
Chronic Effects of Maternal Low-Protein and Low-Quality Protein Diets on Body Composition, Glucose-Homeostasis and Metabolic Factors, Followed by Reversible Changes upon Rehabilitation in Adult Rat Offspring


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Design
2.1.1. Prepregnancy and Gestation
2.1.2. Birth and Lactation
2.1.3. Weaning and Post-Weaning
2.2. Whole-Body Composition Assessment by Dual-Energy X-ray Absorptiometry (DEXA)
2.3. Body and Organ Weights
2.4. Biochemical Analysis
2.5. Statistical Analysis
3. Results
3.1. Effects of the LQP and LP Diets on the Dams
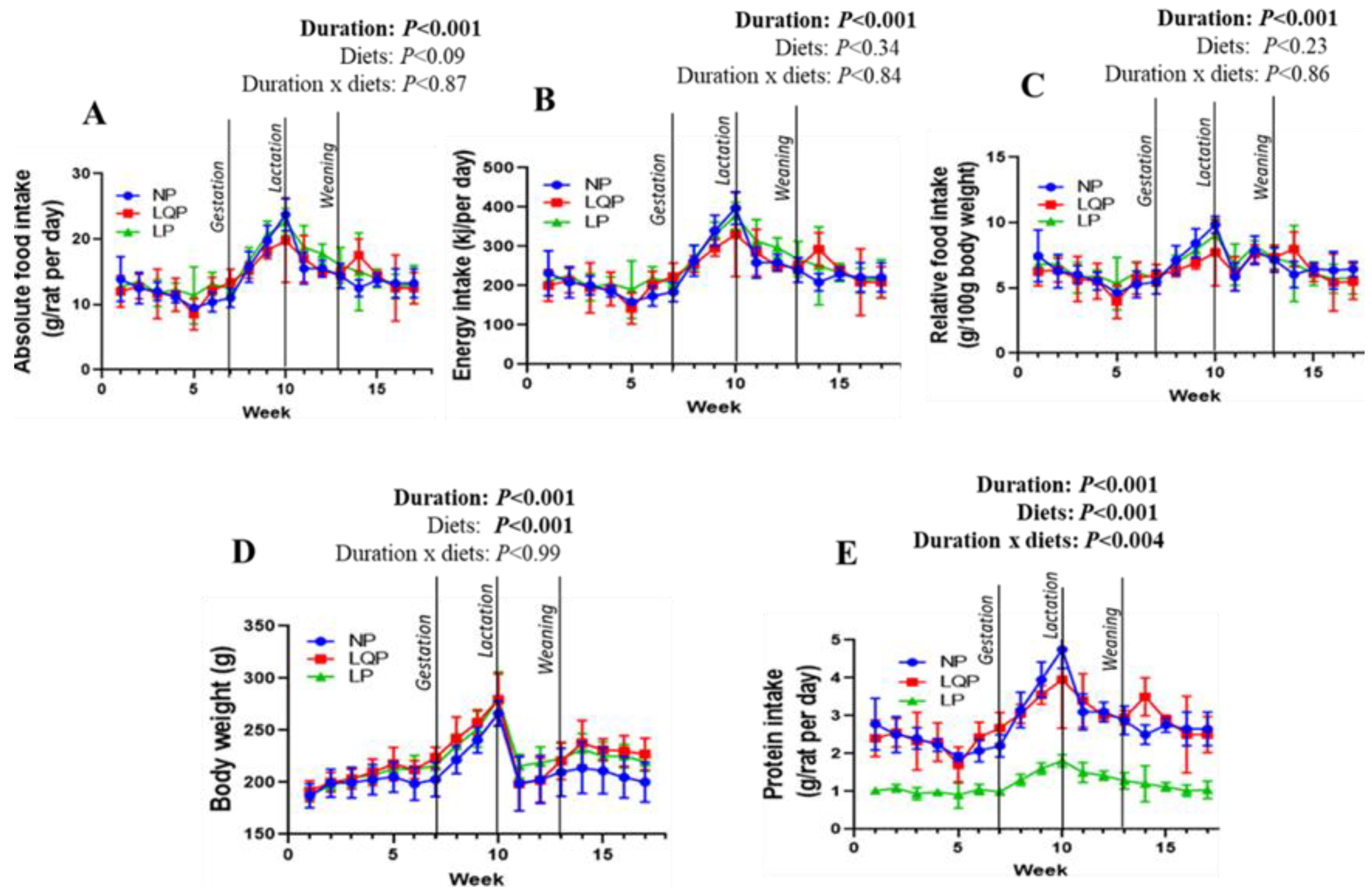
3.1.1. Food Intake and BWs in the Mothers
3.1.2. Body Composition and Metabolic Profile of the Mothers
3.1.3. Mating Success
3.2. Effect of MLQP and MLP Diets on the Offspring
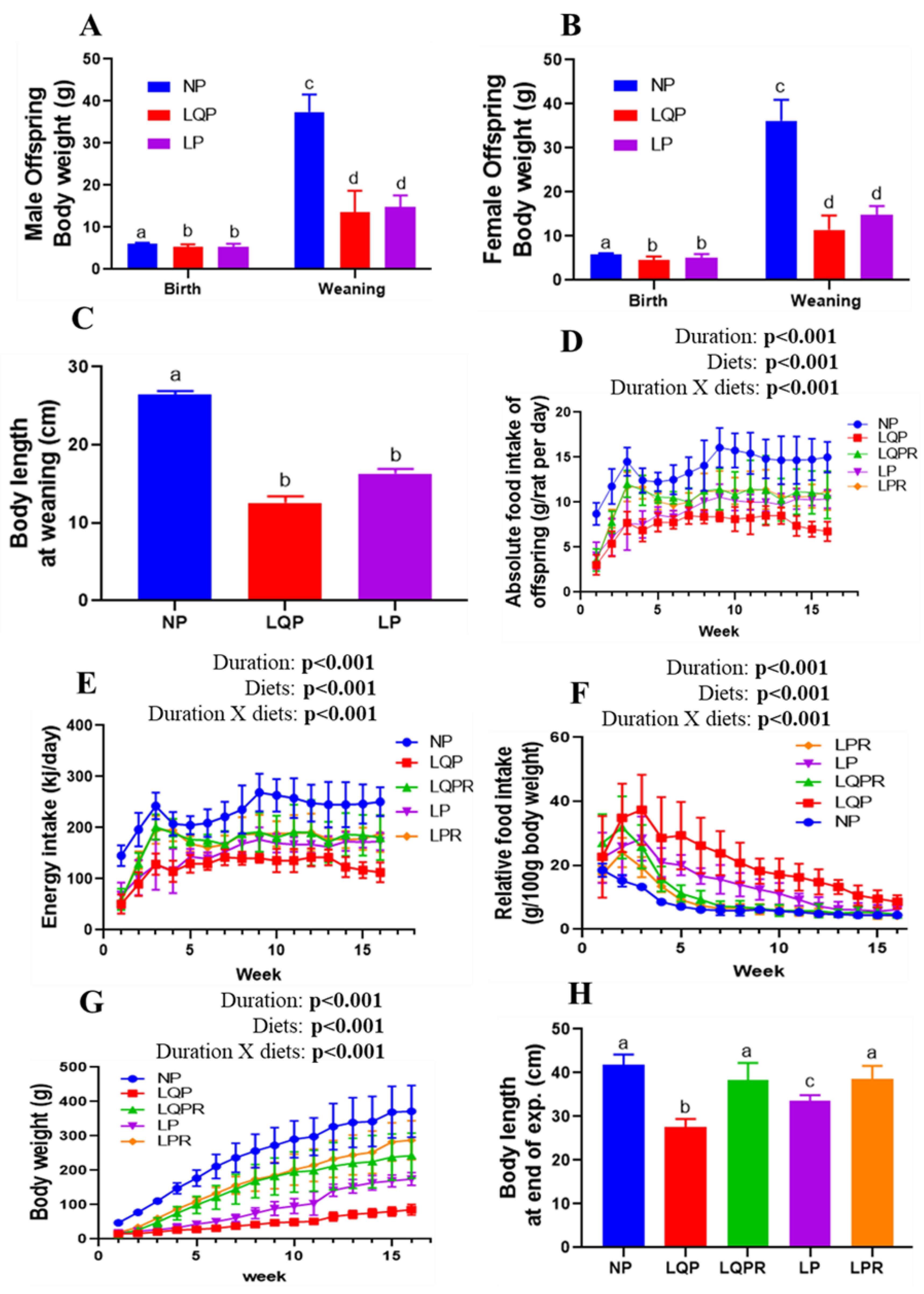
3.2.1. Bodyweight of the Pups at Birth and Weaning
3.2.2. Food Intake and BWs of the Offspring from Weaning to End of the Experiment
3.2.3. Whole-Body Composition of the Offspring
3.2.4. Absolute and Relative Organ Weights of the Offspring
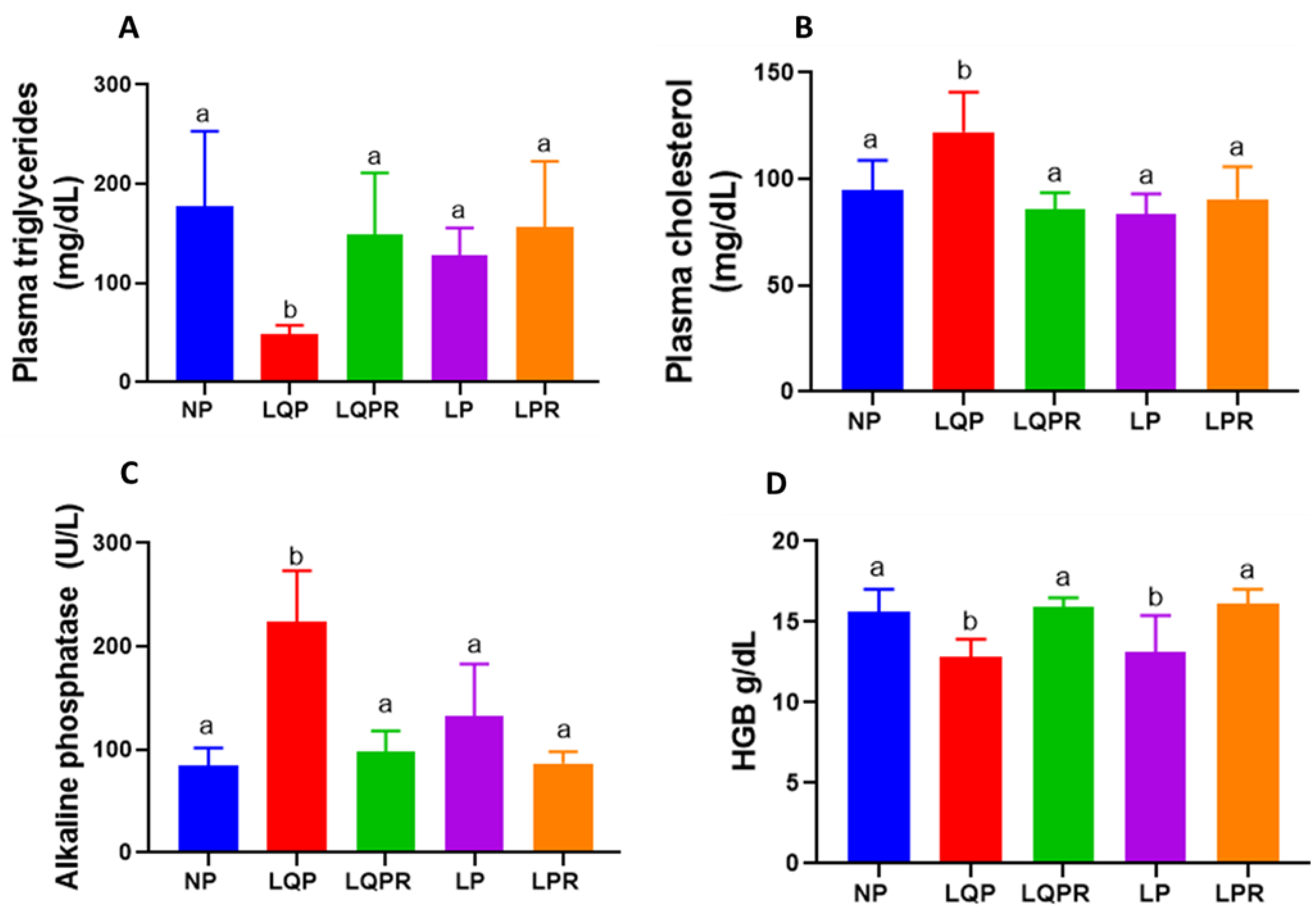
3.2.5. Lipid Profile
3.2.6. Alkaline Phosphatase and Hemoglobin
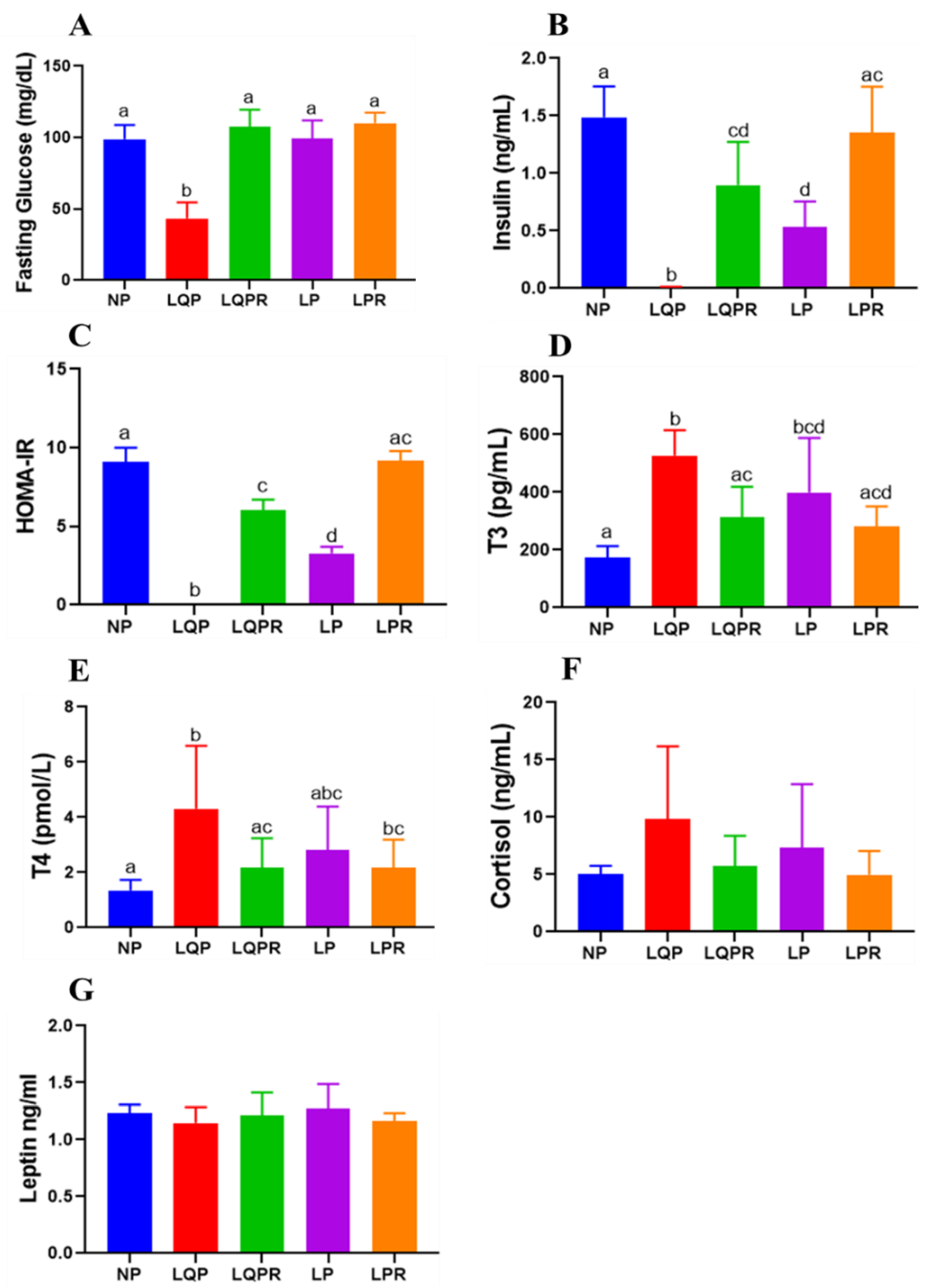
3.2.7. Glucose Homeostasis and HOMA-IR
3.2.8. Hormone Profile
4. Discussion
4.1. Imapct of the MLQP and MLP Diets in the Dams
4.2. Imapct of MLQP and MLP Diets in the Offspring
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Allman, B.R.; Andres, A.; Børsheim, E. The association of maternal protein intake during pregnancy in humans with maternal and offspring insulin sensitivity measures. Curr. Dev. Nutr. 2019, 3, nzz055. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. The fetal and infant origins of adult disease. BMJ 1990, 301, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, D.J. In utero programming of chronic disease. Clin. Sci. 1998, 95, 115–128. [Google Scholar] [CrossRef]
- Remacle, C.; Bieswal, F.; Bol, V.; Reusens, B. Developmental programming of adult obesity and cardiovascular disease in rodents by maternal nutrition imbalance. Am. J. Clin. Nutr. 2011, 94, 1846S–1852S. [Google Scholar] [CrossRef]
- Garcia, A.P.; Palou, M.; Sanchez, J.; Priego, T.; Palou, A.; Pico, C. Moderate caloric restriction during gestation in rats alters adipose tissue sympathetic innervation and later adiposity in offspring. PLoS ONE 2011, 6, e17313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Xiao, X.; Zhang, Q.; Wang, T.; Yu, M.; Xu, J. Maternal Low-Protein Diet Modulates Glucose Metabolism and Hepatic MicroRNAs Expression in the Early Life of Offspring dagger. Nutrients 2017, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Muller, O.; Krawinkel, M. Malnutrition and health in developing countries. CMAJ Can. Med Assoc. J. J. L’assoc. Med. Can. 2005, 173, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petry, C.J.; Ozanne, S.E.; Wang, C.L.; Hales, C.N. Effects of early protein restriction and adult obesity on rat pancreatic hormone content and glucose tolerance. Horm. Metab. Res. Horm. Stoffwechs. Horm. Metab. 2000, 32, 233–239. [Google Scholar] [CrossRef]
