
The Influence of Plants from the Alliaceae Family on Morphological Parameters of the Intestine in Atherogenic Rats


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.1.1. The Content of Bioactive Ingredients in Plants from the Alliaceae Family
Extracts Preparation
Total Polyphenols Content
Flavonoids Content
Flavanols Content
Anthocyanins Content
2.2. Animals
2.2.1. Experimental Design
2.2.2. Animal Sacrifice
2.2.3. Sampling and Measurements
Basic Experimental Data
Biochemical Analysis
The Antioxidant Potential of Plasma
Tissue Sampling Procedure for the Ileum Histology
Light Microscope Examination
2.3. Statistical Analysis
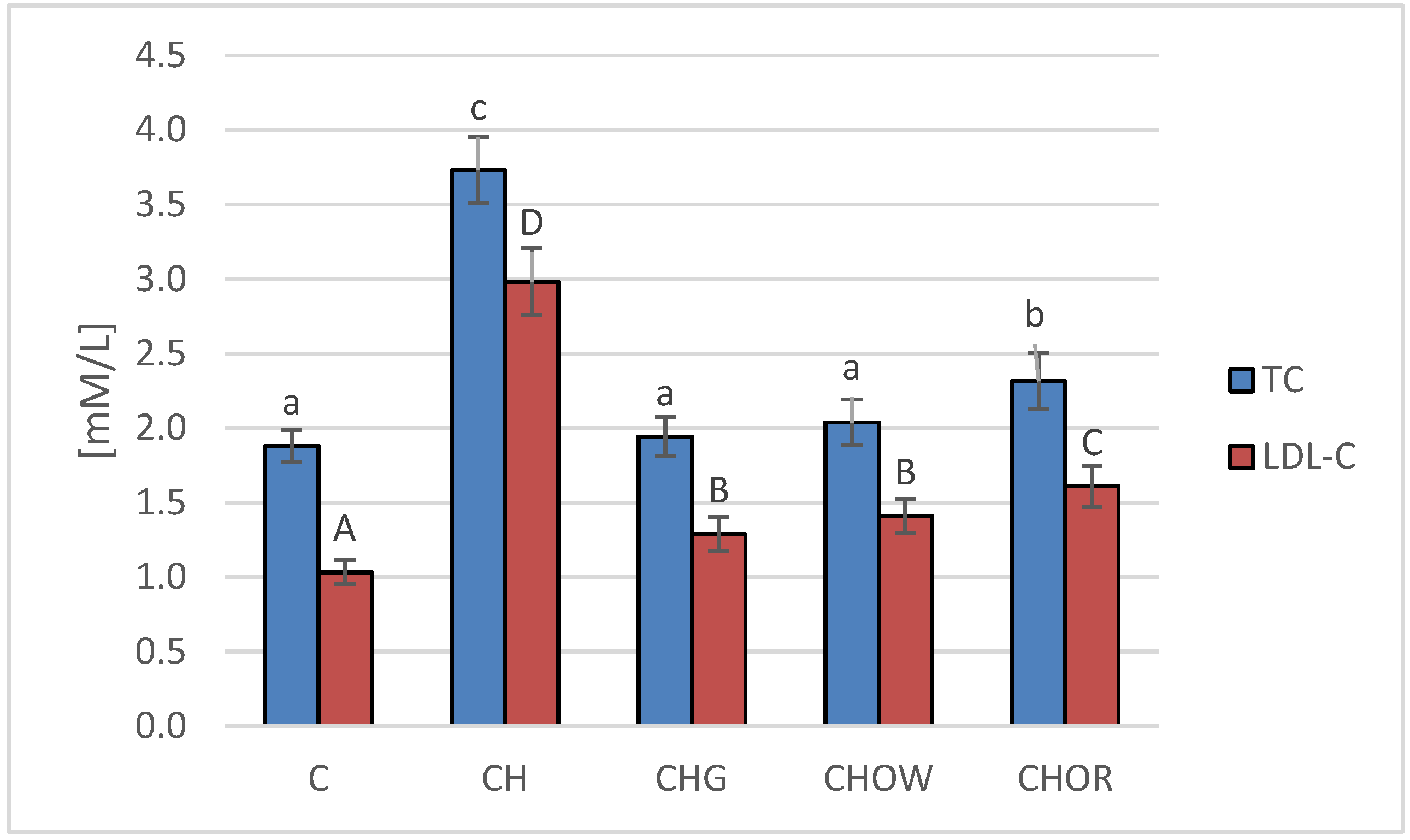
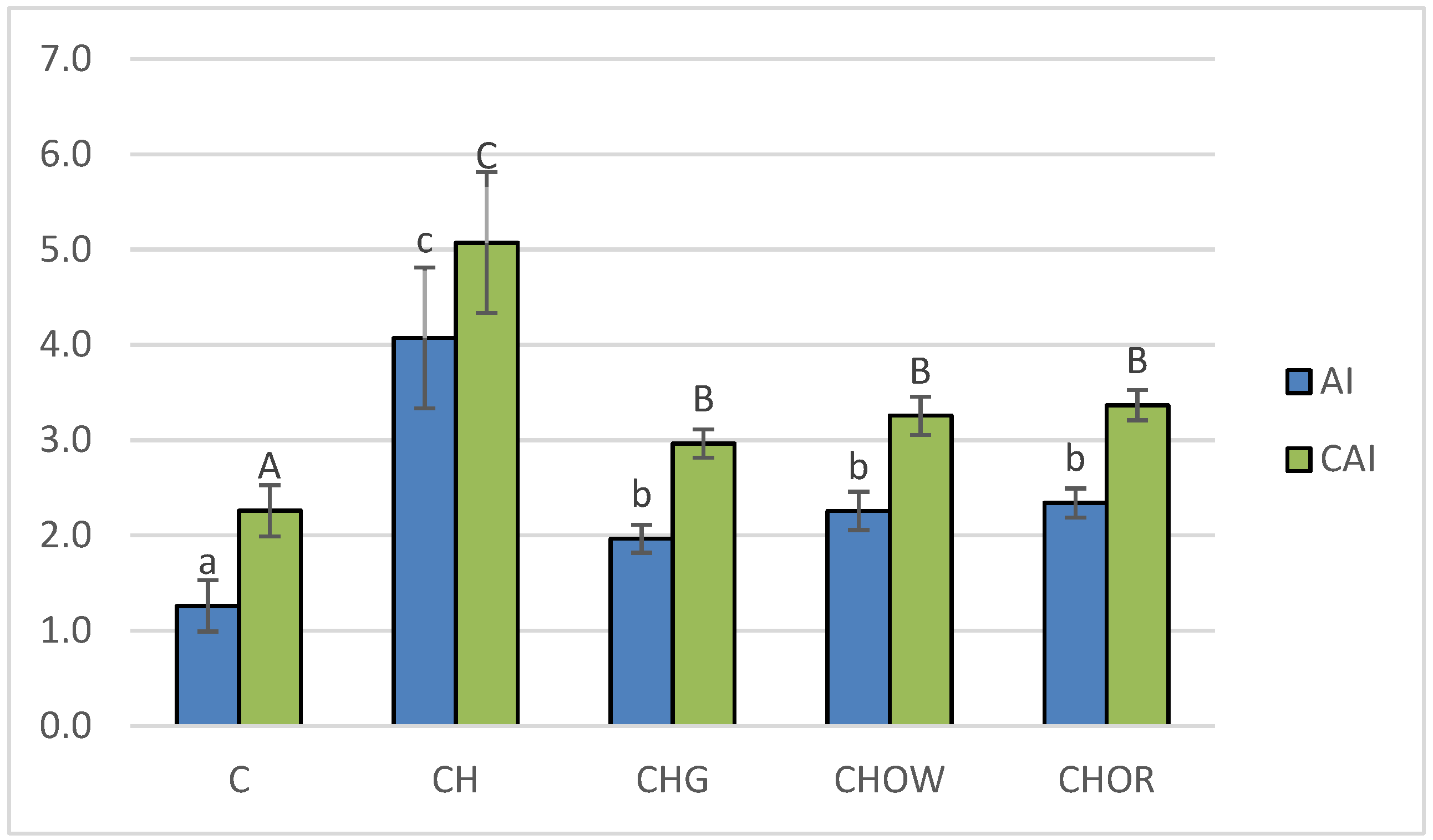
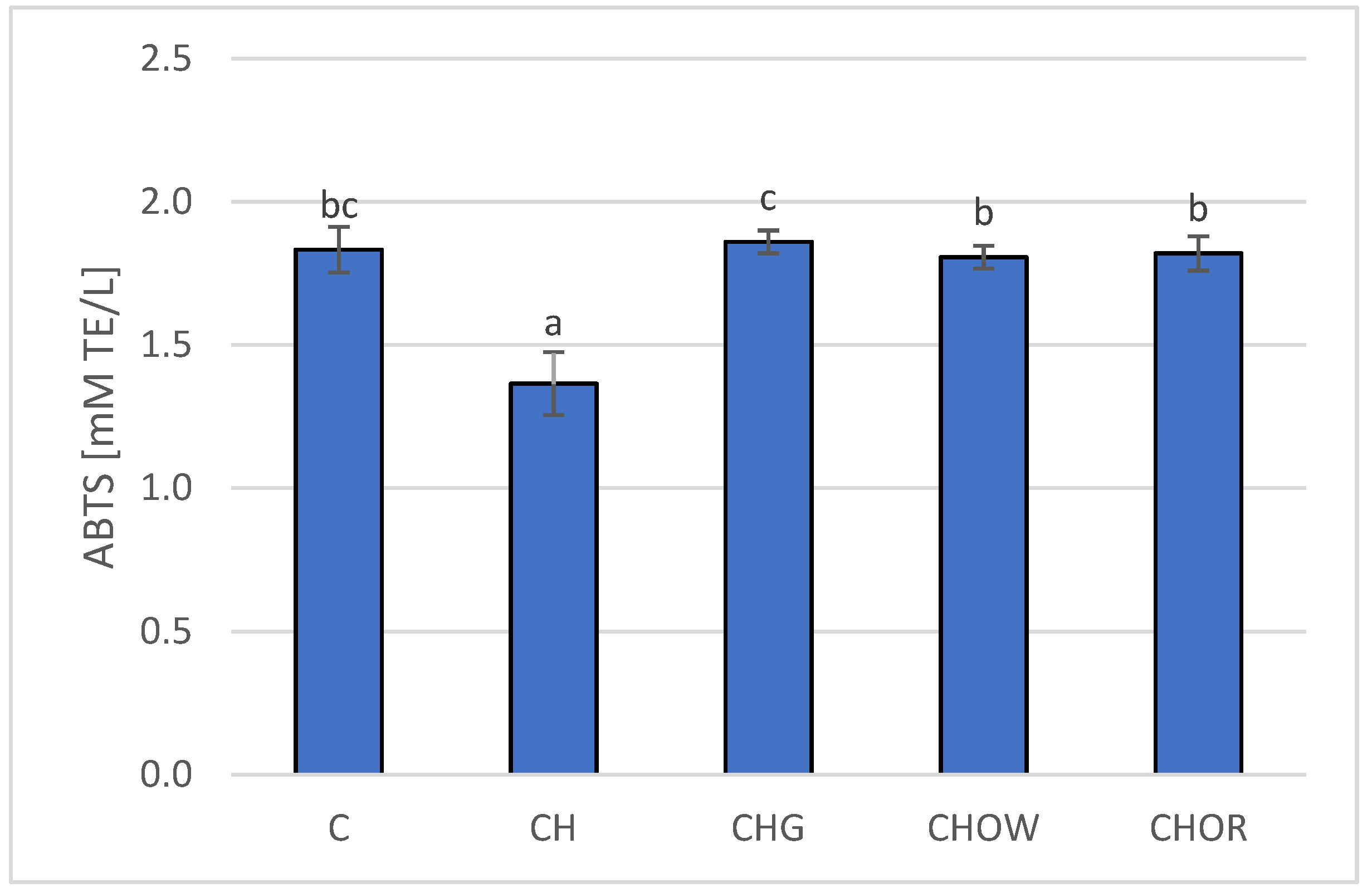
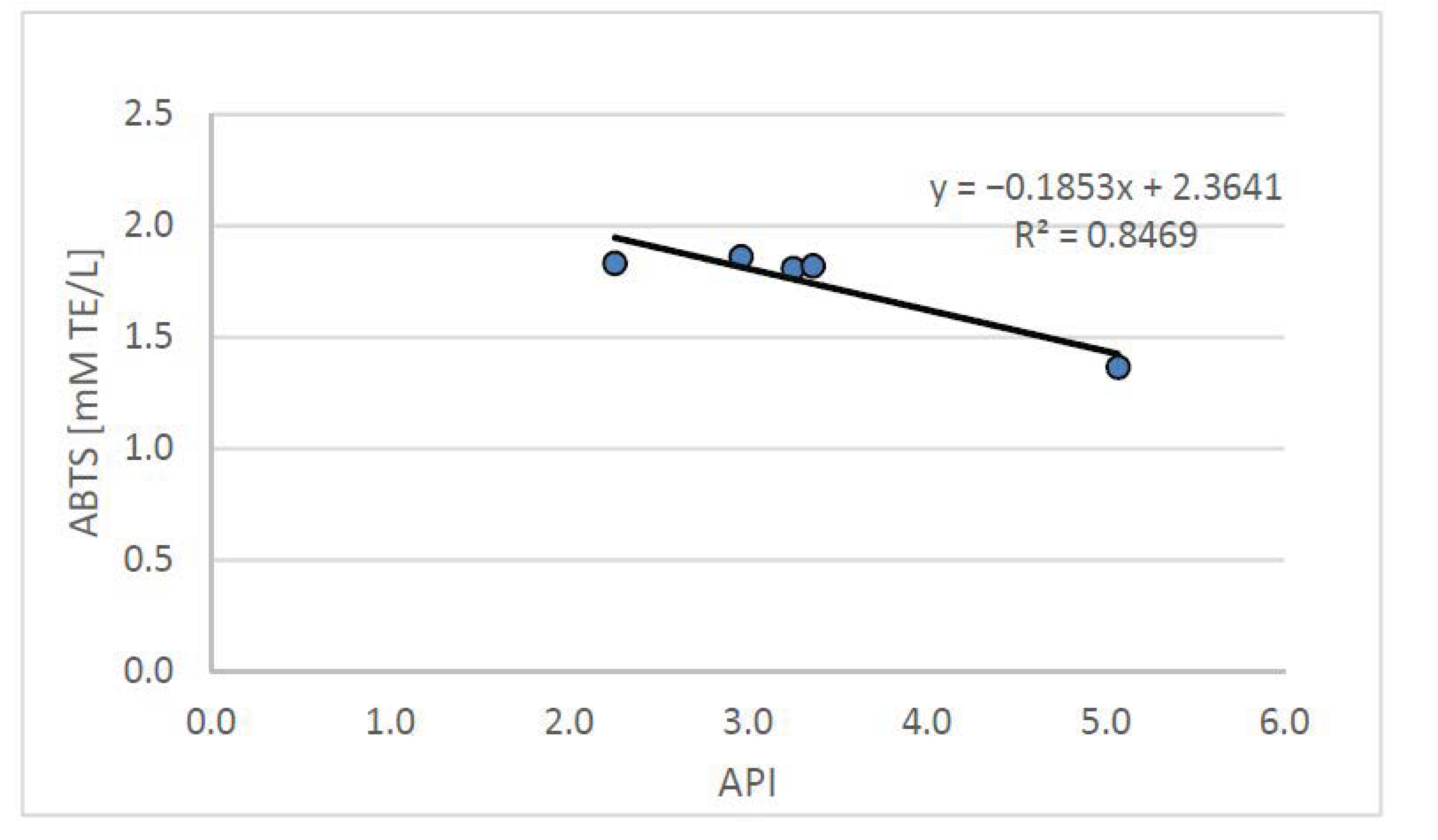
3. Results and Discussion
3.1. Plant Material
3.2. Animal Study
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barzegar, M.; Rajabi, A.; Hassandokht, M.R.; Jabbari, A. Chemical composition of different cultivars of onion (Allium cepa L.) produced in Iran. Hortic. Environ. Biotechnol. 2008, 49, 121–127. [Google Scholar]
- Butt, M.S.; Sultan, M.T.; Iqbal, J. Garlic: Nature’s protection against physiological threats. Crit. Rev. Food Sci. Nutr. 2009, 49, 538–551. [Google Scholar] [CrossRef]
