

Hepatoprotective Effect of Mixture of Dipropyl Polysulfides in Concanavalin A-Induced Hepatitis
 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Synthesis of the Mixture of Dipropyl Polysulfides (DPPS)
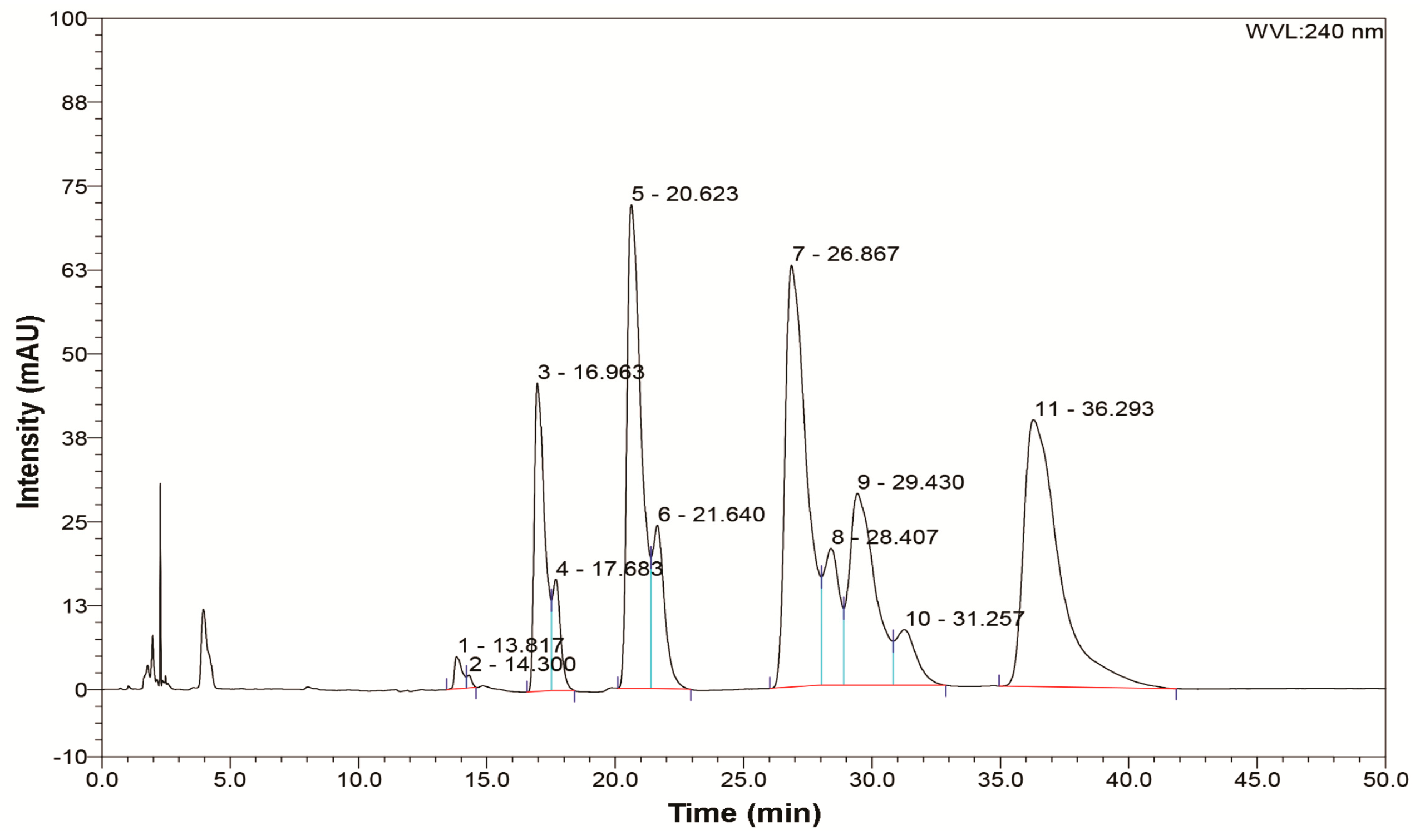
2.3. Analytical Procedure
2.4. Mice, Hepatitis Induction and DPPS Treatment
2.5. Serum Levels of Transaminases
2.6. Liver Histology and TUNEL Staining
2.7. Cytokine Measurements
2.8. Isolation of Hepatic Mononuclear Cells and Flow Cytometry
2.9. RNA Extraction and Real-Time qRT-PCR
2.10. Statistical Analysis
3. Results
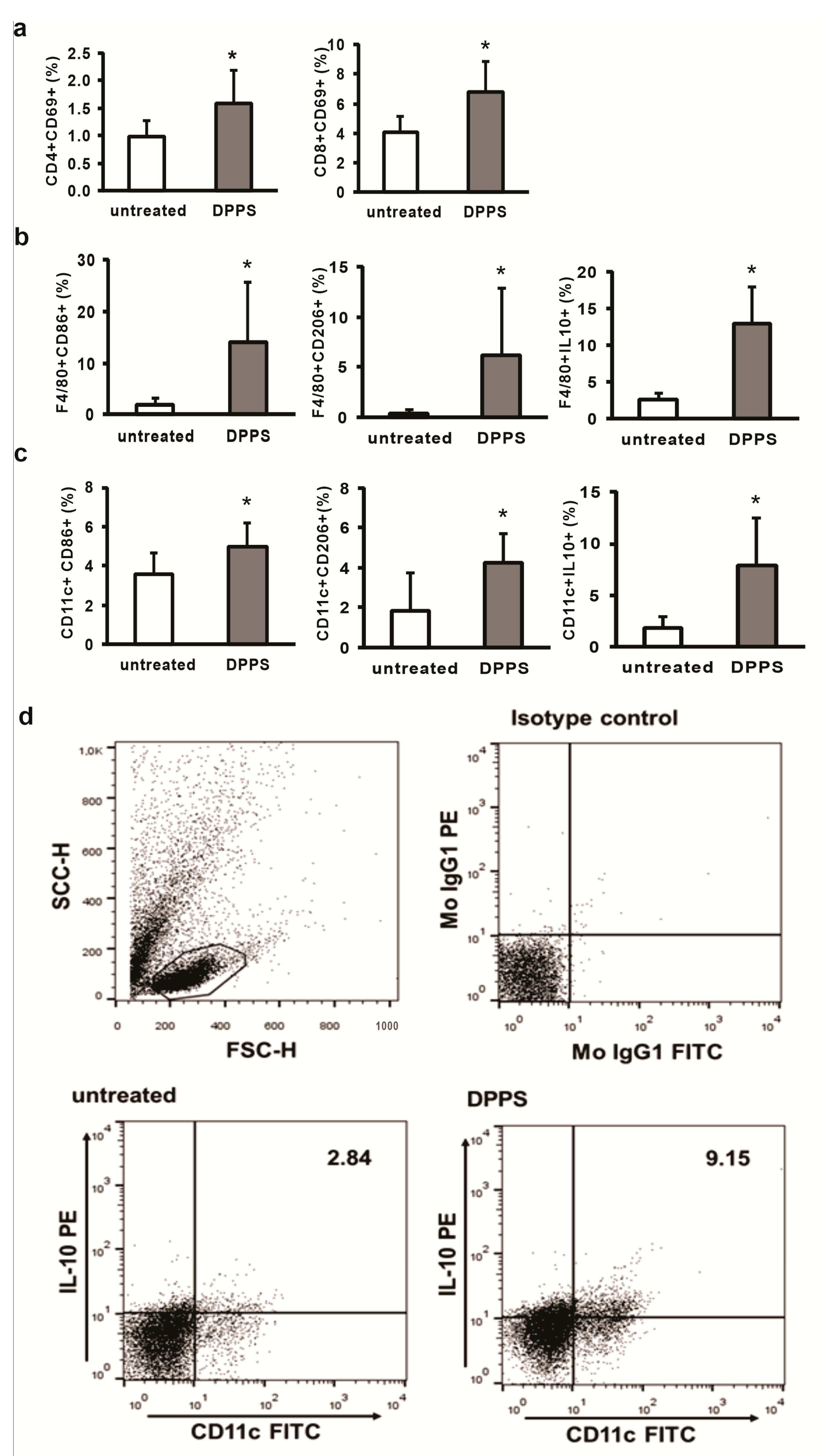
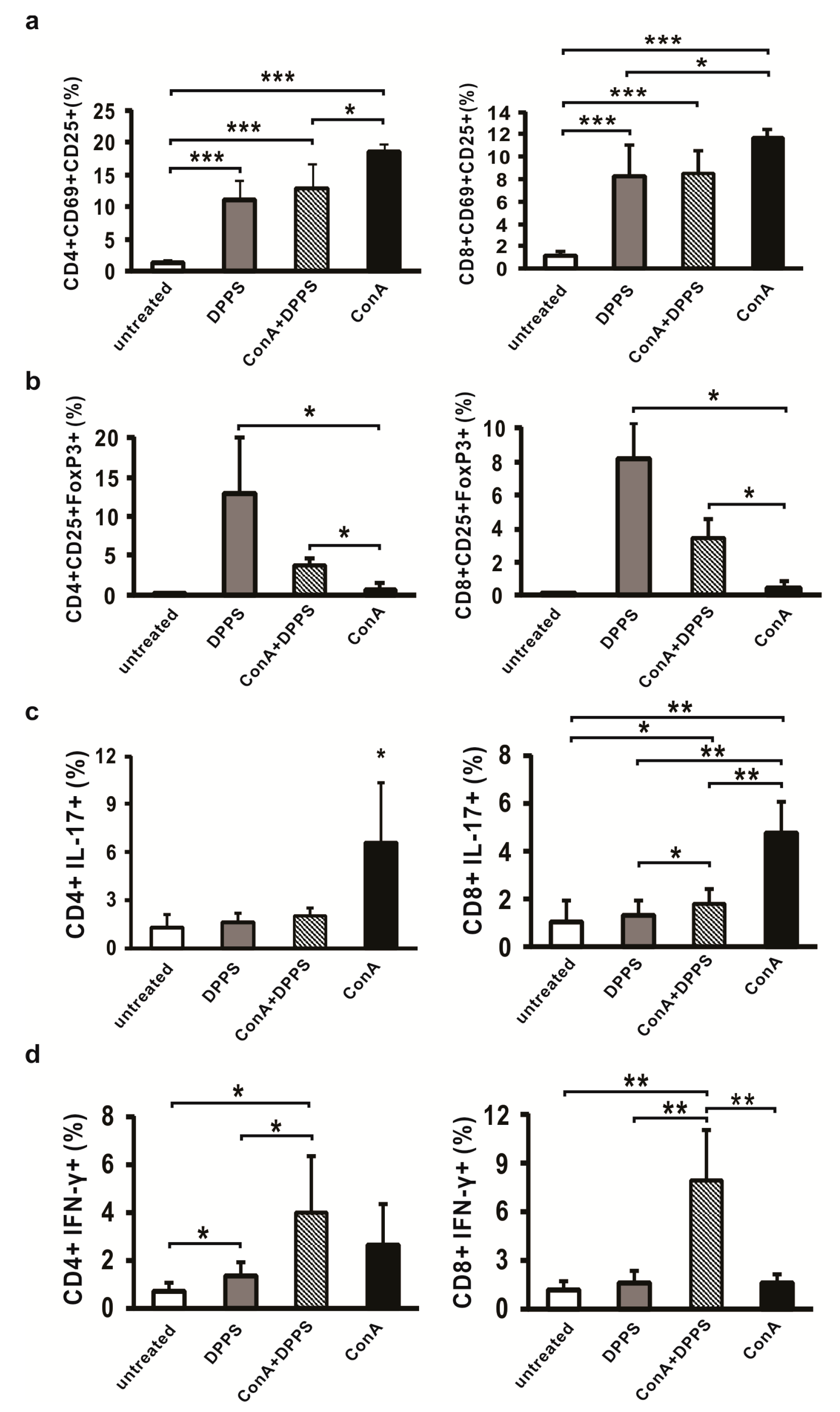
3.1. Mixture of Dipropyl Polysulfides Stimulates Activation and Regulatory Phenotype of Antigen Presenting Cells in the Liver
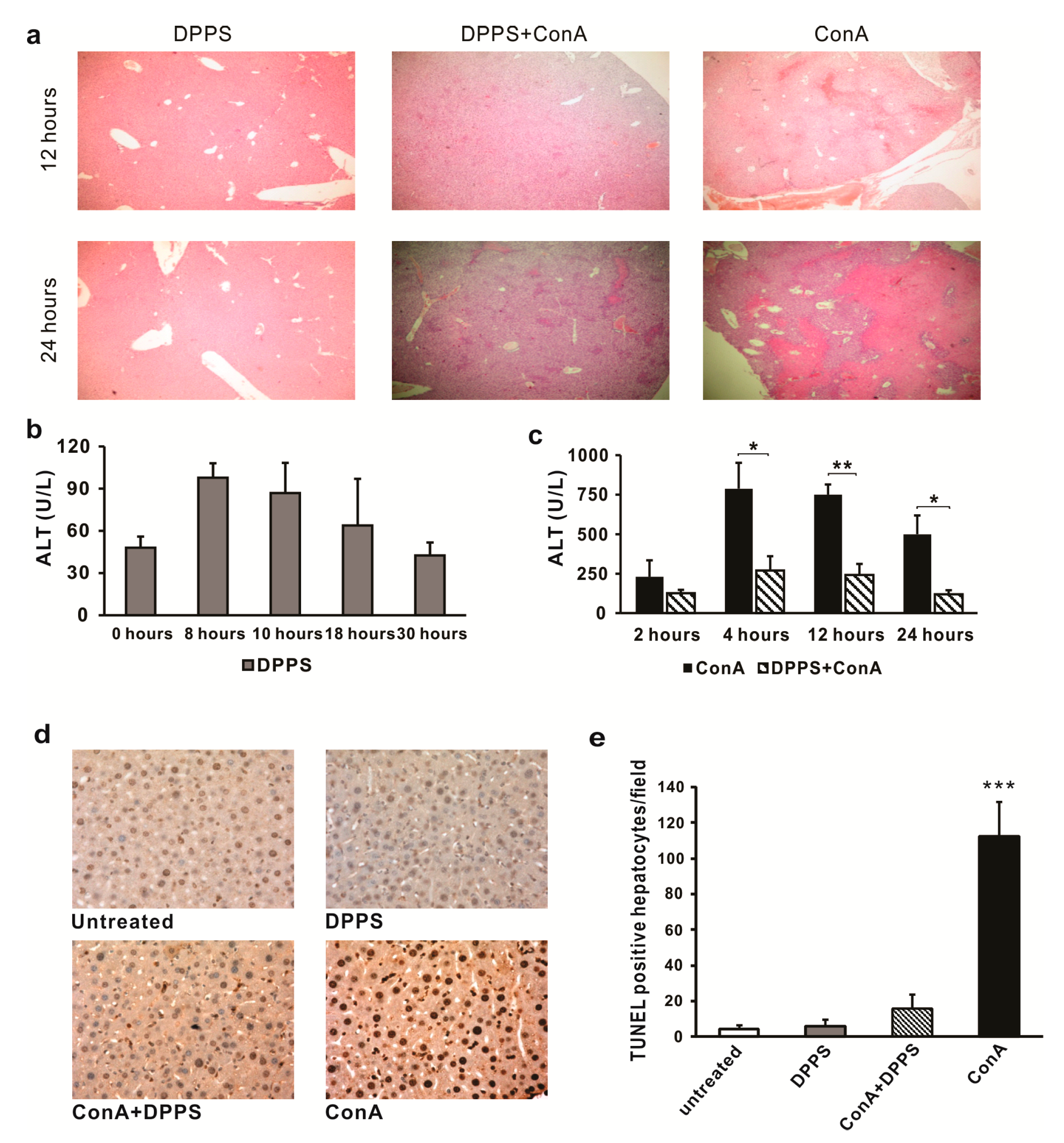
3.2. Mixture of Dipropyl Polysulfides Exerts Hepatoprotective Effects
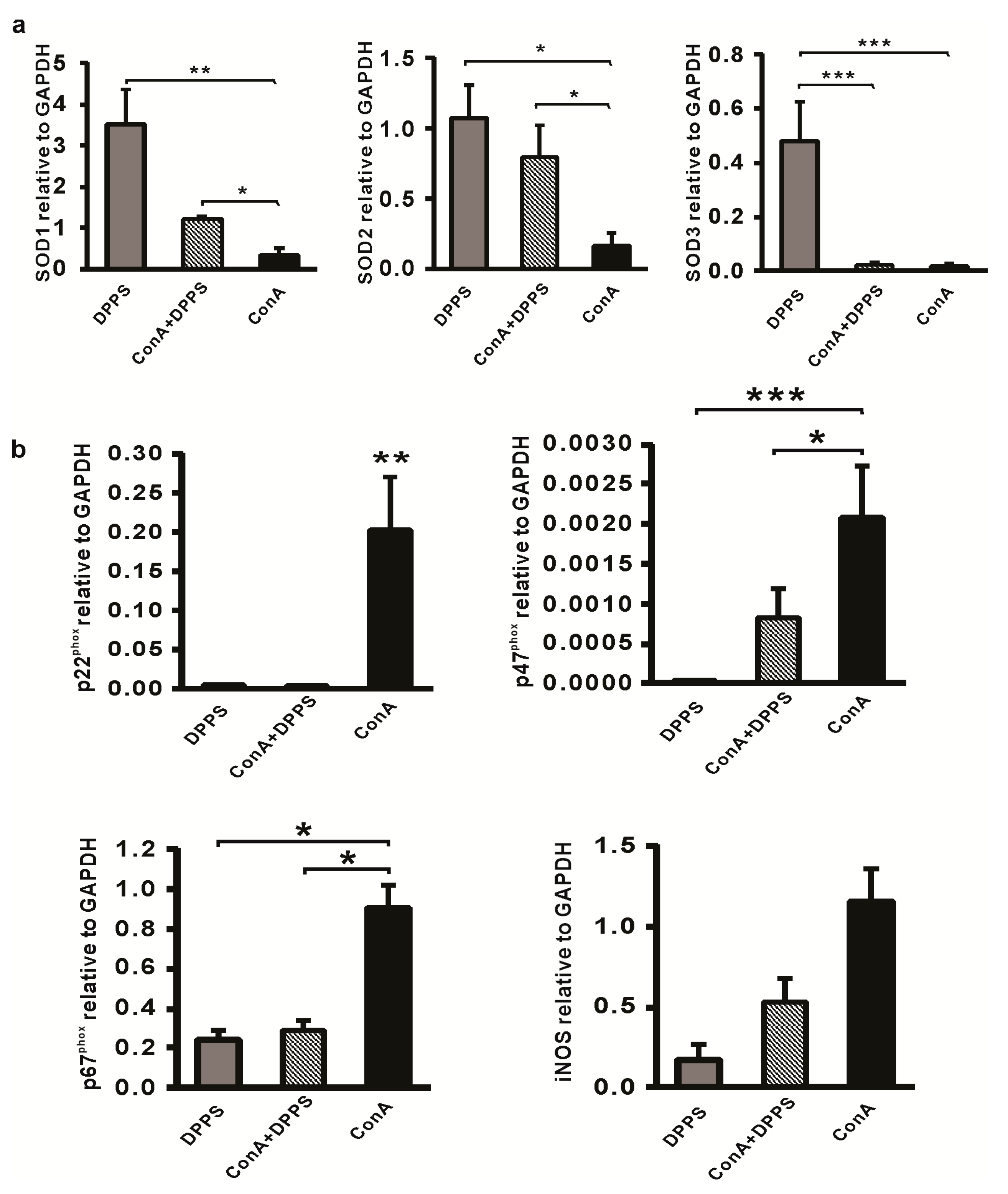
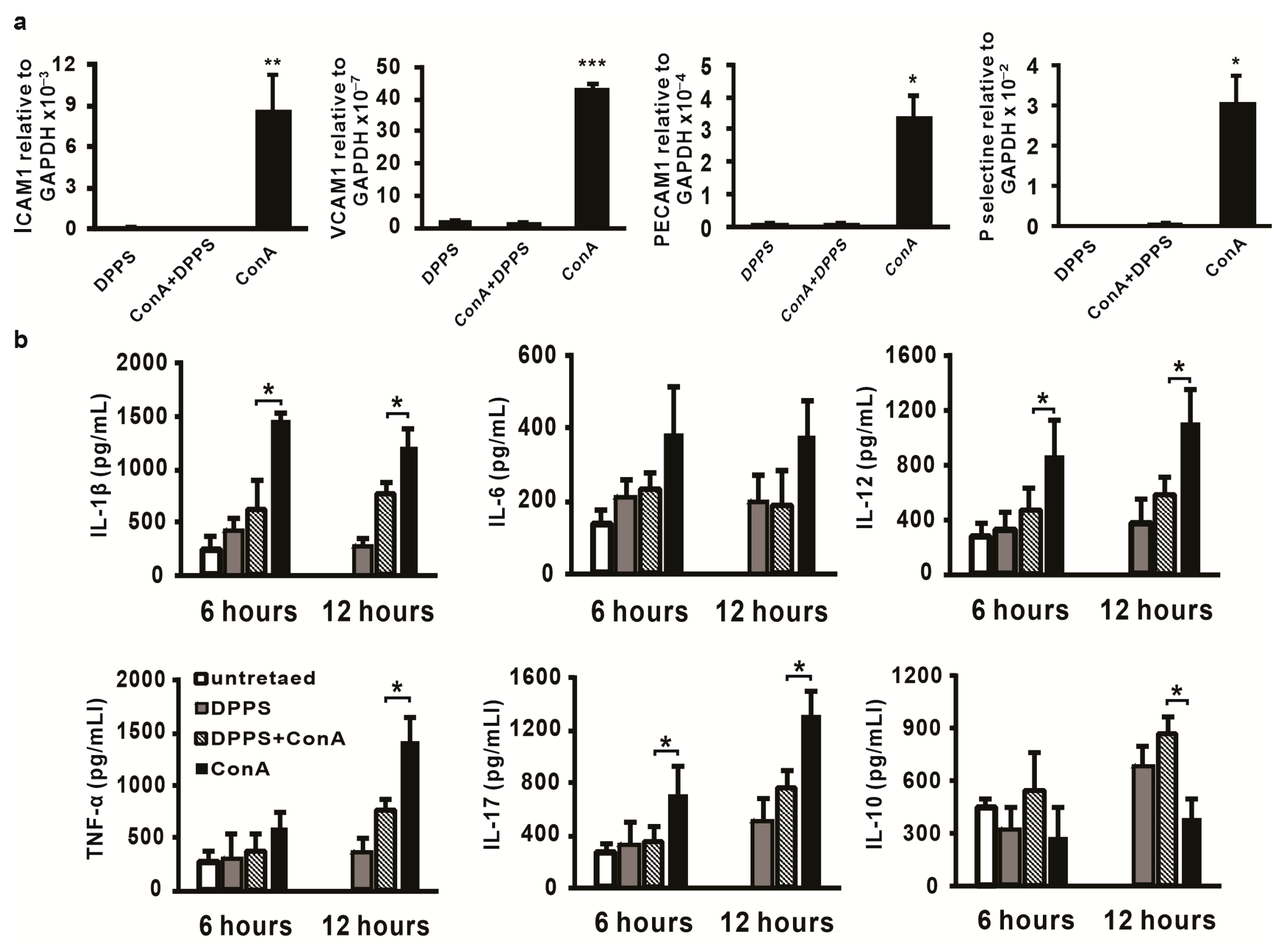
3.3. Anti-Inflammatory Activity of Mixture of Polysulfides in ConA-Induced Hepatitis
3.4. Mixture of Polysulfides Enhances Percentage of Regulatory T Cells and Attenuates Percentage of IL-17-Producing Cells in the Liver
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tiegs, G.; Hentschel, J.; Wendel, A. AT cell-dependent experimental liver injury in mice inducible by concanavalin A. J. Clin. Investig. 1992, 90, 196–203. [Google Scholar] [CrossRef]
- Heymann, F.; Hameschm, K.; Weiskirchen, R.; Tacke, F. The concanavalin A model of acute hepatitis in mice. Lab. Anim. 2015, 49, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Knolle, P.A.; Gerken, G.; Loser, E.; Dienes, H.P.; Gantner, F.; Tiegs, G.; Meyer zum Buschenfelde, K.H.; Lohse, A.W. Role of sinusoidal endothelial cells of the liver in concanavalin A-induced hepatic injury in mice. Hepatology 1996, 24, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Trautwein, C.; Rakemann, T.; Brenner, D.A.; Streetz, K.; Licato, L.; Manns, M.P.; Tiegs, G. Concanavalin A-induced liver cell damage: Activation of intracellular pathways triggered by tumor necrosis factor in mice. Gastroenterology 1998, 114, 1035–1045. [Google Scholar] [CrossRef]
