
Effect and Tolerability of a Nutritional Supplement Based on a Synergistic Combination of β-Glucans and Selenium- and Zinc-Enriched Saccharomyces cerevisiae (ABB C1®) in Volunteers Receiving the Influenza or the COVID-19 Vaccine: A Randomized, Double-Blind, Placebo-Controlled Study

Abstract
:1. Introduction
2. Materials and Methods
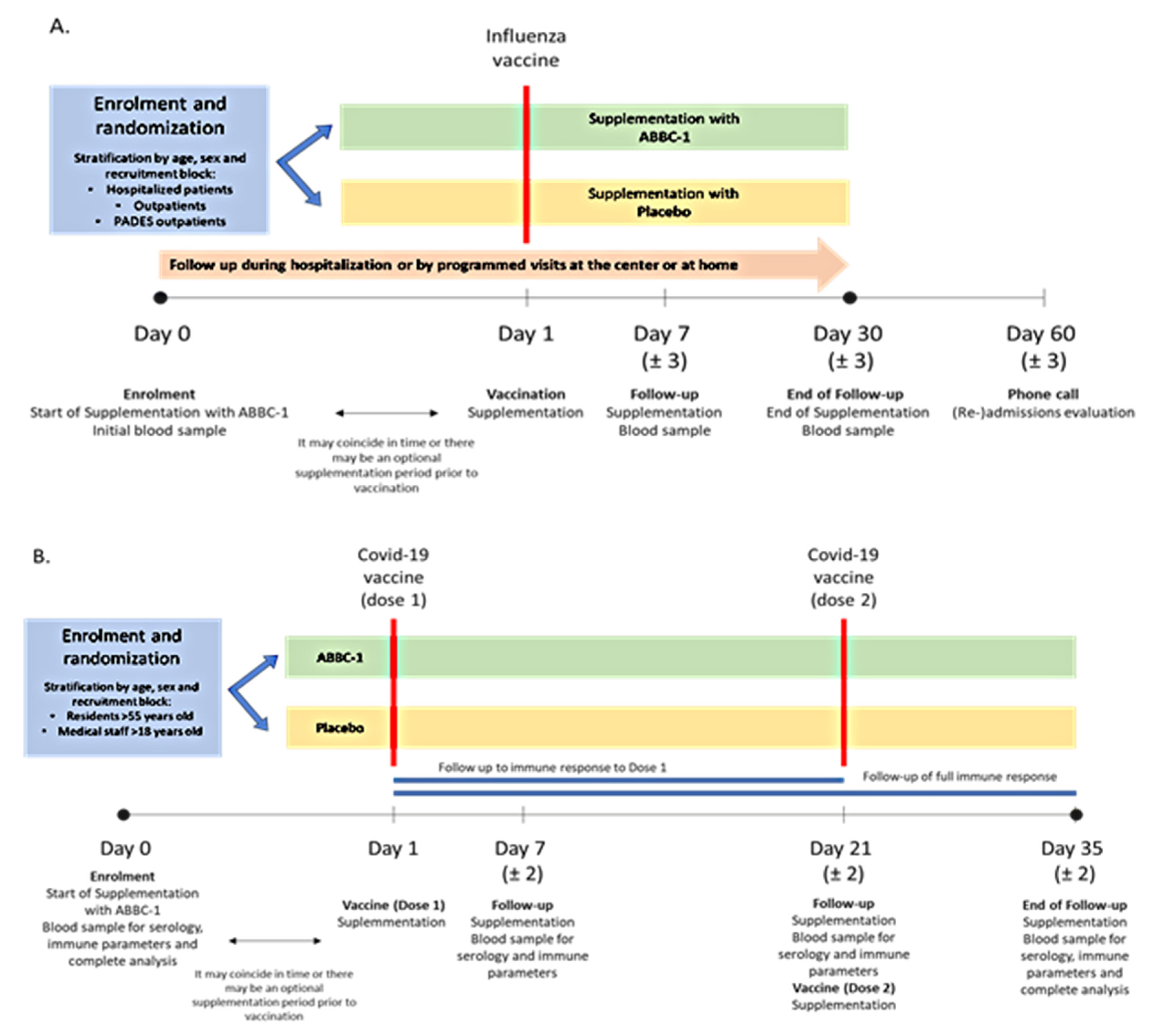
2.1. Study Design
2.2. Participants’ Selection and Randomization
2.3. Intervention and Study Procedures
2.4. Laboratory Analyses
2.5. Statistical Analysis
3. Results
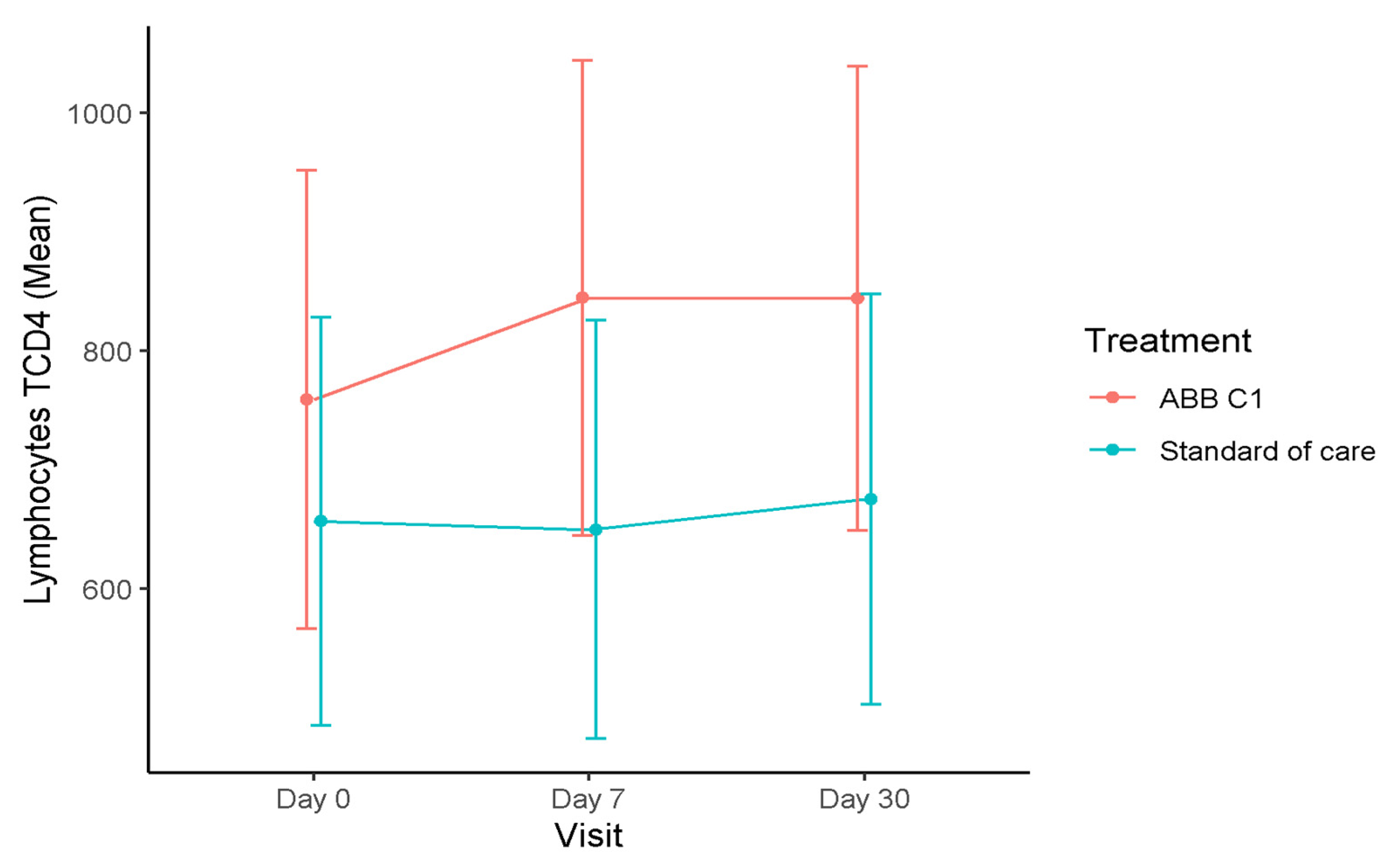
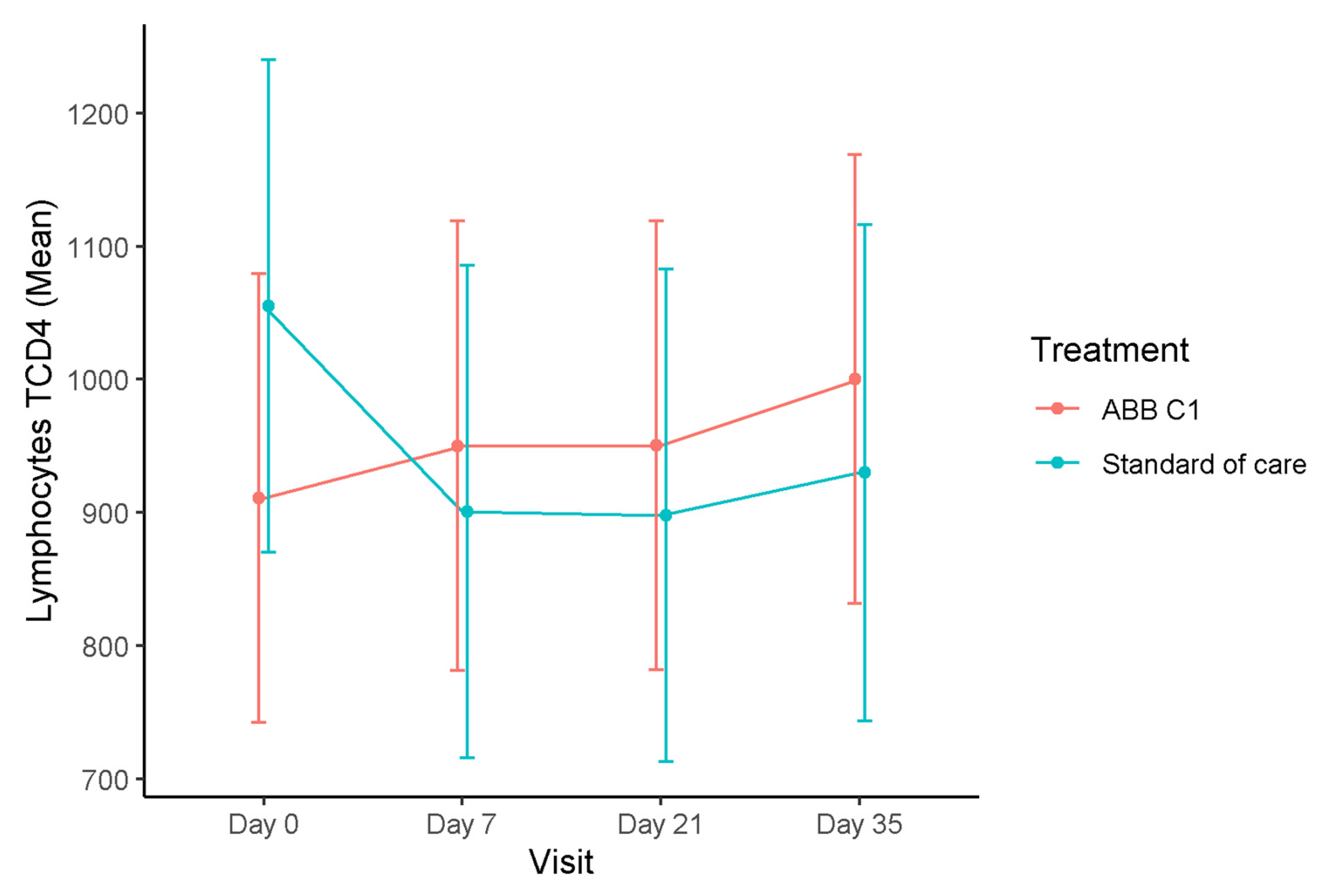
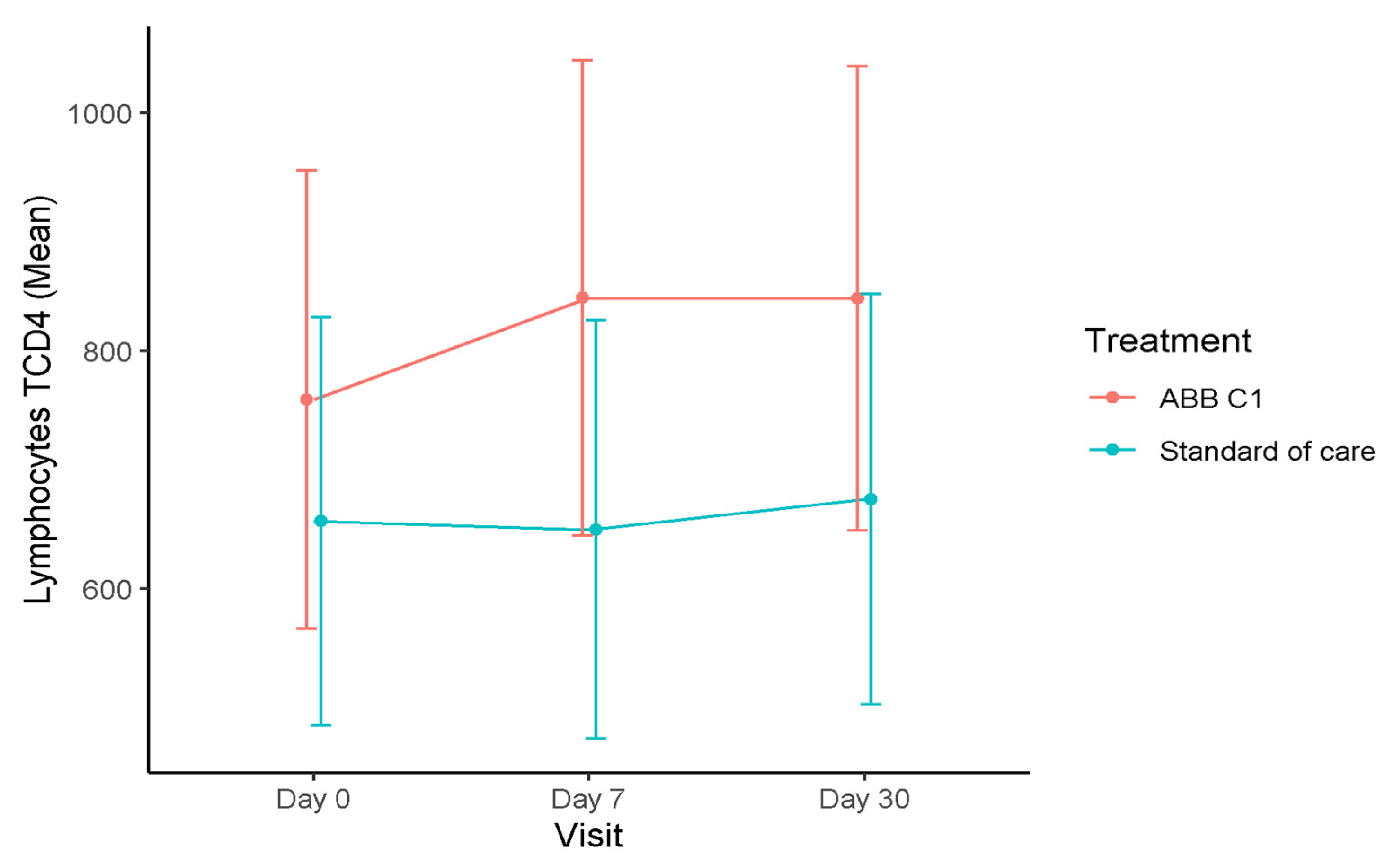
3.1. T Lymphocytes
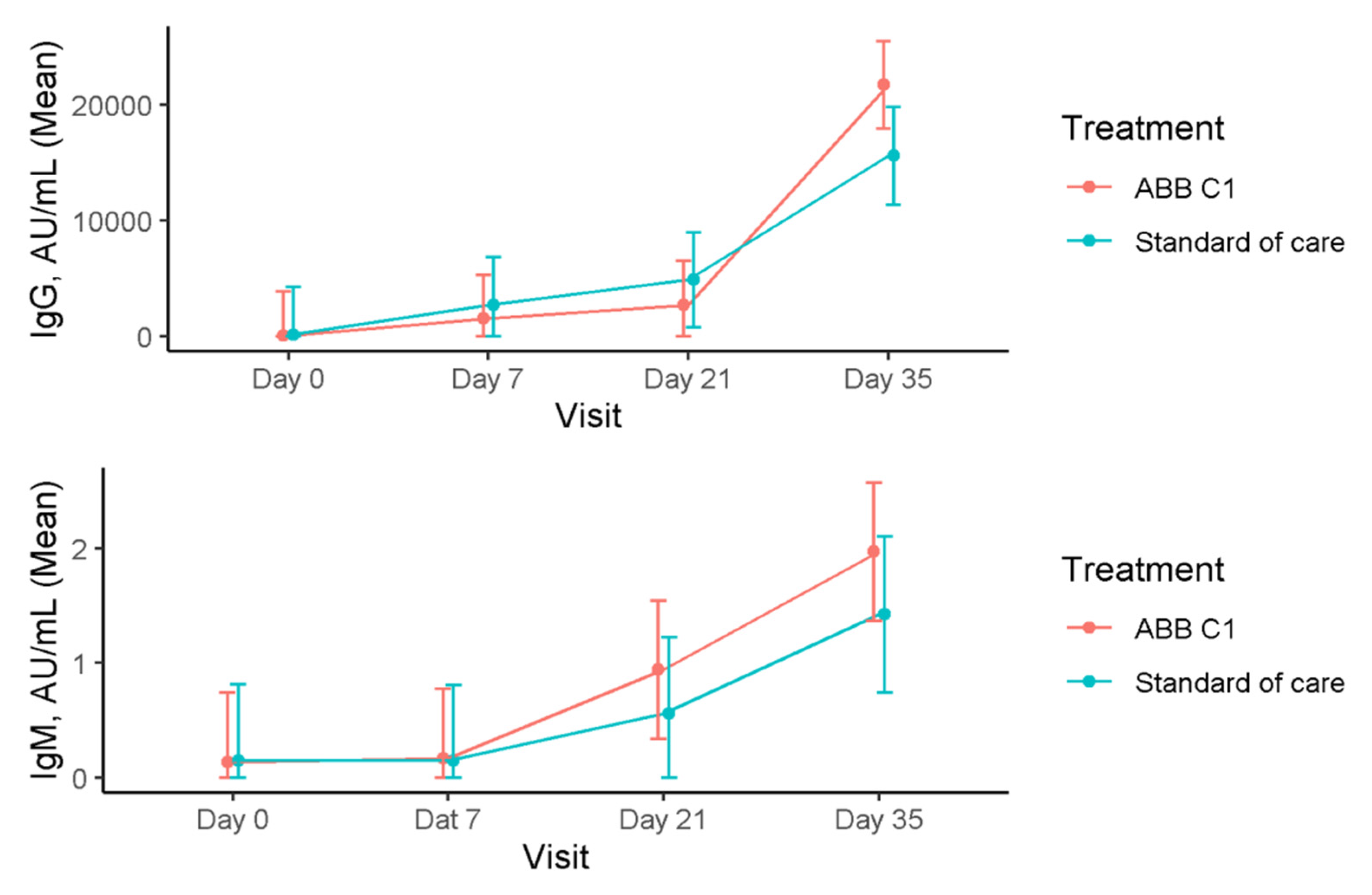
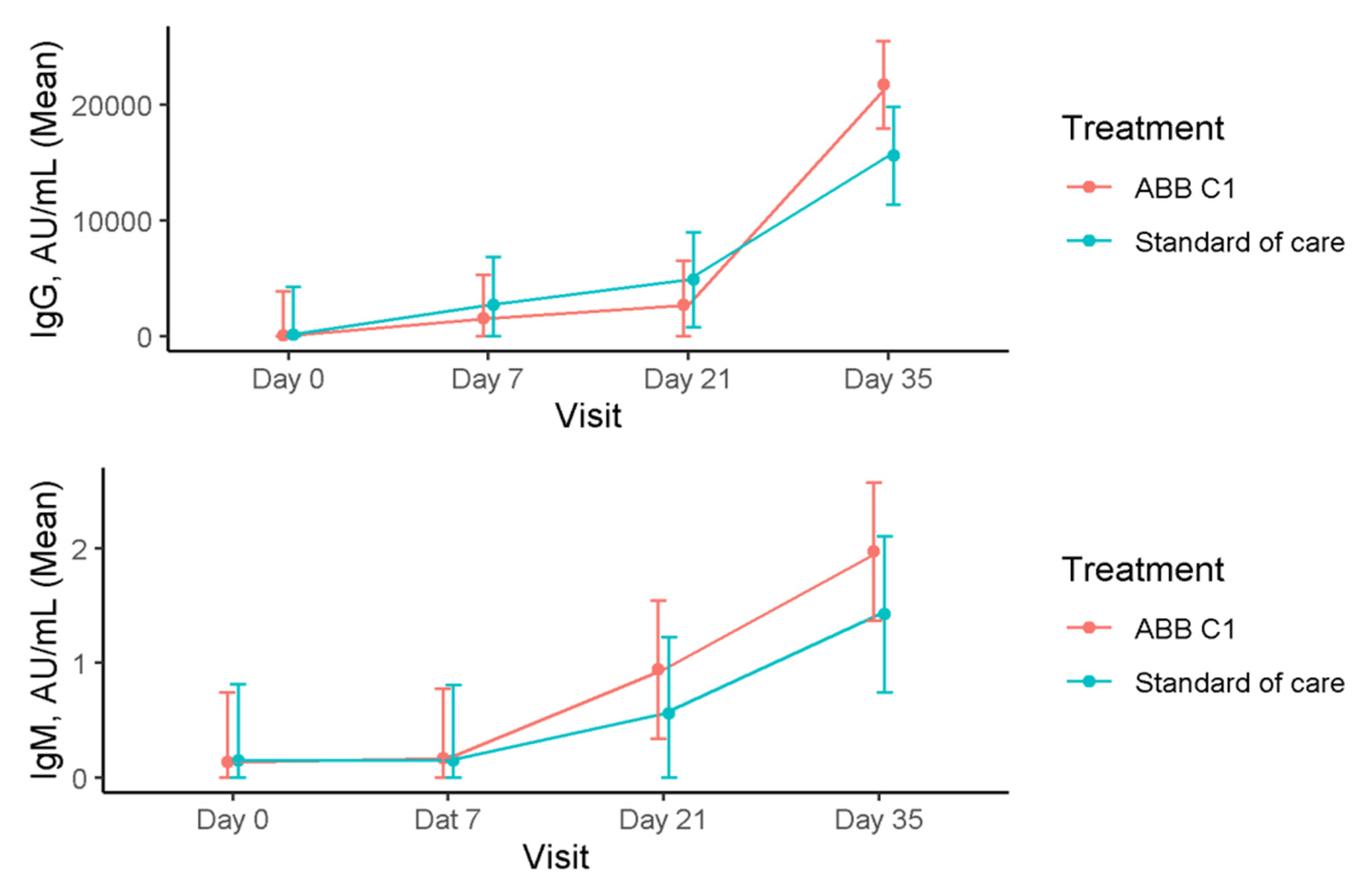
3.2. Immunoglobulins
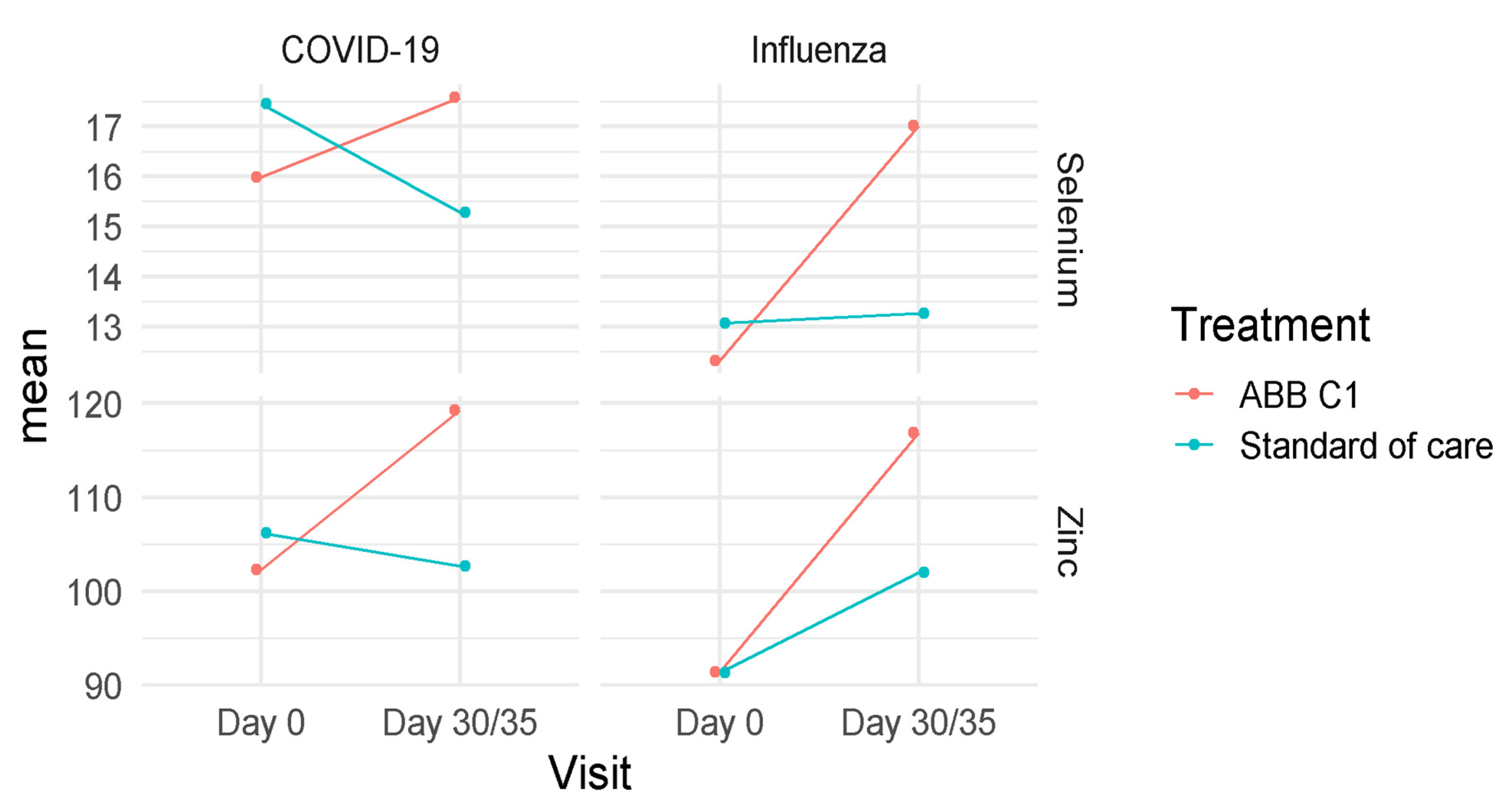
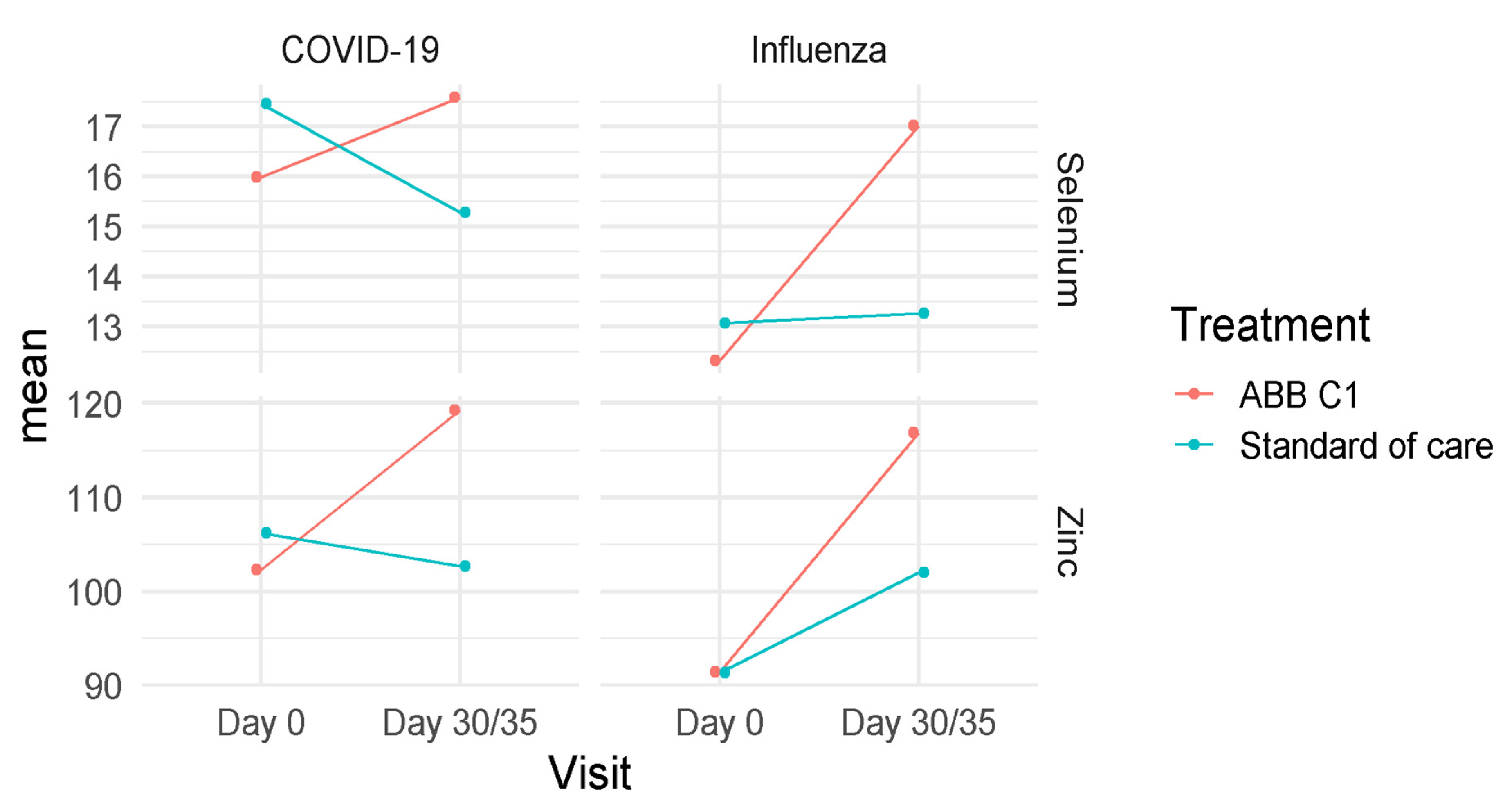
3.3. Selenium and Zinc
3.4. Tolerability and Safety
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singhal, S.; Kumar, P.; Singh, S.; Saha, S.; Dey, A.B. Clinical features and outcomes of COVID-19 in older adults: A systematic review and meta-analysis. BMC Geriatr. 2021, 21, 321. [Google Scholar] [CrossRef]
- Yang, J.; Zheng, Y.; Gou, X.; Pu, K.; Chen, Z.; Guo, Q.; Ji, R.; Wang, H.; Wang, Y.; Zhou, Y. Prevalence of comorbidities and its effects in patients infected with SARS-CoV-2: A systematic review and meta-analysis. Int. J. Infect. Dis. 2020, 94, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Hazeldine, J.; Lord, J.M. Immunesenescence: A predisposing risk factor for the development of COVID-19? Front. Immunol. 2020, 11, 573662. [Google Scholar] [CrossRef] [PubMed]
- Reina, J. La vacunación de la gripe en el tiempo del SARS-CoV-2. Med. Clin. 2020, 156, 17–19. [Google Scholar] [CrossRef]
- Amato, M.; Werba, J.P.; Frigerio, B.; Coggi, D.; Sansaro, D.; Ravani, A.; Ferrante, P.; Veglia, F.; Tremoli, E.; Baldassarre, D. Relationship between influenza vaccination coverage rate and COVID-19 outbreak: An Italian ecological study. Vaccines 2020, 8, 535. [Google Scholar] [CrossRef]