- Petry, C.J.; Dorling, M.W.; Pawlak, D.B.; Ozanne, S.E.; Hales, C.N. Diabetes in old male offspring of rat dams fed a reduced protein diet. Int. J. Exp. Diabetes Res. 2001, 2, 139–143. [Google Scholar] [CrossRef]
- Alejandro, E.U.; Gregg, B.; Wallen, T.; Kumusoglu, D.; Meister, D.; Chen, A.; Merrins, M.J.; Satin, L.S.; Liu, M.; Arvan, P.; et al. Maternal diet-induced microRNAs and mTOR underlie beta cell dysfunction in offspring. J. Clin. Investig. 2014, 124, 4395–4410. [Google Scholar] [CrossRef] [Green Version]
- Langley-Evans, S.C.; Phillips, G.J.; Jackson, A.A. In utero exposure to maternal low protein diets induces hypertension in weanling rats, independently of maternal blood pressure changes. Clin. Nutr. 1994, 13, 319–324. [Google Scholar] [CrossRef]
- Brawley, L.; Itoh, S.; Torrens, C.; Barker, A.; Bertram, C.; Poston, L.; Hanson, M. Dietary protein restriction in pregnancy induces hypertension and vascular defects in rat male offspring. Pediatric Res. 2003, 54, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bautista, C.J.; Rodriguez-Gonzalez, G.L.; Torres, N.; Hernandez-Pando, R.; Ramirez, V.; Rodriguez-Cruz, M.; Nathanielsz, P.W.; Zambrano, E. Protein restriction in the rat negatively impacts long-chain polyunsaturated fatty acid composition and mammary gland development at the end of gestation. Arch. Med. Res. 2013, 44, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Bautista, C.J.; Bautista, R.J.; Montano, S.; Reyes-Castro, L.A.; Rodriguez-Pena, O.N.; Ibanez, C.A.; Nathanielsz, P.W.; Zambrano, E. Effects of maternal protein restriction during pregnancy and lactation on milk composition and offspring development. Br. J. Nutr. 2019, 122, 141–151. [Google Scholar] [CrossRef]
- Fagundes, A.T.; Moura, E.G.; Passos, M.C.; Oliveira, E.; Toste, F.P.; Bonomo, I.T.; Trevenzoli, I.H.; Garcia, R.M.; Lisboa, P.C. Maternal low-protein diet during lactation programmes body composition and glucose homeostasis in the adult rat offspring. Br. J. Nutr. 2007, 98, 922–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahan-mihan, A.; Smith, C.E.; Anderson, G.H. Soy protein- and casein-based weaning diets differ in effects on food intake and blood glucose regulation in male Wistar rats. Nutr. Res. 2011, 31, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Jahan-mihan, A.; Smith, C.E.; Hamedani, A.; Anderson, G.H. Soy protein-based compared with casein-based diets fed during pregnancy and lactation increase food intake and characteristics of metabolic syndrome less in female than male rat offspring. Nutr. Res. 2011, 31, 644–651. [Google Scholar] [CrossRef]
