
Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women



Abstract
:
1. Introduction
2. Menopause Associated Changes in Energy Metabolism
2.1. Composition of the Human Body
Changes in Body Composition in Postmenopausal Women
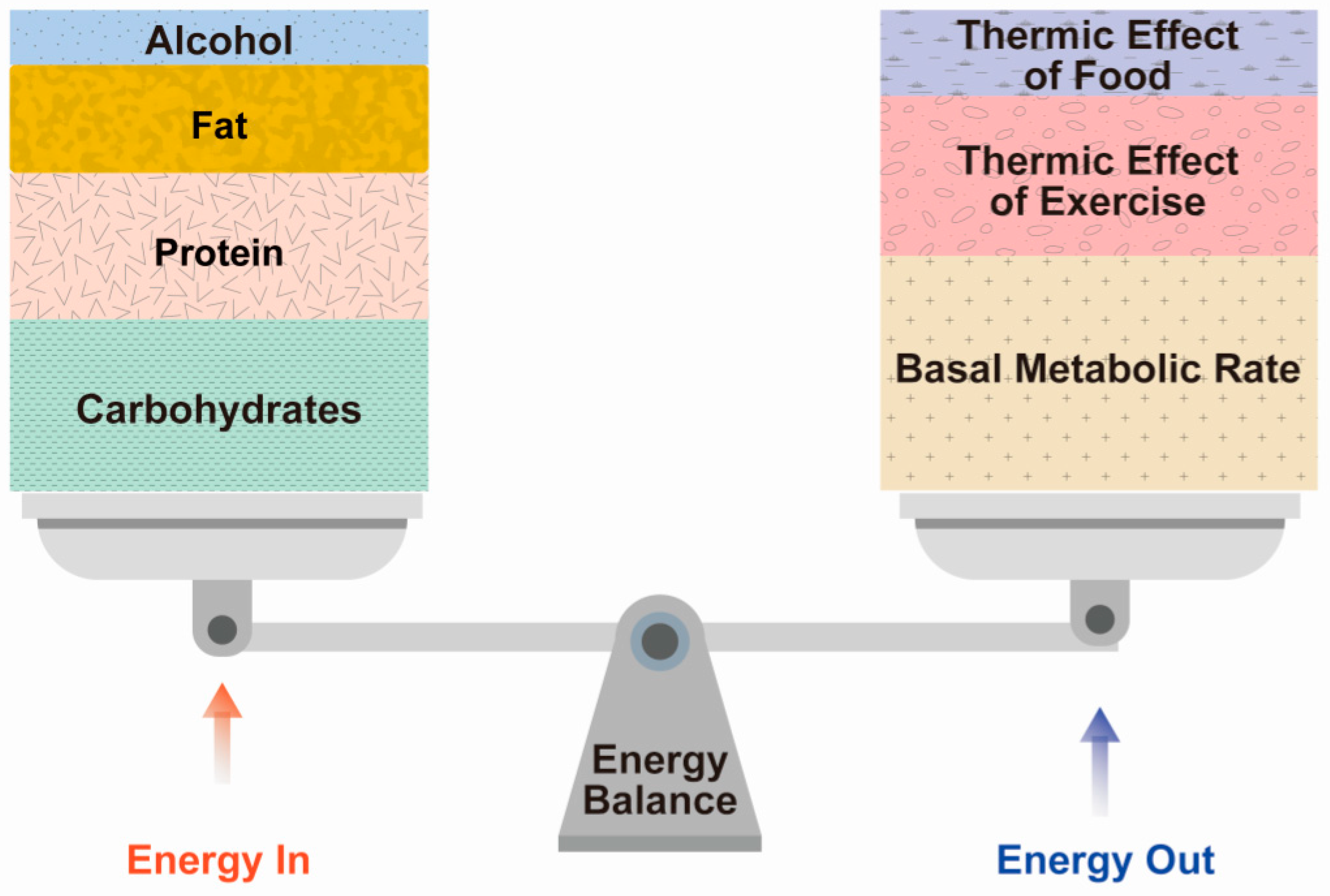
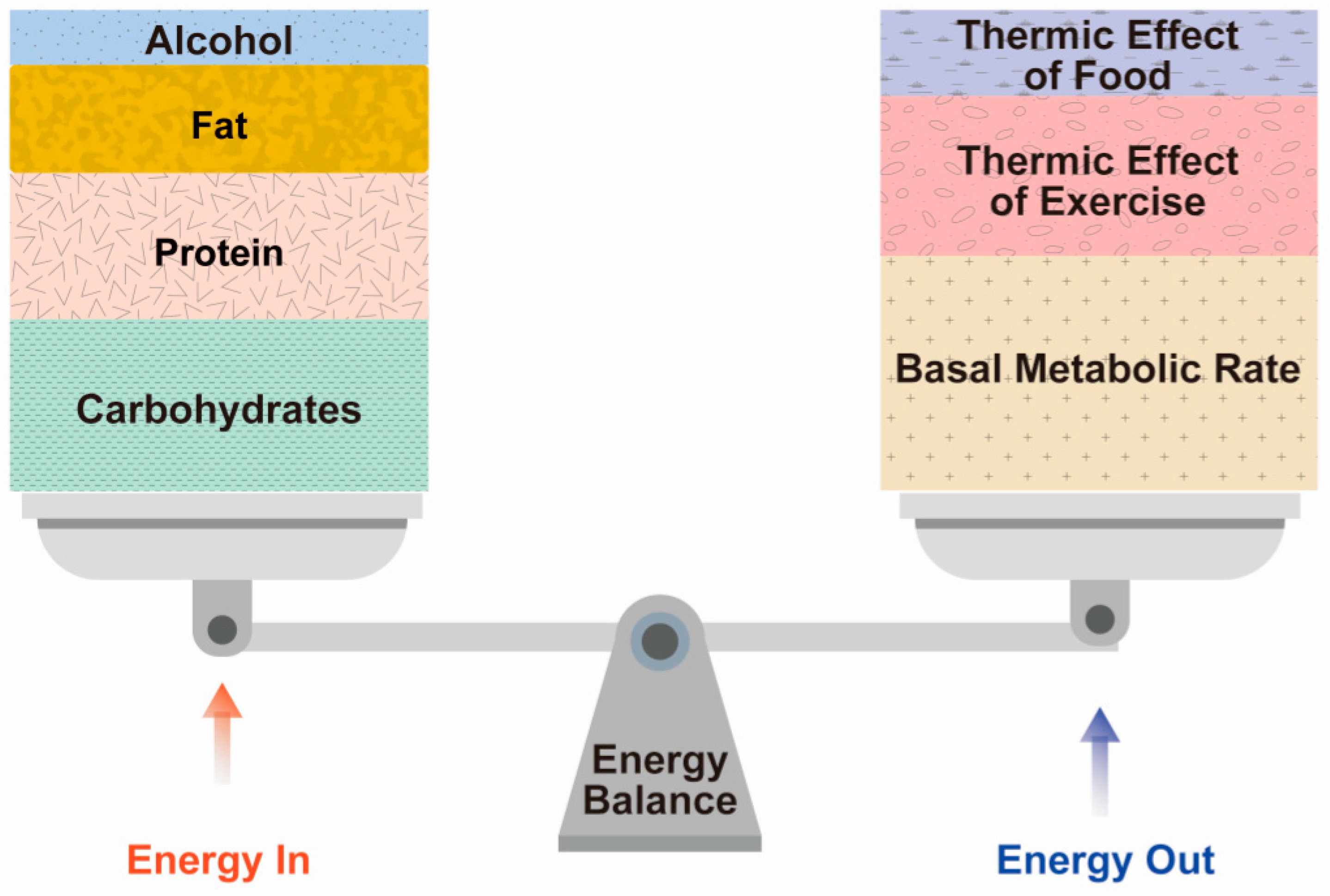
2.2. Component of Energy Expenditure
2.2.1. BMR
2.2.2. TEF
2.2.3. TEA
2.3. Metabolic Rate Difference between Skeletal Muscle and Adipose Tissue
2.3.1. Changes in Body Composition and Energy Expenditure in Postmenopausal Women
2.3.2. Sarcopenia in Postmenopausal Women
3. Background of Weight Gain in Postmenopausal Women
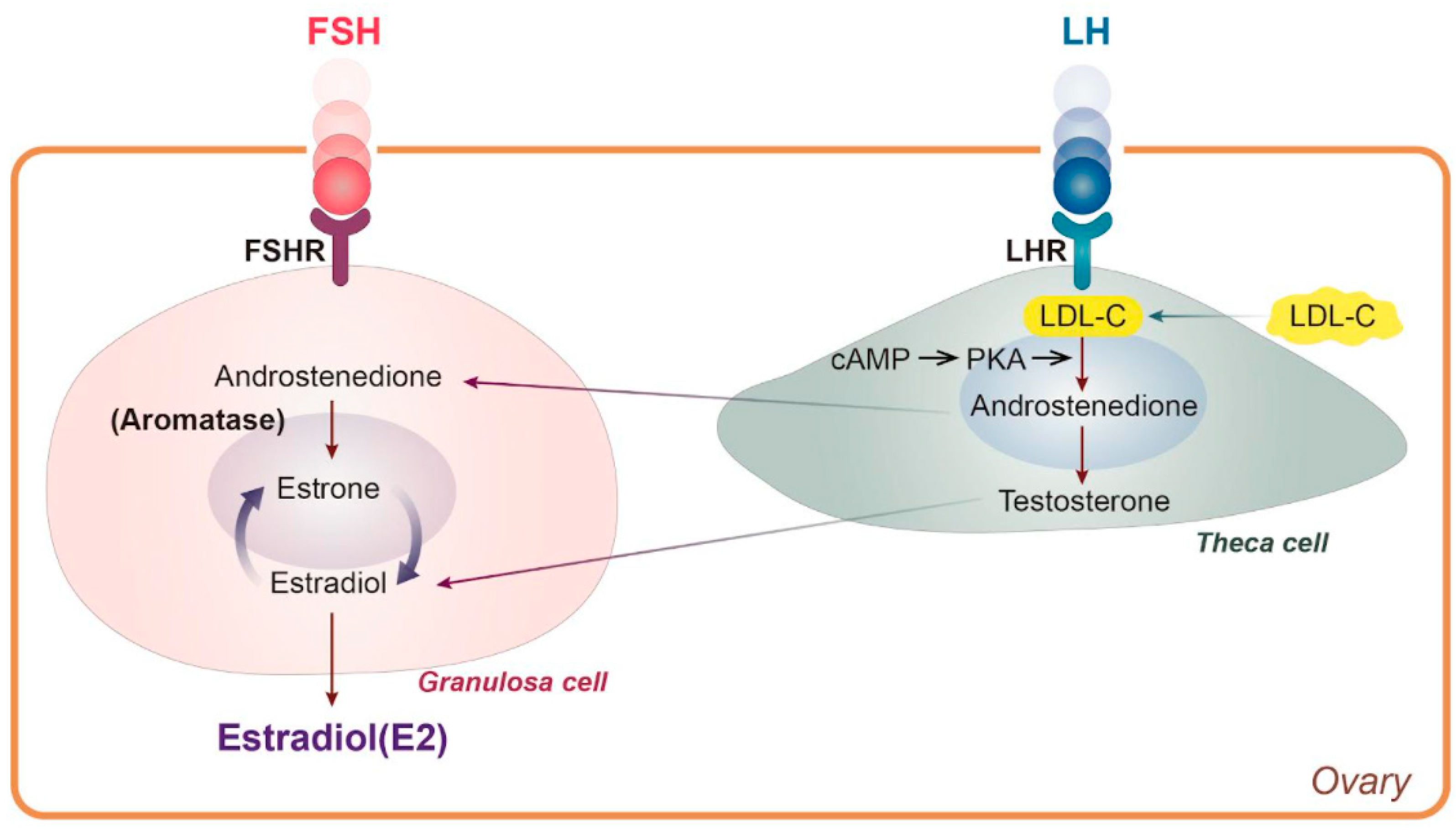
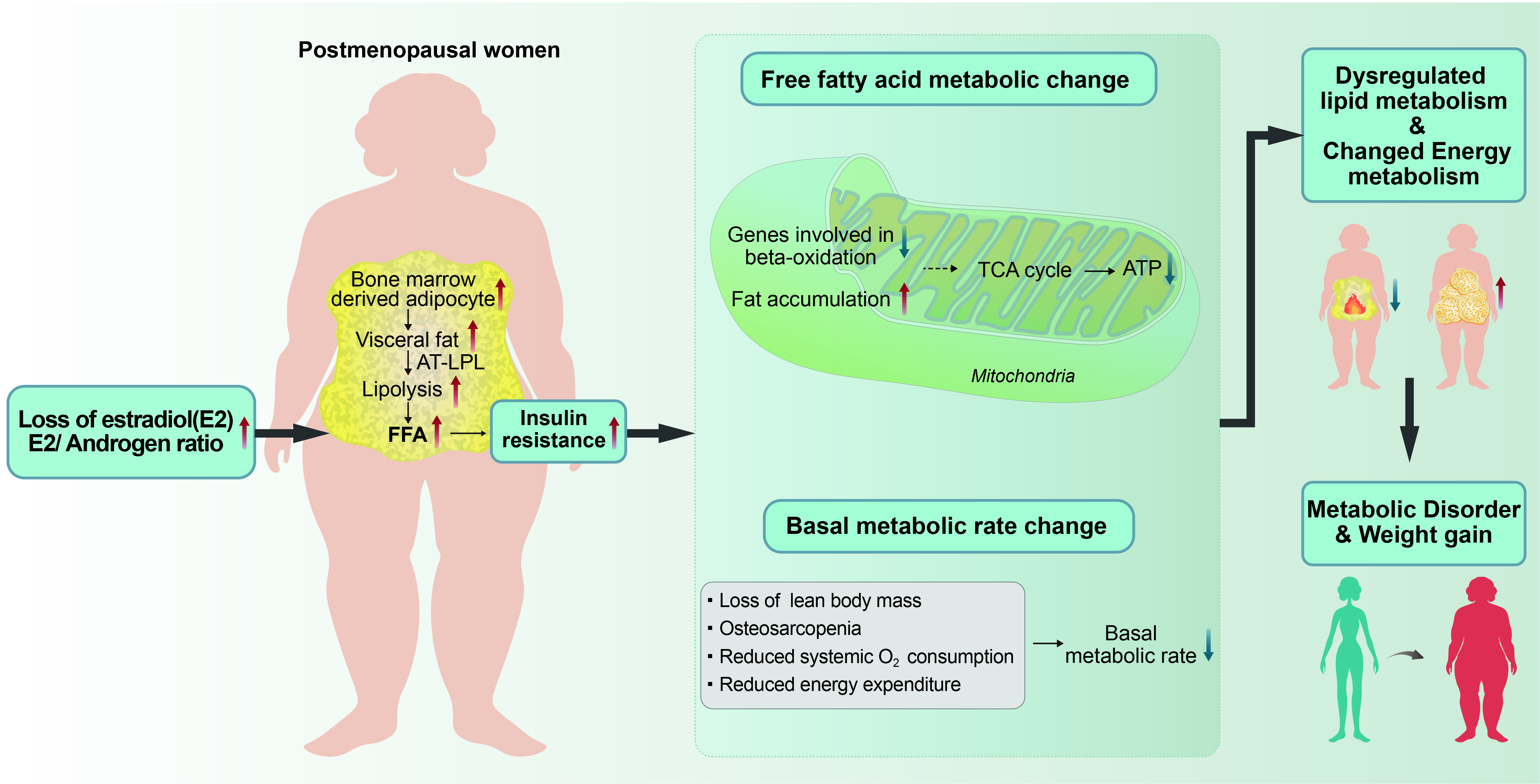
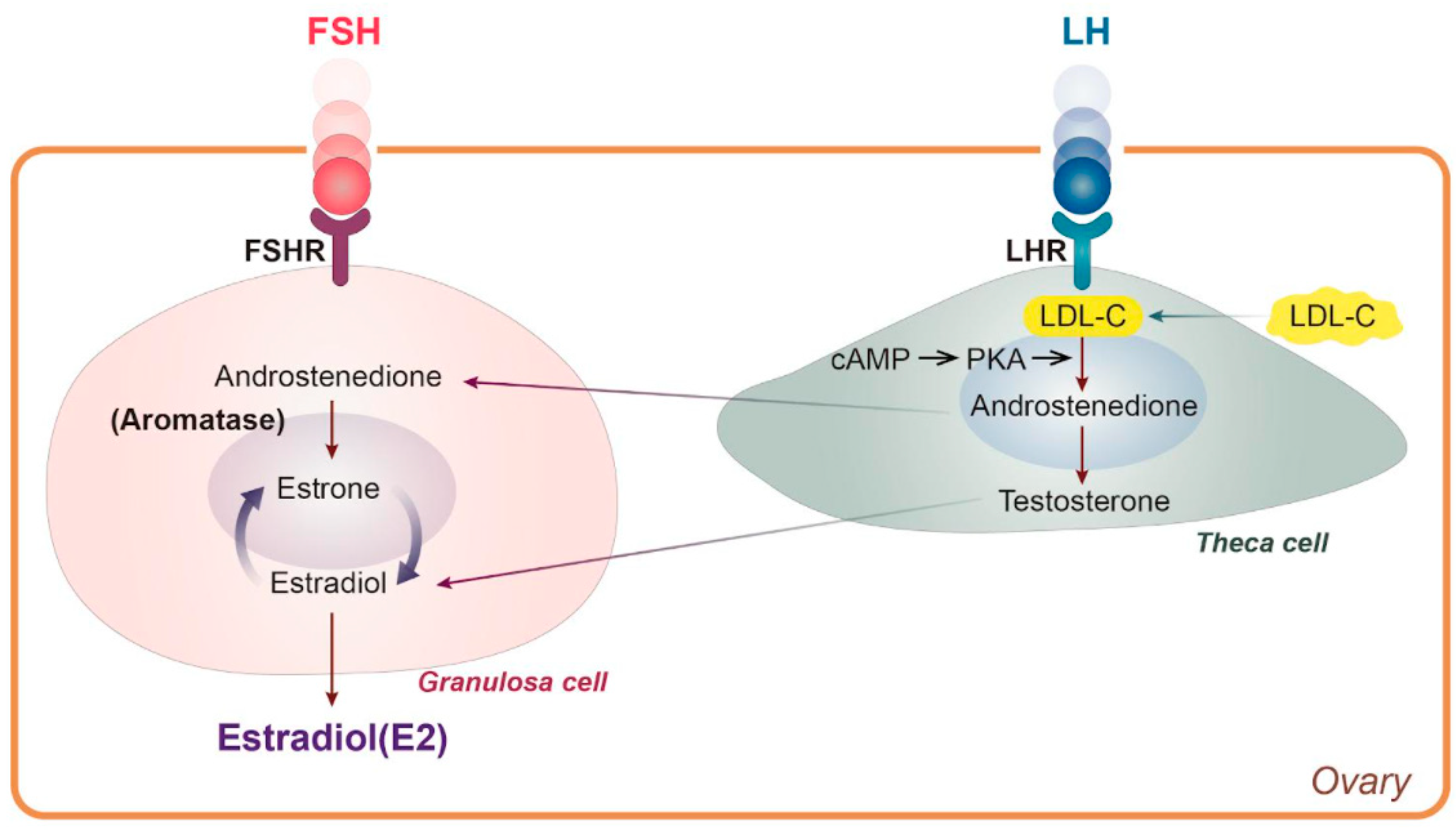
3.1. Lipid Metabolic Abnormality Due to E2 Hormonal Change
3.2. Fat Redistribution in Postmenopausal Women
3.3. Excessive Visceral Abdominal Fat and Metabolic Alterations
3.4. Alterations in Fatty Acid Metabolism
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Landgren, B.M.; Collins, A.; Csemiczky, G.; Burger, H.G.; Baksheev, L.; Robertson, D.M. Menopause transition: Annual changes in serum hormonal patterns over the menstrual cycle in women during a nine-year period prior to menopause. J. Clin. Endocrinol. Metab. 2004, 89, 2763–2769. [Google Scholar] [CrossRef]
- Yoshida, T.; Takahashi, K.; Yamatani, H.; Takata, K.; Kurachi, H. Impact of surgical menopause on lipid and bone metabolism. Climacteric 2011, 14, 445–452. [Google Scholar] [CrossRef]
- Takahashi, T.A.; Johnson, K.M. Menopause. Med. Clin. North Am. 2015, 99, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Patel, S. Disruption of aromatase homeostasis as the cause of a multiplicity of ailments: A comprehensive review. J. Steroid Biochem. Mol. Biol. 2017, 168, 19–25. [Google Scholar] [CrossRef]
- Cui, J.; Shen, Y.; Li, R. Estrogen synthesis and signaling pathways during aging: From periphery to brain. Trends Mol. Med. 2013, 19, 197–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewailly, D.; Robin, G.; Peigne, M.; Decanter, C.; Pigny, P.; Catteau-Jonard, S. Interactions between androgens, FSH, anti-Müllerian hormone and estradiol during folliculogenesis in the human normal and polycystic ovary. Hum. Reprod. Update 2016, 22, 709–724. [Google Scholar] [CrossRef] [Green Version]
- Cervellati, C.; Bergamini, C.M. Oxidative damage and the pathogenesis of menopause related disturbances and diseases. Clin. Chem. Lab. Med. 2016, 54, 739–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, S.H.; Kim, H.S. Menopause-Associated Lipid Metabolic Disorders and Foods Beneficial for Postmenopausal Women. Nutrients 2020, 12, 202. [Google Scholar] [CrossRef] [Green Version]
