
Epigenome-Wide Association Study of Infant Feeding and DNA Methylation in Infancy and Childhood in a Population at Increased Risk for Type 1 Diabetes

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
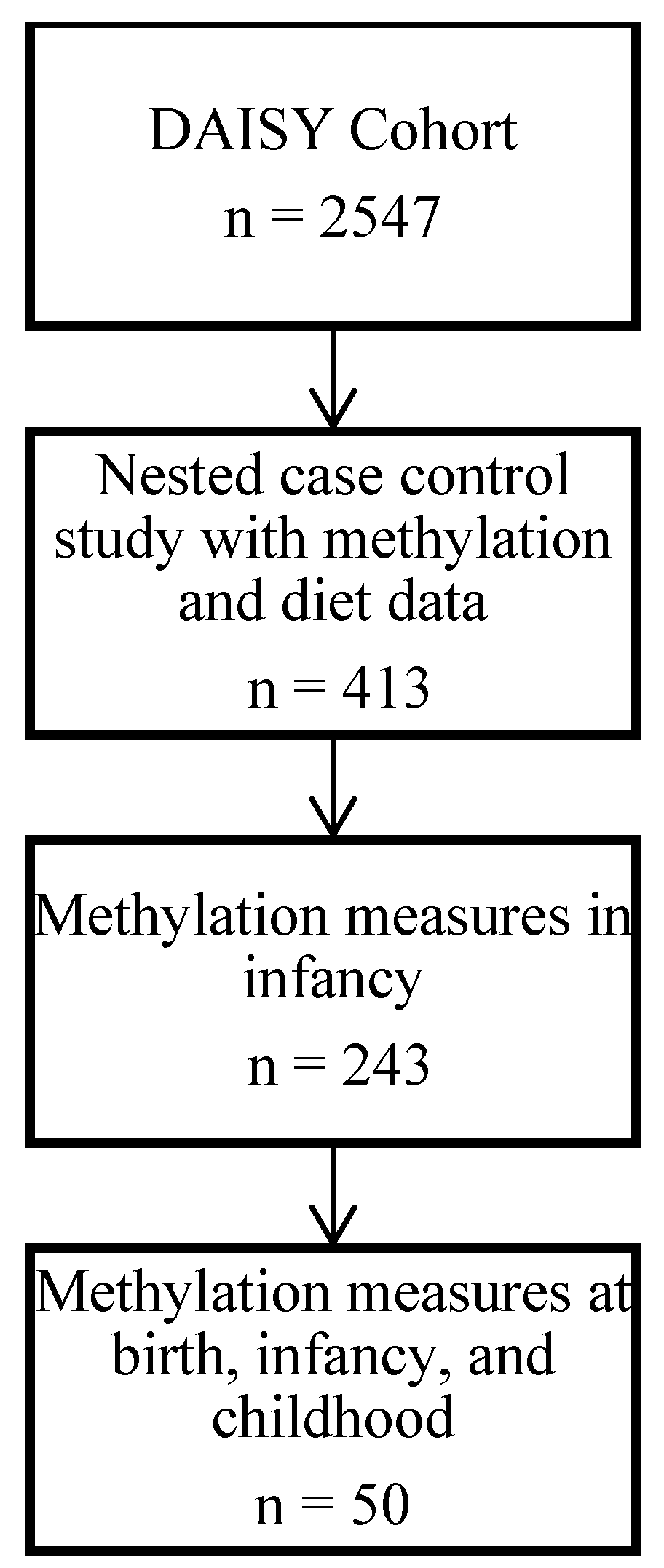
2.1. Study Population
2.2. Measurement of DNA Methylation
2.3. Infant Diet Variables
2.4. Statistical Analyses
2.5. Analysis of Methylation Associations in Childhood and at Birth
3. Results
3.1. Methylation in Infancy
3.2. Confirmation of Associations from Previous Literature
3.3. Methylation in Childhood
3.4. Methylation at Birth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, J.; Liu, L.; Zhu, Y.; Huang, G.; Wang, P.P. The association between breastfeeding and childhood obesity: A meta-analysis. BMC Public Health 2014, 14, 1267. [Google Scholar] [CrossRef] [Green Version]
- Horta, B.L.; de Lima, N.P. Breastfeeding and Type 2 Diabetes: Systematic Review and Meta-Analysis. Curr. Diab. Rep. 2019, 19, 1. [Google Scholar] [CrossRef]
- Gungor, D.; Nadaud, P.; LaPergola, C.C.; Dreibelbis, C.; Wong, Y.P.; Terry, N.; Abrams, S.A.; Beker, L.; Jacobovits, T.; Jarvinen, K.M.; et al. Infant milk-feeding practices and diabetes outcomes in offspring: A systematic review. Am. J. Clin. Nutr. 2019, 109, 817S–837S. [Google Scholar] [CrossRef] [PubMed]
- Butte, N.F.; Lopez-Alarson, M.G.; Garza, C. Expert Consultation on the Optimal Duration of Exclusive Breastfeeding. In Nutrient Adequacy of Exclusive Breastfeeding for the Term Infant during the First Six Months of Life; World Health Organization: Geneva, Switzerland, 2002; pp. 1–49. [Google Scholar]
- Logan, K.; Perkin, M.R.; Marrs, T.; Radulovic, S.; Craven, J.; Flohr, C.; Bahnson, H.T.; Lack, G. Early Gluten Introduction and Celiac Disease in the EAT Study: A Prespecified Analysis of the EAT Randomized Clinical Trial. JAMA Pediatr. 2020, 174, 1041–1047. [Google Scholar] [CrossRef]
- Norris, J.M.; Barriga, K.; Hoffenberg, E.J.; Taki, I.; Miao, D.; Haas, J.E.; Emery, L.M.; Sokol, R.J.; Erlich, H.A.; Eisenbarth, G.S.; et al. Risk of celiac disease autoimmunity and timing of gluten introduction in the diet of infants at increased risk of disease. JAMA 2005, 293, 2343–2351. [Google Scholar] [CrossRef] [Green Version]
- Lionetti, E.; Castellaneta, S.; Francavilla, R.; Pulvirenti, A.; Tonutti, E.; Amarri, S.; Barbato, M.; Barbera, C.; Barera, G.; Bellantoni, A.; et al. Introduction of gluten, HLA status, and the risk of celiac disease in children. N. Engl. J. Med. 2014, 371, 1295–1303. [Google Scholar] [CrossRef] [Green Version]
- Vriezinga, S.L.; Auricchio, R.; Bravi, E.; Castillejo, G.; Chmielewska, A.; Crespo Escobar, P.; Kolacek, S.; Koletzko, S.; Korponay-Szabo, I.R.; Mummert, E.; et al. Randomized feeding intervention in infants at high risk for celiac disease. N. Engl. J. Med. 2014, 371, 1304–1315. [Google Scholar] [CrossRef] [Green Version]