- Millward, D.J.; Layman, D.K.; Tome, D.; Schaafsma, G. Protein quality assessment: Impact of expanding understanding of protein and amino acid needs for optimal health. Am. J. Clin. Nutr. 2008, 87, 1576S–1581S. [Google Scholar] [CrossRef]
- Shivakumar, N.; Minocha, S.; Kurpad, A.V. Protein quality & amino acid requirements in relation to needs in India. Indian J. Med. Res. 2018, 148, 557–568. [Google Scholar] [CrossRef]
- Schonfeldt, H.C.; Gibson Hall, N. Dietary protein quality and malnutrition in Africa. Br. J. Nutr. 2012, 108 (Suppl. 2), S69–S76. [Google Scholar] [CrossRef] [Green Version]
- Gilani, G.S. Background on international activities on protein quality assessment of foods. Br. J. Nutr. 2012, 108 (Suppl. 2), S168–S182. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Suri, D.; Uauy, R. Assessment of protein adequacy in developing countries: Quality matters. Br. J. Nutr. 2012, 108 (Suppl. 2), S77–S87. [Google Scholar] [CrossRef] [Green Version]
- Swaminathan, S.; Vaz, M.; Kurpad, A.V. Protein intakes in India. Br. J. Nutr. 2012, 108, S50–S58. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, S.; Varkey, A.; Shivakumar, N.; Devi, S.; Reddy, B.H.R.; Thomas, T.; Preston, T.; Sreeman, S.; Kurpad, A.V. True ileal digestibility of legumes determined by dual-isotope tracer method in Indian adults. Am. J. Clin. Nutr. 2019, 110, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Shivakumar, N.; Kashyap, S.; Kishore, S.; Thomas, T.; Varkey, A.; Devi, S.; Preston, T.; Jahoor, F.; Sheshshayee, M.; Kurpad, A.V. Protein-quality evaluation of complementary foods in Indian children. Am. J. Clin. Nutr. 2019, 109, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Minocha, S.; Thomas, T.; Kurpad, A.V. Dietary protein and the health–nutrition–agriculture connection in India. J. Nutr. 2017, 147, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altorf-van der Kuil, W.; Engberink, M.F.; Vedder, M.M.; Boer, J.M.; Verschuren, W.M.; Geleijnse, J.M. Sources of dietary protein in relation to blood pressure in a general Dutch population. PLoS ONE 2012, 7, e30582. [Google Scholar] [CrossRef] [Green Version]
- Alfenas Rde, C.; Bressan, J.; Paiva, A.C. Effects of protein quality on appetite and energy metabolism in normal weight subjects. Arq. Bras. Endocrinol. Metabol. 2010, 54, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Ellis, V. The acute effects of four protein meals on insulin, glucose, appetite and energy intake in lean men. Br. J. Nutr. 2010, 104, 1241–1248. [Google Scholar] [CrossRef]
- Alippi, R.M.; Picasso, E.; Huygens, P.; Bozzini, C.E.; Bozzini, C. Growth-dependent effects of dietary protein concentration and quality on the biomechanical properties of the diaphyseal rat femur. Endocrinol. Nutr. Organo Soc. Esp. Endocrinol. Nutr. 2012, 59, 35–43. [Google Scholar] [CrossRef]