- Griffiths, G.; Trueman, L.; Crowther, T.E.; Thomas, B.; Smith, B. Onions—A global benefit to health. Phytother. Res 2002, 16, 819–830. [Google Scholar] [CrossRef]
- Gorinstein, S.; Drzewiecki, J.; Leontowicz, H.; Leontowicz, M.; Najman, K.; Jastrzębski, Z.; Zachwieja, Z.; Barton, H.; Shtabsky, B.; Katrich, E.; et al. Comparison of the bioactive compounds and antioxidant potentials of fresh and cooked Polish, Ukrainian and Israeli gralic. J. Agric. Food Chem. 2005, 53, 2726–2732. [Google Scholar] [CrossRef]
- Nemeth, K.; Piskula, M.K. Food content, processing, absorption and metabolism of onion flavonoids. Crit. Rev. Food Sci. Nutr. 2007, 47, 397–409. [Google Scholar] [CrossRef]
- Gorinstein, S.; Leontowicz, H.; Leontowicz, M.; Namiesnik, J.; Najman, K.; Drzewiecki, J.; Cvikrova, M.; Martincova, O.; Katrich, E.; Trakhtenberg, S. Comparison of the main bioactive compounds and antioxidant activities in garlic and white and red onions after treatment protocols. J. Agric. Food Chem. 2008, 56, 4418–4426. [Google Scholar] [CrossRef]
- Gorinstein, S.; Jastrzębski, Z.; Leontowicz, H.; Leontowicz, M.; Namiesnik, J.; Najman, K.; Park, Y.S.; Heo, B.G.; Cho, J.Y.; Bae, J.H. Comparative control of the bioactivity of some frequently consumed vegetables subjected to different processing conditions. Food Control 2009, 20, 407–413. [Google Scholar] [CrossRef]
