
Urinary Medium-Chained Acyl-Carnitines Sign High Caloric Intake whereas Short-Chained Acyl-Carnitines Sign High -Protein Diet within a High-Fat, Hypercaloric Diet in a Randomized Crossover Design Dietary Trial


 , , , , , and
, , , , , and 
Abstract
1. Introduction
2. Materials and Methods
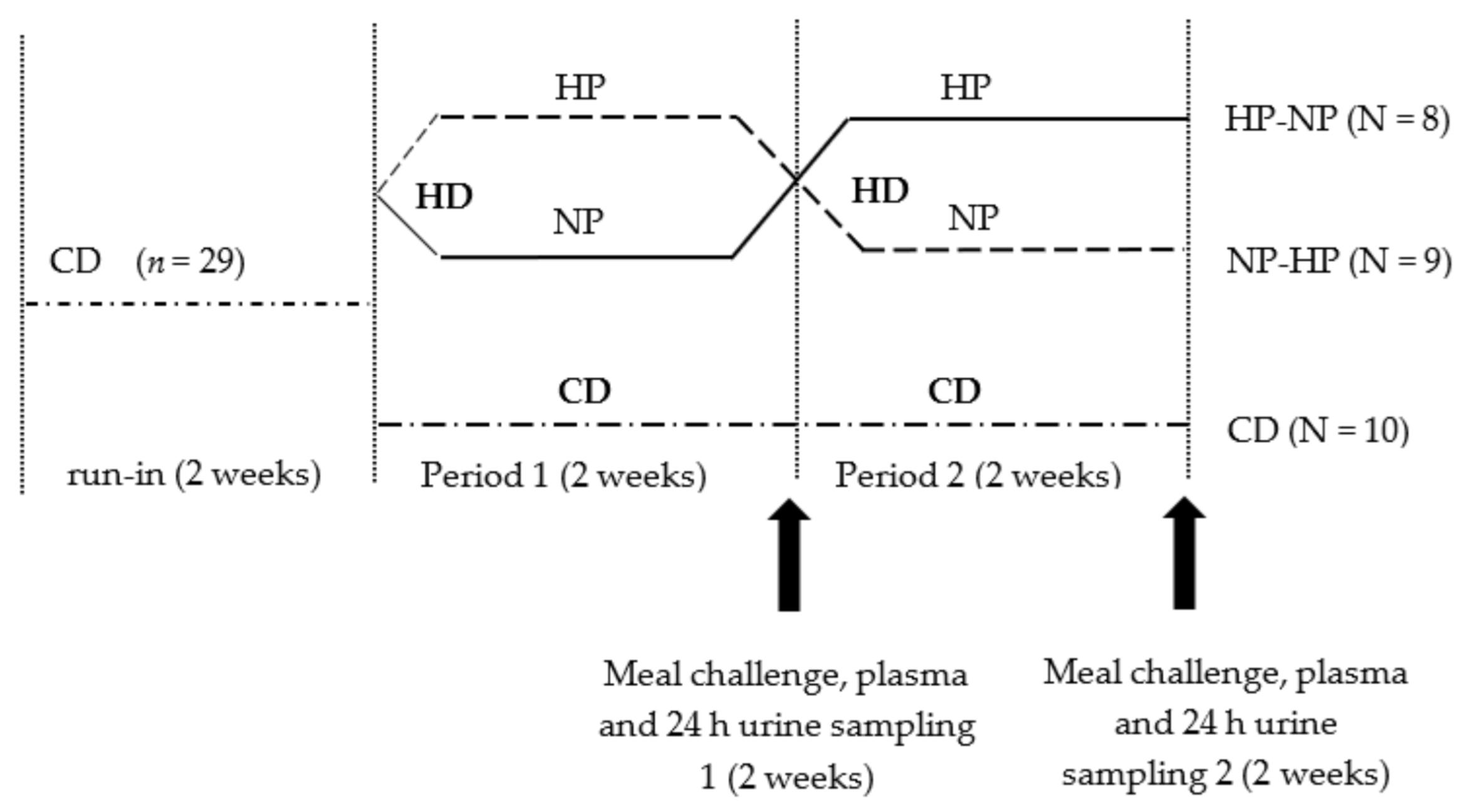
2.1. Participants and Study Design
2.2. Meal Challenge and Urine and Plasma Sampling
2.3. Biochemical Analyses
2.4. 13C Enrichment in Urine
2.5. Metabolomics Sample Preparation
2.6. Liquid Chromatography Mass Spectrometry (LC-MS) Analysis of Urine and Plasma
2.7. LC-MS Data Processing and Analysis
2.8. Statistical Analyses
3. Results
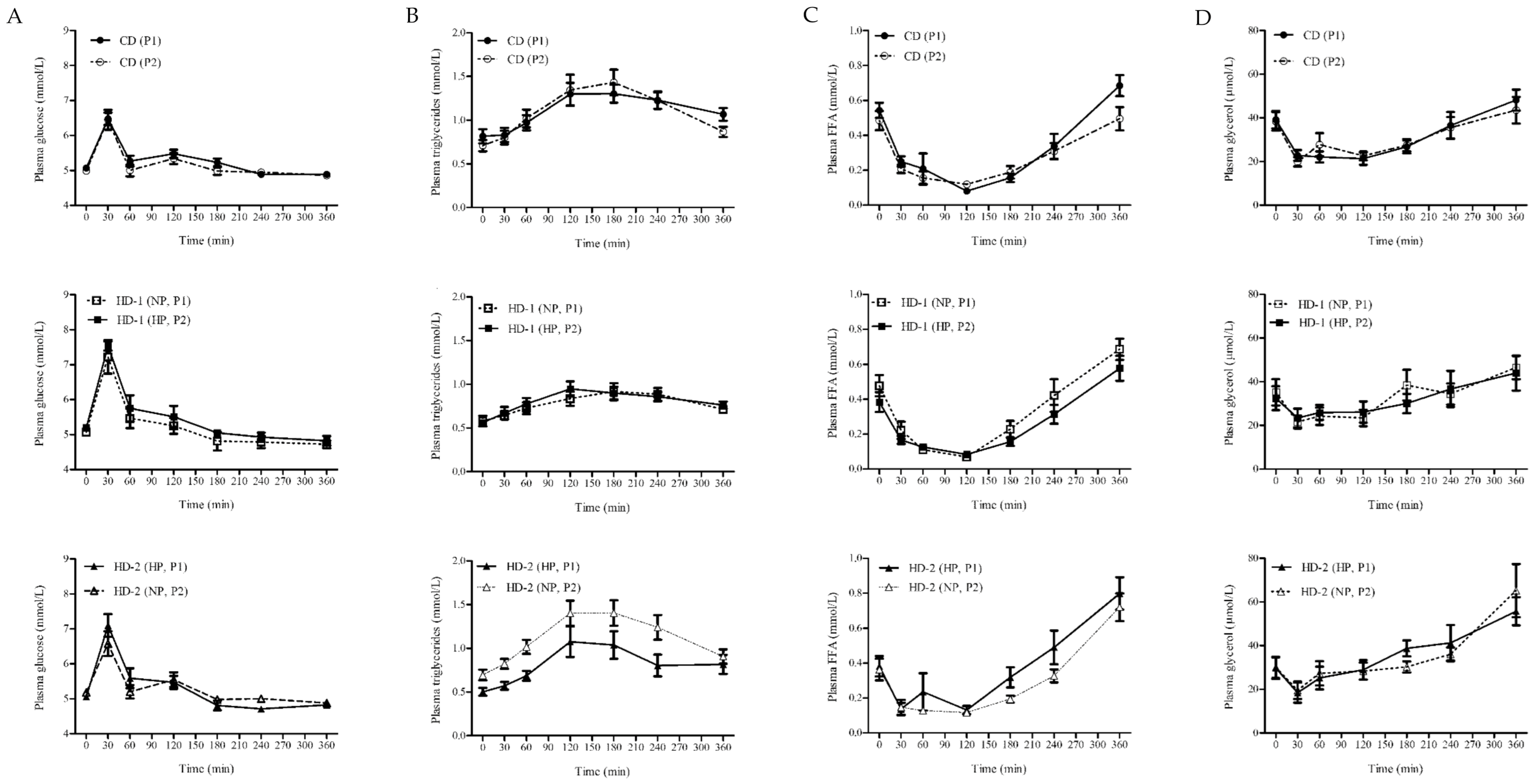
3.1. Fasting Plasma Concentrations of Biomarkers for Lipid and Glucose Metabolism
3.2. Postprandial Blood Biomarkers in Response to a Meal Challenge (MC)
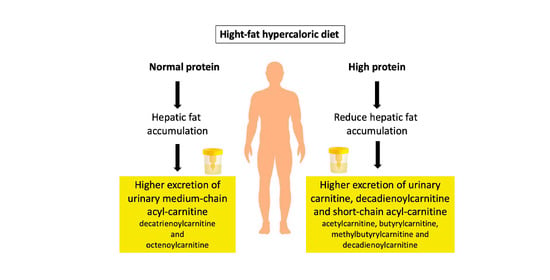
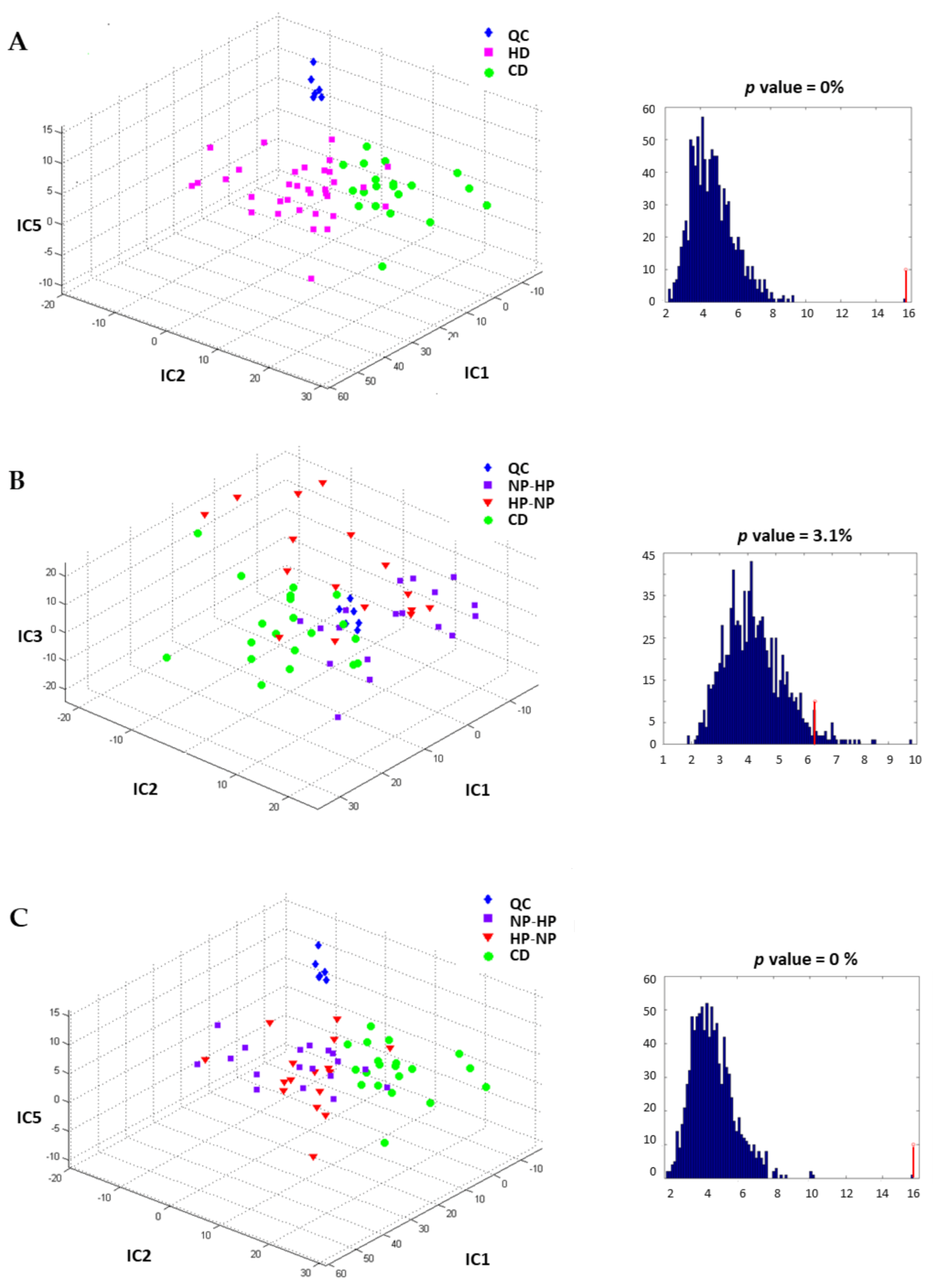
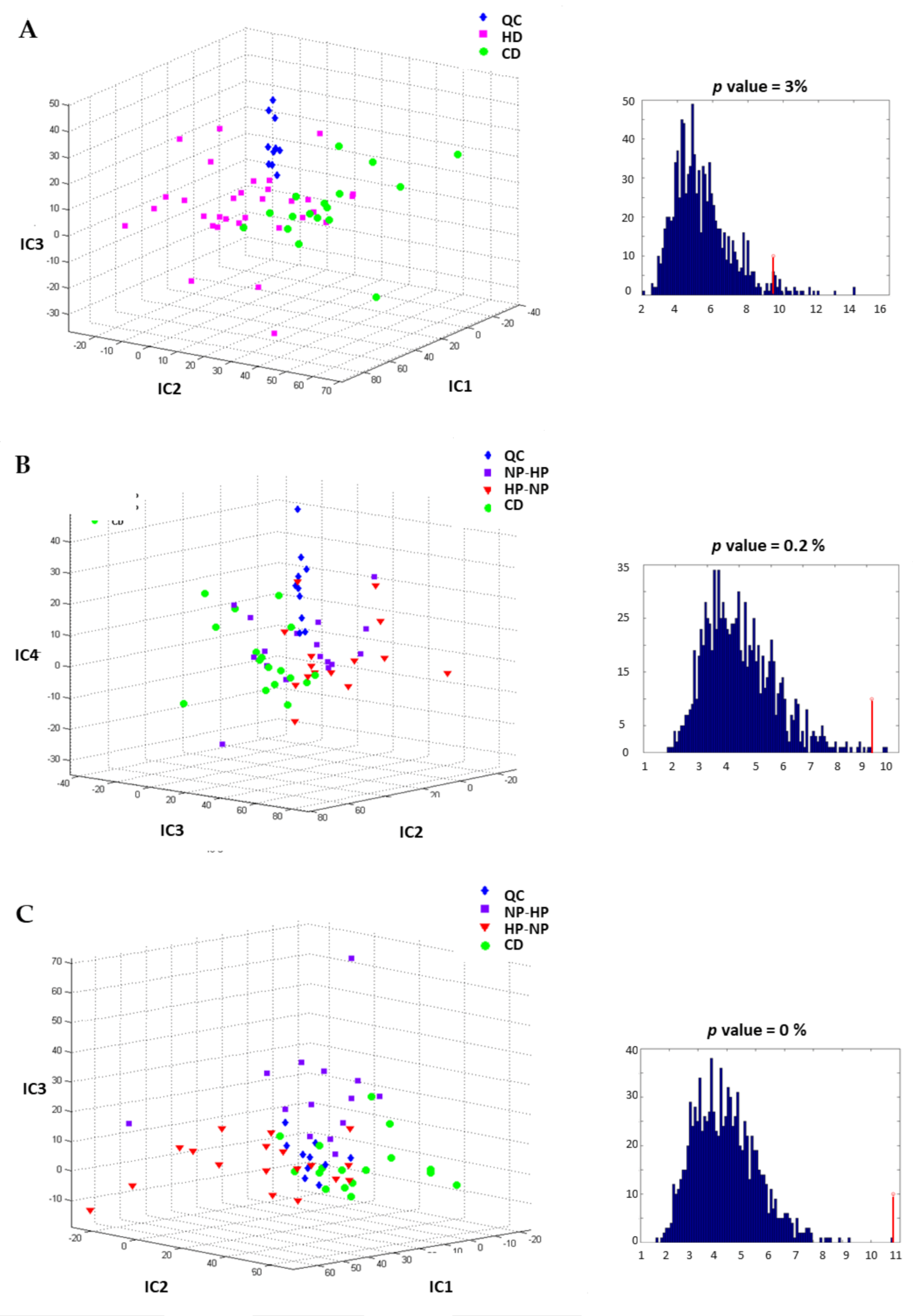
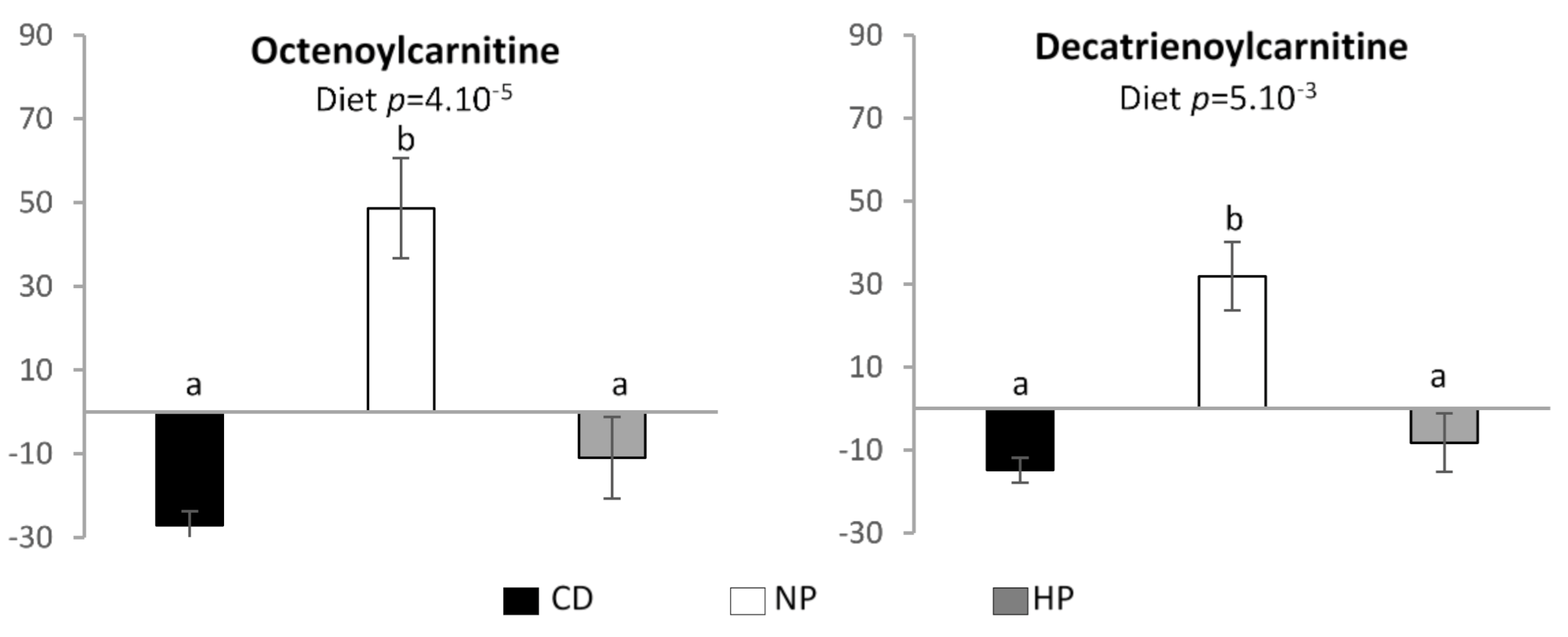
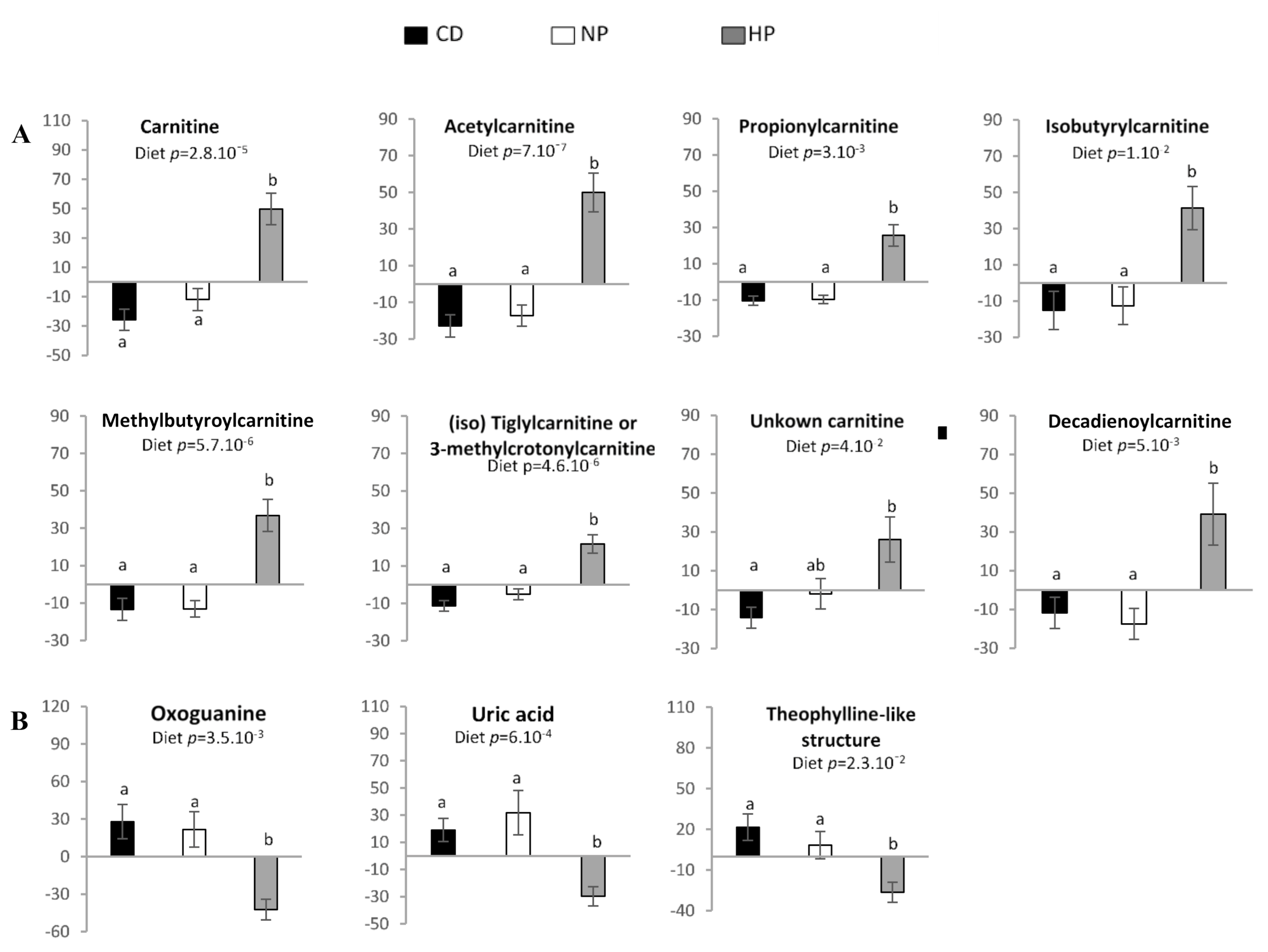
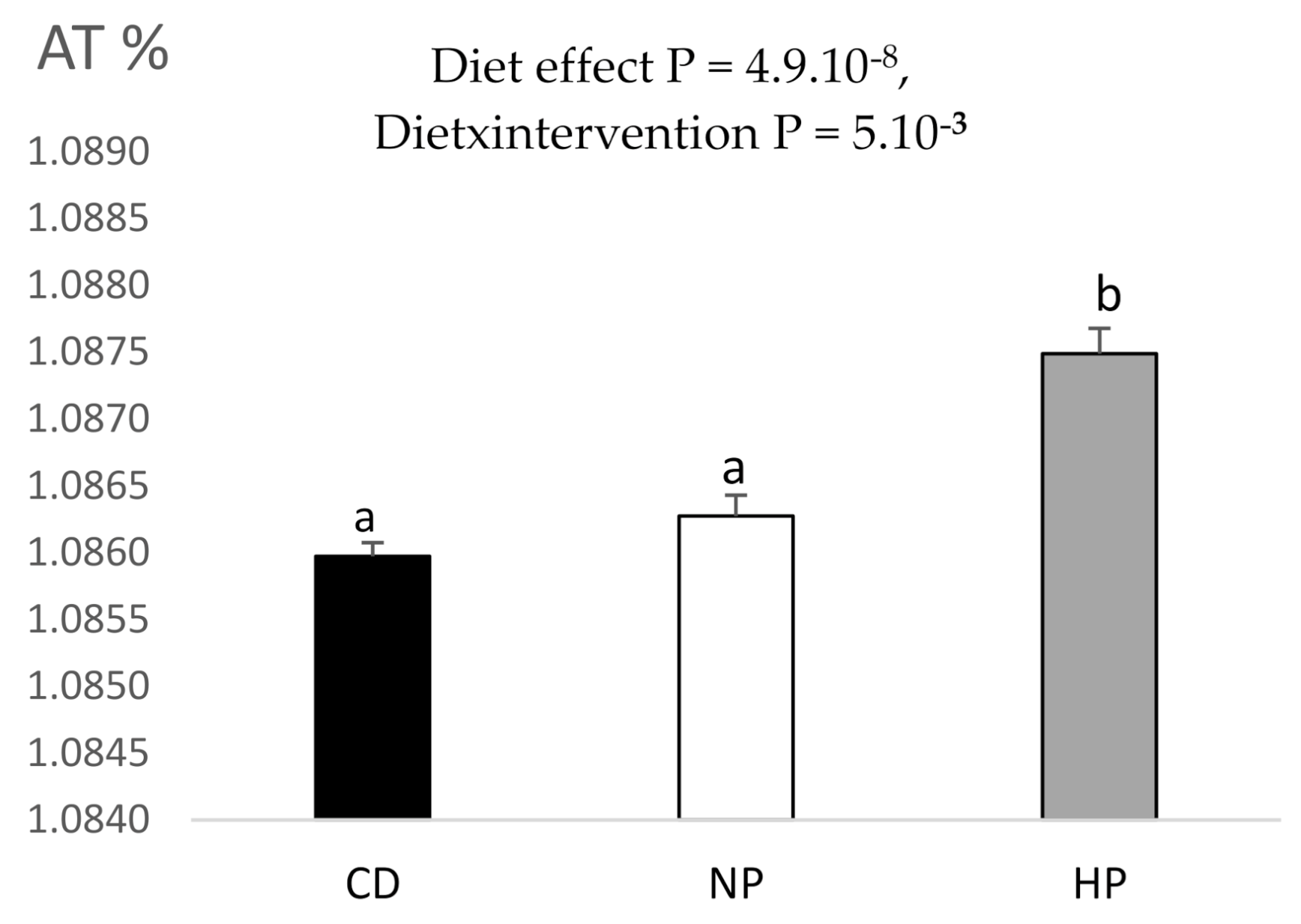
3.3. Urinary and Plasma Metabolites Markers of High Caloric Diet with High or Low Protein Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Parry, S.A.; Hodson, L. Influence of dietary macronutrients on liver fat accumulation and metabolism. J. Investig. Med. Off. Publ. Am. Fed. Clin. Res. 2017, 65, 1102–1115. [Google Scholar] [CrossRef] [PubMed]
- Riazi, K.; Raman, M.; Taylor, L.; Swain, M.G.; Shaheen, A.A. Dietary Patterns and Components in Nonalcoholic Fatty Liver Disease (NAFLD): What Key Messages Can Health Care Providers Offer? Nutrients 2019, 11, 2878. [Google Scholar] [CrossRef] [PubMed]
- DiStefano, J.K. Fructose-mediated effects on gene expression and epigenetic mechanisms associated with NAFLD pathogenesis. Cell Mol. Life Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Joslin, J.; Mularella, J.; Bail, A.; Wojcik, S.; Cooney, D.R. Mandatory Rest Stops Improve Athlete Safety during Event Medical Coverage for Ultramarathons. Prehosp. Disaster Med. 2016, 31, 43–45. [Google Scholar] [CrossRef] [PubMed]
