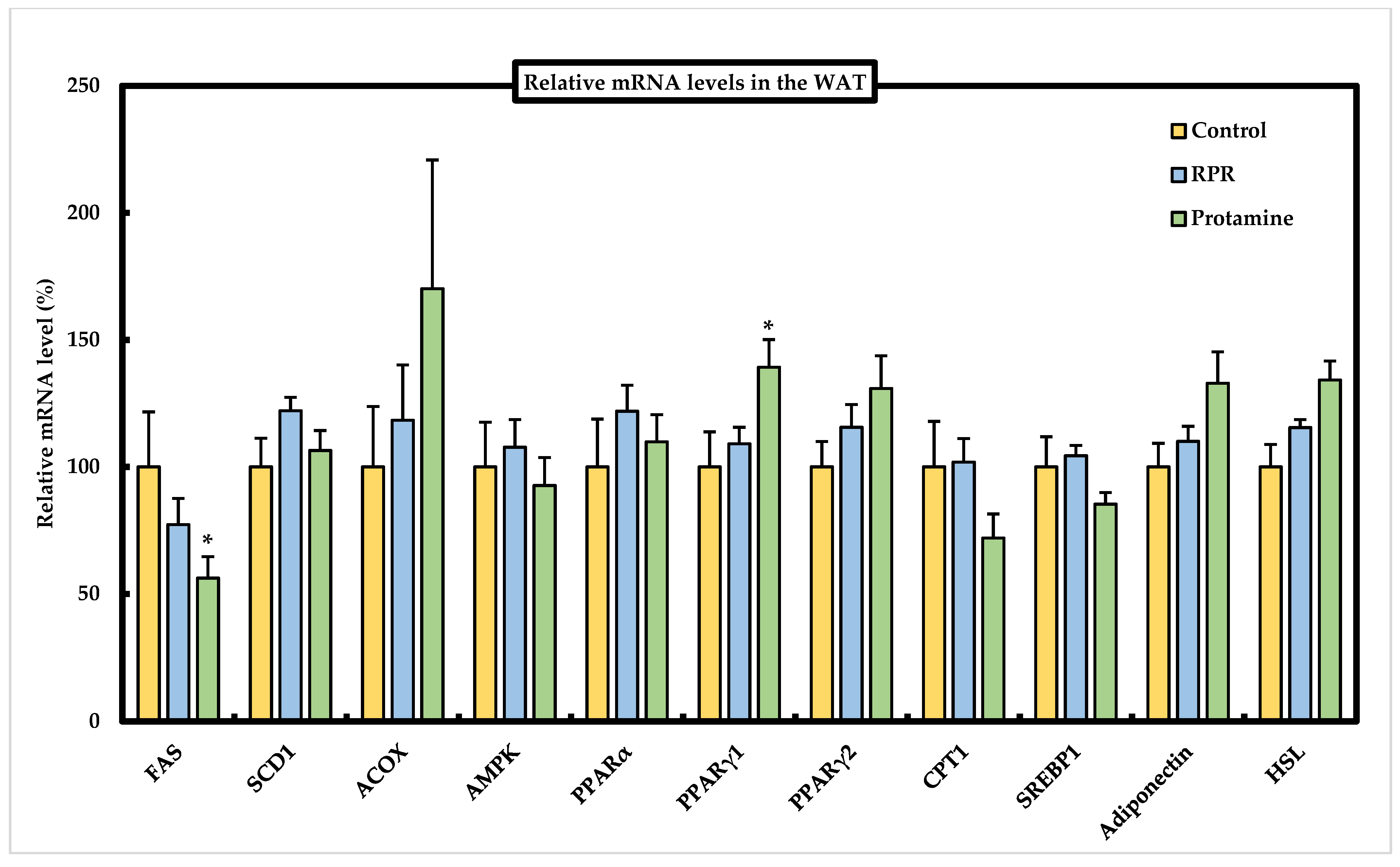
3.7. Effect of RPR or Protamine on the mRNA Levels of Lipid Metabolism-Related Genes in Epididymal WAT of Mice
4. Discussion
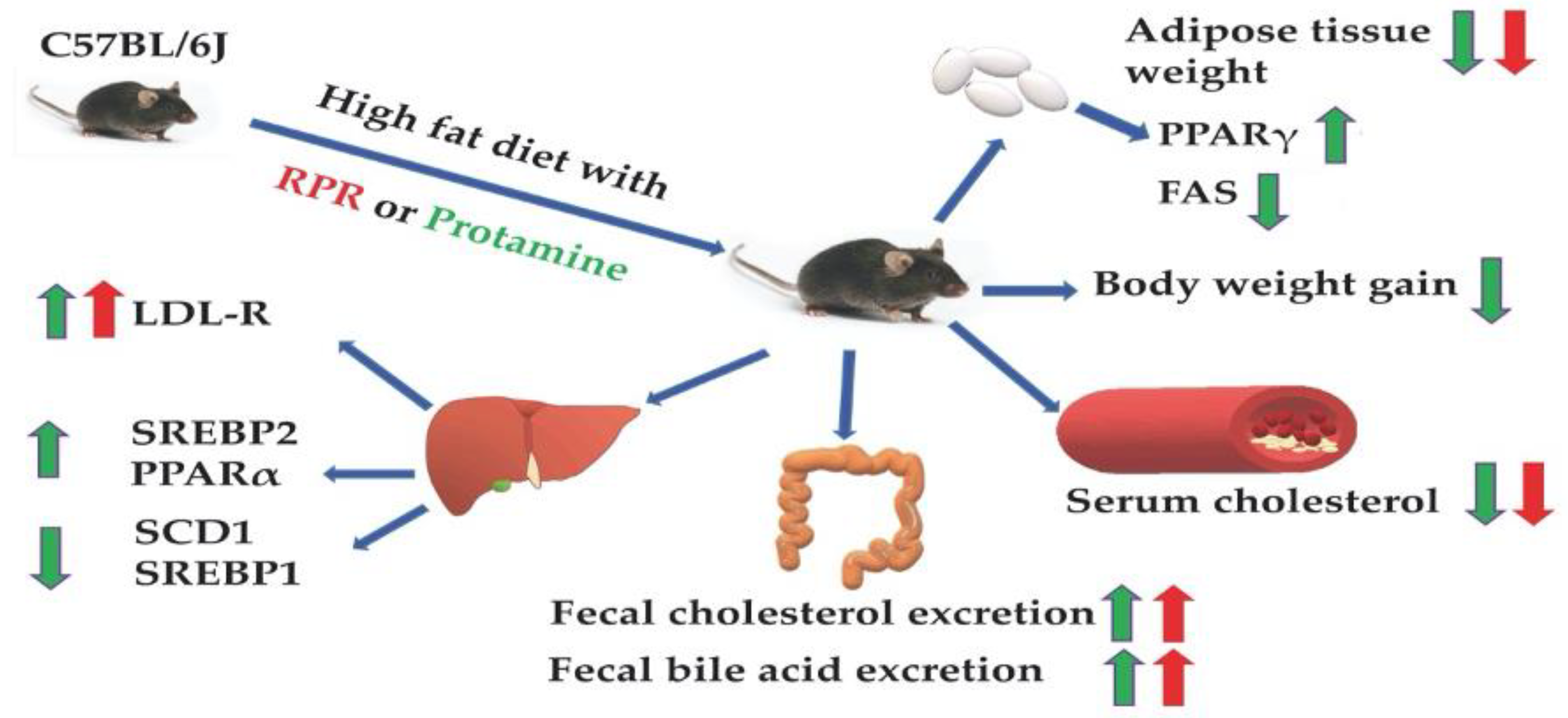
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.A.; Cho, Y.; Hong, S.S.; Ahn, E. Anti-obesity activity of saringosterol isolated from Sargassum muticum (Yendo) Fensholt extract in 3T3-L1 cells. Phytother. Res. 2017, 31, 1694–1701. [Google Scholar] [CrossRef] [PubMed]
- Sevilla-González, M.R.; Quintana-Mendoza, B.M.; Aguilar-Salinas, C.A. Interaction between depression, obesity, and type 2 diabetes: A complex picture. Arch. Med. Res. 2017, 48, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Nurdiantami, Y.; Watanabe, K.; Tanaka, E.; Pradono, J.; Anme, T. Association of general and central obesity with hypertension. Clin. Nutr. 2018, 37, 1259–1263. [Google Scholar] [CrossRef] [PubMed]
- Druso, J.E.; Fischbach, C. Biophysical properties of extracellular matrix: Linking obesity and cancer. Trends Cancer 2018, 4, 271–273. [Google Scholar] [CrossRef]
- Allison, D.B.; Gadbury, G.; Schwartz, L.G.; Murugesan, R.; Kraker, J.L.; Heshka, S.; Fontaine, K.R.; Heymsfield, S.B. A novel soy-based meal replacement formula for weight loss among obese individuals: A randomized controlled clinical trial. Eur. J. Clin. Nutr. 2003, 57, 514–522. [Google Scholar] [CrossRef]
- Velasquez, M.T.; Bhathena, S.J. Role of dietary soy protein in obesity. Int. J. Med. Sci. 2007, 4, 72–82. [Google Scholar] [CrossRef]
- Drotningsvik, A.; Mjøs, S.A.; Pampanin, D.M.; Slizyte, R.; Carvajal, A.; Remman, T.; Høgøy, I.; Gudbrandsen, O.A. Dietary fish protein hydrolysates containing bioactive motifs affect serum and adipose tissue fatty acid compositions, serum lipids, postprandial glucose regulation and growth in obese Zucker fa/fa rats. Br. J. Nutr. 2016, 116, 1336–1345. [Google Scholar] [CrossRef]
- Kim, M.R.; Kim, J.W.; Park, J.B.; Hong, Y.K.; Ku, S.K.; Choi, J.S. Anti-obesity effects of yellow catfish protein hydrolysate on mice fed a 45% kcal high-fat diet. Int. J. Mol. Med. 2017, 40, 784–800. [Google Scholar] [CrossRef]