- Benítez, V.; Mollá, E.; Martín-Cabrejas, M.A.; Aguilera, Y.; López-Andréu, F.J.; Cools, K.; Terry, L.A.; Esteban, M.R. Characterization of industrial onion wastes (Allium cepa L.): Dietary fibre and bioactive compound. Plant Foods Hum. Nutr. 2011, 66, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Bayan, L.; Koulivand, P.H.; Gorji, A. Garlic: A review of potential therapeutic effects. Avicenna J. Phytomedicine 2014, 4, 1–14. [Google Scholar]
- Nile, S.H.; Park, S.W. Total phenolics, antioxidant and xanthine oxidase inhibitory activity of three colored onions (Allium cepa L.). Front. Life Sci. 2014, 7, 224–228. [Google Scholar] [CrossRef]
- Thangasamy, A.; Khade, Y.; Major Singh, M. Nutrient management in onion and garlic—Review. J. Allium Res. 2018, 1, 107–115. [Google Scholar]
- Genkinger, J.M.; Platz, E.A.; Hoffman, S.C.; Comstock, G.W.; Helzsouer, K.J. Fruit, vegetable and antioxidant intake and all-cause, cancer, and cardiovascular disease mortality in a community-dwelling population in Washington County, Maryland. Am. J. Epidemiol. 2004, 160, 1223–1233. [Google Scholar] [CrossRef]