- Ozbey, N.; Sencer, E.; Molvalilar, S.; Orhan, Y. Body fat distribution and cardiovascular disease risk factors in pre-and postmenopausal obese women with similar BMI. Endocr. J. 2002, 49, 503–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldarine, V.T.; Pedroso, A.P.; Brandão-Teles, C.; LoTurco, E.G.; Nascimento, C.M.O.; Oyama, L.M.; Bueno, A.A.; Martins-de-Souza, D.; Ribeiro, E.B. Ovariectomy modifies lipid metabolism of retroperitoneal white fat in rats: A proteomic approach. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E427–E437. [Google Scholar] [CrossRef] [PubMed]
- Maltais, M.L.; Desroches, J.; Dionne, I.J. Changes in muscle mass and strength after menopause. J. Musculoskelet. Neuronal Interact. 2009, 9, 186–197. [Google Scholar]
- Kapoor, E.; Collazo-Clavell, M.L.; Faubion, S.S. Weight Gain in Women at Midlife: A Concise Review of the Pathophysiology and Strategies for Management. Mayo Clin. Proc. 2017, 92, 1552–1558. [Google Scholar] [CrossRef]
- Pannemans, D.L.; Westerterp, K.R. Energy expenditure, physical activity and basal metabolic rate of elderly subjects. Br. J. Nutr. 1995, 73, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Gropper, S.S.; Smith, J.L. Advanced Nutrition and Human Metabolism, 4th ed.; Howe, E., Ed.; Cengage Learning: Boston, MA, USA, 2005. [Google Scholar]
- Lemos, T.; Gallagher, D. Current body composition measurement techniques. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 310–314. [Google Scholar] [CrossRef]
- Luke, A.; Schoeller, D.A. Basal metabolic rate, fat-free mass, and body cell mass during energy restriction. Metabolism 1992, 41, 450–456. [Google Scholar] [CrossRef]
- Choi, B.; Steiss, D.; Garcia-Rivas, J.; Kojaku, S.; Schnall, P.; Dobson, M.; Baker, D. Comparison of body mass index with waist circumference and skinfold-based percent body fat in firefighters: Adiposity classification and associations with cardiovascular disease risk factors. Int. Arch. Occup. Environ. Health 2016, 89, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Greendale, G.A.; Sternfeld, B.; Huang, M.; Han, W.; Karvonen-Gutierrez, C.; Ruppert, K.; Cauley, J.A.; Finkelstein, J.S.; Jiang, S.F.; Karlamangla, A.S. Changes in body composition and weight during the menopause transition. JCI Insight 2019, 4, e124865. [Google Scholar] [CrossRef]
- Ambikairajah, A.; Walsh, E.; Tabatabaei-Jafari, H.; Cherbuin, N. Fat mass changes during menopause: A metaanalysis. Am. J. Obstet. Gynecol. 2019, 221, 393–409.e350. [Google Scholar] [CrossRef]
- Janssen, I.; Powell, L.H.; Jasielec, M.S.; Kazlauskaite, R. Covariation of change in bioavailable testosterone and adiposity in midlife women. Obesity 2015, 23, 488–494. [Google Scholar] [CrossRef]