- Ando, T.; Yamasaki, M.; Suzuki, K. Protamines: Isolation, characterization, structure and function. Mol. Biol. Biochem. Biophys. 1973, 12, 1–114. [Google Scholar]
- Jobgen, W.; Cynthia, J.M.; Scott, C.J.; Li, P.; Lee, M.J.; Stephen, B.S.; Thomas, E.S.; Susan, K.F.; Wu, G.Y. Dietary L-arginine supplementation reduces white fat gain and enhances skeletal muscle and brown fat masses in diet-induced obese rats. J. Nutr. 2009, 139, 230–237. [Google Scholar] [CrossRef]
- Tsujita, T.; Matsuura, Y.; Okuda, H. Studies on the inhibition of pancreatic and carboxylester lipases by protamine. J. Lipid Res. 1996, 37, 1481–1487. [Google Scholar] [CrossRef]
- Nam, H.K.; Won, K.L.; Bo, R.Y.; Hye, R.L.; Min, A.P.; Sang, K.P.; Hyoung, K.P.; Kyung, C.C. Risk of cardiovascular disease is suppressed by dietary supplementation with protamine and chitooligosaccharide in sprague-dawley rats. Mol. Med. Rep. 2013, 1, 127–133. [Google Scholar]
- Duarte-Vazquez, M.A.; Garcia-Padilla, S.; Olvera-Ochoa, L.; Gonzalez-Romero, K.E.; Acosta-Iniguez, J.; Cruz-Corodero, R.D.; Rosado, J.L. Effect of protamine in obesity induced by high-fat diets in rats. Int. J. Obes. 2009, 33, 687–692. [Google Scholar] [CrossRef][Green Version]
- Hosomi, R.; Miyauchi, K.; Yamamoto, D.; Arai, H.; Nishiyama, T.; Yoshida, M.; Fukunaga, K. Salmon protamine decreases serum cholesterol and liver lipid contents by inhibiting lipid absorption in an in vitro gastrointestinal digestion model and in rats. J. Food Sci. 2015, 80, 2346–2353. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Konishi, T.; Enari, H.; Lee, H.S.; Lee, W.K.; Lee, Y.S.; Park, H.K.; Yamamoto, S. Effects of protamine hydrochloride from chum salmon milt on lipid metabolism in rats. Fish Sci. 2011, 77, 1045–1052. [Google Scholar] [CrossRef]