- Kusters, S.; Gantner, F.; Kunstle, G.; Tiegs, G. Interferon gamma plays a critical role in T cell-dependent liver injury in mice initiated by concanavalin A. Gastroenterology 1996, 111, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Mizuhara, H.; O’Neill, E.; Seki, N.; Ogawa, T.; Kusunoki, C.; Otsuka, K.; Satoh, S.; Niwa, M.; Senoh, H.; Fujiwara, H. T cell activation-associated hepatic injury: Mediation by tumor necrosis factors and protection by interleukin 6. J. Exp. Med. 1994, 179, 1529–1537. [Google Scholar] [CrossRef]
- Gomez-Santos, L.; Luka, Z.; Wagner, C.; Fernandez-Alvarez, S.; Lu, S.C.; Mato, J.M.; Martinez-Chantar, M.L.; Beraza, N. Inhibition of natural killer cells protects the liver against acute injury in the absence of glycine N-methyltransferase. Hepatology 2012, 56, 747–759. [Google Scholar] [CrossRef]
- Gantner, F.; Leist, M.; Kusters, S.; Vogt, K.; Volk, H.D.; Tiegs, G. T cell stimulus-induced crosstalk between lymphocytes and liver macrophages results in augmented cytokine release. Exp. Cell. Res. 1996, 229, 137–146. [Google Scholar] [CrossRef]
- Nakashima, H.; Kinoshita, M.; Nakashima, M.; Habu, Y.; Shono, S.; Uchida, T.; Shinomiya, N.; Seki, S. Superoxide produced by Kupffer cells is an essential effector in concanavalin A-induced hepatitis in mice. Hepatology 2008, 48, 1979–1988. [Google Scholar] [CrossRef]
- Zeng, T.; Guo, F.F.; Zhang, C.L.; Zhao, S.; Dou, D.D.; Gao, X.C.; Xie, K.Q. The anti-fatty liver effects of garlic oil on acute ethanol-exposed mice. Chem. Biol. Interact. 2008, 176, 234–242. [Google Scholar] [CrossRef]
- Rose, P.; Whiteman, M.; Moore, P.K.; Zhu, Y.Z. Bioactive S-alk(en)yl cysteine sulfoxide metabolites in the genus Allium: The chemistry of potential therapeutic agents. Nat. Prod. Rep. 2005, 22, 351–368. [Google Scholar] [CrossRef]
- Chu, C.C.; Wu, W.S.; Shieh, J.P.; Chu, H.L.; Lee, C.P.; Duh, P.D. The Anti-Inflammatory and Vasodilating Effects of Three Selected Dietary Organic Sulfur Compounds from Allium Species. J. Funct. Biomater. 2017, 8, 5. [Google Scholar] [CrossRef]