- Kaye, S.A.; Folsom, A.R.; Prineas, R.J.; Potter, J.D.; Gapstur, S.M. The association of body fat distribution with lifestyle and reproductive factors in a population study of postmenopausal women. Int. J. Obes. 1990, 14, 583–591. [Google Scholar] [CrossRef]
- Pasquali, R.; Casimirri, F.; Labate, A.M.; Tortelli, O.; Pascal, G.; Anconetani, B.; Gatto, M.R.; Flamia, R.; Capelli, M.; Barbara, L. Body weight, fat distribution and the menopausal status in women. The VMH Collaborative Group. Int. J. Obes. Relat. Metab. Disord. 1994, 18, 614–621. [Google Scholar]
- Ijuin, H.; Douchi, T.; Oki, T.; Maruta, K.; Nagata, Y. The contribution of menopause to changes in body-fat distribution. J. Obstet. Gynaecol. Res. 1999, 25, 367–372. [Google Scholar] [CrossRef]
- Douchi, T.; Kuwahata, R.; Yamasaki, H.; Yamamoto, S.; Oki, T.; Nakae, M.; Nagata, Y. Inverse relationship between the changes in trunk lean and fat mass during gonadotropin-releasing hormone agonist therapy. Maturitas 2002, 42, 31–35. [Google Scholar] [CrossRef]
- Poehlman, E.T.; Toth, M.J.; Gardner, A.W. Changes in energy balance and body composition at menopause: A controlled longitudinal study. Ann. Intern. Med. 1995, 123, 673–675. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Lee, D.-C. Muscle strength and quality are associated with severity of menopausal symptoms in peri- and post-menopausal women. Maturitas 2013, 76, 88–94. [Google Scholar] [CrossRef]
- Lee, E.S.; Park, H.M. Prevalence of Sarcopenia in Healthy Korean Elderly Women. J. Bone Metab. 2015, 22, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Medeiros Denis, M.; Wildman, R.E.C. Advanced Human Nutrition, 4th ed.; Sheehan, S., Ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2019. [Google Scholar]
- Tur, J.A.; Bibiloni, M.D.M. Anthropometry, Body Composition and Resting Energy Expenditure in Human. Nutrients 2019, 11, 1891. [Google Scholar] [CrossRef] [Green Version]
- Denzer, C.M.; Young, J.C. The effect of resistance exercise on the thermic effect of food. Int. J. Sport Nutr. Exerc. Metab. 2003, 13, 396–402. [Google Scholar] [CrossRef]
- Calcagno, M.; Kahleova, H.; Alwarith, J.; Burgess, N.N.; Flores, R.A.; Busta, M.L.; Barnard, N.D. The Thermic Effect of Food: A Review. J. Am. Coll. Nutr. 2019, 38, 547–551. [Google Scholar] [CrossRef]
- Shaw, E.; Leung, G.K.W.; Jong, J.; Coates, A.M.; Davis, R.; Blair, M.; Huggins, C.E.; Dorrian, J.; Banks, S.; Kellow, N.J.; et al. The Impact of Time of Day on Energy Expenditure: Implications for Long-Term Energy Balance. Nutrients 2019, 11, 2383. [Google Scholar] [CrossRef] [Green Version]
- Welle, S.; Nair, K.S. Relationship of resting metabolic rate to body composition and protein turnover. Am. J. Physiol. 1990, 258, E990–E998. [Google Scholar] [CrossRef]
- Sareen, S.; Gropper, J.L.S.; Groff, J.L. Advanced Nutrition and Human Metabolism, 4th ed.; Howe, E., Ed.; Wsdsworth, Inc.: Belmont, CA, USA, 2005; pp. 159–161. [Google Scholar]