- Candelli, M.; Pignataro, G.; Torelli, E.; Gullì, A.; Nista, E.C.; Petrucci, M.; Saviano, A.; Marchesini, D.; Covino, M.; Ojetti, V.; et al. Effect of influenza vaccine on COVID-19 mortality: A retrospective study. Intern. Emerg. Med. 2021, 20, 1849–1855. [Google Scholar] [CrossRef]
- Fink, G.; Orlova-Fink, N.; Schindler, T.; Grisi, S.; Ferrer, A.P.S.; Daubenberger, C.; Brentani, A. Inactivated trivalent influenza vaccination is associated with lower mortality among patients with COVID-19 in Brazil. BMJ Evid. Based Med. 2021, 26, 192–193. [Google Scholar] [CrossRef] [PubMed]
- Arts, R.J.W.; Moorlag, S.J.C.F.M.; Novakovic, B.; Li, Y.; Wang, S.Y.; Oosting, M.; Kumar, V.; Xavier, R.J.; Wijmenga, C.; Joosten, L.A.B.; et al. BCG Vaccination protects against experimental viral infection in humans through the induction of cytokines associated with trained immunity. Cell Host Microbe 2018, 23, 89–100.e5. [Google Scholar] [CrossRef] [Green Version]
- De Marco Castro, E.; Calder, P.C.; Roche, H.M. β-1,3/1,6-Glucans and immunity: State of the art and future directions. Mol. Nutr. Food Res. 2021, 65, e1901071. [Google Scholar] [CrossRef]
- Netea, M.G.; Domínguez-Andrés, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; van der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Giamarellos-Bourboulis, E.J.; Domínguez-Andrés, J.; Curtis, N.; van Crevel, R.; van de Veerdonk, F.L.; Bonten, M. Trained immunity: A tool for reducing susceptibility to and the severity of SARS-CoV-2 infection. Cell 2020, 181, 969–977. [Google Scholar] [CrossRef]
- Geller, A.; Yan, J. Could the induction of trained immunity by β-glucan serve as a defense against COVID-19? Front. Immunol. 2020, 11, 1782. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, selenoproteins, and immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [Green Version]
- Tinggi, U. Selenium: Its role as antioxidant in human health. Environ. Health Prev. Med. 2008, 13, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohé, R.; Flohé, L. Selenium and redox signaling. Arch. Biochem. Biophys. 2017, 617, 48–59. [Google Scholar] [CrossRef]
- Handy, D.E.; Joseph, J.; Loscalzo, J. Selenium, a micronutrient that modulates cardiovascular health via redox enzymology. Nutrients 2021, 13, 3238. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.; Heller, R.A.; Sun, Q.; Seelig, J.; Cherkezov, A.; Seibert, L.; Hackler, J.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Selenium deficiency is associated with mortality risk from COVID-19. Nutrients 2020, 12, 2098. [Google Scholar] [CrossRef]