- Rietman, A.; Schwarz, J.; Tome, D.; Kok, F.J.; Mensink, M. High dietary protein intake, reducing or eliciting insulin resistance? Eur. J. Clin. Nutr. 2014, 68, 973–979. [Google Scholar] [CrossRef]
- Van Nielen, M.; Feskens, E.J.; Mensink, M.; Sluijs, I.; Molina, E.; Amiano, P.; Ardanaz, E.; Balkau, B.; Beulens, J.W.; Boeing, H.; et al. Dietary protein intake and incidence of type 2 diabetes in Europe: The EPIC-InterAct Case-Cohort Study. Diabetes Care 2014, 37, 1854–1862. [Google Scholar] [CrossRef]
- Smith, G.I.; Yoshino, J.; Stromsdorfer, K.L.; Klein, S.J.; Magkos, F.; Reeds, D.N.; Klein, S.; Mittendorfer, B. Protein Ingestion Induces Muscle Insulin Resistance Independent of Leucine-Mediated mTOR Activation. Diabetes 2015, 64, 1555–1563. [Google Scholar] [CrossRef]
- Campos-Nonato, I.; Hernandez, L.; Barquera, S. Effect of a High-Protein Diet versus Standard-Protein Diet on Weight Loss and Biomarkers of Metabolic Syndrome: A Randomized Clinical Trial. Obes. Facts 2017, 10, 238–251. [Google Scholar] [CrossRef]
- Markova, M.; Pivovarova, O.; Hornemann, S.; Sucher, S.; Frahnow, T.; Wegner, K.; Machann, J.; Petzke, K.J.; Hierholzer, J.; Lichtinghagen, R.; et al. Isocaloric Diets High in Animal or Plant Protein Reduce Liver Fat and Inflammation in Individuals with Type 2 Diabetes. Gastroenterology 2017, 152, 571–585. [Google Scholar] [CrossRef]
- Bortolotti, M.; Kreis, R.; Debard, C.; Cariou, B.; Faeh, D.; Chetiveaux, M.; Ith, M.; Vermathen, P.; Stefanoni, N.; Le, K.A.; et al. High protein intake reduces intrahepatocellular lipid deposition in humans. Am. J. Clin. Nutr. 2009, 90, 1002–1010. [Google Scholar] [CrossRef]
- Rietman, A.; Schwarz, J.; Blokker, B.A.; Siebelink, E.; Kok, F.J.; Afman, L.A.; Tome, D.; Mensink, M. Increasing protein intake modulates lipid metabolism in healthy young men and women consuming a high-fat hypercaloric diet. J. Nutr. 2014, 144, 1174–1180. [Google Scholar] [CrossRef]