- Lee, H.H.; Han, M.H.; Hwang, H.J.; Kim, G.Y.; Moon, S.K.; Hyun, J.W.; Kim, W.J.; Choi, Y.H. Diallyl trisulfide exerts anti-inflammatory effects in lipopolysaccharide-stimulated RAW 264.7 macrophages by suppressing the Toll-like receptor 4/nuclear factor-κB pathway. Int. J. Mol. Med. 2015, 35, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.P.; Huang, S.Y.; Chen, Y.H. Modulation of cytokine secretion by garlic oil derivatives is associated with suppressed nitric oxide production in stimulated macrophages. J. Agric. Food Chem. 2005, 53, 2530–2534. [Google Scholar] [CrossRef]
- You, S.; Nakanishi, E.; Kuwata, H.; Chen, J.; Nakasone, Y.; He, X.; He, J.; Liu, X.; Zhang, S.; Zhang, B.; et al. Inhibitory effects and molecular mechanisms of garlic organosulfur compounds on the production of inflammatory mediators. Mol. Nutr. Food Res. 2013, 57, 2049–2060. [Google Scholar] [CrossRef] [PubMed]
- Efendy, J.L.; Simmons, D.L.; Campbell, G.R.; Campbell, J.H. The effect of the aged garlic extract, ‘Kyolic’, on the development of experimental atherosclerosis. Atherosclerosis 1997, 132, 37–42. [Google Scholar] [CrossRef]
- Xiao, J.; Ching, Y.P.; Liong, E.C.; Nanji, A.A.; Fung, M.L.; Tipoe, G.L. Garlic-derived S-allylmercaptocysteine is a hepato-protective agent in non-alcoholic fatty liver disease in vivo animal model. Eur. J. Nutr. 2013, 52, 179–191. [Google Scholar] [CrossRef]
- Yu, Q.; Lee, Y.Y.; Xia, Z.Y.; Liong, E.C.; Xiao, J.; Tipoe, G.L. S-allylmercaptocysteine improves nonalcoholic steatohepatitis by enhancing AHR/NRF2-mediated drug metabolising enzymes and reducing NF-κB/IκBα and NLRP3/6-mediated inflammation. Eur. J. Nutr. 2021, 60, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Pung, D.; Leong, V.; Hebbar, V.; Shen, G.; Nair, S.; Li, W.; Kong, A.N.T. Induction of Detoxifying Enzymes by Garlic Organosulfur Compounds Through Transcription Factor Nrf2: Effect of Chemical Structure and Stress Signals. Free Radic. Biol. Med. 2004, 37, 1578–1590. [Google Scholar] [CrossRef]
- Na, H.K.; Kim, E.H.; Choi, M.A.; Park, J.M.; Kim, D.H.; Surh, Y.J. Diallyl Trisulfide Induces Apoptosis in Human Breast Cancer Cells Through ROS-mediated Activation of JNK and AP-1. Biochem. Pharmacol. 2012, 84, 1241–1250. [Google Scholar] [CrossRef]