- Zhang, J.; Taylor, E.W.; Bennett, K.; Saad, R.; Rayman, M.P. Association between regional selenium status and reported outcome of COVID-19 cases in China. Am. J. Clin. Nutr. 2020, 111, 1297–1299. [Google Scholar] [CrossRef]
- Guillin, O.M.; Vindry, C.; Ohlmann, T.; Chavatte, L. Selenium, selenoproteins and viral infection. Nutrients 2019, 11, 2101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, S.A.; Obeid, S.; Ahlenstiel, C.; Ahlenstiel, G. The role of zinc in antiviral immunity. Adv. Nutr. 2019, 10, 696–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel-González, M.; Talló-Parra, M.; Herrera-Fernández, V.; Pérez-Vilaró, G.; Chillón, M.; Nogués, X.; Gómez-Zorrilla, S.; López-Montesinos, I.; Arnau-Barrés, I.; Sorli-Redó, M.L.; et al. Low zinc levels at admission associates with poor clinical outcomes in SARS-CoV-2 infection. Nutrients 2021, 13, 562. [Google Scholar] [CrossRef] [PubMed]
- Goldson, A.J.; Fairweather-Tait, S.J.; Armah, C.N.; Bao, Y.; Broadley, M.R.; Dainty, J.R.; Furniss, C.; Hart, D.J.; Teucher, B.; Hurst, R. Effects of selenium supplementation on selenoprotein gene expression and response to influenza vaccine challenge: A randomised controlled trial. PLoS ONE 2011, 6, e14771. [Google Scholar] [CrossRef]
- Girodon, F.; Galan, P.; Monget, A.L.; Boutron-Ruault, M.C.; Brunet-Lecomte, P.; Preziosi, P.; Arnaud, J.; Manuguerra, J.C.; Herchberg, S. Impact of trace elements and vitamin supplementation on immunity and infections in institutionalized elderly patients: A randomized controlled trial. MIN. VIT. AOX. geriatric network. Arch. Intern. Med. 1999, 159, 748–754. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.; Tinkov, A.; Strand, T.A.; Alehagen, U.; Skalny, A.; Aaseth, J. Early nutritional interventions with zinc, selenium and vitamin d for raising anti-viral resistance against progressive COVID-19. Nutrients 2020, 12, 2358. [Google Scholar] [CrossRef]