- Bozzini, C.E.; Champin, G.M.; Alippi, R.M.; Bozzini, C. Biomechanical properties of the mandible, as assessed by bending test, in rats fed a low-quality protein. Arch. Oral Biol. 2013, 58, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Kabasakal Cetin, A.; Dasgin, H.; Gulec, A.; Onbasilar, I.; Akyol, A. Maternal Low Quality Protein Diet Alters Plasma Amino Acid Concentrations of Weaning Rats. Nutrients 2015, 7, 9847–9859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akyol, A.; Cetin, A.K.; Gulec, A.; Dasgin, H.; Ayaz, A.; Onbasilar, I. Maternal low-quality protein diet exerts sex-specific effects on plasma amino acid profile and alters hepatic expression of methyltransferases in adult rat offspring. J. Dev. Orig. Health Dis. 2018, 9, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Carlin, G.; Chaumontet, C.; Blachier, F.; Barbillon, P.; Darcel, N.; Delteil, C.; van der Beek, E.M.; Kodde, A.; van de Heijning, B.J.M.; Tome, D.; et al. Perinatal exposure of rats to a maternal diet with varying protein quantity and quality affects the risk of overweight in female adult offspring. J. Nutr. Biochem. 2020, 79, 108333. [Google Scholar] [CrossRef]
- Zohdi, V.; Lim, K.; Pearson, J.T.; Black, M.J. Developmental programming of cardiovascular disease following intrauterine growth restriction: Findings utilising a rat model of maternal protein restriction. Nutrients 2014, 7, 119–152. [Google Scholar] [CrossRef] [Green Version]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Pezeshki, A.; Chelikani, P.K. Low Protein Diets and Energy Balance: Mechanisms of Action on Energy Intake and Expenditure. Front. Nutr. 2021, 8, 655833. [Google Scholar] [CrossRef]
- Clowes, E.J.; Aherne, F.X.; Baracos, V.E. Skeletal muscle protein mobilization during the progression of lactation. Am. J. Physiol. Endocrinol. Metab. 2005, 288, E564–E572. [Google Scholar] [CrossRef]
- Jansen, G.R.; Binard, R.; Longenecker, J.B. Protein quality and quantity influence free amino acid levels in the brain and serum of rats during lactation. J. Nutr. 1991, 121, 1187–1194. [Google Scholar] [CrossRef] [Green Version]
- Zambrano, E.; Bautista, C.J.; Deas, M.; Martinez-Samayoa, P.M.; Gonzalez-Zamorano, M.; Ledesma, H.; Morales, J.; Larrea, F.; Nathanielsz, P.W. A low maternal protein diet during pregnancy and lactation has sex- and window of exposure-specific effects on offspring growth and food intake, glucose metabolism and serum leptin in the rat. J. Physiol. 2006, 571, 221–230. [Google Scholar] [CrossRef]
- Shiraki, N.; Shiraki, Y.; Tsuyama, T.; Obata, F.; Miura, M.; Nagae, G.; Aburatani, H.; Kume, K.; Endo, F.; Kume, S. Methionine metabolism regulates maintenance and differentiation of human pluripotent stem cells. Cell Metab. 2014, 19, 780–794. [Google Scholar] [CrossRef] [Green Version]