- Thivel, D.; Tremblay, A.; Genin, P.M.; Panahi, S.; Rivière, D.; Duclos, M. Physical Activity, Inactivity, and Sedentary Behaviors: Definitions and Implications in Occupational Health. Front. Public Health 2018, 6, 288. [Google Scholar] [CrossRef]
- Colpani, V.; Oppermann, K.; Spritzer, P.M. Association between habitual physical activity and lower cardiovascular risk in premenopausal, perimenopausal, and postmenopausal women: A population-based study. Menopause 2013, 20, 525–531. [Google Scholar] [CrossRef]
- Schubert, C.M.; Rogers, N.L.; Remsberg, K.E.; Sun, S.S.; Chumlea, W.C.; Demerath, E.W.; Czerwinski, S.A.; Towne, B.; Siervogel, R.M. Lipids, lipoproteins, lifestyle, adiposity and fat-free mass during middle age: The Fels Longitudinal Study. Int. J. Obes. 2006, 30, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.N.; Kauffman, T.G.; Cooney, P.T.; Ramseur, K.R.; Brown, L.M. Comparison of DEXA and QMR for assessing fat and lean body mass in adult rats. Physiol. Behav. 2011, 103, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ying, Z.; Bosy-Westphal, A.; Zhang, J.; Heller, M.; Later, W.; Heymsfield, S.B.; Müller, M.J. Evaluation of specific metabolic rates of major organs and tissues: Comparison between men and women. Am. J. Hum. Biol. 2011, 23, 333–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polotsky, H.N.; Polotsky, A.J. Metabolic implications of menopause. Semin. Reprod. Med. 2010, 28, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Lazzer, S.; Bedogni, G.; Lafortuna, C.L.; Marazzi, N.; Busti, C.; Galli, R.; De Col, A.; Agosti, F.; Sartorio, A. Relationship between basal metabolic rate, gender, age, and body composition in 8,780 white obese subjects. Obesity 2010, 18, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Sabounchi, N.S.; Rahmandad, H.; Ammerman, A. Best-fitting prediction equations for basal metabolic rate: Informing obesity interventions in diverse populations. Int. J. Obes. 2013, 37, 1364–1370. [Google Scholar] [CrossRef] [Green Version]
- Kalyani, R.R.; Corriere, M.; Ferrucci, L. Age-related and disease-related muscle loss: The effect of diabetes, obesity, and other diseases. Lancet Diabetes Endocrinol. 2014, 2, 819–829. [Google Scholar] [CrossRef] [Green Version]
- Wildman, R.E.; Medeiros, D.M. Advanced Human Nutrition, 4th ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2019; p. 212. [Google Scholar]
- Gavin, K.M.; Sullivan, T.M.; Kohrt, W.M.; Majka, S.M.; Klemm, D.J. Ovarian Hormones Regulate the Production of Adipocytes From Bone Marrow-Derived Cells. Front. Endocrinol. 2018, 9, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.J.; Kew, K.A.; Ryan, T.E.; Pennington, E.R.; Lin, C.T.; Buddo, K.A.; Fix, A.M.; Smith, C.A.; Gilliam, L.A.; Karvinen, S.; et al. 17β-Estradiol Directly Lowers Mitochondrial Membrane Microviscosity and Improves Bioenergetic Function in Skeletal Muscle. Cell Metab. 2018, 27, 167–179.e167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [Green Version]