- Rayman, M.P.; Calder, P.C. Optimising COVID-19 vaccine efficacy by ensuring nutritional adequacy. Br. J. Nutr. 2021, 126, 1919–1920. [Google Scholar] [CrossRef] [PubMed]
- Peretz, A.; Nève, J.; Desmedt, J.; Duchateau, J.; Dramaix, M.; Famaey, J.P. Lymphocyte response is enhanced by supplementation of elderly subjects with selenium-enriched yeast. Am. J. Clin. Nutr. 1991, 53, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Tompkins, T.A.; Renard, N.E.; Kiuchi, A. Clinical evaluation of the bioavailability of zinc-enriched yeast and zinc gluconate in healthy volunteers. Biol. Trace Elem. Res. 2007, 120, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.L.; Zamith-Miranda, D.; Martins, F.S.; Bozza, F.A.; Nimrichter, L.; Montero-Lomeli, M.; Marques, E.T., Jr.; Douradinha, B. Probiotic Saccharomyces cerevisiae strains as biotherapeutic tools: Is there room for improvement? Appl. Microbiol. Biotechnol. 2015, 99, 6563–6570. [Google Scholar] [CrossRef] [PubMed]
- Trottein, F.; Sokol, H. Potential causes and consequences of gastrointestinal disorders during a SARS-CoV-2 Infection. Cell Rep. 2020, 32, 107915. [Google Scholar] [CrossRef]
- Zhou, Y.; Shi, X.; Fu, W.; Xiang, F.; He, X.; Yang, B.; Wang, X.; Ma, W.L. Gut microbiota dysbiosis correlates with abnormal immune response in moderate COVID-19 patients with fever. J. Inflamm. Res. 2021, 14, 2619–2631. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, J.; Zhang, D.; Ma, W.L.; Wang, X. Linking the gut microbiota to persistent symptoms in survivors of COVID-19 after discharge. J. Microbiol. 2021, 59, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Vetvickova, J. Addition of selenium improves immunomodulative effects of glucan. N. Am. J. Med. Sci. 2016, 8, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Vetvicka, V.; Pinatto-Botelho, M.F.; Dos Santos, A.A.; De Oliveira, C.A. Evaluation of a special combination of glucan with organic selenium derivative in different murine tumor models. Anticancer Res. 2014, 34, 6939–6944. [Google Scholar]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.; Jacobs, C.; Xavier, R.J.; van der Meer, J.W.; van Crevel, R.; Netea, M.G. BCG-induced trained immunity in NK cells: Role for non-specific protection to infection. Clin. Immunol. 