- Banerjee, S.K.; Maulik, S.K. Effect of garlic on cardiovascular disorders: A review. Nutr. J. 2002, 19, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Mnayer, D.G.; Tabanelli, G.; Stojanović-Radić, Z.Z.; Sharifi-Rad, M.; Yousaf, Z.; Vallone, L.; Setzer, W.N.; Iriti, M. Plants of the genus Allium as antibacterial agents: From tradition to pharmacy. Cell. Mol. Biol. 2016, 62, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, C.S.; Sekhar, C.C. A review on integrated nutrient management in bulbous vegetable crops. J. Pharmacogn. Phyto. Chem. 2018, 7, 411–415. [Google Scholar]
- Izotti, A.; Di Marco, B.; De Floras, S.; Sacca, S. The role of oxidative stress oxidative stress. Mutat. Res. 2006, 612, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Katsuda, S.; Kaji, T. Atherosclerosis and extracellular matrix. J. Atheroscler. Thromb. 2003, 10, 267–274. [Google Scholar] [CrossRef]
- Gorinstein, S.; Leontowicz, M.; Leontowicz, H.; Najman, K.; Namiesnik, J.; Park, Y.S.; Jung, S.T.; Kang, S.G.; Trakhtenberg, S. Supplementation of garlic lowers lipids and increases antioxidant capacity in plasma of rats. Nutr. Res. 2006, 26, 362–368. [Google Scholar] [CrossRef]