- Münchberg, U.; Anwar, A.; Mecklenburg, S.; Jacob, C. Polysulfides as biologically active ingredients of garlic. Org. Biomol. Chem. 2007, 5, 1505–1518. [Google Scholar] [CrossRef]
- Kim, J.W.; Huh, J.E.; Kyung, S.H.; Kyung, K.H. Antimicrobial activity of alk(en)yl sulfides found in essential oils of garlic and onion. Food Sci. Biotechnol. 2004, 13, 235–239. [Google Scholar]
- Yun, H.M.; Ban, J.O.; Park, K.R.; Lee, C.K.; Jeong, H.S.; Han, S.B.; Hong, J.T. Potential therapeutic effects of functionally active compounds isolated from garlic. Pharmacol. Ther. 2014, 142, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Blom, K.G.; Qazi, M.R.; Matos, J.B.; Nelson, B.D.; DePierre, J.W.; Abedi-Valugerdi, M. Isolation of murine intrahepatic immune cells employing a modified procedure for mechanical disruption and functional characterization of the B, T and natural killer T cells obtained. Clin. Exp. Immunol. 2009, 155, 320–329. [Google Scholar] [CrossRef]
- Germain, E.; Semon, E.; Siess, M.H.; Teyssier, C. Disposition and metabolism of dipropyl disulphide in vivo in rat. Xenobiotica 2008, 38, 87–97. [Google Scholar] [CrossRef]
- Teyssier, C.; Siess, M.H. Metabolism of dipropyl disulfide by rat liver phase I and phase II enzymes and by isolated perfused rat liver. Drug Metab. Dispos. 2000, 28, 648–654. [Google Scholar]
- Rodrigues, C.; Percival, S.S. Immunomodulatory Effects of Glutathione, Garlic Derivatives, and Hydrogen Sulfide. Nutrients 2019, 11, 295. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhang, C.; Ji, J.; Wang, C.; Yang, J.; Geng, B.; Zhao, T.; Zhou, H.; Mu, X.; Pan, J.; et al. CD36 deficiency attenuates immune-mediated hepatitis in mice by modulating the proapoptotic effects of CXC chemokine ligand 10. Hepatology 2018, 67, 1943–1955. [Google Scholar] [CrossRef]
- Kim, K.M.; Chun, S.B.; Koo, M.S.; Choi, W.J.; Kim, T.W.; Kwon, Y.G.; Chung, H.T.; Billiar, T.R.; Kim, Y.M. Differential regulation of NO availability from macrophages and endothelial cells by the garlic component S-allyl cysteine. Free Radic. Biol. Med. 2001, 30, 747–756. [Google Scholar] [CrossRef]