- Bezerra Duarte, S.M.; Faintuch, J.; Stefano, J.T.; Sobral de Oliveira, M.B.; de Campos Mazo, D.F.; Rabelo, F.; Vanni, D.; Nogueira, M.A.; Carrilho, F.J.; Marques Souza de Oliveira, C.P. Hypocaloric high-protein diet improves clinical and biochemical markers in patients with nonalcoholic fatty liver disease (NAFLD). Nutr. Hosp. 2014, 29, 94–101. [Google Scholar] [CrossRef]
- Skov, A.R.; Toubro, S.; Ronn, B.; Holm, L.; Astrup, A. Randomized trial on protein vs carbohydrate in ad libitum fat reduced diet for the treatment of obesity. Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 1999, 23, 528–536. [Google Scholar] [CrossRef]
- Due, A.; Toubro, S.; Skov, A.R.; Astrup, A. Effect of normal-fat diets, either medium or high in protein, on body weight in overweight subjects: A randomised 1-year trial. Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 2004, 28, 1283–1290. [Google Scholar] [CrossRef]
- Green, C.J.; Hodson, L. The influence of dietary fat on liver fat accumulation. Nutrients 2014, 6, 5018–5033. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: Improving the biological information content of metabolomics data. BMC Genom. 2006, 7, 142. [Google Scholar] [CrossRef]
- Fiehn, O.; Robertson, D.; Griffin, J.; Werf, M.; Nikolau, B.; Morrison, N.; Lloyd, W.; Sumner, L.W.; Goodacre, R.; Hardy, N.W.; et al. The metabolomics standards initiative (MSI). Metab. Off. J. Metab. Soc. 2007, 3, 175–178. [Google Scholar] [CrossRef]
- Wang, G.Q.; Ding, Q.Z.; Hou, Z.Y. Independent component analysis and its applications in signal processing for analytical chemistry. TrAC-Trends Anal. Chem. 2008, 27, 368–376. [Google Scholar] [CrossRef]
- Gustafsson, M.G. Independent component analysis yields chemically interpretable latent variables in multivariate regression. J. Chem. Inf. Model. 2005, 45, 1244–1255. [Google Scholar] [CrossRef]
- Habchi, B.; Alves, S.; Jouan-Rimbaud Bouveresse, D.; Moslah, B.; Paris, A.; Lécluse, Y.; Gauduchon, P.; Lebailly, P.; Rutledge, D.N.; Rathahao-Paris, E. An innovative chemometric method for processing direct introduction high resolution mass spectrometry metabolomic data: Independent component–discriminant analysis (IC–DA). Metab. Off. J. Metab. Soc. 2017, 13. [Google Scholar] [CrossRef]