- Frederiksen, B.; Kroehl, M.; Lamb, M.M.; Seifert, J.; Barriga, K.; Eisenbarth, G.S.; Rewers, M.; Norris, J.M. Infant exposures and development of type 1 diabetes mellitus: The Diabetes Autoimmunity Study in the Young (DAISY). JAMA Pediatr. 2013, 167, 808–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, J.M.; Barriga, K.; Klingensmith, G.; Hoffman, M.; Eisenbarth, G.S.; Erlich, H.A.; Rewers, M. Timing of initial cereal exposure in infancy and risk of islet autoimmunity. JAMA 2003, 290, 1713–1720. [Google Scholar] [CrossRef]
- Virtanen, S.M.; Kenward, M.G.; Erkkola, M.; Kautiainen, S.; Kronberg-Kippila, C.; Hakulinen, T.; Ahonen, S.; Uusitalo, L.; Niinisto, S.; Veijola, R.; et al. Age at introduction of new foods and advanced beta cell autoimmunity in young children with HLA-conferred susceptibility to type 1 diabetes. Diabetologia 2006, 49, 1512–1521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virtanen, S.M.; Takkinen, H.M.; Nevalainen, J.; Kronberg-Kippila, C.; Salmenhaara, M.; Uusitalo, L.; Kenward, M.G.; Erkkola, M.; Veijola, R.; Simell, O.; et al. Early introduction of root vegetables in infancy associated with advanced ss-cell autoimmunity in young children with human leukocyte antigen-conferred susceptibility to Type 1 diabetes. Diabet. Med. 2011, 28, 965–971. [Google Scholar] [CrossRef]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Bedogni, G.; Brambilla, P.; Cianfarani, S.; Nobili, V.; Pietrobelli, A.; Agostoni, C. Epigenetic mechanisms elicited by nutrition in early life. Nutr. Res. Rev. 2011, 24, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Hartwig, F.P.; Loret de Mola, C.; Davies, N.M.; Victora, C.G.; Relton, C.L. Breastfeeding effects on DNA methylation in the offspring: A systematic literature review. PLoS ONE 2017, 12, e0173070. [Google Scholar] [CrossRef] [Green Version]
- Pauwels, S.; Symons, L.; Vanautgaerden, E.L.; Ghosh, M.; Duca, R.C.; Bekaert, B.; Freson, K.; Huybrechts, I.; Langie, S.A.S.; Koppen, G.; et al. The Influence of the Duration of Breastfeeding on the Infant’s Metabolic Epigenome. Nutrients 2019, 11, 1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherwood, W.B.; Bion, V.; Lockett, G.A.; Ziyab, A.H.; Soto-Ramirez, N.; Mukherjee, N.; Kurukulaaratchy, R.J.; Ewart, S.; Zhang, H.; Arshad, S.H.; et al. Duration of breastfeeding is associated with leptin (LEP) DNA methylation profiles and BMI in 10-year-old children. Clin. Epigenet. 2019, 11, 128. [Google Scholar] [CrossRef]
- Sherwood, W.B.; Kothalawala, D.M.; Kadalayil, L.; Ewart, S.; Zhang, H.; Karmaus, W.; Arshad, S.H.; Holloway, J.W.; Rezwan, F.I. Epigenome-Wide Association Study Reveals Duration of Breastfeeding Is Associated with Epigenetic Differences in Children. Int. J. Environ. Res. Public Health 2020, 17, 3569. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, V.V.; Hagenbeek, F.A.; Suderman, M.; Caramaschi, D.; van Beijsterveldt, C.E.M.; Kallsen, N.A.; Ehli, E.A.; Davies, G.E.; Sukhikh, G.T.; Fanos, V.; et al. DNA Methylation Signatures of Breastfeeding in Buccal Cells Collected in Mid-Childhood. Nutrients 2019, 11, 2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwig, F.P.; Davey Smith, G.; Simpkin, A.J.; Victora, C.G.; Relton, C.L.; Caramaschi, D. Association between Breastfeeding and DNA Methylation over the Life Course: Findings from the Avon Longitudinal Study of Parents and Children (ALSPAC). Nutrients 2020, 12, 3309. [Google Scholar] [CrossRef] [PubMed]
- Rewers, M.; Bugawan, T.L.; Norris, J.M.; Blair, A.; Beaty, B.; Hoffman, M.; McDuffie, R.S., Jr.; Hamman, R.F.; Klingensmith, G.; Eisenbarth, G.S.; et al. Newborn screening for HLA markers associated with IDDM: Diabetes autoimmunity study in the young (DAISY). Diabetologia 1996, 39, 807–812. [Google Scholar] [CrossRef]
- Rewers, M.; Norris, J.M.; Eisenbarth, G.S.; Erlich, H.A.; Beaty, B.; Klingensmith, G.; Hoffman, M.; Yu, L.; Bugawan, T.L.; Blair, A.; et al. Beta-cell autoantibodies in infants and toddlers without IDDM relatives: Diabetes autoimmunity study in the young (DAISY). J. Autoimmun. 1996, 9, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.D. Assessing intrauterine influences on offspring health outcomes: Can epidemiological studies yield robust findings? Basic Clin. Pharmacol. Toxicol. 2008, 102, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Triche, T.J., Jr.; Laird, P.W.; Shen, H. SeSAMe: Reducing artifactual detection of DNA methylation by Infinium BeadChips in genomic deletions. Nucleic Acids Res. 2018, 46, e123. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.K.; Vanderlinden, L.A.; Dong, F.; Carry, P.M.; Seifert, J.; Waugh, K.; Shorrosh, H.; Fingerlin, T.; Frohnert, B.I.; Yang, I.V.; et al. Longitudinal DNA methylation differences precede type 1 diabetes. Sci. Rep. 2020, 10, 3721. [Google Scholar] [CrossRef] [Green Version]
- Vanderlinden, L.A.; Johnson, R.K.; Carry, P.M.; Dong, F.; DeMeo, D.L.; Yang, I.V.; Norris, J.M.; Kechris, K. An effective processing pipeline for harmonizing DNA methylation data from Illumina’s 450K and EPIC platforms for epidemiological studies. BMC Res. Notes 2021, 14, 352. [Google Scholar] [CrossRef] [PubMed]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinform. 2012, 13, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffe, A.E.; Irizarry, R.A. Accounting for cellular heterogeneity is critical in epigenome-wide association studies. Genome Biol. 2014, 15, R31. [Google Scholar] [CrossRef] [PubMed]