- Van Winkle, L.J.; Galat, V.; Iannaccone, P.M. Lysine Deprivation during Maternal Consumption of Low-Protein Diets Could Adversely Affect Early Embryo Development and Health in Adulthood. Int. J. Environ. Res. Public Health 2020, 17, 5462. [Google Scholar] [CrossRef]
- Aissa, A.F.; Gomes, T.D.; Almeida, M.R.; Hernandes, L.C.; Darin, J.D.; Bianchi, M.L.; Antunes, L.M. Methionine concentration in the diet has a tissue-specific effect on chromosomal stability in female mice. Food Chem. Toxicol. 2013, 62, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Lee, K.S.; Kwon, D.H.; Bong, J.J.; Jeong, J.Y.; Nam, Y.S.; Lee, M.S.; Liu, X.; Baik, M. Severe dietary lysine restriction affects growth and body composition and hepatic gene expression for nitrogen metabolism in growing rats. J. Anim. Physiol. Anim. Nutr. 2014, 98, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter, J.E.; O’Brien, K.O.; Insogna, K.L. Dietary protein, calcium metabolism, and skeletal homeostasis revisited. Am. J. Clin. Nutr. 2003, 78, 584S–592S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerstetter, J.E.; O’Brien, K.O.; Insogna, K.L. Low protein intake: The impact on calcium and bone homeostasis in humans. J. Nutr. 2003, 133, 855S–861S. [Google Scholar] [CrossRef] [Green Version]
- Marin, M.C.; De Tomas, M.E.; Serres, C.; Mercuri, O. Protein-energy malnutrition during gestation and lactation in rats affects growth rate, brain development and essential fatty acid metabolism. J. Nutr. 1995, 125, 1017–1024. [Google Scholar] [CrossRef]
- Desai, M.; Crowther, N.J.; Lucas, A.; Hales, C.N. Organ-selective growth in the offspring of protein-restricted mothers. Br. J. Nutr. 1996, 76, 591–603. [Google Scholar] [CrossRef] [Green Version]
- Kwon, D.-H.; Kang, W.; Nam, Y.S.; Lee, M.S.; Lee, I.Y.; Kim, H.J.; Rajasekar, P.; Lee, J.-H.; Baik, M. Dietary protein restriction induces steatohepatitis and alters leptin/signal transducers and activators of transcription 3 signaling in lactating rats. J. Nutr. Biochem. 2012, 23, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Erhuma, A.; Salter, A.M.; Sculley, D.V.; Langley-Evans, S.C.; Bennett, A.J. Prenatal exposure to a low-protein diet programs disordered regulation of lipid metabolism in the aging rat. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1702–E1714. [Google Scholar] [CrossRef] [Green Version]
- Fetoui, H.; Mahjoubi-Samet, A.; Guermazi, F.; Zeghal, N. Maternal low-protein diet affects bone mass and mineral metabolism in suckling rats. J. Anim. Physiol. Anim. Nutr. 2008, 92, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Kanakis, I.; Alameddine, M.; Scalabrin, M.; van ‘t Hof, R.J.; Liloglou, T.; Ozanne, S.E.; Goljanek-Whysall, K.; Vasilaki, A. Low protein intake during reproduction compromises the recovery of lactation-induced bone loss in female mouse dams without affecting skeletal muscles. FASEB J. 2020, 34, 11844–11859. [Google Scholar] [CrossRef] [PubMed]