2014, 155, 213–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walk, J.; de Bree, L.C.J.; Graumans, W.; Stoter, R.; van Gemert, G.J.; van de Vegte-Bolmer, M.; Teelen, K.; Hermsen, C.C.; Arts, R.J.W.; Behet, M.C.; et al. Outcomes of controlled human malaria infection after BCG vaccination. Nat. Commun. 2019, 10, 874. [Google Scholar] [CrossRef] [Green Version]
- Taghioff, S.M.; Slavin, B.R.; Holton, T.; Singh, D. Examining the potential benefits of the influenza vaccine against SARS-CoV-2: A retrospective cohort analysis of 74,754 patients. PLoS ONE 2021, 16, e0255541. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Murasko, D.M.; Bernstein, E.D.; Gardner, E.M.; Gross, P.; Munk, G.; Dran, S.; Abrutyn, E. Role of humoral and cell-mediated immunity in protection from influenza disease after immunization of healthy elderly. Exp. Gerontol. 2002, 37, 427–439. [Google Scholar] [CrossRef]
- Burlington, D.B.; Clements, M.L.; Meiklejohn, G.; Phelan, M.; Murphy, B.R. Hemagglutinin-specific antibody responses in immunoglobulin G, A, and M isotypes as measured by enzyme-linked immunosorbent assay after primary or secondary infection of humans with influenza a virus. Infect. Immun. 1983, 41, 540–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Science, M.; Johnstone, J.; Roth, D.E.; Guyatt, G.; Loeb, M. Zinc for the treatment of the common cold: A systematic review and meta-analysis of randomized controlled trials. Can. Med Assoc. J. 2012, 184, E551–E561. [Google Scholar] [CrossRef] [Green Version]
- Paget, J.; Caini, S.; Cowling, B.; Esposito, S.; Falsey, A.R.; Gentile, A.; Kyncl, J.; MacIntyre, C.; Pitman, R.; Lina, B. The impact of influenza vaccination on the COVID-19 pandemic? Evidence and lessons for public health policies. Vaccine 2020, 38, 6485–6486. [Google Scholar] [CrossRef]
- Woldemeskel, B.A.; Karaba, A.H.; Garliss, C.C.; Beck, E.J.; Wang, K.H.; Laeyendecker, O.; Cox, A.L.; Blankson, J.N. The BNT162b2 mRNA vaccine elicits robust humoral and cellular immune responses in people living with HIV. Clin. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Muntjewerff, E.M.; Meesters, L.D.; van den Bogaart, G. Antigen cross-presentation by macrophages. Front. Immunol. 2020, 11, 1276. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, S.; Prommersberger, S.; Pfeiffer, I.A.; Schuler-Thurner, B.; Schuler, G.; Dörrie, J.; Schaft, N. Concurrent interaction of DCs with CD4(+) and CD8(+) T cells improves secondary CTL expansion: It takes three to tango. Eur. J. Immunol. 2014, 44, 3543–3559. [Google Scholar] [CrossRef]