- van Iterson, M.; van Zwet, E.W.; Consortium, B.; Heijmans, B.T. Controlling bias and inflation in epigenome- and transcriptome-wide association studies using the empirical null distribution. Genome Biol. 2017, 18, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juvinao-Quintero, D.L.; Marioni, R.E.; Ochoa-Rosales, C.; Russ, T.C.; Deary, I.J.; van Meurs, J.B.J.; Voortman, T.; Hivert, M.F.; Sharp, G.C.; Relton, C.L.; et al. DNA methylation of blood cells is associated with prevalent type 2 diabetes in a meta-analysis of four European cohorts. Clin. Epigenet. 2021, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Bae, J.H.; Park, K.S.; Sung, J.; Kwak, S.H. DNA Methylation Changes Associated with Type 2 Diabetes and Diabetic Kidney Disease in an East Asian Population. J. Clin. Endocrinol. Metab. 2021, 106, e3837–e3851. [Google Scholar] [CrossRef]
- Jhun, M.A.; Mendelson, M.; Wilson, R.; Gondalia, R.; Joehanes, R.; Salfati, E.; Zhao, X.; Braun, K.V.E.; Do, A.N.; Hedman, A.K.; et al. A multi-ethnic epigenome-wide association study of leukocyte DNA methylation and blood lipids. Nat. Commun. 2021, 12, 3987. [Google Scholar] [CrossRef] [PubMed]
- Richard, M.A.; Huan, T.; Ligthart, S.; Gondalia, R.; Jhun, M.A.; Brody, J.A.; Irvin, M.R.; Marioni, R.; Shen, J.; Tsai, P.C.; et al. DNA Methylation Analysis Identifies Loci for Blood Pressure Regulation. Am. J. Hum. Genet. 2017, 101, 888–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, J.; Priego, T.; Pico, C.; Ahrens, W.; De Henauw, S.; Fraterman, A.; Marild, S.; Molnar, D.; Moreno, L.A.; Peplies, J.; et al. Blood cells as a source of transcriptional biomarkers of childhood obesity and its related metabolic alterations: Results of the IDEFICS study. J. Clin. Endocrinol. Metab. 2012, 97, E648–E652. [Google Scholar] [CrossRef] [Green Version]
- Cheshmeh, S.; Nachvak, S.M.; Rezvani, N.; Saber, A. Effects of Breastfeeding and Formula Feeding on the Expression Level of FTO, CPT1A and PPAR-alpha Genes in Healthy Infants. Diabetes Metab. Syndr. Obes. 2020, 13, 2227–2237. [Google Scholar] [CrossRef] [PubMed]
- Obermann-Borst, S.A.; Eilers, P.H.; Tobi, E.W.; de Jong, F.H.; Slagboom, P.E.; Heijmans, B.T.; Steegers-Theunissen, R.P. Duration of breastfeeding and gender are associated with methylation of the LEPTIN gene in very young children. Pediatr. Res. 2013, 74, 344–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paracchini, V.; Pedotti, P.; Taioli, E. Genetics of leptin and obesity: A HuGE review. Am. J. Epidemiol. 2005, 162, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Felix, J.F.; Joubert, B.R.; Baccarelli, A.A.; Sharp, G.C.; Almqvist, C.; Annesi-Maesano, I.; Arshad, H.; Baiz, N.; Bakermans-Kranenburg, M.J.; Bakulski, K.M.; et al. Cohort Profile: Pregnancy And Childhood Epigenetics (PACE) Consortium. Int. J. Epidemiol. 2018, 47, 22–23u. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Children with Methylation Data at the Infancy Timepoint (n = 243) | Children with Methylation Data at Birth, Infancy, and Childhood (n = 50) | |
|---|---|---|
| Demographic Factors | N(%) unless otherwise specified | N(%) unless otherwise specified |
| Maternal age (years) at birth, mean (SD) a | 30.1 (5.2) | 30.1 (4.7) |
| Maternal education | ||
| >12 years | 194 (80.8%) | 42 (84.0%) |
| ≤12 years | ||
| Sex (Female) | ||