- Khodorova, N.V.; Rutledge, D.N.; Oberli, M.; Mathiron, D.; Marcelo, P.; Benamouzig, R.; Tome, D.; Gaudichon, C.; Pilard, S. Urinary Metabolomics Profiles Associated to Bovine Meat Ingestion in Humans. Mol. Nutr. Food Res. 2019, 63, 1700834. [Google Scholar] [CrossRef]
- Cardoso, J.F. High-order contrasts for independent component analysis. Neural Comput. 1999, 11, 157–192. [Google Scholar] [CrossRef]
- Rutledge, D.N.; Jouan-Rimbaud Bouveresse, D. Independent Components Analysis with the JADE algorithm. TrAC-Trends Anal. Chem. 2013, 50, 22–32. [Google Scholar] [CrossRef]
- Rutledge, D.N.; Jouan-Rimbaud Bouveresse, D. Corrigendum to “Independent Components Analysis with the Jade Algorithm”. TrAC-Trends Anal. Chem. 2015, 67, 220. [Google Scholar] [CrossRef]
- Newsholme, P.; Cruzat, V.; Arfuso, F.; Keane, K. Nutrient regulation of insulin secretion and action. J. Endocrinol. 2014, 221, 105–120. [Google Scholar] [CrossRef]
- Wiklund, S.; Johansson, E.; Sjostrom, L.; Mellerowicz, E.J.; Edlund, U.; Shockcor, J.P.; Gottfries, J.; Moritz, T.; Trygg, J. Visualization of GC/TOF-MS-based metabolomics data for identification of biochemically interesting compounds using OPLS class models. Anal. Chem. 2008, 80, 115–122. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, D.; Anderson, G.H. Recent advances in dietary proteins and lipid metabolism. Curr. Opin. Lipidol. 2013, 24, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Westphal, S.; Kastner, S.; Taneva, E.; Leodolter, A.; Dierkes, J.; Luley, C. Postprandial lipid and carbohydrate responses after the ingestion of a casein-enriched mixed meal. Am. J. Clin. Nutr. 2004, 80, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Brader, L.; Holm, L.; Mortensen, L.; Thomsen, C.; Astrup, A.; Holst, J.J.; De Vrese, M.; Schrezenmeir, J.; Hermansen, K. Acute effects of casein on postprandial lipemia and incretin responses in type 2 diabetic subjects. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 101–109. [Google Scholar] [CrossRef]