- Hivroz, C.; Chemin, K.; Tourret, M.; Bohineust, A. Crosstalk between T lymphocytes and dendritic cells. Crit. Rev. Immunol. 2012, 32, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Chetty, R.; Gatter, K. CD3: Structure, function, and role of immunostaining in clinical practice. J. Pathol. 1994, 173, 303–307. [Google Scholar] [CrossRef] [PubMed]
- McElhaney, J.E.; Upshaw, C.M.; Hooton, J.W.; Lechelt, K.E.; Meneilly, G.S. Responses to influenza vaccination in different T-cell subsets: A comparison of healthy young and older adults. Vaccine 1998, 16, 1742–1747. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Cohort | CD4+T, Cells/µL Mean (95% CI) | CD3+T, Cells/µL Mean (95% CI) | CD8+T, Cells/µL Mean (95% CI) |
|---|---|---|---|
| COVID-19 vaccine | |||
| ABB C1® supplement | |||
| Baseline (n = 18) | 910.67 (741.90–1079.44) | 1508.67 (1278.05–1739.28) | 585.50 (464.27–706.73) |
| Day 7 (n = 18) | 950.0 (781.23–1118.77) | 1589.44 (1358.83–1820.06) | 606.50 (485.27–727.73) |
| Day 21 (n = 18) | 950.28 (781.51–1119.05) | 1598.06 (1367.44–1828.67) | 621.11 (499.88–742.34) |
| Day 35 (n = 18) | 1000.17 (831.40–1168.94) | 1699.56 (1468.94–1930.17) | 653.44 (532.21–774.68) |
| Placebo | |||
| Baseline (n = 15) | 1055.07 (869.96–1240.18) | 1587.47 (1334.52–1840.41) | 496.73 (363.76–629.71) |
| Day 7 (n = 15) | 900.60 (715.49–1085.71) | 1367.20 (1114.25–1620.15) | 435.73 (302.76–568.71) |
| Day 21 (n = 15) | 897.80 (712.69–1082.91) | 1391.67 (1138.72–1644.61) | 450.87 (317.89–583.84) |
| Day 35 (n = 14) | 929.80 (743.15–1116.46) | 1407.23 (1151.75–1662.71) | 450.10 (316.16–584.03) |
| Study Cohort | CD4+T, Cells/µL Mean (95% CI) | CD3+T, Cells/µL Mean (95% CI) | CD8+T, Cells/µL Mean (95% CI) |
|---|---|---|---|
| Influenza vaccine | |||
| ABB C1® supplement | |||
| Baseline (n = 15) | 759.40 (566.85–951.45) | 1195.20 (955.85–1434.55) | 396.0 (291.55–500.45) |
