
Ghrelin and Glucagon-Like Peptide-1: A Gut-Brain Axis Battle for Food Reward


{kind=link}
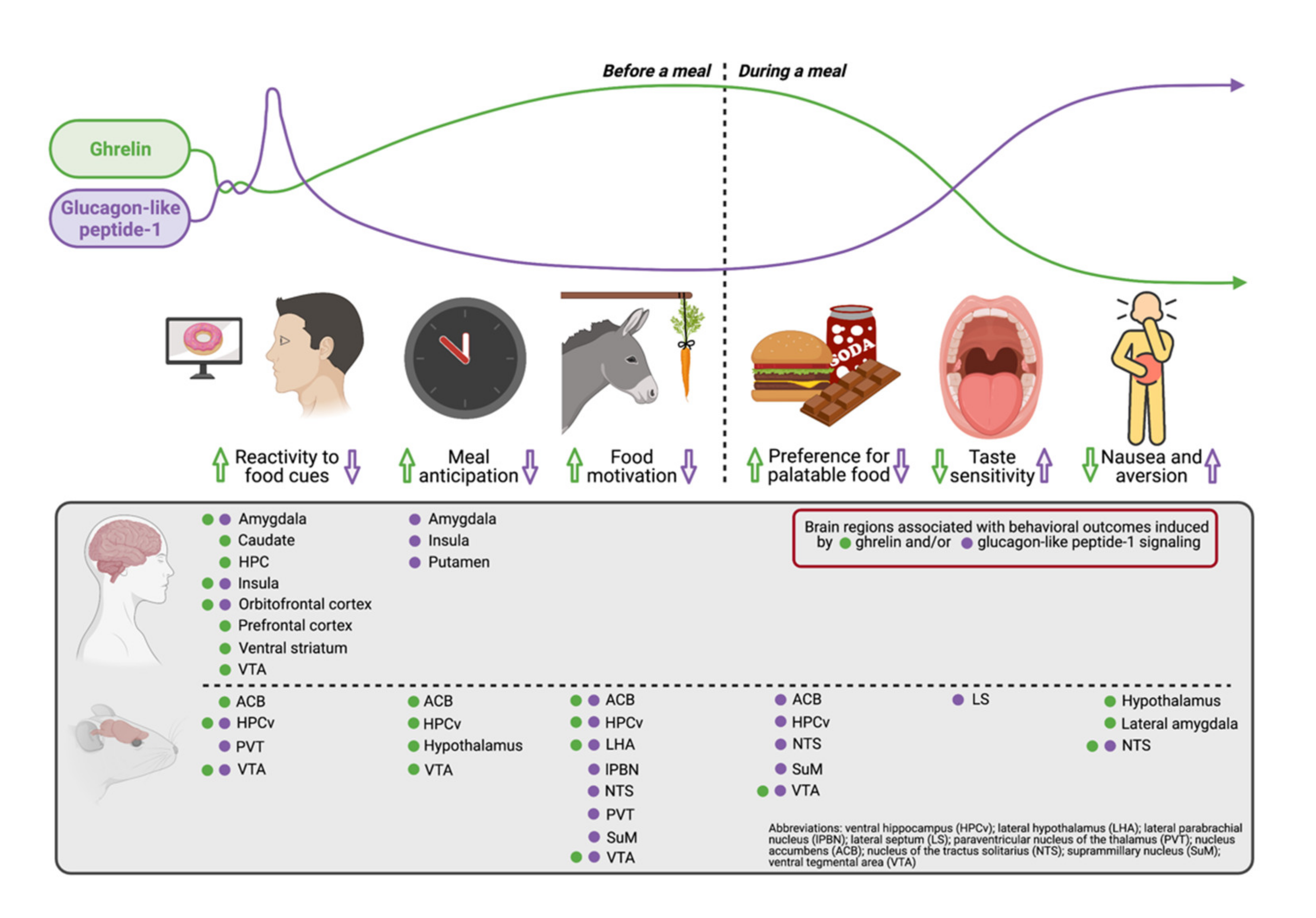
Abstract
1. Introduction
2. Reactivity to Food Cues
2.1. Rodent Studies
2.2. Human Studies
2.3. Summary
3. Meal Anticipation
3.1. Rodent Studies
3.2. Human Studies
3.3. Summary
4. Food Motivation
4.1. Rodent Studies
4.2. Human Studies
4.3. Summary
5. Nutrient Preference
5.1. Rodent Studies
5.2. Human Studies
5.3. Summary
6. Flavor
6.1. Rodent Studies
6.2. Human Studies
6.3. Summary
7. Aversion
7.1. Rodent Studies
7.2. Human Studies
7.3. Summary
8. Future Directions
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sun, Y.; Wang, P.; Zheng, H.; Smith, R.G. Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 4679–4684. [Google Scholar] [CrossRef]
- Drazen, D.L.; Vahl, T.P.; D’Alessio, D.A.; Seeley, R.J.; Woods, S.C. Effects of a fixed meal pattern on ghrelin secretion: Evidence for a learned response independent of nutrient status. Endocrinology 2006, 147, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Callahan, H.S.; Cummings, D.E.; Pepe, M.S.; Breen, P.A.; Matthys, C.C.; Weigle, D.S. Postprandial suppression of plasma ghrelin level is proportional to ingested caloric load but does not predict intermeal interval in humans. J. Clin. Endocrinol. Metab. 2004, 89, 1319–1324. [Google Scholar] [CrossRef]
- Davis, E.A.; Wald, H.S.; Suarez, A.N.; Zubcevic, J.; Liu, C.M.; Cortella, A.M.; Kamitakahara, A.K.; Polson, J.W.; Arnold, M.; Grill, H.J.; et al. Ghrelin signaling regulates feeding behavior, metabolism, and memory through the vagus nerve. Curr. Biol. 2020. [Google Scholar] [CrossRef]
- Krieger, J.P.; Arnold, M.; Pettersen, K.G.; Lossel, P.; Langhans, W.; Lee, S.J. Knockdown of GLP-1 receptors in vagal afferents affects normal food intake and glycemia. Diabetes 2016, 65, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Kanoski, S.E.; Hayes, M.R.; Skibicka, K.P. GLP-1 and weight loss: Unraveling the diverse neural circuitry. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R885–R895. [Google Scholar] [CrossRef]
- Boswell, R.G.; Kober, H. Food cue reactivity and craving predict eating and weight gain: A meta-analytic review. Obes. Rev. 2016, 17, 159–177. [Google Scholar] [CrossRef]
- Walker, A.K.; Ibia, I.E.; Zigman, J.M. Disruption of cue-potentiated feeding in mice with blocked ghrelin signaling. Physiol. Behav. 2012, 108, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Kanoski, S.E.; Fortin, S.M.; Ricks, K.M.; Grill, H.J. Ghrelin signaling in the ventral hippocampus stimulates learned and motivational aspects of feeding via PI3K-Akt signaling. Biol. Psychiatry 2013, 73, 915–923. [Google Scholar] [CrossRef] [PubMed]
- St-Onge, V.; Watts, A.; Abizaid, A. Ghrelin enhances cue-induced bar pressing for high fat food. Horm. Behav. 2016, 78, 141–149. [Google Scholar] [CrossRef]
- Sommer, S.; Hauber, W. Ghrelin receptor activation in the ventral tegmental area amplified instrumental responding but not the excitatory influence of Pavlovian stimuli on instrumental responding. Neurobiol. Learn. Mem. 2016, 134, 210–215. [Google Scholar] [CrossRef]
- Davidson, T.L.; Kanoski, S.E.; Tracy, A.L.; Walls, E.K.; Clegg, D.; Benoit, S.C. The interoceptive cue properties of ghrelin generalize to cues produced by food deprivation. Peptides 2005, 26, 1602–1610. [Google Scholar] [CrossRef] [PubMed]
- Suarez, A.N.; Liu, C.M.; Cortella, A.M.; Noble, E.E.; Kanoski, S.E. Ghrelin and Orexin Interact to Increase Meal Size Through a Descending Hippocampus to Hindbrain Signaling Pathway. Biol. Psychiatry 2020, 87, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Van Der Plasse, G.; Van Zessen, R.; Luijendijk, M.C.M.; Erkan, H.; Stuber, G.D.; Ramakers, G.M.J.; Adan, R.A.H. Modulation of cue-induced firing of ventral tegmental area dopamine neurons by leptin and ghrelin. Int. J. Obes. 2015, 39, 1742–1749. [Google Scholar] [CrossRef] [PubMed]
- Abizaid, A.; Liu, Z.-W.; Andrews, Z.B.; Shanabrough, M.; Borok, E.; Elsworth, J.D.; Roth, R.H.; Sleeman, M.W.; Picciotto, M.R.; Tschöp, M.H.; et al. Ghrelin modulates the activity and synaptic input organization of midbrain dopamine neurons while promoting appetite. J. Clin. Investig. 2006, 116, 3229–3239. [Google Scholar] [CrossRef] [PubMed]
- Roseberry, A.G. Acute fasting increases somatodendritic dopamine release in the ventral Tegmental area. J. Neurophysiol. 2015, 114, 1072–1082. [Google Scholar] [CrossRef]
- Godfrey, N.; Borgland, S.L. Sex differences in the effect of acute fasting on excitatory and inhibitory synapses onto ventral tegmental area dopamine neurons. J. Physiol. 2020, 598, 5523–5539. [Google Scholar] [CrossRef]
- Van Der Plasse, G.; Merkestein, M.; Luijendijk, M.C.M.; Van Der Roest, M.; Westenberg, H.G.M.; Mulder, A.B.; Adan, R.A.H. Food cues and ghrelin recruit the same neuronal circuitry. Int. J. Obes. 2013, 37, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.W.; Canter, R.; Gallagher, M.; Holland, P.C. Assessing the Role of the Growth Hormone Secretagogue Receptor in Motivational Learning and Food Intake. Behav. Neurosci. 2009, 123, 1058–1065. [Google Scholar] [CrossRef]
- Konanur, V.R.; Hsu, T.M.; Kanoski, S.E.; Hayes, M.R.; Roitman, M.F. Phasic dopamine responses to a food-predictive cue are suppressed by the glucagon-like peptide-1 receptor agonist Exendin-4. Physiol. Behav. 2020, 215, 112771. [Google Scholar] [CrossRef] [PubMed]
- Ong, Z.Y.; Liu, J.J.; Pang, Z.P.; Grill, H.J. Paraventricular thalamic control of food intake and reward: Role of glucagon-like peptide-1 receptor signaling. Neuropsychopharmacology 2017, 42, 2387–2397. [Google Scholar] [CrossRef]