- Holmer-Jensen, J.; Mortensen, L.S.; Astrup, A.; De Vrese, M.; Holst, J.J.; Thomsen, C.; Hermansen, K. Acute differential effects of dietary protein quality on postprandial lipemia in obese non-diabetic subjects. Nutr. Res. 2013, 33, 34–40. [Google Scholar] [CrossRef]
- Linn, T.; Santosa, B.; Gronemeyer, D.; Aygen, S.; Scholz, N.; Busch, M.; Bretzel, R.G. Effect of long-term dietary protein intake on glucose metabolism in humans. Diabetologia 2000, 43, 1257–1265. [Google Scholar] [CrossRef]
- Zhao, S.; Wang, J.; Song, X.; Zhang, X.; Ge, C.; Gao, S. Impact of dietary protein on lipid metabolism-related gene expression in porcine adipose tissue. Nutr. Metab. 2010, 7, 6. [Google Scholar] [CrossRef]
- Fu, Z.; Gilbert, E.R.; Liu, D. Regulation of insulin synthesis and secretion and pancreatic Beta-cell dysfunction in diabetes. Curr. Diabetes Rev. 2013, 9, 25–53. [Google Scholar] [CrossRef]
- Wang, H.; Eckel, R.H. Lipoprotein lipase: From gene to obesity. Am. J. Physiol. Endocrinol. Metab. 2009, 297, 271–288. [Google Scholar] [CrossRef]
- Schwarz, J.; Tome, D.; Baars, A.; Hooiveld, G.J.; Muller, M. Dietary protein affects gene expression and prevents lipid accumulation in the liver in mice. PLoS ONE 2012, 7, 47303. [Google Scholar] [CrossRef]
- Stadler, D.D.; Chenard, C.A.; Rebouche, C.J. Effect of dietary macronutrient content on carnitine excretion and efficiency of carnitine reabsorption. Am. J. Clin. Nutr. 1993, 58, 868–872. [Google Scholar] [CrossRef]
- Lake, A.D.; Novak, P.; Shipkova, P.; Aranibar, N.; Robertson, D.G.; Reily, M.D.; Lehman-McKeeman, L.D.; Vaillancourt, R.R.; Cherrington, N.J. Branched chain amino acid metabolism profiles in progressive human nonalcoholic fatty liver disease. Amino acids 2015, 47, 603–615. [Google Scholar] [CrossRef]
- Kalhan, S.C.; Guo, L.; Edmison, J.; Dasarathy, S.; McCullough, A.J.; Hanson, R.W.; Milburn, M. Plasma metabolomic profile in nonalcoholic fatty liver disease. Metab. Clin. Exp. 2011, 60, 404–413. [Google Scholar] [CrossRef]