| Day 7 (n = 12) | 844.41 (644.80–1044.03) | 1278.40 (1024.89–1531.91) | 402.45 (291.22–513.69) |
| Day 30 (n = 14) | 843.87 (648.94–1038.80) | 1311.45 (1067.43–1555.46) | 414.20 (307.54–520.87) |
| Placebo | |||
| Baseline (n = 19) | 656.84 (485.55–828.13) | 1114.63 (901.71–1327.55) | 422.68 (329.77–515.60) |
| Day 7 (n = 16) | 649.93 (474.20–825.65) | 1069.96 (847.99–1291.92) | 390.68 (293.41–487.95) |
| Day 30 (n = 18) | 675.42 (502.81–848.04) | 1065.32 (849.68–1280.96) | 359.61 (265.38–453.83) |
| Study Cohort | IgG, AU/mL Mean (95% CI) | IgM, AU/mL Mean (95% CI) |
|---|---|---|
| COVID-19 vaccine | ||
| ABB C1® supplement | ||
| Baseline (n = 18) | 114.34 (−3634.77–3863.45) | 0.14 (−0.46–0.74) |
| Day 7 (n = 18) | 1560.97 (−2188.14–5310.08) | 0.17 (−0.43–0.77) |
| Day 21 (n = 18) | 2758.97 (−990.14–6508.08) | 0.94 (0.34–1.54) |
| Day 35 (n = 18) | 21,716.64 (17,967.53–25,465.75) | 1.97 (1.37–2.58) |
| Placebo | ||
| Baseline (n = 15) | 136.95 (−3975.20–4249.10) | 0.15 (−0.51–0.81) |
| Day 7 (n = 15) | 2733.02 (−1379.13–6845.16) | 0.15 (−0.51–0.81) |
| Day 21 (n = 15) | 4892.81 (780.67–9004.96) | 0.56 (−0.10–1.22) |
| Day 35 (n = 14) | 15,595.70 (11,369.80–19,821.60) | 1.42 (0.74–2.11) |
| Study Cohort | Anti-Influenza A IgG, AI Mean (95% CI) | Anti-Influenza B IgG, AI Mean (95% CI) |
|---|---|---|
| Influenza vaccine | ||
| ABB C1® supplement | ||
| Baseline (n = 15) | 2.29 (1.08–3.50) | 2.35 (0.92–3.78) |
| Day 7 (n = 12) | 2.41 (1.19–3.63) | 1.95 (0.48–3.42) |
| Day 30 (n = 14) | 2.22 (1.01–3.43) | 2.31 (0.86–3.75) |
| Placebo | ||
| Baseline (n = 19) | 2.67 (1.60–3.75) | 2.54 (1.27–3.81) |
| Day 7 (n = 16) | 2.37 (1.28–3.45) | 2.42 (1.12–3.71) |
| Day 30 (n = 18) | 2.26 (1.18–3.33) | 2.42 (1.14–3.70) |
| Study Cohort | Selenium, µg/dL Mean (95% CI) | Zinc, µg/dL Mean (95% CI) |
|---|---|---|
| COVID-19 vaccine | ||
| ABB C1® supplement | ||
| Baseline (n = 18) | 15.98 (13.99–17.98) | 102.33 (91.79–112.88) |
| Day 35 (n = 18) | 17.57 (15.57–19.57) | 119.28 (108.71–129.85) |
| Placebo | ||
| Baseline (n = 15) | 17.45 (15.26–19.64) | 106.20 (94.63–117.77) |
| Day 35 (n = 14) | 14.90 (12.67–17.13) | 102.64 (90.66–114.63) |
| Influenza vaccine | ||
| ABB C1® supplement | ||