| Female | 114 (46.9%) | 25 (50.0%) |
| Male | ||
| Race/Ethnicity | ||
| Non-Hispanic White | 193 (79.4%) | 39 (78.0%) |
| Other | ||
| Perinatal Factors | ||
| Birth weight (g), mean (SD) b | 3336.7 (516.0) | 3380.3 (487.3) |
| Birth Delivery type | ||
| Uncomplicated vaginal | 146 (60.8%) | 35 (70.0%) |
| Complicated vaginal | 36 (15.0%) | 8 (16.0%) |
| Cesarean section | 58 (24.2%) | 7 (14.0%) |
| Gestational Age Category c | ||
| Pre-term | 42 (17.7%) | 9 (18.0%) |
| Term | 173 (72.7%) | 32 (64.0%) |
| Post-term | 23 (9.7%) | 9 (18.0%) |
| Exposure | Age at Introduction | Children with Methylation Data at the Infancy Timepoint (n = 243) | Children with Methylation Data at Birth, Infancy, and Childhood (n = 50) |
|---|---|---|---|
| Exclusive breastfeeding duration in months, mean (SD) a | n.a. | 1.9 (2.0) | 2.0 (2.1) |
| Breastfeeding duration in months, mean (SD) | n.a. | 7.3 (7.2) | 9.0 (9.4) |
| N (%) | N (%) | ||
| Age introduced to gluten-containing cereals (wheat/barley/rye) | <6 months | 90 (37.0%) | 14 (28.0%) |
| ≥6 months | 153 (63.0%) | 36 (72.0%) | |
| Age introduced to non-gluten-containing cereals (rice/oat) | <4 months | 91 (37.5%) | 15 (30.0%) |
| 4–5 months | 129 (53.1%) | 28 (56.0%) | |
| ≥6 months | 23 (9.5%) | 7 (14.0%) | |
| Age introduced to fruit, excluding fruit juice | <4 months | 34 (14.0%) | 3 (6.0%) |
| 4–5 months | 136 (56.0%) | 30 (60.0%) | |
| ≥6 months | 73 (30.0%) | 17 (34.0%) | |
| Age introduced to vegetables | <4 months | 22 (9.1%) | 4 (8.0%) |
| 4–5 months | 142 (58.4%) | 27 (54.0%) | |
| ≥6 months | 79 (32.5%) | 19 (38.0% | |
| Age introduced to meat | <6 months | 34 (14.0%) | 4 (8.0%) |
| ≥6 months | 209 (86.0%) | 46 (92.0%) |
| Position | Nearest Gene | Gene Region | Breastfeeding Duration (Months, Continuous) | Age at Introduction to Meat | ||||
|---|---|---|---|---|---|---|---|---|
| cgID | Chr | Beta Estimate | Nominal p-Value | Beta Estimate | Nominal p-Value | |||
| Significant (FDR < 0.10) CpGs from the Discovery EWAS | ||||||||
| cg00574958 | 11 | 68607622 | CPT1A | 5′UTR | −0.01741 | 8.34 × 10−10 | ||
| cg19693031 | 1 | 145441552 | TXNIP | 3′UTR | −0.01574 | 2.16 × 10−6 | ||
| cg22369607 | 18 | 13821885 | AP001525.1 (miRNA) | −0.01507 | 9.76 × 10−7 | |||
| cg23307264 | 19 | 6424217 | KHSRP | Body | 0.025026 | 1.03 × 10−6 | ||
| cg24092000 | 17 | 80839375 | TBCD | Body | −0.02497 | 1.37 × 10−6 | ||
| cg27173510 | 1 | 230468168 | PGBD5 | Body (in enhancer) | −0.01134 | 2.74 × 10−6 | ||
| Significant (p < 0.05) CpGs of Candidates from the Previous Literature | ||||||||
| cg13381984 | 7 | 127881344 | LEP | 1st exon, 5′UTR | −0.006 | 0.0121 | ||
| cg26814075 | 7 | 127881298 | LEP | TSS200 | −0.005 | 0.0323 | ||
| cg23753947 | 7 | 127889701 | LEP | 5′UTR | <6 m: 0.088 ≥6 m: ref | 0.0482 | ||
| cg00666422 | 7 | 127881440 | LEP | 5′UTR | <6 m: 0.095 ≥6 m: ref | 0.0417 | ||