- Gavin, K.M.; Kohrt, W.M.; Klemm, D.J.; Melanson, E.L. Modulation of Energy Expenditure by Estrogens and Exercise in Women. Exerc. Sport Sci. Rev. 2018, 46, 232–239. [Google Scholar] [CrossRef]
- Dube, M.C.; Lemieux, S.; Piche, M.E.; Corneau, L.; Bergeron, J.; Riou, M.E.; Weisnagel, S.J. The contribution of visceral adiposity and mid-thigh fat-rich muscle to the metabolic profile in postmenopausal women. Obesity 2011, 19, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Leon, A.S. Attenuation of Adverse Effects of Aging on Skeletal Muscle by Regular Exercise and Nutritional Support. Am. J. Lifestyle Med. 2017, 11, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.C.; Laakkonen, E.K.; Lowe, D.A. Aging of the musculoskeletal system: How the loss of estrogen impacts muscle strength. Bone 2019, 123, 137–144. [Google Scholar] [CrossRef]
- Moreira, M.A.; Zunzunegui, M.V.; Vafaei, A.; da Camara, S.M.; Oliveira, T.S.; Maciel, A.C. Sarcopenic obesity and physical performance in middle aged women: A cross-sectional study in Northeast Brazil. BMC Public Health 2016, 16, 43. [Google Scholar] [CrossRef] [Green Version]
- Agostini, D.; Zeppa Donati, S.; Lucertini, F.; Annibalini, G.; Gervasi, M.; Ferri Marini, C.; Piccoli, G.; Stocchi, V.; Barbieri, E.; Sestili, P. Muscle and Bone Health in Postmenopausal Women: Role of Protein and Vitamin D Supplementation Combined with Exercise Training. Nutrients 2018, 10, 1103. [Google Scholar] [CrossRef] [Green Version]
- Kirk, B.; Al Saedi, A.; Duque, G. Osteosarcopenia: A case of geroscience. Aging Med. 2019, 2, 147–156. [Google Scholar] [CrossRef]
- Hamad, B.; Basaran, S.; Coskun Benlidayi, I. Osteosarcopenia among postmenopausal women and handgrip strength as a practical method for predicting the risk. Aging Clin. Exp. Res. 2020, 32, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.I.; Kim, H.; Ha, Y.C.; Kwon, H.B.; Koo, K.H. Osteosarcopenia in Patients with Hip Fracture Is Related with High Mortality. J. Korean Med Sci. 2018, 33, e27. [Google Scholar] [CrossRef]
- Greco, E.A.; Pietschmann, P.; Migliaccio, S. Osteoporosis and Sarcopenia Increase Frailty Syndrome in the Elderly. Front. Endocrinol. 2019, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic, V.; van der Mooren, M.J.; Stehouwer, C.D.; Netelenbos, J.C.; Kenemans, P. Postmenopausal hormone replacement, risk estimators for coronary artery disease and cardiovascular protection. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 1999, 13, 130–144. [Google Scholar] [CrossRef]