- Gorinstein, S.; Drzewiecki, J.; Leontowicz, H.; Leontowicz, M.; Najman, K.; Katrich, E.; Barasch, D.; Yamamoto, K.; Trakhtenberg, S. Raw and boiled garlic enhances plasma antioxidant activity and improves plasma lipid metabolism in cholesterol—Fed rats. Life Sci. 2006, 78, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, R. Beneficial effect of Allium sativum and Allium tuberosum on experimental hyperlipidemia and atherosclerosis. Pak. J Physiol. 2008, 4, 7–9. [Google Scholar]
- Iciek, M.; Kwiecień, I.; Włodek, L. Biological properties of garlic-derived organosulfur compounds. Environ. Mol. 2009, 50, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Gorinstein, S.; Leontowicz, H.; Leontowicz, M.; Jastrzębski, Z.; Najman, K.; Tashma, Z.; Katrich, E.; Heo, B.G.; Cho, J.Y.; Park, Y.J.; et al. The Influence of raw and processed garlic and onions on plasma classical and non-classical atherosclerosis indices: Investigations in vitro and in vivo. Phytother. Res. 2010, 24, 706–714. [Google Scholar] [CrossRef]
- Gorinstein, S.; Leontowicz, H.; Leontowicz, M.; Najman, K.; Bielecki, W.; Ham, K.S.; Kang, S.G.; Paredes-Lopez, O.; Martinez-Ayala, A.L.; Trakhtenberg, S. Aorta and liver changes in rats fed cholesterol-containing and raw vegetables-supplemented diets: Experiments in vitro and in vivo. J. Agric. Food Chem. 2011, 59, 7441–7451. [Google Scholar] [CrossRef]
- El-Sabban, F. Garlic as an antiplatelet aggregation agent. J. Chin. Clin. Med. 2009, 4, 288–294. [Google Scholar]