- Cruz, C.; Correa-Rotter, R.; Sánchez-González, D.J.; Hernández-Pando, R.; Maldonado, P.D.; Martínez-Martínez, C.M.; Medina-Campos, O.N.; Tapia, E.; Aguilar, D.; Chirino, Y.I.; et al. Renoprotective and antihypertensive effects of S-allylcysteine in 5/6 nephrectomized rats. Am. J. Physiol. Ren. Physiol. 2007, 293, F1691–F1698. [Google Scholar] [CrossRef] [PubMed]
- Swart, P.J.; Hirano, T.; Kuipers, M.E.; Ito, Y.; Smit, C.; Hashida, M.; Nishikawa, M.; Beljaars, L.; Meijer, D.K.; Poelstra, K. Targeting of superoxide dismutase to the liver results in anti-inflammatory effects in rats with fibrotic livers. J. Hepatol. 1999, 31, 1034–1043. [Google Scholar] [CrossRef]
- Kawasuji, A.; Hasegawa, M.; Horikawa, M.; Fujita, T.; Matsushita, Y.; Matsushita, T.; Fujimoto, M.; Steeber, D.A.; Tedder, T.F.; Takehara, K.; et al. L-selectin and intercellular adhesion molecule-1 regulate the development of Concanavalin A-induced liver injury. J. Leukoc. Biol. 2006, 79, 696–705. [Google Scholar] [CrossRef]
- Volarevic, V.; Milovanovic, M.; Ljujic, B.; Pejnovic, N.; Arsenijevic, N.; Nilsson, U.; Leffler, H.; Lukic, M.L. Galectin-3 deficiency prevents concanavalin A-induced hepatitis in mice. Hepatology 2012, 55, 1954–1964. [Google Scholar] [CrossRef] [PubMed]
- Kato, J.; Okamoto, T.; Motoyama, H.; Uchiyama, R.; Kirchhofer, D.; Van Rooijen, N.; Enomoto, H.; Nishiguchi, S.; Kawada, N.; Fujimoto, J.; et al. Interferon-gamma-mediated tissue factor expression contributes to T-cell-mediated hepatitis through induction of hypercoagulation in mice. Hepatology 2013, 57, 362–372. [Google Scholar] [CrossRef]
- Massaguer, A.; Perez-Del-Pulgar, S.; Engel, P.; Serratosa, J.; Bosch, J.; Pizcueta, P. Concanavalin-A-induced liver injury is severely impaired in mice deficient in P-selectin. J. Leukoc. Biol. 2002, 72, 262–270. [Google Scholar]
- Jaruga, B.; Hong, F.; Kim, W.H.; Gao, B. IFN-gamma/STAT1 acts as a proinflammatory signal in T cell-mediated hepatitis via induction of multiple chemokines and adhesion molecules: A critical role of IRF-1. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G1044–G1052. [Google Scholar] [CrossRef] [PubMed]
