
Fasting-Induced Upregulation of MKP-1 Modulates the Hepatic Response to Feeding



Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Animal Studies
2.3. Cell Culture, Nuclear-Cytoplasmic Isolation and Transfections
2.4. RNA Extraction and Real-Time Quantitative PCR Analysis
2.5. Measurement of Fasting Blood Glucose and Hepatic Lipids
2.6. Histological Analysis of Tissue Sections
2.7. Immunoblotting
3. Statistical Analysis
4. Results
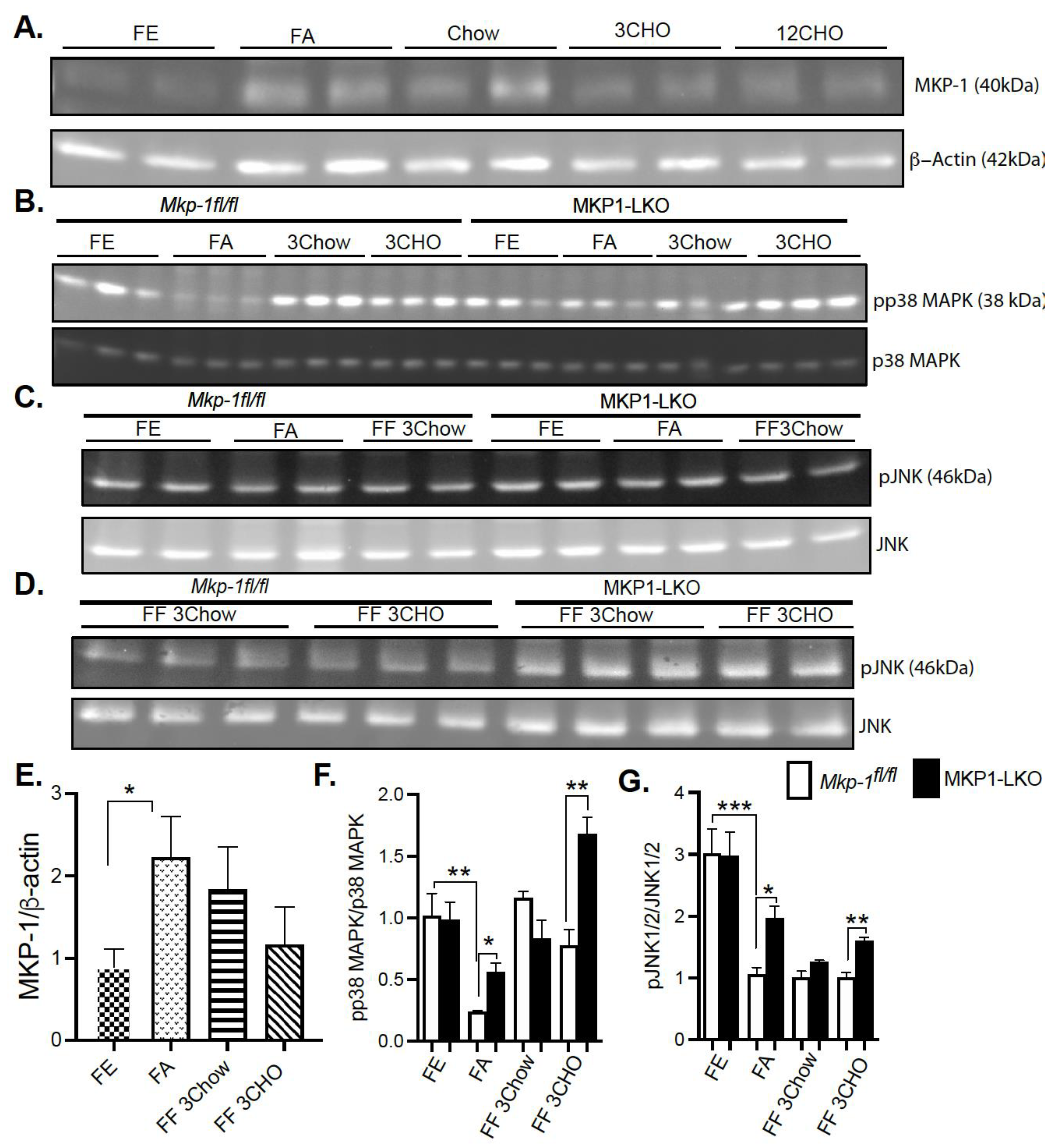
4.1. Fasting Induces MKP-1 Expression in the Liver
4.2. Fasting-Induced Downregulation of p38 MAPK and JNK in the Liver
4.3. Physiologic Parameters of MKP-1-Deficient Liver after Fasting and Refeeding
4.4. Protection from Fasting-Induced Hepatic Steatosis in MKP-1-Deficient Liver
4.5. Regulation of Srebf2 and Its Target Genes during Fasting in MKP-1-Deficient Liver
4.6. Reduced Expression of Hepatic Lipogenic Genes in Fasted and Refed MKP-1-Defcient Liver
4.7. Enhanced Hepatic Inflammatory Response after Fasting and Refeeding a High CHO Diet in MKP-1-Deficient Liver
4.8. p38 MAPK Mediates MKP-1 Regulation of Srebf2
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Arroyo-Johnson, C.; Mincey, K.D. Obesity Epidemiology Worldwide. Gastroenterol. Clin. N. Am. 2016, 45, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, C.; Fisichella, P.M. Epidemiology of Obesity and Associated Comorbidities. J. Laparoendosc. Adv. Surg. Tech. A 2018, 28, 919–924. [Google Scholar] [CrossRef]
- Lawan, A.; Bennett, A.M. Mitogen-Activated Protein Kinase Regulation in Hepatic Metabolism. Trends Endocrinol. Metab. 2017, 28, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Hillgartner, F.B.; Salati, L.M.; Goodridge, A.G. Physiological and molecular mechanisms involved in nutritional regulation of fatty acid synthesis. Physiol. Rev. 1995, 75, 47–76. [Google Scholar] [CrossRef]
- Shimano, H. SREBPs: Physiology and pathophysiology of the SREBP family. FEBS J. 2009, 276, 616–621. [Google Scholar] [CrossRef] [PubMed]
- DeBose-Boyd, R.A.; Ye, J. SREBPs in Lipid Metabolism, Insulin Signaling, and Beyond. Trends Biochem. Sci. 2018, 43, 358–368. [Google Scholar] [CrossRef]
- Johnson, B.M.; DeBose-Boyd, R.A. Underlying mechanisms for sterol-induced ubiquitination and ER-associated degradation of HMG CoA reductase. Semin. Cell Dev. Biol. 2018, 81, 121–128. [Google Scholar] [CrossRef]
- Brown, A.E.; Palsgaard, J.; Borup, R.; Avery, P.; Gunn, D.A.; De Meyts, P.; Yeaman, S.J.; Walker, M. p38 MAPK activation upregulates proinflammatory pathways in skeletal muscle cells from insulin-resistant type 2 diabetic patients. Am. J. Physiol.-Endocrinol. Metab. 2015, 308, E63–E70. [Google Scholar] [CrossRef]