- Kroemer, N.B.; Krebs, L.; Kobiella, A.; Grimm, O.; Pilhatsch, M.; Bidlingmaier, M.; Zimmermann, U.S.; Smolka, M.N. Fasting levels of ghrelin covary with the brain response to food pictures. Addict. Biol. 2013, 18, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Holsen, L.M.; Lawson, E.A.; Christensen, K.; Klibanski, A.; Goldstein, J.M. Abnormal relationships between the neural response to high- and low-calorie foods and endogenous acylated ghrelin in women with active and weight-recovered anorexia nervosa. Psychiatry Res. Neuroimaging 2014, 223, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Schüssler, P.; Kluge, M.; Yassouridis, A.; Dresler, M.; Uhr, M.; Steiger, A. Ghrelin levels increase after pictures showing food. Obesity 2012, 20, 1212–1217. [Google Scholar] [CrossRef]
- Dorton, H.M.; Luo, S.; Monterosso, J.R.; Page, K.A. Influences of dietary added sugar consumption on striatal food-cue reactivity and postprandial GLP-1 response. Front. Psychiatry 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Heni, M.; Kullmann, S.; Gallwitz, B.; Häring, H.U.; Preissl, H.; Fritsche, A. Dissociation of GLP-1 and insulin association with food processing in the brain: GLP-1 sensitivity despite insulin resistance in obese humans. Mol. Metab. 2015, 4, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Maurer, L.; Mai, K.; Krude, H.; Haynes, J.D.; Weygandt, M.; Spranger, J. Interaction of circulating GLP-1 and the response of the dorsolateral prefrontal cortex to food-cues predicts body weight development. Mol. Metab. 2019, 29, 136–144. [Google Scholar] [CrossRef]
- Goldstone, A.P.; Prechtl, C.G.; Scholtz, S.; Miras, A.D.; Chhina, N.; Durighel, G.; Deliran, S.S.; Beckmann, C.; Ghatei, M.A.; Ashby, D.R.; et al. Ghrelin mimics fasting to enhance human hedonic, orbitofrontal cortex, and hippocampal responses to food. Am. J. Clin. Nutr. 2014, 99, 1319–1330. [Google Scholar] [CrossRef]
- Han, J.E.; Frasnelli, J.; Zeighami, Y.; Larcher, K.; Boyle, J.; McConnell, T.; Malik, S.; Jones-Gotman, M.; Dagher, A. Ghrelin Enhances Food Odor Conditioning in Healthy Humans: An fMRI Study. Cell Rep. 2018, 25, 2643–2652. [Google Scholar] [CrossRef]
- Malik, S.; McGlone, F.; Bedrossian, D.; Dagher, A. Ghrelin Modulates Brain Activity in Areas that Control Appetitive Behavior. Cell Metab. 2008, 7, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Van Bloemendaal, L.; IJzerman, R.G.; Ten Kulve, J.S.; Barkhof, F.; Konrad, R.J.; Drent, M.L.; Veltman, D.J.; Diamant, M. GLP-1 receptor activation modulates appetite- and reward-related brain areas in humans. Diabetes 2014, 63, 4186–4196. [Google Scholar] [CrossRef]
- Van Bloemendaal, L.; Veltman, D.J.; Ten Kulve, J.S.; Drent, M.L.; Barkhof, F.; Diamant, M.; Ijzerman, R.G. Emotional eating is associated with increased brain responses to food-cues and reduced sensitivity to GLP-1 receptor activation. Obesity 2015, 23, 2075–2082. [Google Scholar] [CrossRef]
- Ten Kulve, J.S.; Veltman, D.J.; van Bloemendaal, L.; Barkhof, F.; Deacon, C.F.; Holst, J.J.; Konrad, R.J.; Sloan, J.H.; Drent, M.L.; Diamant, M.; et al. Endogenous GLP-1 mediates postprandial reductions in activation in central reward and satiety areas in patients with type 2 diabetes. Diabetologia 2015, 58, 2688–2698. [Google Scholar] [CrossRef]
- Farr, O.M.; Li, C.R.; Mantzoros, C.S. Central nervous system regulation of eating: Insights from human brain imaging. Metabolism 2016, 65, 699–713. [Google Scholar] [CrossRef]
- Farr, O.M.; Tsoukas, M.A.; Triantafyllou, G.; Dincer, F.; Filippaios, A.; Ko, B.J.; Mantzoros, C.S. Short-term administration of the GLP-1 analog liraglutide decreases circulating leptin and increases GIP levels and these changes are associated with alterations in CNS responses to food cues: A randomized, placebo-controlled, crossover study. Metabolism 2016, 65, 945–953. [Google Scholar] [CrossRef]
- Farr, O.M.; Upadhyay, J.; Rutagengwa, C.; DiPrisco, B.; Ranta, Z.; Adra, A.; Bapatla, N.; Douglas, V.P.; Douglas, K.A.A.; Nolen-Doerr, E.; et al. Longer-term liraglutide administration at the highest dose approved for obesity increases reward-related orbitofrontal cortex activation in response to food cues: Implications for plateauing weight loss in response to anti-obesity therapies. Diabetes Obes. Metab. 2019, 21, 2459–2464. [Google Scholar] [CrossRef]
- Sinclair, P.; Brennan, D.J.; le Roux, C.W. Gut adaptation after metabolic surgery and its influences on the brain, liver and cancer. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 606–624. [Google Scholar] [CrossRef] [PubMed]
- Madsbad, S.; Dirksen, C.; Holst, J.J. Mechanisms of changes in glucose metabolism and bodyweight after bariatric surgery. Lancet Diabetes Endocrinol. 2014, 2, 152–164. [Google Scholar] [CrossRef]
- Hutch, C.R.; Sandoval, D. The role of GLP-1 in the metabolic success of bariatric surgery. Endocrinology 2017, 158, 4139–4151. [Google Scholar] [CrossRef] [PubMed]
- Lepping, R.J.; Bruce, A.S.; Francisco, A.; Yeh, H.W.; Martin, L.E.; Powell, J.N.; Hancock, L.; Patrician, T.M.; Breslin, F.J.; Selim, N.; et al. Resting-state brain connectivity after surgical and behavioral weight loss. Obesity 2015. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, G.; Li, G.; Hu, Y.; Liu, L.; Jin, Q.; Meng, Q.; Zhao, J.; Yuan, K.; Liu, J.; et al. Ghrelin reductions following bariatric surgery were associated with decreased resting state activity in the hippocampus. Int. J. Obes. 2019, 43, 842–851. [Google Scholar] [CrossRef]
- Hu, Y.; Ji, G.; Li, G.; Manza, P.; Zhang, W.; Wang, J.; Lv, G.; He, Y.; Zhang, Z.; Yuan, K.; et al. Brain Connectivity, and Hormonal and Behavioral Correlates of Sustained Weight Loss in Obese Patients after Laparoscopic Sleeve Gastrectomy. Cereb. Cortex 2021, 31, 1284–1295. [Google Scholar] [CrossRef]
- Giel, K.E.; Rieber, N.; Enck, P.; Friederich, H.C.; Meile, T.; Zipfel, S.; Teufel, M. Effects of laparoscopic sleeve gastrectomy on attentional processing of food-related information: Evidence from eye-tracking. Surg. Obes. Relat. Dis. 2014. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Ji, G.; Hu, Y.; Liu, L.; Jin, Q.; Zhang, W.; Liu, L.; Wang, Y.; Zhao, J.; von Deneen, K.M.; et al. Reduced plasma ghrelin concentrations are associated with decreased brain reactivity to food cues after laparoscopic sleeve gastrectomy. Psychoneuroendocrinology 2019, 100, 229–236. [Google Scholar] [CrossRef]