- Chosay, J.G.; Fisher, M.A.; Farhood, A.; Ready, K.A.; Dunn, C.J.; Jaeschke, H. Role of PECAM-1 (CD31) in neutrophil transmigration in murine models of liver and peritoneal inflammation. Am. J. Physiol. 1998, 274, G776–G782. [Google Scholar] [CrossRef]
- Luo, Q.; Zhu, L.; Ding, J.; Zhuang, X.; Xu, L.; Chen, F. Protective effect of galangin in Concanavalin A-induced hepatitis in mice. Drug Des. Dev. Ther. 2015, 9, 2983–2992. [Google Scholar]
- Luo, Q.; Ding, J.; Zhu, L.; Chen, F.; Xu, L. Hepatoprotective Effect of Wedelolactone against Concanavalin A-Induced Liver Injury in Mice. Am. J. Chin. Med. 2018, 46, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Sang, X.X.; Wang, R.L.; Zhang, C.E.; Liu, S.J.; Shen, H.H.; Guo, Y.M.; Zhang, Y.M.; Niu, M.; Wang, J.B.; Bai, Z.F.; et al. Sophocarpine Protects Mice from ConA-Induced Hepatitis via Inhibition of the IFN-Gamma/STAT1 Pathway. Front. Pharmacol. 2017, 8, 140. [Google Scholar] [CrossRef]
- Luan, J.; Zhang, X.; Wang, S.; Li, Y.; Fan, J.; Chen, W.; Zai, W.; Wang, S.; Wang, Y.; Chen, M.; et al. NOD-Like Receptor Protein 3 Inflammasome-Dependent IL-1β Accelerated ConA-Induced Hepatitis. Front. Immunol. 2018, 9, 758. [Google Scholar] [CrossRef] [PubMed]
- Zenewicz, L.A.; Yancopoulos, G.D.; Valenzuela, D.M.; Murphy, A.J.; Karow, M.; Flavell, R.A. Interleukin-22 but not interleukin-17 provides protection to hepatocytes during acute liver inflammation. Immunity 2007, 27, 647–659. [Google Scholar] [CrossRef]
- Nicoletti, F.; Di Marco, R.; Zaccone, P.; Salvaggio, A.; Magro, G.; Bendtzen, K.; Meroni, P. Murine concanavalin A-induced hepatitis is prevented by interleukin 12 (IL-12) antibody and exacerbated by exogenous IL-12 through an interferon-gamma-dependent mechanism. Hepatology 2000, 32, 728–733. [Google Scholar] [CrossRef] [PubMed]
- Filliol, A.; Piquet-Pellorce, C.; Le Seyec, J.; Farooq, M.; Genet, V.; Lucas-Clerc, C.; Bertin, J.; Gough, P.J.; Dimanche-Boitrel, M.T.; Vandenabeele, P.; et al. RIPK1 protects from TNF-α-mediated liver damage during hepatitis. Cell Death Dis. 2016, 7, e2462. [Google Scholar] [CrossRef] [PubMed]