- Bennett, A.M.; Lawan, A. Improving Obesity and Insulin Resistance by Targeting Skeletal Muscle MKP-1. J. Cell. Signal. 2020, 1, 160–168. [Google Scholar]
- Kotzka, J.; Knebel, B.; Haas, J.; Kremer, L.; Jacob, S.; Hartwig, S.; Nitzgen, U.; Muller-Wieland, D. Preventing phosphorylation of sterol regulatory element-binding protein 1a by MAP-kinases protects mice from fatty liver and visceral obesity. PLoS ONE 2012, 7, e32609. [Google Scholar] [CrossRef] [PubMed]
- Lawan, A.; Shi, H.; Gatzke, F.; Bennett, A.M. Diversity and specificity of the mitogen-activated protein kinase phosphatase-1 functions. Cell. Mol. Life Sci. 2013, 70, 223–237. [Google Scholar] [CrossRef] [PubMed]
- DeBose-Boyd, R.A. Significance and regulation of lipid metabolism. Semin. Cell Dev. Biol. 2018, 81, 97. [Google Scholar] [CrossRef]
- Goldstein, I.; Hager, G.L. Transcriptional and Chromatin Regulation during Fasting—The Genomic Era. Trends Endocrinol. Metab. 2015, 26, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; Sarraf, P.; Wright, M.; Yao, K.M.; Mueller, E.; Solanes, G.; Lowell, B.B.; Spiegelman, B.M. Nutritional and insulin regulation of fatty acid synthetase and leptin gene expression through ADD1/SREBP1. J. Clin. Invest. 1998, 101, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, I.; Shimano, H.; Korn, B.S.; Bashmakov, Y.; Horton, J.D. Nuclear sterol regulatory element-binding proteins activate genes responsible for the entire program of unsaturated fatty acid biosynthesis in transgenic mouse liver. J. Biol. Chem. 1998, 273, 35299–35306. [Google Scholar] [CrossRef]
- Weber, L.W.; Boll, M.; Stampfl, A. Maintaining cholesterol homeostasis: Sterol regulatory element-binding proteins. World J. Gastroenterol. 2004, 10, 3081–3087. [Google Scholar] [CrossRef] [PubMed]
- Pawar, A.; Botolin, D.; Mangelsdorf, D.J.; Jump, D.B. The role of liver X receptor-alpha in the fatty acid regulation of hepatic gene expression. J. Biol. Chem. 2003, 278, 40736–40743. [Google Scholar] [CrossRef] [PubMed]
- Linden, A.G.; Li, S.; Choi, H.Y.; Fang, F.; Fukasawa, M.; Uyeda, K.; Hammer, R.E.; Horton, J.D.; Engelking, L.J.; Liang, G. Interplay between ChREBP and SREBP-1c coordinates postprandial glycolysis and lipogenesis in livers of mice. J. Lipid Res. 2018, 59, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Haas, J.T.; Manthena, P.; Wang, Y.; Zhao, E.; Vaitheesvaran, B.; Kurland, I.J.; Biddinger, S.B. Hepatic insulin receptor deficiency impairs the SREBP-2 response to feeding and statins. J. Lipid Res. 2014, 55, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Lawan, A.; Zhang, L.; Gatzke, F.; Min, K.; Jurczak, M.J.; Al-Mutairi, M.; Richter, P.; Camporez, J.P.; Couvillon, A.; Pesta, D.; et al. Hepatic mitogen-activated protein kinase phosphatase 1 selectively regulates glucose metabolism and energy homeostasis. Mol. Cell. Biol. 2015, 35, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Raingeaud, J.; Whitmarsh, A.J.; Barrett, T.; Dérijard, B.; Davis, R.J. MKK3- and MKK6-regulated gene expression is mediated by the p38 mitogen-activated protein kinase signal transduction pathway. Mol. Cell. Biol. 1996, 16, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, S.; Zhou, Y.; Su, W.; Ruan, X.; Wang, B.; Zheng, F.; Warner, M.; Gustafsson, J.; Guan, Y. Ablation of cytochrome P450 omega-hydroxylase 4A14 gene attenuates hepatic steatosis and fibrosis. Proc. Natl. Acad. Sci. USA 2017, 114, 3181–3185. [Google Scholar] [CrossRef] [PubMed]
- Ajabnoor, G.M.; Bahijri, S.; Shaik, N.A.; Borai, A.; Alamoudi, A.A.; Al-Aama, J.Y.; Chrousos, G.P. Ramadan fasting in Saudi Arabia is associated with altered expression of CLOCK, DUSP and IL-1alpha genes, as well as changes in cardiometabolic risk factors. PLoS ONE 2017, 12, e0174342. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, Y.; Ueyama, E.; Senba, E. Fasting-induced activation of mitogen-activated protein kinases (ERK/p38) in the mouse hypothalamus. J. Neuroendocrinol. 2004, 16, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Nishio, H.; Kuwabara, H.; Mori, H.; Suzuki, K. Repeated fasting stress causes activation of mitogen-activated protein kinases (ERK/JNK) in rat liver. Hepatology 2002, 36, 72–80. [Google Scholar] [CrossRef]
- Lee, C.; Raffaghello, L.; Brandhorst, S.; Safdie, F.M.; Bianchi, G.; Martin-Montalvo, A.; Pistoia, V.; Wei, M.; Hwang, S.; Merlino, A.; et al. Fasting cycles retard growth of tumors and sensitize a range of cancer cell types to chemotherapy. Sci. Transl. Med. 2012, 4, 124ra27. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.W.; Adams, G.B.; Perin, L.; Wei, M.; Zhou, X.; Lam, B.S.; Da Sacco, S.; Mirisola, M.; Quinn, D.I.; Dorff, T.B.; et al. Prolonged fasting reduces IGF-1/PKA to promote hematopoietic-stem-cell-based regeneration and reverse immunosuppression. Cell Stem Cell 2014, 14, 810–823. [Google Scholar] [CrossRef] [PubMed]
- Gross, S.; Rahal, R.; Stransky, N.; Lengauer, C.; Hoeflich, K.P. Targeting cancer with kinase inhibitors. J. Clin. Investig. 2015, 125, 1780–1789. [Google Scholar] [CrossRef] [PubMed]