- Mori, R.D.; Dixon, G.H. Characterization of a protamine gene from the chum salmon (Oncorhynchus keta). J. Mol. Evol. 1988, 27, 8–16. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Nagaoka, S.; Miwa, K.; Eto, M.; Kuzuya, Y.; Hori, G.; Yamamoto, K. Soy protein peptic hydrolysate with bound phospholipids decrease micellar solubility and cholesterol absorption in rats and Caco-2 cells. J. Nutr. 1999, 129, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Dunnett, C.W. New tables for multiple comparisons with a control. Biometrics 1964, 20, 482–491. [Google Scholar] [CrossRef]
- David, L.T.; Huang, Y.; Vicki, H.; Wysocki-John, R.Y. Influence of basic residue content on fragment ion peak intensities in low-energy collision-induced dissociation spectra of peptides. Anal. Chem. 2004, 76, 1243–1248. [Google Scholar]
- Bergstrom, R.W.; Newell-Morris, L.L.; Leonetti, D.L.; Shuman, W.P.; Wahl, P.W.; Fujimoto, W.Y. Association of elevated fasting C-peptide level and increased intra-abdominal fat distribution with development of NIDDM in Japanese-American men. Diabetes 1990, 39, 104–111. [Google Scholar] [CrossRef]
- Ingelsson, E.; Schaefer, E.J.; Contois, J.H.; McNamara, J.R.; Sullivan, L.; Keyes, M.J.; Pencina, M.J.; Schoonmaker, C.; Wilson, P.W.; D’Agostino, R.B.; et al. Clinical utility of different lipid measures for prediction of coronary heart disease in men and women. JAMA 2007, 298, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Leal-Díaz, A.M.; Noriega, L.G.; Torre-Villalvazo, I.; Torres, N.; Alemán-Escondrillas, G.; López-Romero, P.; Sánchez-Tapia, M.; Aguilar-López, M.; Furuzawa-Carballeda, J.; Velázquez-Villegas, L.A.; et al. Aguamiel concentrate from Agave salmiana and its extracted saponins attenuated obesity and hepatic steatosis and increased Akkermansia muciniphila in C57BL6 mice. Sci. Rep. 2016, 6, 34242. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, I.; Bashmakov, Y.; Ikemoto, S.; Horton, J.D.; Brown, M.S.; Goldstein, J.L. Insulin selectively increase SREBP-1c mRNA in the livers of rats with streptozotocin- induced diabetes. Proc. Natl. Acad. Sci. USA 1999, 96, 13656–13661. [Google Scholar] [CrossRef] [PubMed]