- Ochner, C.N.; Laferrère, B.; Afifi, L.; Atalayer, D.; Geliebter, A.; Teixeira, J. Neural responsivity to food cues in fasted and fed states pre and post gastric bypass surgery. Neurosci. Res. 2012, 74, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Zoon, H.F.A.; de Bruijn, S.E.M.; Jager, G.; Smeets, P.A.M.; de Graaf, C.; Janssen, I.M.C.; Schijns, W.; Deden, L.; Boesveldt, S. Altered neural inhibition responses to food cues after Roux-en-Y Gastric Bypass. Biol. Psychol. 2018, 137, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Van Duinkerken, E.; Bernardes, G.; van Bloemendaal, L.; Veltman, D.J.; Barkhof, F.; Mograbi, D.C.; Gerdes, V.E.A.; Deacon, C.F.; Holst, J.J.; Drent, M.L.; et al. Cerebral effects of glucagon-like peptide-1 receptor blockade before and after Roux-en-Y gastric bypass surgery in obese women: A proof-of-concept resting-state functional MRI study. Diabetes Obes. Metab. 2021, 23, 415–424. [Google Scholar] [CrossRef]
- Vahl, T.P.; Drazen, D.L.; Seeley, R.J.; D’Alessio, D.A.; Woods, S.C. Meal-anticipatory glucagon-like peptide-1 secretion in rats. Endocrinology 2010, 151, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Ott, V.; Friedrich, M.; Zemlin, J.; Lehnert, H.; Schultes, B.; Born, J.; Hallschmid, M. Meal anticipation potentiates postprandial ghrelin suppression in humans. Psychoneuroendocrinology 2012, 37, 1096–1100. [Google Scholar] [CrossRef]
- Davis, J.F.; Choi, D.L.; Clegg, D.J.; Benoit, S.C. Signaling through the ghrelin receptor modulates hippocampal function and meal anticipation in mice. Physiol. Behav. 2011, 103, 39–43. [Google Scholar] [CrossRef]
- Lesauter, J.; Hoque, N.; Weintraub, M.; Pfaff, D.W.; Silver, R. Stomach ghrelin-secreting cells as food-entrainable circadian clocks. Proc. Natl. Acad. Sci. USA 2009, 106, 13582–13587. [Google Scholar] [CrossRef]
- Blum, I.D.; Patterson, Z.; Khazall, R.; Lamont, E.W.; Sleeman, M.W.; Horvath, T.L.; Abizaid, A. Reduced anticipatory locomotor responses to scheduled meals in ghrelin receptor deficient mice. Neuroscience 2009, 164, 351–359. [Google Scholar] [CrossRef]
- Lamont, E.W.; Patterson, Z.; Rodrigues, T.; Vallejos, O.; Blum, I.D.; Abizaid, A. Ghrelin-deficient mice have fewer orexin cells and reduced cFOS expression in the mesolimbic dopamine pathway under a restricted feeding paradigm. Neuroscience 2012, 218, 12–19. [Google Scholar] [CrossRef]
- Hsu, T.M.; Hahn, J.D.; Konanur, V.R.; Noble, E.E.; Suarez, A.N.; Thai, J.; Nakamoto, E.M.; Kanoski, S.E. Hippocampus ghrelin signaling mediates appetite through lateral hypothalamic orexin pathways. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Merkestein, M.; Brans, M.A.D.; Luijendijk, M.C.M.; De Jong, J.W.; Egecioglu, E.; Dickson, S.L.; Adan, R.A.H. Ghrelin mediates anticipation to a palatable meal in rats. Obesity 2012, 20, 963–971. [Google Scholar] [CrossRef]
- Wei, X.J.; Sun, B.; Chen, K.; Lv, B.; Luo, X.; Yan, J.Q. Ghrelin signaling in the ventral tegmental area mediates both reward-based feeding and fasting-induced hyperphagia on high-fat diet. Neuroscience 2015, 300, 53–62. [Google Scholar] [CrossRef]
- Bake, T.; Hellgren, K.T.; Dickson, S.L. Acute ghrelin changes food preference from a high-fat diet to chow during binge-like eating in rodents. J. Neuroendocrinol. 2017, 29. [Google Scholar] [CrossRef] [PubMed]
- Pierce-Messick, Z.; Pratt, W.E. Glucagon-like peptide-1 receptors modulate the binge-like feeding induced by µ-opioid receptor stimulation of the nucleus accumbens in the rat. Neuroreport 2020, 31, 1283–1288. [Google Scholar] [CrossRef]
- Cummings, D.E.; Purnell, J.Q.; Frayo, R.S.; Schmidova, K.; Wisse, B.E.; Weigle, D.S. A Preprandial Rise in Plasma Ghrelin Levels Suggests a Role in Meal Initiation in Humans. Diabetes 2001, 50, 1714–1719. [Google Scholar] [CrossRef] [PubMed]
- Frecka, J.M.; Mattes, R.D. Possible entrainment of ghrelin to habitual meal patterns in humans. Am. J. Physiol. Liver Physiol. 2008, 294, G699–G707. [Google Scholar] [CrossRef]
- Rigamonti, A.E.; Piscitelli, F.; Aveta, T.; Agosti, F.; De Col, A.; Bini, S.; Cella, S.G.; Di Marzo, V.; Sartorio, A. Anticipatory and consummatory effects of (hedonic) chocolate intake are associated with increased circulating levels of the orexigenic peptide ghrelin and endocannabinoids in obese adults. Food Nutr. Res. 2015, 59. [Google Scholar] [CrossRef]
- Van Bloemendaal, L.; Veltman, D.J.; Ten Kulve, J.S.; Groot, P.F.C.; Ruhé, H.G.; Barkhof, F.; Sloan, J.H.; Diamant, M.; Ijzerman, R.G. Brain reward-system activation in response to anticipation and consumption of palatable food is altered by glucagon-like peptide-1 receptor activation in humans. Diabetes Obes. Metab. 2015, 17, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Perello, M.; Sakata, I.; Birnbaum, S.; Chuang, J.C.; Osborne-Lawrence, S.; Rovinsky, S.A.; Woloszyn, J.; Yanagisawa, M.; Lutter, M.; Zigman, J.M. Ghrelin Increases the Rewarding Value of High-Fat Diet in an Orexin-Dependent Manner. Biol. Psychiatry 2010, 67, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Egecioglu, E.; Jerlhag, E.; Salomé, N.; Skibicka, K.P.; Haage, D.; Bohlooly-Y, M.; Andersson, D.; Bjursell, M.; Perrissoud, D.; Engel, J.A.; et al. Ghrelin increases intake of rewarding food in rodents. Addict. Biol. 2010, 15, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Bake, T.; Le May, M.V.; Edvardsson, C.E.; Vogel, H.; Bergström, U.; Albers, M.N.; Skibicka, K.P.; Farkas, I.; Liposits, Z.; Dickson, S.L. Ghrelin Receptor Stimulation of the Lateral Parabrachial Nucleus in Rats Increases Food Intake but not Food Motivation. Obesity 2020, 28, 1503–1511. [Google Scholar] [CrossRef]
- Dickson, S.L.; Shirazi, R.H.; Hansson, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. The glucagon-like peptide 1 (GLP-1) analogue, exendin-4, decreases the rewarding value of food: A new role for mesolimbic GLP-1 receptors. J. Neurosci. 2012, 32, 4812–4820. [Google Scholar] [CrossRef] [PubMed]