- Caffa, I.; Longo, V.D.; Nencioni, A. Fasting plus tyrosine kinase inhibitors in cancer. Aging (Albany NY) 2015, 7, 1026–1027. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moslehi, A.; Hamidi-Zad, Z. Role of SREBPs in Liver Diseases: A Mini-review. J. Clin. Transl. Hepatol. 2018, 6, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Purves, T.; Middlemas, A.; Agthong, S.; Jude, E.B.; Boulton, A.J.; Fernyhough, P.; Tomlinson, D.R. A role for mitogen-activated protein kinases in the etiology of diabetic neuropathy. FASEB J. 2001, 15, 2508–2514. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | kcal./g | g/kg | kcal./kg |
|---|---|---|---|
| Casein | 3.58 | 140 | 501.2 |
| L-Cystine | 4 | 1.8 | 7.2 |
| Sucrose | 4 | 100 | 400 |
| Cornstarch | 3.6 | 465.692 | 1676.4912 |
| Dyetrose | 3.8 | 155 | 589 |
| Soybean Oil | 9 | 40 | 360 |
| t-Butylhydroquinone | 0.008 | 0 | |
| Cellulose | 0 | 50 | 0 |
| Mineral Mix | 210,050 0.84 | 35 | 29.4 |
| Vitamin Mix | 310,025 3.87 | 10 | 38.7 |
| Choline Bitartrate | 2.5 | 0 | |
| 1000 | 3601.9912 |
| Ingredient | kcal./g | g/kg | kcal./kg |
|---|---|---|---|
| Casein | 3.58 | 200 | 716 |
| L-Cystine | 4 | 3 | 12 |
| Sucrose | 4 | 350 | 1400 |
| Cornstarch | 3.6 | 315 | 1134 |
| Dyetrose | 3.8 | 35 | 133 |
| Soybean Oil | 9 | 25 | 225 |
| t-Butylhydroquinone | 0 | 0.005 | 0 |
| Lard | 9 | 20 | 180 |
| Cellulose | 0 | 50 | 0 |
| Mineral Mix #210088 | 1.6 | 10 | 16 |
| Dicalcium Phosphate | 0 | 13 | 0 |
| Calcium Carbonate | 0 | 5.5 | 0 |
| Potassium Citrate H2O | 0 | 16.5 | 0 |
| Vitamin Mix # 300050 | 3.92 | 10 | 39.2 |
| Choline Bitartrate | 0 | 2 | 0 |
| 1055.005 | 3855.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sellers, J.; Brooks, A.; Fernando, S.; Westenberger, G.; Junkins, S.; Smith, S.; Min, K.; Lawan, A. Fasting-Induced Upregulation of MKP-1 Modulates the Hepatic Response to Feeding. Nutrients 2021, 13, 3941. https://doi.org/10.3390/nu13113941
Sellers J, Brooks A, Fernando S, Westenberger G, Junkins S, Smith S, Min K, Lawan A. Fasting-Induced Upregulation of MKP-1 Modulates the Hepatic Response to Feeding. Nutrients. 2021; 13(11):3941. https://doi.org/10.3390/nu13113941
Chicago/Turabian StyleSellers, Jacob, Abigail Brooks, Savanie Fernando, Gabrielle Westenberger, Sadie Junkins, Shauri Smith, Kisuk Min, and Ahmed Lawan. 2021. "Fasting-Induced Upregulation of MKP-1 Modulates the Hepatic Response to Feeding" Nutrients 13, no. 11: 3941. https://doi.org/10.3390/nu13113941
APA StyleSellers, J., Brooks, A., Fernando, S., Westenberger, G., Junkins, S., Smith, S., Min, K., & Lawan, A. (2021). Fasting-Induced Upregulation of MKP-1 Modulates the Hepatic Response to Feeding. Nutrients, 13(11), 3941. https://doi.org/10.3390/nu13113941