| Breastfeeding Duration (Months, Continuous) | Age at Introduction to Meat | ||||||
|---|---|---|---|---|---|---|---|
| Birth | Infancy | Childhood | Birth | Infancy | Childhood | ||
| cgID | Nearest Gene | Beta Estimate (p-Value) | Beta Estimate (p-Value) | Beta Estimate (p-Value) | Beta Estimate (p-Value) | Beta Estimate (p-Value) | Beta Estimate (p-Value) |
| Significant CpGs from the Discovery EWAS | |||||||
| cg00574958 | CPT1A | −0.002 (0.6062) | −0.020 (2.35 × 10−6) | −0.015 (0.0011) | |||
| cg19693031 | TXNIP | 0.001 (0.8688) | −0.013 (0.0111) | 0.003 (0.5905) | |||
| cg22369607 | AP001525.1 | −0.029 (0.0028) | −0.037 (9.58 × 10−5) | −0.025 (0.0031) | |||
| cg23307264 | KHSRP | −0.003 (0.7816) | 0.073 (4.88 × 10−5) | −0.010 (0.5385) | |||
| cg24092000 | TBCD | −0.046 (0.0064) | −0.068 (1.18 × 10−5) | −0.064 (0.0001) | |||
| cg27173510 | PGBD5 | −0.023 (0.0010) | −0.027 (0.0002) | −0.020 (0.0051) | |||
| Significant CpGs of Candidates from the Previous Literature | |||||||
| cg13381984 | LEP | −0.005 (0.3665) | −0.003 (0.4189) | −0.003 (0.5692) | |||
| cg26814075 | LEP | −0.009 (0.0732) | −0.006 (0.1424) | −0.003 (0.5622) | |||
| cg23753947 | LEP | <6 m: −0.335 (0.1119) ≥ 6 m: ref | <6 m: 0.065 (0.6018) ≥6 m: ref | <6 m: −0.158 (0.1963) ≥6 m: ref | |||
| cg00666422 | LEP | <6 m: 0.190 (0.2065) ≥6 m: ref | <6 m: 0.248 (0.0685) ≥6 m: ref | <6 m: 0.218 (0.1324) ≥6 m: ref | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walker-Short, E.; Buckner, T.; Vigers, T.; Carry, P.; Vanderlinden, L.A.; Dong, F.; Johnson, R.K.; Yang, I.V.; Kechris, K.; Rewers, M.; et al. Epigenome-Wide Association Study of Infant Feeding and DNA Methylation in Infancy and Childhood in a Population at Increased Risk for Type 1 Diabetes. Nutrients 2021, 13, 4057. https://doi.org/10.3390/nu13114057
Walker-Short E, Buckner T, Vigers T, Carry P, Vanderlinden LA, Dong F, Johnson RK, Yang IV, Kechris K, Rewers M, et al. Epigenome-Wide Association Study of Infant Feeding and DNA Methylation in Infancy and Childhood in a Population at Increased Risk for Type 1 Diabetes. Nutrients. 2021; 13(11):4057. https://doi.org/10.3390/nu13114057
Chicago/Turabian StyleWalker-Short, Elizabeth, Teresa Buckner, Timothy Vigers, Patrick Carry, Lauren A. Vanderlinden, Fran Dong, Randi K. Johnson, Ivana V. Yang, Katerina Kechris, Marian Rewers, and et al. 2021. "Epigenome-Wide Association Study of Infant Feeding and DNA Methylation in Infancy and Childhood in a Population at Increased Risk for Type 1 Diabetes" Nutrients 13, no. 11: 4057. https://doi.org/10.3390/nu13114057
APA StyleWalker-Short, E., Buckner, T., Vigers, T., Carry, P., Vanderlinden, L. A., Dong, F., Johnson, R. K., Yang, I. V., Kechris, K., Rewers, M., & Norris, J. M. (2021). Epigenome-Wide Association Study of Infant Feeding and DNA Methylation in Infancy and Childhood in a Population at Increased Risk for Type 1 Diabetes. Nutrients, 13(11), 4057. https://doi.org/10.3390/nu13114057