- Osborne, T.F. Sterol regulatory element-binding proteins (SREBPs): Key regulators of nutritional homeostasis and insulin action. J. Biol. Chem. 2000, 275, 32379–32382. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Kim, Y.C.; Ntambi, J.M. A lipogenic diet in mice with a disruption of the stearoyl-CoA desaturase 1 gene reveals a stringent requirement of endogenous monounsaturated fatty acids for triglyceride synthesis. J. Lipid Res. 2001, 42, 1018–1024. [Google Scholar] [CrossRef]
- Lounis, M.A.; Bergeron, K.F.; Burhans, M.S.; Ntambi, J.M.; Mounier, C. Oleate activates SREBP-1 signaling activity in SCD1-deficient hepatocytes. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E710–E720. [Google Scholar] [CrossRef]
- Miyazaki, M.; Dobrzyn, A.; Sampath, H.; Lee, S.H.; Man, W.C.; Chu, K.; Jeffrey, M.P.; Frank, J.G.; James, M.N. Reduced adiposity and liver steatosis by stearoyl-CoA desaturase deficiency are independent of peroxi-some proliferator-activated receptor-alpha. J. Biol. Chem. 2004, 279, 35017–35024. [Google Scholar] [CrossRef]
- Lodhi, I.J.; Yin, L.; Jensen-Urstad, A.P.; Funai, K.; Coleman, T.; Baird, J.H.; El Ramahi, M.K.; Razani, B.; Song, H.; Fu-Hsu, F.; et al. Inhibiting adipose tissue lipogenesis reprograms thermogenesis and PPARγ activation to decrease diet-induced obesity. Cell Metab. 2012, 16, 189–201. [Google Scholar] [CrossRef]
- Kersten, S.; Desvergne, B.; Wahli, W. Roles of PPARs in health and disease. Nature 2000, 405, 421–424. [Google Scholar] [CrossRef]
- Braissant, O.; Foufelle, F.; Scotto, C.; Dauca, M.; Wahli, W. Differential expression of peroxisome proliferator-activated receptors (PPARs): Tissue distribution of PPAR-α, -β, and -γ in the adult rat. Endocrinology 1996, 137, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Iwase, M.; Yamamoto, T.; Nishimura, K.; Takahashi, H.; Mohri, S.; Li, Y.; Jheng, H.F.; Nomura, W.; Takahashi, N.; Kim, C.S.; et al. Suksdorfin promotes adipocyte differentiation and improves abnormalities in glucose metabolism via PPARγ activation. Lipids 2017, 52, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S.; Seydoux, J.; Peters, J.M.; Gonzalez, F.J.; Desvergne, B.; Wahli, W. Peroxisome proliferator-activated receptor alpha mediates the adaptive response to fasting. J. Clin. Investig. 1999, 103, 1489–1498. [Google Scholar] [CrossRef]