- Alhadeff, A.L.; Grill, H.J. Hindbrain nucleus tractus solitarius glucagon-like peptide-1 receptor signaling reduces appetitive and motivational aspects of feeding. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307. [Google Scholar] [CrossRef]
- Richard, J.E.; Anderberg, R.H.; Göteson, A.; Gribble, F.M.; Reimann, F.; Skibicka, K.P. Activation of the GLP-1 receptors in the nucleus of the solitary tract reduces food reward behavior and targets the mesolimbic system. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Hsu, T.M.; Hahn, J.D.; Konanur, V.R.; Lam, A.; Kanoski, S.E. Hippocampal GLP-1 receptors influence food intake, meal size, and effort-based responding for food through volume transmission. Neuropsychopharmacology 2015, 40, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.F.; Perello, M.; Choi, D.L.; Magrisso, I.J.; Kirchner, H.; Pfluger, P.T.; Tschoep, M.; Zigman, J.M.; Benoit, S.C. GOAT induced ghrelin acylation regulates hedonic feeding. Horm. Behav. 2012, 62, 598–604. [Google Scholar] [CrossRef]
- Finger, B.C.; Dinan, T.G.; Cryan, J.F. Diet-induced obesity blunts the behavioural effects of ghrelin: Studies in a mouse-progressive ratio task. Psychopharmacol. 2012, 220, 173–181. [Google Scholar] [CrossRef]
- Landgren, S.; Simms, J.A.; Thelle, D.S.; Strandhagen, E.; Bartlett, S.E.; Engel, J.A.; Jerlhag, E. The ghrelin signalling system is involved in the consumption of sweets. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Bake, T.; Edvardsson, C.E.; Cummings, C.J.; Dickson, S.L. Ghrelin’s effects on food motivation in rats are not limited to palatable foods. J. Neuroendocrinol. 2019, 31. [Google Scholar] [CrossRef] [PubMed]
- Overduin, J.; Figlewicz, D.P.; Bennett-Jay, J.; Kittleson, S.; Cummings, D.E. Ghrelin increases the motivation to eat, but does not alter food palatability. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, 259–269. [Google Scholar] [CrossRef]
- Salamone, J.D.; Correa, M. The Mysterious Motivational Functions of Mesolimbic Dopamine. Neuron 2012, 76, 470–485. [Google Scholar] [CrossRef]
- Skibicka, K.P.; Shirazi, R.H.; Rabasa-Papio, C.; Alvarez-Crespo, M.; Neuber, C.; Vogel, H.; Dickson, S.L. Divergent circuitry underlying food reward and intake effects of ghrelin: Dopaminergic VTA-accumbens projection mediates ghrelin’s effect on food reward but not food intake. Neuropharmacology 2013, 73, 274–283. [Google Scholar] [CrossRef]
- King, S.J.; Isaacs, A.M.; O’Farrell, E.; Abizaid, A. Motivation to obtain preferred foods is enhanced by ghrelin in the ventral tegmental area. Horm. Behav. 2011, 60, 572–580. [Google Scholar] [CrossRef]
- Weinberg, Z.Y.; Nicholson, M.L.; Currie, P.J. 6-Hydroxydopamine lesions of the ventral tegmental area suppress ghrelin’s ability to elicit food-reinforced behavior. Neurosci. Lett. 2011, 499, 70–73. [Google Scholar] [CrossRef]
- Skibicka, K.P.; Hansson, C.; Alvarez-Crespo, M.; Friberg, P.A.; Dickson, S.L. Ghrelin directly targets the ventral tegmental area to increase food motivation. Neuroscience 2011, 180, 129–137. [Google Scholar] [CrossRef] [PubMed]
- López-Ferreras, L.; Richard, J.E.; Anderberg, R.H.; Nilsson, F.H.; Olandersson, K.; Kanoski, S.E.; Skibicka, K.P. Ghrelin’s control of food reward and body weight in the lateral hypothalamic area is sexually dimorphic. Physiol. Behav. 2017, 176, 40–49. [Google Scholar] [CrossRef]
- Cone, J.J.; McCutcheon, J.E.; Roitman, M.F. Ghrelin acts as an interface between physiological state and phasic dopamine signaling. J. Neurosci. 2014, 34, 4905–4913. [Google Scholar] [CrossRef]
- Terrill, S.J.; Wall, K.D.; Medina, N.D.; Maske, C.B.; Williams, D.L. Lateral septum growth hormone secretagogue receptor affects food intake and motivation for sucrose reinforcement. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R76–R83. [Google Scholar] [CrossRef] [PubMed]
- Howell, E.; Baumgartner, H.M.; Zallar, L.J.; Selva, J.A.; Engel, L.; Currie, P.J. Glucagon-like peptide-1 (GLP-1) and 5-hydroxytryptamine 2c (5-HT2c) receptor agonists in the ventral tegmental area (VTA) inhibit ghrelin-stimulated appetitive reward. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Bernosky-Smith, K.A.; Stanger, D.B.; Trujillo, A.J.; Mitchell, L.R.; España, R.A.; Bass, C.E. The GLP-1 agonist exendin-4 attenuates self-administration of sweetened fat on fixed and progressive ratio schedules of reinforcement in rats. Pharmacol. Biochem. Behav. 2016, 142, 48–55. [Google Scholar] [CrossRef]
- Anderberg, R.H.; Anefors, C.; Bergquist, F.; Nissbrandt, H.; Skibicka, K.P. Dopamine signaling in the amygdala, increased by food ingestion and GLP-1, regulates feeding behavior. Physiol. Behav. 2014, 136, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Wolf, S.; Rabasa, C.; Rodriguez-Pacheco, F.; Babaei, C.S.; Stöber, F.; Goldschmidt, J.; DiMarchi, R.D.; Finan, B.; Tschöp, M.H.; et al. GLP-1 and estrogen conjugate acts in the supramammillary nucleus to reduce food-reward and body weight. Neuropharmacology 2016, 110, 396–406. [Google Scholar] [CrossRef]
- Alhadeff, A.L.; Baird, J.P.; Swick, J.C.; Hayes, M.R.; Grill, H.J. Glucagon-like peptide-1 receptor signaling in the lateral parabrachial nucleus contributes to the control of food intake and motivation to feed. Neuropsychopharmacology 2014, 39, 2233–2243. [Google Scholar] [CrossRef]
- Terrill, S.J.; Holt, M.K.; Maske, C.B.; Abrams, N.; Reimann, F.; Trapp, S.; Williams, D.L. Endogenous GLP-1 in lateral septum promotes satiety and suppresses motivation for food in mice. Physiol. Behav. 2019, 206, 191–199. [Google Scholar] [CrossRef] [PubMed]