- Azzout-Marniche, D.; Gaudichon, C.; Tome, D. Dietary protein and blood glucose control. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 349–354. [Google Scholar] [CrossRef]
- Odle, J.; Adams, S.H.; Vockley, J. Carnitine. Adv. Nutr. 2014, 5, 289–290. [Google Scholar] [CrossRef]
- Bieber, L.L.; Emaus, R.; Valkner, K.; Farrell, S. Possible functions of short-chain and medium-chain carnitine acyltransferases. Fed. Proc. 1982, 41, 2858–2862. [Google Scholar]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef]
- O’Brien, D.M. Stable Isotope Ratios as Biomarkers of Diet for Health Research. Annu. Rev. Nutr. 2015, 35, 565–594. [Google Scholar] [CrossRef]
- Tea, I.; Martineau, E.; Antheaume, I.; Lalande, J.; Mauve, C.; Gilard, F.; Barille-Nion, S.; Blackburn, A.C.; Tcherkez, G. 13C and 15N natural isotope abundance reflects breast cancer cell metabolism. Sci. Rep. 2016, 6, 34251. [Google Scholar] [CrossRef]
- Noland, R.C.; Woodlief, T.L.; Whitfield, B.R.; Manning, S.M.; Evans, J.R.; Dudek, R.W.; Lust, R.M.; Cortright, R.N. Peroxisomal-mitochondrial oxidation in a rodent model of obesity-associated insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2007, 293, 986–1001. [Google Scholar] [CrossRef]
- Galgani, J.E.; Heilbronn, L.K.; Azuma, K.; Kelley, D.E.; Albu, J.B.; Pi-Sunyer, X.; Smith, S.R.; Ravussin, E.; Look, A.A.R.G. Metabolic flexibility in response to glucose is not impaired in people with type 2 diabetes after controlling for glucose disposal rate. Diabetes 2008, 57, 841–845. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT, min (RP/HILIC) | Experimental m/z | RP/HILIC | Elemental Formula | MS/MS Fragment Ions | Collision Energy, eV | MSI MI Level | Compound |
|---|---|---|---|---|---|---|---|
| 0.48/5.40 | 162.1114 | RP/HILIC | C7H16NO3 | 103.0396, 85.0295 | 20 | 1 | carnitine |
| 0.50/4.49 | 204.1216 | RP/HILIC | C9H18NO4 | 144.1015, 85.0291 | 15 | 1 | acetylcarnitine (C2) |
| 0.59/3.53 | 218.1374 | RP | C10H20NO4 | 159.0656, 144.1039, 85.0289 | 15 | 1 | propionylcarnitine (C4) |