- Morikawa, K.; Kondo, I.; Kanamaru, Y.; Nagaoka, S. A novel regulatory pathway for cholesterol degradation via lactostatin. Biochem. Biophys. Res. Commun. 2007, 352, 697–702. [Google Scholar] [CrossRef]
- Takeuchi, A.; Hisamatsu, K.; Okumura, N.; Sugimitsu, Y.; Yanase, E.; Ueno, Y.; Nagaoka, S. IIAEK targets intestinal alkaline phosphatase (IAP) to improve cholesterol metabolism with a specific activation of IAP and downregulation of ABCA1. Nutrients 2020, 12, 2859. [Google Scholar] [CrossRef]
- Banno, A.; Wang, J.; Okada, K.; Mori, R.; Maihemuti, M.; Nagaoka, S. Identification of a novel cholesterol-lowering dipeptide, phenylalanine-proline (FP), and its down-regulation of intestinal ABCA1 in hypercholesterolemic rats and Caco-2 cells. Sci. Rep. 2019, 9, 19416. [Google Scholar] [CrossRef]
- Toyobuku, H.; Sai, Y.; Tamai, I.; Tsuji, A. Enhanced delivery of drugs to the liver by adenovirous-mediated heterologous expression of the human oligopeptide transporter PEPT1. J. Pharmacol. Exp. Ther. 2002, 301, 812–819. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (1) control (Control group) | 25% (w/w) casein | HFD | n = 8 |
| (2) 0.5% RPR (RPR group) | 24.5% (w/w) casein + 0.5% (w/w) RPR * | HFD | n = 8 |
| (3) 5% protamine (Protamine group) | 20% (w/w) casein + 5% (w/w) protamine ** | HFD | n = 8 |
| Gene | Forword Primers (5′➝3′) | Reverse Primers (5′➝3′) | NCBI Gen Bank |
|---|---|---|---|
| ABCA1 | CTTCCCACATTTTTGCCTGG | AAGGTTCCGTCCTACCAAGTCC | NM_013454.3 |
| ACOX | AGCGAGCCAGAGCCCCAG | TCAGGCAGCTCACTCAGG | NM_015729.3 |
| Adiponectin | TACAACCAACCAACAGAATCATTATGACGG | GAAAGCCAGTAAATGTAGAGTCGTTGA | NM_009605.5 |
| AMPK | CATGGCTGAGAAGCAGAAGCAC | CTTAACTGCCACTTTATGGCCTG | NM_178143.2 |
| CPT1 | GGATCTACAATTCCCCTCTGC | GCAAAATAGGTCTGCCGACA | NM_013495.2 |
| CYP7A1 | AGCAACTAAACAACCTGCCAGTACTA | GTCCGGATATTCAAGGATGCA | NM_007824.3 |
| FAS | TTCCAAGACGAAAATGATGC | AATTGTGGGATCAGGAGAGC | NM_007988.3 |