- Kirigiti, M.A.; Frazee, T.; Bennett, B.; Arik, A.; Blundell, P.; Bader, L.; Bagley, J.; Frias, A.E.; Sullivan, E.L.; Roberts, C.T., Jr.; et al. Effects of pre- and postnatal protein restriction on maternal and offspring metabolism in the nonhuman primate. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R929–R939. [Google Scholar] [CrossRef]
- Santos, E.W.; Oliveira, D.C.; Silva, G.B.; Tsujita, M.; Beltran, J.O.; Hastreiter, A.; Fock, R.A.; Borelli, P. Hematological alterations in protein malnutrition. Nutr. Rev. 2017, 75, 909–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palkowska-Gozdzik, E.; Lachowicz, K.; Rosolowska-Huszcz, D. Effects of Dietary Protein on Thyroid Axis Activity. Nutrients 2017, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Passos, M.C.; da Fonte Ramos, C.; Dutra, S.C.; Mouco, T.; de Moura, E.G. Long-term effects of malnutrition during lactation on the thyroid function of offspring. Horm. Metab. Res. Horm Stoffwechs. Horm. Metab. 2002, 34, 40–43. [Google Scholar] [CrossRef] [PubMed]
- Ekelund, U.; Ong, K.; Linné, Y.; Neovius, M.; Brage, S.; Dunger, D.B.; Wareham, N.J.; Rössner, S. Upward weight percentile crossing in infancy and early childhood independently predicts fat mass in young adults: The Stockholm Weight Development Study (SWEDES). Am. J. Clin. Nutr. 2006, 83, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Sachdev, H.S.; Fall, C.H.; Osmond, C.; Lakshmy, R.; Dey Biswas, S.K.; Leary, S.D.; Reddy, K.S.; Barker, D.J.; Bhargava, S.K. Anthropometric indicators of body composition in young adults: Relation to size at birth and serial measurements of body mass index in childhood in the New Delhi birth cohort. Am. J. Clin. Nutr. 2005, 82, 456–466. [Google Scholar] [CrossRef]
- Ozanne, S.; Olsen, G.; Hansen, L.; Tingey, K.; Nave, B.; Wang, C.; Hartil, K.; Petry, C.; Buckley, A.; Mosthaf-Seedorf, L. Early growth restriction leads to down regulation of protein kinase C zeta and insulin resistance in skeletal muscle. J. Endocrinol. 2003, 177, 235–242. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | NP | LQP | LP |
|---|---|---|---|
| Metabolizable energy (kcal/g of diet) | 3.9 | 3.9 | 3.9 |
| kJ/g of diet | 16.3 | 16.3 | 16.3 |
| Proteins (% energy) | 20 | 20 | 8 |
| Carbohydrates (% energy) | 64 | 64 | 76 |
| Lipids (% energy) | 16 | 16 | 16 |
| Ingredients (g/kg of diet) | |||
| Starch | 495 | 495 | 536.5 |
| Casein | 200 | 0 | 80 |
| Wheat gluten | 0 | 200 | 0 |
| Sucrose | 132.5 | 132.5 | 212.5 |
| Ground Nut Oil | 70 | 70 | 70 |
| Cellulose | 50 | 50 | 50 |
| Mineral Mix (AIN-93G-MX) 1 | 35 | 35 | 35 |
| Vitamin Mix (AIN-93-VX) 2 | 10 | 10 | 10 |
| L.Cystine | 3 | 3 | 1.5 |
| Choline Chloride | 2.5 | 2.5 | 2.5 |
| PABA | 1 | 1 | 1 |