- Mumusoglu, S.; Yildiz, B.O. Metabolic Syndrome During Menopause. Curr. Vasc. Pharmacol. 2019, 17, 595–603. [Google Scholar] [CrossRef]
- Al-Safi, Z.A.; Polotsky, A.J. Obesity and menopause. Best Pract. Res. Clin. Obstet. Gynaecol. 2015, 29, 548–553. [Google Scholar] [CrossRef]
- Razmjou, S.; Abdulnour, J.; Bastard, J.P.; Fellahi, S.; Doucet, E.; Brochu, M.; Lavoie, J.M.; Rabasa-Lhoret, R.; Prud’homme, D. Body composition, cardiometabolic risk factors, physical activity, and inflammatory markers in premenopausal women after a 10-year follow-up: A MONET study. Menopause 2018, 25, 89–97. [Google Scholar] [CrossRef]
- Lovejoy, J.C.; Bray, G.A.; Bourgeois, M.O.; Macchiavelli, R.; Rood, J.C.; Greeson, C.; Partington, C. Exogenous androgens influence body composition and regional body fat distribution in obese postmenopausal women—A clinical research center study. J. Clin. Endocrinol. Metab. 1996, 81, 2198–2203. [Google Scholar] [CrossRef]
- Perry, A.; Wang, X.; Goldberg, R.; Ross, R.; Jackson, L. Androgenic sex steroids contribute to metabolic risk beyond intra-abdominal fat in overweight/obese black and white women. Obesity 2013, 21, 1618–1624. [Google Scholar] [CrossRef] [PubMed]
- Hammes, S.R.; Levin, E.R. Impact of estrogens in males and androgens in females. J. Clin. Investig. 2019, 129, 1818–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, A.P.; de Souza Santos, R.; Palmer, B.F.; Clegg, D.J. Determinants of body fat distribution in humans may provide insight about obesity-related health risks. J. Lipid Res. 2019, 60, 1710–1719. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.M. Lipid metabolism in women. Proc. Nutr. Soc. 2004, 63, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Sutton-Tyrrell, K.; Wildman, R.P.; Matthews, K.A.; Chae, C.; Lasley, B.L.; Brockwell, S.; Pasternak, R.C.; Lloyd-Jones, D.; Sowers, M.F.; Torrens, J.I.; et al. Sex-hormone-binding globulin and the free androgen index are related to cardiovascular risk factors in multiethnic premenopausal and perimenopausal women enrolled in the Study of Women Across the Nation (SWAN). Circulation 2005, 111, 1242–1249. [Google Scholar] [CrossRef] [Green Version]
- Torrens, J.I.; Sutton-Tyrrell, K.; Zhao, X.; Matthews, K.; Brockwell, S.; Sowers, M.; Santoro, N. Relative androgen excess during the menopausal transition predicts incident metabolic syndrome in midlife women: Study of Women’s Health Across the Nation. Menopause 2009, 16, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.H. Androgens, estrogens, and cardiovascular disease: Considerations for women with polycystic ovary syndrome. Fertil. Steril. 