| HMGCR | CCTGGGCCCCACATTCA | GACATGGTGCCAACTCCAATC | NM_001360165.1 |
| HSL | ACCGAGACAGGCCTCAGTGTG | GAATCGGCCACCGGTAAAGAG | NM_001039507.2 |
| LDLR | TGTGAAAATGACTCAGACGAACAA | GGAGATGCACTTGCCATCCT | NM_001252658.1 |
| PPARα | TCAGGGTACCACTACGGAGTTCA | CCGAATAGTTCGCCGAAAGA | NM_001113418.1 |
| PPARγ1 | GGTGAACCACTGATATTCAGGACA | TGTGTCAACCATGGTAATTTCAGT | NM_001127330.2 |
| PPARγ2 | TGAGCACTTCACAAGAAATTACC | TGCGAGTGGTCTTCCATCAC | NM_011146.3 |
| SCD1 | TTCTTGCGATACACTCTGGTGC | CGGGATTGAATGTTCTTGTCGT | NM_009127.4 |
| SREBP1 | CTTTGGCCTCGCTTTTCGG | TGGGTCCAATTAGAGCCATCTC | NM_001313979.1 |
| SREBP2 | GTGCGCTCTCGTTTTACTGAAGT | GTATAGAAGACGGCCTTCACCAA | NM_033218.1 |
| β-actin | TGTCCACCTTCCAGCAGATGT | AGCTCAGTAACAGTCCGCCTAGA | NM_007393.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mijiti, M.; Mori, R.; Huang, B.; Tsukamoto, K.; Kiriyama, K.; Sutoh, K.; Nagaoka, S. Anti-Obesity and Hypocholesterolemic Actions of Protamine-Derived Peptide RPR (Arg-Pro-Arg) and Protamine in High-Fat Diet-Induced C57BL/6J Mice. Nutrients 2021, 13, 2501. https://doi.org/10.3390/nu13082501
Mijiti M, Mori R, Huang B, Tsukamoto K, Kiriyama K, Sutoh K, Nagaoka S. Anti-Obesity and Hypocholesterolemic Actions of Protamine-Derived Peptide RPR (Arg-Pro-Arg) and Protamine in High-Fat Diet-Induced C57BL/6J Mice. Nutrients. 2021; 13(8):2501. https://doi.org/10.3390/nu13082501
Chicago/Turabian StyleMijiti, Maihemuti, Ryosuke Mori, Bingyu Huang, Kenichiro Tsukamoto, Keisuke Kiriyama, Keita Sutoh, and Satoshi Nagaoka. 2021. "Anti-Obesity and Hypocholesterolemic Actions of Protamine-Derived Peptide RPR (Arg-Pro-Arg) and Protamine in High-Fat Diet-Induced C57BL/6J Mice" Nutrients 13, no. 8: 2501. https://doi.org/10.3390/nu13082501
APA StyleMijiti, M., Mori, R., Huang, B., Tsukamoto, K., Kiriyama, K., Sutoh, K., & Nagaoka, S. (2021). Anti-Obesity and Hypocholesterolemic Actions of Protamine-Derived Peptide RPR (Arg-Pro-Arg) and Protamine in High-Fat Diet-Induced C57BL/6J Mice. Nutrients, 13(8), 2501. https://doi.org/10.3390/nu13082501