| Inositol | 1 | 1 | 1 |
| Four Weeks | NP (n = 4) | LQP (n = 4) | LP (n = 4) |
|---|---|---|---|
| Area (cm2) | 45.68 ± 3.65 | 47.06 ± 1.20 | 46.64 ± 1.20 |
| BMC(g) | 7.24 ± 0.44 | 7.39 ± 0.43 | 7.28 ± 0.25 |
| BMD (g/cm2) | 0.15 ± 0.004 | 0.15 ± 0.003 | 0.15 ± 0.004 |
| Total mass (g) | 197.72 ± 16.17 | 211 ± 13.65 | 211.77 ± 12.97 |
| Lean mass (g) | 148.80 ± 8.87 | 150.58 ± 2.46 | 147.88 ± 4.12 |
| % LBM | 79.07 ± 3.42 | 75.06 ± 4.17 | 73.41 ± 3.40 |
| Fat mass (g) | 41.7 ± 9.63 | 53.05 ± 11.77 | 56.57 ± 10.34 |
| % BF | 20.95 ± 3.41 | 24.95 ± 4.18 | 26.5 ± 3.40 |
| Fasting Glucose (mg/dL) | 77.25 ± 6.07 | 74.5 ± 5.97 | 73.75 ± 7.5 |
| At weaning | |||
| Area (cm2) | 48.02 ± 2.30 | 48.92 ± 1.88 | 49.4 ± 2.07 |
| BMC(g) | 6.95 ± 0.54 | 7.24 ± 0.56 | 7.21 ± 0.51 |
| BMD (g/cm2) | 0.14 ± 0.005 | 0.148 ± 0.007 | 0.146 ± 0.007 |
| Total mass (g) | 207.15 ± 25.34 | 207.27 ± 17.01 | 229.02 ± 13.61 |
| Lean mass (g) | 180.32 ± 18.94 | 160.82 ± 8.43 | 173.6 ± 3.05 |
| % LBM | 90.58 ± 2.17 a | 81.22 ± 2.35 b | 79.12 ± 4.08 b |
| Fat mass (g) | 19.85 ± 6.3 a | 39.22 ± 8.35 b | 48.2 ± 11.74 b |
| % BF | 9.4 ± 2.18 a | 18.77 ± 2.33 b | 20.87 ± 4.11 b |
| Fasting Glucose (mg/dL) | 80.5 ± 12.12 | 62.75 ± 3.30 | 72.5 ± 12.92 |
| Plasma cholesterol (mg/dL) | 88.61 ± 15.84 | 84.81 ± 22.87 | 76.23 ± 8.69 |
| Plasma triglycerides (mg/dL) | 85.43 ± 55.01 | 126.37 ± 66.19 | 230.31 ± 95.09 |
| NP (n = 6) | LQP (n = 6) | LP (n = 6) | |
|---|---|---|---|
| Area (cm2) | 26.2 ± 1.82 a | 4.32 ± 0.59 b | 7.84 ± 1.20 c |
| BMC(g) | 2.15 ± 0.14 a | 0.34 ± 0.04 b | 0.66 ± 0.10 c |
| BMD (g/cm2) | 0.082 ± 0.001 ab | 0.080 ± 0.003 a | 0.085 ± 0.02 b |
| Total mass (g) | 98.4 ± 5.94 a | 12.18 ± 1.53 b | 25.43 ± 2.53 c |
| Lean mass (g) | 78.17 ± 5.07 a | 10.93 ± 1.17 b | 21.80 ± 2.00 c |
| % LBM | 81.64 ± 2.06 a | 92.79 ± 2.64 b | 88.40 ± 2.13 c |
| Fat mass (g) | 18.08 ± 2.28 a | 0.91 ± 0.40 b | 2.95 ± 0.65 b |
| % BF | 18.4 ± 2.05 a | 7.28 ± 2.56 b | 11.58 ± 2.15 b |
| NP (n = 6) | LQP (n = 6) | LQPR (n = 6) | LP (n = 6) | LPR (n = 6) | |
|---|---|---|---|---|---|
| Area (cm2) | 57.48 ± 8.33 a | 15.02 ± 3.39 b | 44.29 ± 5.12 c | 30.81 ± 2.57 d | 46.92 ± 8.28 c |
| BMC(g) | 8.43 ± 1.32 a | 1.51 ± 0.38 b | 6.05 ± 0.92 c | 3.33 ± 0.42 d | 6.5 ± 1.16 c |
| BMD (g/cm2) | 0.14 ± 0.007 a | 0.10 ± 0.01 b | 0.13 ± 0.006 a | 0.10 ± 0.006 b | 0.13 ± 0.007 a |
| Total mass (g) | 311.58 ± 64.93 a | 52.2 ± 9.38 b | 205.75 ± 49.25 c | 119.45 ± 18.03 b | 234.11 ± 55.01 cd |
| Lean mass (g) | 233.74 ± 49.05 a | 45.48 ± 8.06 b | 164.06 ± 39.9 c | 87.48 ± 12.2 b | 180.71 ± 43.53 ac |
| % LBM | 77.76 ± 2.77 a | 90.13 ± 4.49 b | 82.83 ± 4.07 ac | 76.19 ± 2.21 ac | 79.92 ± 1.85 ac |
| Fat mass (g) | 69.41 ± 18.36 a | 5.21 ± 2.85 b | 35.63 ± 12.52 c | 28.65 ± 5.98 c | 46.88 ± 11.37 c |
| % BF | 22.23 ± 2.75 a | 9.91 ± 4.42 b | 17.16 ± 4.05 ac | 23.81 ± 2.1 ac | 20.08 ± 1.87 ac |