2019, 112, 478–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douchi, T.; Yoshimitsu, N.; Nagata, Y. Relationships among serum testosterone levels, body fat and muscle mass distribution in women with polycystic ovary syndrome. Endocr. J. 2001, 48, 685–689. [Google Scholar] [CrossRef] [Green Version]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of lipolysis in adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, A.; Mottillo, E.P. Adipocyte lipolysis: From molecular mechanisms of regulation to disease and therapeutics. Biochem. J. 2020, 477, 985–1008. [Google Scholar] [CrossRef]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [Green Version]
- Luong, Q.; Huang, J.; Lee, K.Y. Deciphering White Adipose Tissue Heterogeneity. Biology 2019, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Berman, D.M.; Nicklas, B.J.; Rogus, E.M.; Dennis, K.E.; Goldberg, A.P. Regional differences in adrenoceptor binding and fat cell lipolysis in obese, postmenopausal women. Metabolism 1998, 47, 467–473. [Google Scholar] [CrossRef]
- Stefanska, A.; Bergmann, K.; Sypniewska, G. Metabolic Syndrome and Menopause: Pathophysiology, Clinical and Diagnostic Significance. Adv. Clin. Chem. 2015, 72, 1–75. [Google Scholar] [CrossRef]
- Yamatani, H.; Takahashi, K.; Yoshida, T.; Soga, T.; Kurachi, H. Differences in the fatty acid metabolism of visceral adipose tissue in postmenopausal women. Menopause 2014, 21, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Goettems-Fiorin, P.B.; Costa-Beber, L.C.; Dos Santos, J.B.; Friske, P.T.; Sulzbacher, L.M.; Frizzo, M.N.; Ludwig, M.S.; Rhoden, C.R.; Heck, T.G. Ovariectomy predisposes female rats to fine particulate matter exposure’s effects by altering metabolic, oxidative, pro-inflammatory, and heat-shock protein levels. Environ. Sci. Pollut. Res. Int. 2019, 26, 20581–20594. [Google Scholar] [CrossRef]
- de Souza, C.F.; Stopa, L.R.S.; Santos, G.F.; Takasumi, L.C.N.; Martins, A.B.; Garnica-Siqueira, M.C.; Ferreira, R.N.; de Andrade, F.G.; Leite, C.M.; Zaia, D.A.M.; et al. Estradiol protects against ovariectomy-induced susceptibility to the anabolic effects of glucocorticoids in rats. Life Sci. 2019, 218, 185–196. [Google Scholar] [CrossRef]
- Moise, A.R.; Kuksa, V.; Imanishi, Y.; Palczewski, K. Identification of all-trans-retinol:all-trans-13,14-dihydroretinol saturase. J. Biol. Chem. 2004, 279, 50230–50242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soucy, G.; Boivin, G.; Labrie, F.; Rivest, S. Estradiol is required for a proper immune response to bacterial and viral pathogens in the female brain. J. Immunol. 2005, 174, 6391–6398. [Google Scholar] [CrossRef] [Green Version]
- Costa-Beber, L.C.; Goettems-Fiorin, P.B.; Dos Santos, J.B.; Friske, P.T.; Frizzo, M.N.; Heck, T.G.; Hirsch, G.E.; Ludwig, M.S. Ovariectomy enhances female rats’ susceptibility to metabolic, oxidative, and heat shock response effects induced by a high-fat diet and fine particulate matter. Exp. Gerontol. 2021, 145, 111215. [Google Scholar] [CrossRef]