- López-Ferreras, L.; Eerola, K.; Mishra, D.; Shevchouk, O.T.; Richard, J.E.; Nilsson, F.H.; Hayes, M.R.; Skibicka, K.P. GLP-1 modulates the supramammillary nucleus-lateral hypothalamic neurocircuit to control ingestive and motivated behavior in a sex divergent manner. Mol. Metab. 2019, 20, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.E.; Anderberg, R.H.; López-Ferreras, L.; Olandersson, K.; Skibicka, K.P. Sex and estrogens alter the action of glucagon-like peptide-1 on reward. Biol. Sex Differ. 2016, 7, 1–6. [Google Scholar] [CrossRef]
- López-Ferreras, L.; Richard, J.E.; Noble, E.E.; Eerola, K.; Anderberg, R.H.; Olandersson, K.; Taing, L.; Kanoski, S.E.; Hayes, M.R.; Skibicka, K.P. Lateral hypothalamic GLP-1 receptors are critical for the control of food reinforcement, ingestive behavior and body weight. Mol. Psychiatry 2018, 23, 1157–1168. [Google Scholar] [CrossRef]
- Maske, C.B.; Loney, G.C.; Lilly, N.; Terrill, S.J.; Williams, D.L. Intragastric nutrient infusion reduces motivation for food in male and female rats. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E81–E90. [Google Scholar] [CrossRef] [PubMed]
- Anderberg, R.H.; Hansson, C.; Fenander, M.; Richard, J.E.; Dickson, S.L.; Nissbrandt, H.; Bergquist, F.; Skibicka, K.P. The Stomach-Derived Hormone Ghrelin Increases Impulsive Behavior. Neuropsychopharmacology 2016, 41, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.M.; Noble, E.E.; Liu, C.M.; Cortella, A.M.; Konanur, V.R.; Suarez, A.N.; Reiner, D.J.; Hahn, J.D.; Hayes, M.R.; Kanoski, S.E. A hippocampus to prefrontal cortex neural pathway inhibits food motivation through glucagon-like peptide-1 signaling. Mol. Psychiatry 2018, 23, 1555–1565. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Sample, C.H.; Davidson, T.L. The effects of a GLP-1 analog liraglutide on reward value and the learned inhibition of appetitive behavior in male and female rats. Int. J. Obes. 2019, 43, 1875–1879. [Google Scholar] [CrossRef]
- Noble, E.E.; Wang, Z.; Liu, C.M.; Davis, E.A.; Suarez, A.N.; Stein, L.M.; Tsan, L.; Terrill, S.J.; Hsu, T.M.; Jung, A.-H.; et al. Hypothalamus-hippocampus circuitry regulates impulsivity via melanin-concentrating hormone. Nat. Commun. 2019, 10, 4923. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, R.; Kretschmer, A.C.; Rigoux, L.; Albus, K.; Thanarajah, S.E.; Sitnikow, T.; Melzer, C.; Cornely, O.A.; Brüning, J.C.; Tittgemeyer, M. GLP-1 and hunger modulate incentive motivation depending on insulin sensitivity in humans. Mol. Metab. 2021, 45, 101163. [Google Scholar] [CrossRef] [PubMed]
- Miras, A.D.; Jackson, R.N.; Jackson, S.N.; Goldstone, A.P.; Olbers, T.; Hackenberg, T.; Spector, A.C.; Le Roux, C.W. Gastric bypass surgery for obesity decreases the reward value of a sweet-fat stimulus as assessed in a progressive ratio task. Am. J. Clin. Nutr. 2012, 96, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, A.P.; Miras, A.D.; Scholtz, S.; Jackson, S.; Neff, K.J.; Pénicaud, L.; Geoghegan, J.; Chhina, N.; Durighel, G.; Bell, J.D.; et al. Link between increased satiety gut hormones and reduced food reward after gastric bypass surgery for obesity. J. Clin. Endocrinol. Metab. 2016, 101, 599–609. [Google Scholar] [CrossRef]
- Elliott, J.A.; Docherty, N.G.; Haag, J.; Eckhardt, H.G.; Ravi, N.; Reynolds, J.V.; Le Roux, C.W. Attenuation of satiety gut hormones increases appetitive behavior after curative esophagectomy for esophageal cancer. Am. J. Clin. Nutr. 2019, 109, 335–344. [Google Scholar] [CrossRef]
- Disse, E.; Bussier, A.L.; Veyrat-Durebex, C.; Deblon, N.; Pfluger, P.T.; Tschöp, M.H.; Laville, M.; Rohner-Jeanrenaud, F. Peripheral ghrelin enhances sweet taste food consumption and preference, regardless of its caloric content. Physiol. Behav. 2010, 101, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, J.R.; Francomacaro, L.M.; Bohidar, A.E.; Young, K.A.; Pesarchick, B.F.; Buirkle, J.M.; McMahon, E.K.; O’Bryan, C.M. Ghrelin receptor (GHS-R1A) antagonism alters preference for ethanol and sucrose in a concentration-dependent manner in prairie voles. Physiol. Behav. 2016, 155, 231–236. [Google Scholar] [CrossRef]
- Lockie, S.H.; Dinan, T.; Lawrence, A.J.; Spencer, S.J.; Andrews, Z.B. Diet-induced obesity causes ghrelin resistance in reward processing tasks. Psychoneuroendocrinology 2015, 62, 114–120. [Google Scholar] [CrossRef]
- Pierre, A.; Regin, Y.; Van Schuerbeek, A.; Fritz, E.M.; Muylle, K.; Beckers, T.; Smolders, I.J.; Singewald, N.; De Bundel, D. Effects of disrupted ghrelin receptor function on fear processing, anxiety and saccharin preference in mice. Psychoneuroendocrinology 2019, 110. [Google Scholar] [CrossRef] [PubMed]
- Furudono, Y.; Ando, C.; Yamamoto, C.; Kobashi, M.; Yamamoto, T. Involvement of specific orexigenic neuropeptides in sweetener-induced overconsumption in rats. Behav. Brain Res. 2006, 175, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Schéle, E.; Bake, T.; Rabasa, C.; Dickson, S.L. Centrally administered ghrelin acutely influences food choice in rodents. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- Yamaguchi, E.; Yasoshima, Y.; Shimura, T. Systemic administration of anorexic gut peptide hormones impairs hedonic-driven sucrose consumption in mice. Physiol. Behav. 2017, 171, 158–164. [Google Scholar] [CrossRef]
- Alhadeff, A.L.; Rupprecht, L.E.; Hayes, M.R. GLP-1 neurons in the nucleus of the solitary tract project directly to the ventral tegmental area and nucleus accumbens to control for food intake. Endocrinology 2012, 153, 647–658. [Google Scholar] [CrossRef]
- Mietlicki-Baase, E.G.; McGrath, L.E.; Koch-Laskowski, K.; Krawczyk, J.; Pham, T.; Lhamo, R.; Reiner, D.J.; Hayes, M.R. Hindbrain DPP-IV inhibition improves glycemic control and promotes negative energy balance. Physiol. Behav. 2017, 173, 9–14. [Google Scholar] [CrossRef]
- Hyde, K.M.; Blonde, G.D.; le Roux, C.W.; Spector, A.C. Liraglutide suppression of caloric intake competes with the intake-promoting effects of a palatable cafeteria diet, but does not impact food or macronutrient selection. Physiol. Behav. 2017, 177, 4–12. [Google Scholar] [CrossRef]