| 0.66 | 169.0355 | RP | C5H5N4O3 | 152.0097, 141.0423, 126.0302, 98.0357, 96.0194, 70.0409, 69.0080 | 20 | 1 | uric acid |
| 0.70 | 167.0559 | RP | C6H7N4O2 | 149.0470, 124.0518, 110.0356, 96.0554, 82.0407 | 20 | 2 | oxoguanine |
| 0.70 | 181.0711 | RP | C7H9N4O | 163.0618, 135.0715, 124.0516, 67.0287 | 20 | 3 | theophylline-like structure |
| 0.70 | 244.1525 | RP | C12H22NO4 | 185.0824, 144.1022, 85.0288 | 20 | 2 | (iso)tiglylcarnitine or 3-methylcrotonylcarnitine (C5:1) |
| 0.70/2.90 | 232.1529 | RP/HILIC | C11H22NO4 | 173.0833, 144.1036, 128.0701, 85.0285 | 15 | 1 | isobutyrylcarnitine (C4) |
| 0.70 | 246.1683 | RP | C12H24NO4 | 187.0973, 144.1012, 85.0289 | 15 | 2 | 2-methylbutyroylcarnitine (C5) |
| 3.97/1.72 | 286.2013 | RP/HILIC | C15H28NO4 | 227.1272, 144.1019, 125.0972, 97.1009, 85.0290 | 20 | 2 | octenoylcarnitine (C8:1) |
| 4.35/1.48 | 310.2013 | RP/HILIC | C17H28NO4 | 251.1277, 149.0961, 144.1022, 121.1018, 93.0714, 85.0294 | 20 | 2 | decatrienoylcarnitine (C10:3) |
| 5.13 | 312.2169 | RP | C17H30NO4 | 253.1436, 151.1120, 144.1032, 123.1174, 85.0290 | 20 | 2 | decadienoylcarnitine (C10:2) |
| 5.68 | 328.2485 | RP | C18H34NO4 | 269.1742, 144.1016, 85.0284 | 20 | -- | unknown acylcarnitine |
| CD | NP-HP | HP-NP | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| P1 | P2 | P1 (NP) | P2 (HP) | P1 (HP) | P2 (NP) | Diet | Period | |
| Glucose (mmol/L) | 5.07 ± 0.04 | 5.08 ± 0.06 | 4.98 ± 0.11 | 5.14 ± 0.12 | 4.97 ± 0.13 | 5.09 ± 0.11 | 0.90 | 0.04 |
| Insulin (mU/L) | 4.0 1± 0.62 | 4.22 ± 0.85 | 4.74 ± 1.14 | 4.93 ± 1.13 | 3.08 ± 0.57 | 3.74 ± 0.62 | 0.81 | 0.25 |
| TG (mmol/L) | 1.03 ± 0.10 | 0.85 ± 0.08 | 0.69 ± 0.06 | 0.70 ± 0.04 | 0.61 ± 0.06 | 0.84 ± 0.06 | 0.003 * | 0.89 |
| FFA (mmol/L) | 0.40 ± 0.07 | 0.25 ± 0.03 | 0.41 ± 0.08 | 0.22 ± 0.05 | 0.32 ± 0.08 | 0.21 ± 0.03 | 0.65 | 0.003 |
| β-HB (mmol/L) | 0.07 ± 0.02 | 0.05 ± 0.01 | 0.09 ± 0.03 | 0.05 ± 0.01 | 0.12 ± 0.04 | 0.04 ± 0.01 | 0.45 | 0.02 |
| Glycerol (umol/L) | 40.8 ± 4.43 | 34.9 ± 4.69 | 45.2 ± 6.84 | 34.2 ± 6.22 | 36.0 ± 6.37 | 37.2 ± 6.57 | 0.45 | 0.18 |