| Baseline (n = 15) | 12.33 (11.15–13.51) | 91.40 (79.07–103–73) |
| Day 30 (n = 14) | 16.94 (15.74–18.14) | 116.75 (104.04–139.47) |
| Placebo | ||
| Baseline (n = 19) | 13.07 (12.02–14.12) | 91.32 (80.35–102.28) |
| Day 30 (n = 18) | 13.41 (12.35–14.47) | 102.23 (91.01–113.45) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodriguez, J.A.M.; Bifano, M.; Roca Goma, E.; Plasencia, C.M.; Torralba, A.O.; Font, M.S.; Millán, P.R. Effect and Tolerability of a Nutritional Supplement Based on a Synergistic Combination of β-Glucans and Selenium- and Zinc-Enriched Saccharomyces cerevisiae (ABB C1®) in Volunteers Receiving the Influenza or the COVID-19 Vaccine: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients 2021, 13, 4347. https://doi.org/10.3390/nu13124347
Rodriguez JAM, Bifano M, Roca Goma E, Plasencia CM, Torralba AO, Font MS, Millán PR. Effect and Tolerability of a Nutritional Supplement Based on a Synergistic Combination of β-Glucans and Selenium- and Zinc-Enriched Saccharomyces cerevisiae (ABB C1®) in Volunteers Receiving the Influenza or the COVID-19 Vaccine: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients. 2021; 13(12):4347. https://doi.org/10.3390/nu13124347
Chicago/Turabian StyleRodriguez, Julián Andrés Mateus, Mónica Bifano, Elvira Roca Goma, Carlos Méndez Plasencia, Anna Olivé Torralba, Mercè Santó Font, and Pedro Roy Millán. 2021. "Effect and Tolerability of a Nutritional Supplement Based on a Synergistic Combination of β-Glucans and Selenium- and Zinc-Enriched Saccharomyces cerevisiae (ABB C1®) in Volunteers Receiving the Influenza or the COVID-19 Vaccine: A Randomized, Double-Blind, Placebo-Controlled Study" Nutrients 13, no. 12: 4347. https://doi.org/10.3390/nu13124347
APA StyleRodriguez, J. A. M., Bifano, M., Roca Goma, E., Plasencia, C. M., Torralba, A. O., Font, M. S., & Millán, P. R. (2021). Effect and Tolerability of a Nutritional Supplement Based on a Synergistic Combination of β-Glucans and Selenium- and Zinc-Enriched Saccharomyces cerevisiae (ABB C1®) in Volunteers Receiving the Influenza or the COVID-19 Vaccine: A Randomized, Double-Blind, Placebo-Controlled Study. Nutrients, 13(12), 4347. https://doi.org/10.3390/nu13124347