| Organs | NP (n = 8) | LQP (n = 8) | LQPR (n = 8) | LP (n = 8) | LPR (n = 8) |
|---|---|---|---|---|---|
| Bodyweight (BW; g) | 353.12 ± 78.09 a | 85.62 ± 15.37 b | 241. ± 63.43 c | 168.12 ± 22.75 c,d | 272.5 ± 57.07 c |
| Brain (g) | 1.8 ± 0.12 a | 1.43 ± 0.13 b | 1.69 ± 0.16 a,c | 1.52 ± 0.13 b,c,d | 1.69 ± 0.13 a,c,d |
| Liver (g) | 12.05 ± 2.87 a | 3.35 ± 0.06 b | 8.88 ± 2.24 c | 7.2 ± 1.04 c,d | 10.53 ± 2.5b a,c |
| Kidney (g) | 2.62 ± 0.6 a | 0.8 ± 0.15 b | 1.88 ± 0.47 c | 1.17 ± 0.14 b | 1.89 ± 0.34 c |
| Heart (g) | 1.12 ± 0.15 a | 0.47 ± 0.07 b | 0.91 ± 0.19 a,c | 0.79 ± 0.15 c | 0.95 ± 0.14 a,c |
| Pancreas (g) | 0.81 ± 0.3 a | 0.29 ± 0.08 b | 0.66 ± 0.2 a,c | 0.43 ± 0.12 b,c,d | 0.61 ± 0.19 a,c,d |
| Brain weight/BW (%) | 0.53 ± 0.13 a | 1.7 ± 0.19 b | 0.73 ± 0.16 a,c | 0.91 ± 0.14 cd | 0.64 ± 0.15b a,c |
| Liver weight/BW (%) | 3.41 ± 0.24 a | 3.9 ± 0.61 ab | 3.7 ± 0.22 a | 4.36 ± 0.66 b | 3.83 ± 0.32 a,b |
| Kidney weight/BW (%) | 0.74 ± 0.06 a | 0.94 ± 0.11 b | 0.78 ± 0.03 a | 0.7 ± 0.07 a | 0.7 ± 0.04 a |
| Heart weight/BW (%) | 0.32 ± 0.04 a | 0.56 ± 0.08 b | 0.38 ± 0.04 a | 0.46 ± 0.05 c | 0.35 ± 0.03 a |
| Pancreas weight/BW (%) | 0.22 ± 0.06 a | 0.35 ± 0.11 b | 0.28 ± 0.1 a,b | 0.25 ± 0.07 a,b | 0.22 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savitikadi, P.; Pullakhandam, R.; Kulkarni, B.; Kumar, B.N.; Reddy, G.B.; Reddy, V.S. Chronic Effects of Maternal Low-Protein and Low-Quality Protein Diets on Body Composition, Glucose-Homeostasis and Metabolic Factors, Followed by Reversible Changes upon Rehabilitation in Adult Rat Offspring. Nutrients 2021, 13, 4129. https://doi.org/10.3390/nu13114129
Savitikadi P, Pullakhandam R, Kulkarni B, Kumar BN, Reddy GB, Reddy VS. Chronic Effects of Maternal Low-Protein and Low-Quality Protein Diets on Body Composition, Glucose-Homeostasis and Metabolic Factors, Followed by Reversible Changes upon Rehabilitation in Adult Rat Offspring. Nutrients. 2021; 13(11):4129. https://doi.org/10.3390/nu13114129
Chicago/Turabian StyleSavitikadi, Pandarinath, Raghu Pullakhandam, Bharati Kulkarni, Boiroju Naveen Kumar, Geereddy Bhanuprakash Reddy, and Vadde Sudhakar Reddy. 2021. "Chronic Effects of Maternal Low-Protein and Low-Quality Protein Diets on Body Composition, Glucose-Homeostasis and Metabolic Factors, Followed by Reversible Changes upon Rehabilitation in Adult Rat Offspring" Nutrients 13, no. 11: 4129. https://doi.org/10.3390/nu13114129
APA StyleSavitikadi, P., Pullakhandam, R., Kulkarni, B., Kumar, B. N., Reddy, G. B., & Reddy, V. S. (2021). Chronic Effects of Maternal Low-Protein and Low-Quality Protein Diets on Body Composition, Glucose-Homeostasis and Metabolic Factors, Followed by Reversible Changes upon Rehabilitation in Adult Rat Offspring. Nutrients, 13(11), 4129. https://doi.org/10.3390/nu13114129