- Monteleone, P.; Piscitelli, F.; Scognamiglio, P.; Monteleone, A.M.; Canestrelli, B.; Di Marzo, V.; Maj, M. Hedonic eating is associated with increased peripheral levels of ghrelin and the endocannabinoid 2-arachidonoyl-glycerol in healthy humans: A pilot study. J. Clin. Endocrinol. Metab. 2012, 97. [Google Scholar] [CrossRef]
- Monteleone, P.; Scognamiglio, P.; Monteleone, A.M.; Perillo, D.; Canestrelli, B.; Maj, M. Gastroenteric hormone responses to hedonic eating in healthy humans. Psychoneuroendocrinology 2013, 38, 1435–1441. [Google Scholar] [CrossRef]
- Basolo, A.; Heinitz, S.; Stinson, E.J.; Begaye, B.; Hohenadel, M.; Piaggi, P.; Krakoff, J.; Votruba, S.B. Fasting glucagon-like peptide 1 concentration is associated with lower carbohydrate intake and increases with overeating. J. Endocrinol. Investig. 2019, 42, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Hankir, M.K.; Seyfried, F.; Hintschich, C.A.; Diep, T.A.; Kleberg, K.; Kranz, M.; Deuther-Conrad, W.; Tellez, L.A.; Rullmann, M.; Patt, M.; et al. Gastric Bypass Surgery Recruits a Gut PPAR-α-Striatal D1R Pathway to Reduce Fat Appetite in Obese Rats. Cell Metab. 2017, 25, 335–344. [Google Scholar] [CrossRef]
- Søndergaard Nielsen, M.; Rasmussen, S.; Just Christensen, B.; Ritz, C.; le Roux, C.W.; Berg Schmidt, J.; Sjödin, A. Bariatric Surgery Does Not Affect Food Preferences, but Individual Changes in Food Preferences May Predict Weight Loss. Obesity 2018, 26, 1879–1887. [Google Scholar] [CrossRef] [PubMed]
- Kittrell, H.; Graber, W.; Mariani, E.; Czaja, K.; Hajnal, A.; Di Lorenzo, P.M. Taste and odor preferences following roux-en-Y surgery in humans. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Le Roux, C.W.; Bueter, M.; Theis, N.; Werling, M.; Ashrafian, H.; Löwenstein, C.; Athanasiou, T.; Bloom, S.R.; Spector, A.C.; Olbers, T.; et al. Gastric bypass reduces fat intake and preference. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R1057. [Google Scholar] [CrossRef]
- Shin, Y.K.; Martin, B.; Kim, W.; White, C.M.; Ji, S.; Sun, Y.; Smith, R.G.; Sévigny, J.; Tschöp, M.H.; Maudsley, S.; et al. Ghrelin is produced in taste cells and ghrelin receptor null mice show reduced taste responsivity to salty (NaCl) and sour (Citric Acid) tastants. PLoS ONE 2010, 5, e12729. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Cong, W.N.; Daimon, C.M.; Wang, R.; Tschöp, M.H.; Sévigny, J.; Martin, B.; Maudsley, S. Altered Lipid and Salt Taste Responsivity in Ghrelin and GOAT Null Mice. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Shin, Y.K.; Martin, B.; Golden, E.; Dotson, C.D.; Maudsley, S.; Kim, W.; Jang, H.J.; Mattson, M.P.; Drucker, D.J.; Egan, J.M.; et al. Modulation of taste sensitivity by GLP-1 signaling. J. Neurochem. 2008, 106, 455–463. [Google Scholar] [CrossRef]
- Martin, B.; Dotson, C.D.; Shin, Y.K.; Ji, S.; Drucker, D.J.; Maudsley, S.; Munger, S.D. Modulation of taste sensitivity by GLP-1 signaling in taste buds. In Annals of the New York Academy of Sciences; Wiley-Blackwell: Hoboken, NJ, USA, 2009; Volume 1170, pp. 98–101. [Google Scholar]
- Takai, S.; Yasumatsu, K.; Inoue, M.; Iwata, S.; Yoshida, R.; Shigemura, N.; Yanagawa, Y.; Drucker, D.J.; Margolskee, R.F.; Ninomiya, Y. Glucagon-like peptide-1 is specifically involved in sweet taste transmission. FASEB J. 2015, 29, 2268–2280. [Google Scholar] [CrossRef]
- Asarian, L.; Corp, E.S.; Hrupka, B.; Geary, N. Intracerebroventricular glucagon-like peptide-1 (7-36) amide inhibits sham feeding in rats without eliciting satiety. Physiol. Behav. 1998. [Google Scholar] [CrossRef]
- Douton, J.E.; Norgren, R.; Grigson, P.S. Effects of a glucagon-like peptide-1 analog on appetitive and consummatory behavior for rewarding and aversive gustatory stimuli in rats. Physiol. Behav. 2021, 229. [Google Scholar] [CrossRef] [PubMed]
- Arosio, M.; Ronchi, C.L.; Beck-Peccoz, P.; Gebbia, C.; Giavoli, C.; Cappiello, V.; Conte, D.; Peracchi, M. Effects of modified sham feeding on ghrelin levels in healthy human subjects. J. Clin. Endocrinol. Metab. 2004, 89, 5101–5104. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Hsu, W.H.; Hollis, J.H. Modified sham feeding of foods with different macronutrient compositions differentially influences cephalic change of insulin, ghrelin, and NMR-based metabolomic profiles. Physiol. Behav. 2014, 135, 135–142. [Google Scholar] [CrossRef]
- Uygun, B.; Kiyici, S.; Ozmen, S.; Gul, Z.; Sigirli, D.; Cavun, S. The Association between Olfaction and Taste Functions with Serum Ghrelin and Leptin Levels in Obese Women. Metab. Syndr. Relat. Disord. 2019, 17, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Bueter, M.; Miras, A.D.; Chichger, H.; Fenske, W.; Ghatei, M.A.; Bloom, S.R.; Unwin, R.J.; Lutz, T.A.; Spector, A.C.; Le Roux, C.W. Alterations of sucrose preference after Roux-en-Y gastric bypass. Physiol. Behav. 2011, 104, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Kadouh, H.; Chedid, V.; Halawi, H.; Burton, D.D.; Clark, M.M.; Khemani, D.; Vella, A.; Acosta, A.; Camilleri, M. GLP-1 analog modulates appetite, taste preference, gut hormones, and regional body fat stores in adults with obesity. J. Clin. Endocrinol. Metab. 2020, 105, 1552. [Google Scholar] [CrossRef] [PubMed]
- Brindisi, M.C.; Brondel, L.; Meillon, S.; Barthet, S.; Grall, S.; Fenech, C.; Liénard, F.; Schlich, P.; Astruc, K.; Mouillot, T.; et al. Proof of concept: Effect of GLP-1 agonist on food hedonic responses and taste sensitivity in poor controlled type 2 diabetic patients. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 2489–2494. [Google Scholar] [CrossRef]
- Baretić, M.; Kušec, V.; Uroić, V.; Pavlić-Renar, I.; Altabas, V. Glucagon-like peptide-1 affects taste perception differently in women: A randomized, placebo-controlled crossover study. Acta Clin. Croat. 2019, 58, 240–248. [Google Scholar] [CrossRef]
- Rodriguez, J.A.; Fehrentz, J.A.; Martinez, J.; Ben Haj Salah, K.; Wellman, P.J. The GHR-R antagonist JMV 2959 neither induces malaise nor alters the malaise property of LiCl in the adult male rat. Physiol. Behav. 2018, 183, 46–48. [Google Scholar] [CrossRef]