- Kastaniotis, A.J.; Autio, K.J.; Kerätär, J.M.; Monteuuis, G.; Mäkelä, A.M.; Nair, R.R.; Pietikäinen, L.P.; Shvetsova, A.; Chen, Z.; Hiltunen, J.K. Mitochondrial fatty acid synthesis, fatty acids and mitochondrial physiology. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2017, 1862, 39–48. [Google Scholar] [CrossRef]
- Kushwaha, P.; Wolfgang, M.J.; Riddle, R.C. Fatty acid metabolism by the osteoblast. Bone 2018, 115, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; Carneiro-Freire, N.; Seco-Filgueira, M.; Fernández-Fernández, C.; Mouriño-Bayolo, D. Mitochondrial β-oxidation of saturated fatty acids in humans. Mitochondrion 2019, 46, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Karwi, Q.G.; Uddin, G.M.; Ho, K.L.; Lopaschuk, G.D. Loss of Metabolic Flexibility in the Failing Heart. Front. Cardiovasc. Med. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holloway, G.P.; Bonen, A.; Spriet, L.L. Regulation of skeletal muscle mitochondrial fatty acid metabolism in lean and obese individuals. Am. J. Clin. Nutr. 2009, 89, 455s–462s. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Men | Women | |
|---|---|---|
| Age, years | 20–24 | 20–24 |
| Height, in | 68.5 | 64.5 |
| Weight, lb | 154 | 125 |
| Total fat, lb (% body weight) | 23.1 (15.0%) | 33.8 (27.0%) |
| Storage fat, lb (% body weight) | 18.5 (12.0%) | 18.8 (15.0%) |
| Essential fat, lb (% body weight) | 4.6 (3.0%) | 15.0 (12.0%) |
| Muscle, lb (% body weight) | 69 (44.8%) | 45 (36.0%) |
| Bone, lb (% body weight) | 21 (14.9%) | 15 (12.0%) |
| Remainder, lb (% body weight) | 38.9 (25.3%) | 31.2 (25.3%) |
| Average body density | 1.070 g/mL | 1.040 g/mL |
| REE | MEN | WOMEN | |
|---|---|---|---|
| kcal/kg/Day | % Total REE | % Total REE | |
| Liver | 200 | 17 | 18 |
| Brain | 240 | 19 | 21 |
| Heart | 440 | 9 | 8 |
| Kidneys | 440 | 8 | 8 |
| Skeletal muscle a | 13 | 24 | 20 |
| Adipose tissue | 4.5 | 4 | 7 |
| Other b | 12 | 19 | 18 |
| Total | 100 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, S.-H.; Jung, Y. Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women. Nutrients 2021, 13, 4556. https://doi.org/10.3390/nu13124556
Ko S-H, Jung Y. Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women. Nutrients. 2021; 13(12):4556. https://doi.org/10.3390/nu13124556
Chicago/Turabian StyleKo, Seong-Hee, and YunJae Jung. 2021. "Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women" Nutrients 13, no. 12: 4556. https://doi.org/10.3390/nu13124556
APA StyleKo, S.-H., & Jung, Y. (2021). Energy Metabolism Changes and Dysregulated Lipid Metabolism in Postmenopausal Women. Nutrients, 13(12), 4556. https://doi.org/10.3390/nu13124556