| Total Cholesterol (mmol/L) | 4.79 ± 0.30 | 4.33 ± 0.25 | 4.69 ± 0.42 | 4.35 ± 0.18 | 4.98 ± 0.32 | 5.51 ± 0.59 | 0.20 | 0.61 |
| HDL Cholesterol (mmol/L) | 1.19 ± 0.09 | 1.11 ± 0.07 | 1.33 ± 0.08 | 1.32 ± 0.15 | 1.24 ± 0.09 | 1.25 ± 0.07 | 0.20 | 0.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khodorova, N.V.; Rietman, A.; Rutledge, D.N.; Schwarz, J.; Piedcoq, J.; Pilard, S.; Siebelink, E.; Kok, F.J.; Tomé, D.; Mensink, M.; et al. Urinary Medium-Chained Acyl-Carnitines Sign High Caloric Intake whereas Short-Chained Acyl-Carnitines Sign High -Protein Diet within a High-Fat, Hypercaloric Diet in a Randomized Crossover Design Dietary Trial. Nutrients 2021, 13, 1191. https://doi.org/10.3390/nu13041191
Khodorova NV, Rietman A, Rutledge DN, Schwarz J, Piedcoq J, Pilard S, Siebelink E, Kok FJ, Tomé D, Mensink M, et al. Urinary Medium-Chained Acyl-Carnitines Sign High Caloric Intake whereas Short-Chained Acyl-Carnitines Sign High -Protein Diet within a High-Fat, Hypercaloric Diet in a Randomized Crossover Design Dietary Trial. Nutrients. 2021; 13(4):1191. https://doi.org/10.3390/nu13041191
Chicago/Turabian StyleKhodorova, Nadezda V., Annemarie Rietman, Douglas N. Rutledge, Jessica Schwarz, Julien Piedcoq, Serge Pilard, Els Siebelink, Frans J. Kok, Daniel Tomé, Marco Mensink, and et al. 2021. "Urinary Medium-Chained Acyl-Carnitines Sign High Caloric Intake whereas Short-Chained Acyl-Carnitines Sign High -Protein Diet within a High-Fat, Hypercaloric Diet in a Randomized Crossover Design Dietary Trial" Nutrients 13, no. 4: 1191. https://doi.org/10.3390/nu13041191
APA StyleKhodorova, N. V., Rietman, A., Rutledge, D. N., Schwarz, J., Piedcoq, J., Pilard, S., Siebelink, E., Kok, F. J., Tomé, D., Mensink, M., & Azzout-Marniche, D. (2021). Urinary Medium-Chained Acyl-Carnitines Sign High Caloric Intake whereas Short-Chained Acyl-Carnitines Sign High -Protein Diet within a High-Fat, Hypercaloric Diet in a Randomized Crossover Design Dietary Trial. Nutrients, 13(4), 1191. https://doi.org/10.3390/nu13041191