- Song, L.; Zhu, Q.; Liu, T.; Yu, M.; Xiao, K.; Kong, Q.; Zhao, R.; Li, G.D.; Zhou, Y. Ghrelin Modulates Lateral Amygdala Neuronal Firing and Blocks Acquisition for Conditioned Taste Aversion. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Li, N.; Song, G.; Wang, Y.; Zhu, Q.; Han, F.; Zhang, C.; Zhou, Y. Blocking constitutive activity of GHSR1a in the lateral amygdala facilitates acquisition of conditioned taste aversion. Neuropeptides 2018, 68, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Zhu, Q.; Han, F.; Liu, S.; Zhao, C.; Zhou, Y. Local infusion of ghrelin into the lateral amygdala blocks extinction of conditioned taste aversion in rats. Neurosci. Lett. 2018, 662, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Abegg, K.; Bernasconi, L.; Hutter, M.; Whiting, L.; Pietra, C.; Giuliano, C.; Lutz, T.A.; Riediger, T. Ghrelin receptor inverse agonists as a novel therapeutic approach against obesity-related metabolic disease. Diabetes Obes. Metab. 2017. [Google Scholar] [CrossRef]
- Yakabi, K.; Sadakane, C.; Noguchi, M.; Ohno, S.; Ro, S.; Chinen, K.; Aoyama, T.; Sakurada, T.; Takabayashi, H.; Hattori, T. Reduced ghrelin secretion in the hypothalamus of rats due to cisplatin-induced anorexia. Endocrinology 2010, 151, 3773–3782. [Google Scholar] [CrossRef]
- Malik, N.M.; Moore, G.B.T.; Kaur, R.; Liu, Y.L.; Wood, S.L.; Morrow, R.W.; Sanger, G.J.; Andrews, P.L.R. Adaptive upregulation of gastric and hypothalamic ghrelin receptors and increased plasma ghrelin in a model of cancer chemotherapy-induced dyspepsia. Regul. Pept. 2008, 148, 33–38. [Google Scholar] [CrossRef]
- Liu, Y.L.; Malik, N.M.; Sanger, G.J.; Andrews, P.L.R. Ghrelin alleviates cancer chemotherapy-associated dyspepsia in rodents. Cancer Chemother. Pharmacol. 2006, 58, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.L.; Liu, F.; Jin, H.; Xu, L.; Guo, F.F. Involvement of ghrelin in nucleus tractus solitaries on gastric signal afferent and gastric motility in cisplatin-treated rats. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 3480–3489. [Google Scholar]
- Rudd, J.A.; Chan, S.W.; Ngan, M.P.; Tu, L.; Lu, Z.; Giuliano, C.; Lovati, E.; Pietra, C. Anti-emetic action of the brain-penetrating new ghrelin agonist, HM01, alone and in combination with the 5-HT3 antagonist, palonosetron and with the NK1 antagonist, netupitant, against cisplatin- and motion-induced emesis in Suncus murinus house musk shr. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Thiele, T.E.; Van Dijk, G.; Campfield, L.A.; Smith, F.J.; Burn, P.; Woods, S.C.; Bernstein, J.L.; Seeley, R.J. Central infusion of GLP-1, but not leptin, produces conditioned taste aversions in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1997, 272. [Google Scholar] [CrossRef]
- Kinzig, K.P.; D’Alessio, D.A.; Seeley, R.J. The diverse roles of specific GLP-1 receptors in the control of food intake and the response to visceral illness. J. Neurosci. 2002, 22, 10470–10476. [Google Scholar] [CrossRef] [PubMed]
- Baraboi, E.D.; St-Pierre, D.H.; Shooner, J.; Timofeeva, E.; Richard, D. Brain activation following peripheral administration of the GLP-1 receptor agonist exendin-4. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011. [Google Scholar] [CrossRef]
- Kanoski, S.E.; Rupprecht, L.E.; Fortin, S.M.; De Jonghe, B.C.; Hayes, M.R. The role of nausea in food intake and body weight suppression by peripheral GLP-1 receptor agonists, exendin-4 and liraglutide. Neuropharmacology 2012. [Google Scholar] [CrossRef] [PubMed]
- Punjabi, M.; Arnold, M.; Rüttimann, E.; Graber, M.; Geary, N.; Pacheco-López, G.; Langhans, W. Circulating Glucagon-like Peptide-1 (GLP-1) Inhibits Eating in Male Rats by Acting in the Hindbrain and Without Inducing Avoidance. Endocrinology 2014. [Google Scholar] [CrossRef] [PubMed]
- McMahon, L.R.; Wellman, P.J. PVN infusion of GLP-1-(7-36) amide suppresses feeding but does not induce aversion or alter locomotion in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 274. [Google Scholar] [CrossRef] [PubMed]
- Dossat, A.M.; Lilly, N.; Kay, K.; Williams, D.L. Glucagon-like peptide 1 receptors in nucleus accumbens affect food intake. J. Neurosci. 2011, 31, 14453–14457. [Google Scholar] [CrossRef] [PubMed]
- Reiner, D.J.; Leon, R.M.; McGrath, L.E.; Koch-Laskowski, K.; Hahn, J.D.; Kanoski, S.E.; Mietlicki-Baase, E.G.; Hayes, M.R. Glucagon-Like Peptide-1 Receptor Signaling in the Lateral Dorsal Tegmental Nucleus Regulates Energy Balance. Neuropsychopharmacology 2018, 43, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Tang-Christensen, M.; Larsen, P.J.; Göke, R.; Fink-Jensen, A.; Jessop, D.S.; Møller, M.; Sheikh, S.P. Central administration of GLP-1-(7-36) amide inhibits food and water intake in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1996, 271. [Google Scholar] [CrossRef] [PubMed]
- Labouesse, M.A.; Stadlbauer, U.; Weber, E.; Arnold, M.; Langhans, W.; Pacheco-López, G. Vagal Afferents Mediate Early Satiation and Prevent Flavour Avoidance Learning in Response to Intraperitoneally Infused Exendin-4. J. Neuroendocrinol. 2012. [Google Scholar] [CrossRef]
- Quigley, K.S.; Kanoski, S.; Grill, W.M.; Barrett, L.F.; Tsakiris, M. Functions of Interoception: From Energy Regulation to Experience of the Self. Trends Neurosci. 2021, 44, 29–38. [Google Scholar] [CrossRef]
- Borner, T.; Liberini, C.G.; Lutz, T.A.; Riediger, T. Brainstem GLP-1 signalling contributes to cancer anorexia-cachexia syndrome in the rat. Neuropharmacology 2018, 131, 282–290. [Google Scholar] [CrossRef] [PubMed]
- De Jonghe, B.C.; Holland, R.A.; Olivos, D.R.; Rupprecht, L.E.; Kanoski, S.E.; Hayes, M.R. Hindbrain GLP-1 receptor mediation of cisplatin-induced anorexia and nausea. Physiol. Behav. 2016, 153, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Farmer, A.D.; Ban, V.F.; Coen, S.J.; Sanger, G.J.; Barker, G.J.; Gresty, M.A.; Giampietro, V.P.; Williams, S.C.; Webb, D.L.; Hellström, P.M.; et al. Visually induced nausea causes characteristic changes in cerebral, autonomic and endocrine function in humans. J. Physiol. 2015, 593, 1183–1196. [Google Scholar] [CrossRef] [PubMed]