- Bruck, R.; Aeed, H.; Brazovsky, E.; Noor, T.; Hershkoviz, R. Allicin, the active component of garlic, prevents immune-mediated, concanavalin A-induced hepatic injury in mice. Liver Int. 2005, 25, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Pemberton, P.W.; Aboutwerat, A.; Smith, A.; Burrows, P.C.; McMahon, R.F.; Warnes, T.W. Oxidant stress in type I autoimmune hepatitis: The link between necroinflammation and fibrogenesis? Biochim. Biophys. Acta 2004, 1689, 182–189. [Google Scholar] [CrossRef]
- Colín-González, A.L.; Santana, R.A.; Silva-Islas, C.A.; Chánez-Cárdenas, M.E.; Santamaría, A.; Maldonado, P.D. The antioxidant mechanisms underlying the aged garlic extract- and S-allylcysteine-induced protection. Oxid. Med. Cell. Longev. 2012, 2012, 907162. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abdelkhalek, N.K.; Hassan, A.M. Antagonistic activity of dietary allicin against deltamethrin-induced oxidative damage in freshwater Nile tilapia; Oreochromis niloticus. Ecotoxicol. Environ. Saf. 2015, 111, 146–152. [Google Scholar] [CrossRef]
- Sharabi, A.; Tsokos, M.G.; Ding, Y.; Malek, T.R.; Klatzmann, D.; Tsokos, G.C. Regulatory T cells in the treatment of disease. Nat. Rev. Drug Discov. 2018, 17, 823–844. [Google Scholar] [CrossRef]
- Huang, H.; Deng, Z. Adoptive transfer of regulatory T cells stimulated by Allogeneic Hepatic Stellate Cells mitigates liver injury in mice with concanavalin A-induced autoimmune hepatitis. Biochem. Biophys. Res. Commun. 2019, 512, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Santner-Nanan, B.; Hu, M.; Skarratt, K.; Lee, C.H.; Stormon, M.; Wong, M.; Fuller, S.J.; Nanan, R. IL-10 Potentiates Differentiation of Human Induced Regulatory T Cells via STAT3 and Foxo1. J. Immunol. 2015, 195, 3665–3674. [Google Scholar] [CrossRef] [PubMed]
- Makris, A.; Thornton, C.E.; Xu, B.; Hennessy, A. Garlic increases IL-10 and inhibits TNFalpha and IL-6 production in endotoxin-stimulated human placental explants. Placenta 2005, 26, 828–834. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sense and Antisense |
|---|---|
| ICAM-1 | forward 5′-CAATTTCTCATGCCGCACAG-3′, reverse 5′-AGCTGGAAGATCGAAAGTCCG-3′ |
| VCAM-1 | forward 5′-TGAACCCAAACAGAGGCAGAGT-3′, reverse 5′-GGTATCCCATCACTTGAGCAGG-3′), |