- Hiura, Y.; Takiguchi, S.; Yamamoto, K.; Kurokawa, Y.; Yamasaki, M.; Nakajima, K.; Miyata, H.; Fujiwara, Y.; Mori, M.; Doki, Y. Fall in plasma ghrelin concentrations after cisplatin-based chemotherapy in esophageal cancer patients. Int. J. Clin. Oncol. 2012, 17, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Ohno, T.; Yanai, M.; Ando, H.; Toyomasu, Y.; Ogawa, A.; Morita, H.; Ogata, K.; Mochiki, E.; Asao, T.; Kuwano, H. Rikkunshito, a traditional Japanese medicine, suppresses cisplatin-induced anorexia in humans. Clin. Exp. Gastroenterol. 2011, 4, 291–296. [Google Scholar] [CrossRef][Green Version]
- Hiura, Y.; Takiguchi, S.; Yamamoto, K.; Takahashi, T.; Kurokawa, Y.; Yamasaki, M.; Nakajima, K.; Miyata, H.; Fujiwara, Y.; Mori, M.; et al. Effects of ghrelin administration during chemotherapy with advanced esophageal cancer patients: A prospective, randomized, placebo-controlled phase 2 study. Cancer 2012, 118, 4785–4794. [Google Scholar] [CrossRef]
- Harada, T.; Amano, T.; Ikari, T.; Takamura, K.; Ogi, T.; Fujikane, T.; Fujita, Y.; Taima, K.; Tanaka, H.; Sasaki, T.; et al. Rikkunshito for preventing chemotherapy-induced nausea and vomiting in lung cancer patients: Results from 2 prospective, randomized phase 2 trials. Front. Pharmacol. 2018, 8. [Google Scholar] [CrossRef]
- Hamai, Y.; Yoshiya, T.; Hihara, J.; Emi, M.; Furukawa, T.; Yamakita, I.; Ibuki, Y.; Okada, M. Traditional Japanese herbal medicine rikkunshito increases food intake and plasma acylated ghrelin levels in patients with esophageal cancer treated by cisplatin-based chemotherapy. J. Thorac. Dis. 2019, 11, 2470–2478. [Google Scholar] [CrossRef]
- Wo, J.M.; Ejskjaer, N.; Hellström, P.M.; Malik, R.A.; Pezzullo, J.C.; Shaughnessy, L.; Charlton, P.; Kosutic, G.; McCallum, R.W. Randomised clinical trial: Ghrelin agonist TZP-101 relieves gastroparesis associated with severe nausea and vomiting—Randomised clinical study subset data. Aliment. Pharmacol. Ther. 2011, 33, 679–688. [Google Scholar] [CrossRef]
- Mccallum, R.W.; Lembo, A.; Esfandyari, T.; Bhandari, B.R.; Ejskjaer, N.; Cosentino, C.; Helton, N.; Mondou, E.; Quinn, J.; Rousseau, F. Phase 2b, randomized, double-blind 12-week studies of TZP-102, a ghrelin receptor agonist for diabetic gastroparesis. Neurogastroenterol. Motil. 2013, 25, e705–e717. [Google Scholar] [CrossRef]
- Camilleri, M.; McCallum, R.W.; Tack, J.; Spence, S.C.; Gottesdiener, K.; Fiedorek, F.T. Efficacy and Safety of Relamorelin in Diabetics With Symptoms of Gastroparesis: A Randomized, Placebo-Controlled Study. Gastroenterology 2017, 153, 1240–1250. [Google Scholar] [CrossRef]
- Hong, S.W.; Chun, J.; Kim, J.; Lee, J.; Lee, H.J.; Chung, H.; Cho, S.J.; Im, J.P.; Kim, S.G.; Kim, J.S. Efficacy and safety of ghrelin agonists in patients with diabetic gastroparesis: A systematic review and meta-analysis. Gut Liver 2020, 14, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Bettge, K.; Kahle, M.; Abd El Aziz, M.S.; Meier, J.J.; Nauck, M.A. Occurrence of nausea, vomiting and diarrhoea reported as adverse events in clinical trials studying glucagon-like peptide-1 receptor agonists: A systematic analysis of published clinical trials. Diabetes Obes. Metab. 2017, 19, 336–347. [Google Scholar] [CrossRef]
- Shiomi, M.; Takada, T.; Tanaka, Y.; Yajima, K.; Isomoto, A.; Sakamoto, M.; Otori, K. Clinical factors associated with the occurrence of nausea and vomiting in type 2 diabetes patients treated with glucagon-like peptide-1 receptor agonists. J. Diabetes Investig. 2019, 10, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Lean, M.E.J.; Carraro, R.; Finer, N.; Hartvig, H.; Lindegaard, M.L.; Rössner, S.; Van Gaal, L.; Astrup, A. Tolerability of nausea and vomiting and associations with weight loss in a randomized trial of liraglutide in obese, non-diabetic adults. Int. J. Obes. 2014, 38, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Rayner, C.K.; Wu, T.; Aroda, V.R.; Whittington, C.; Kanters, S.; Guyot, P.; Shaunik, A.; Horowitz, M. Gastrointestinal adverse events with insulin glargine/lixisenatide fixed-ratio combination versus glucagon-like peptide-1 receptor agonists in people with type 2 diabetes mellitus: A network meta-analysis. Diabetes Obes. Metab. 2021, 23, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Mietlicki-Baase, E.G.; Liberini, C.G.; Workinger, J.L.; Bonaccorso, R.L.; Borner, T.; Reiner, D.J.; Koch-Laskowski, K.; McGrath, L.E.; Lhamo, R.; Stein, L.M.; et al. A vitamin B12 conjugate of exendin-4 improves glucose tolerance without associated nausea or hypophagia in rodents. Diabetes Obes. Metab. 2018, 20, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Borner, T.; Workinger, J.L.; Tinsley, I.C.; Fortin, S.M.; Stein, L.M.; Chepurny, O.G.; Holz, G.G.; Wierzba, A.J.; Gryko, D.; Nexø, E.; et al. Corrination of a GLP-1 Receptor Agonist for Glycemic Control without Emesis. Cell Rep. 2020, 31. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Decarie-Spain, L.; Kanoski, S.E. Ghrelin and Glucagon-Like Peptide-1: A Gut-Brain Axis Battle for Food Reward. Nutrients 2021, 13, 977. https://doi.org/10.3390/nu13030977
Decarie-Spain L, Kanoski SE. Ghrelin and Glucagon-Like Peptide-1: A Gut-Brain Axis Battle for Food Reward. Nutrients. 2021; 13(3):977. https://doi.org/10.3390/nu13030977
Chicago/Turabian StyleDecarie-Spain, Lea, and Scott E. Kanoski. 2021. "Ghrelin and Glucagon-Like Peptide-1: A Gut-Brain Axis Battle for Food Reward" Nutrients 13, no. 3: 977. https://doi.org/10.3390/nu13030977
APA StyleDecarie-Spain, L., & Kanoski, S. E. (2021). Ghrelin and Glucagon-Like Peptide-1: A Gut-Brain Axis Battle for Food Reward. Nutrients, 13(3), 977. https://doi.org/10.3390/nu13030977