| PECAM-1 | forward 5′-CAAACAGAAACCCGTGGAGATG-3′, reverse 5′-ACCGTAATGGCTGTTGGCTTC-3′ |
| P-selectine | forward 5′-TCATCCCGGTGAAGCAATGT3′, reverse, 5′-TGGAGAACGCAAGGACAGGTAT-3′ |
| p47phox | forward 5′-ATGACCTCAATGGCTTCACC-3′, reverse 5′-CTATCTGGAGCCCCTTGACA-3′ |
| p67phox | forward 5′-CTATCAGCTGGTTCCCACGA-3′, reverse 5′-GCAGTGGCCTACTTCCAGAG-3′ |
| iNOS | forward 5′-CGAAACGCTTCACTTCCAA-3′, reverse 5′-TGAGCCTATATTGCTGTGGCT-3′ |
| SOD1 | 5′-AATGTGTCCATTGAAGATCGTGTGA-3′, reverse 5′-GCTTCCAGCATTTCCAGTCTTTGTA-3′ |
| SOD2 | forward 5′-AGGGCCTGTCCCATGATGTC-3′, reverse 5′-AGAAACCCGTTTGCCTCTACTGAA-3′ |
| SOD3 | forward 5′-GGGTCTGTCCTGTACTTCACCAGAG-3′, reverse 5′-CTGACATGGTCCAGGTGACAGAG-3′ |
| GAPDH | forward 5′-CATCACTGCCACCCAGAAGACTG-3′, reverse 5′-ATGCCAGTGAGCTTCCCGTTCAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arsenijevic, D.; Stojanovic, B.; Milovanovic, J.; Arsenijevic, A.; Simic, M.; Pergal, M.; Kodranov, I.; Cvetkovic, O.; Vojvodic, D.; Ristanovic, E.; et al. Hepatoprotective Effect of Mixture of Dipropyl Polysulfides in Concanavalin A-Induced Hepatitis. Nutrients 2021, 13, 1022. https://doi.org/10.3390/nu13031022
Arsenijevic D, Stojanovic B, Milovanovic J, Arsenijevic A, Simic M, Pergal M, Kodranov I, Cvetkovic O, Vojvodic D, Ristanovic E, et al. Hepatoprotective Effect of Mixture of Dipropyl Polysulfides in Concanavalin A-Induced Hepatitis. Nutrients. 2021; 13(3):1022. https://doi.org/10.3390/nu13031022
Chicago/Turabian StyleArsenijevic, Dragana, Bojana Stojanovic, Jelena Milovanovic, Aleksandar Arsenijevic, Milos Simic, Marija Pergal, Igor Kodranov, Olga Cvetkovic, Danilo Vojvodic, Elizabeta Ristanovic, and et al. 2021. "Hepatoprotective Effect of Mixture of Dipropyl Polysulfides in Concanavalin A-Induced Hepatitis" Nutrients 13, no. 3: 1022. https://doi.org/10.3390/nu13031022
APA StyleArsenijevic, D., Stojanovic, B., Milovanovic, J., Arsenijevic, A., Simic, M., Pergal, M., Kodranov, I., Cvetkovic, O., Vojvodic, D., Ristanovic, E., Manojlovic, D., Milovanovic, M., & Arsenijevic, N. (2021). Hepatoprotective Effect of Mixture of Dipropyl Polysulfides in Concanavalin A-Induced Hepatitis. Nutrients, 13(3), 1022. https://doi.org/10.3390/nu13031022