- Qi, R.; Liao, F.; Inouke, K.; Yatomi, Y.; Sato, K.; Ozaki, Y. Inhibition by diallyl trisulfide, a garlic component, of intacellular Ca2+ mobilization without affecting inositol-1,4,5-triphosphate (IP3) formation in activated platelets. Biochem. Pharmacol. 2000, 60, 1475–1480. [Google Scholar] [CrossRef]
- Liu, L.; Yeh, Y.Y. Water-soluble organosulfur compounds of garlic inhibit fatty acid and triglycerides synthesis in cultured rat hepatocytes. Lipids 2001, 36, 395–400. [Google Scholar] [CrossRef]
- Yeh, Y.Y.; Liu, L. Cholesterol-lowering effects of garlic extracts and organosulfur compounds: Human and animal studies. J Nutr. 2001, 131, 989S–993S. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yeh, Y.Y. S-alk(en)yl cysteines of garlic inhibit cholesterol synthesis by deactivating HMG-CoA reductase in cultured rat hepatocytes. J. Nutr. 2002, 132, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Augusti, K.T.; Chackery, J.; Jacob, J.; Kuriakose, S.; George, S.; Nair, S.S. Beneficial effects of a polar fraction of garlic (Allium sativum L.) oil in rats fed with two different high fat diets. Indian J. Exp. Biol. 2005, 43, 76–83. [Google Scholar] [PubMed]
- Rose, P.; Whiteman, M.; Moore, P.K.; Zhu, Y.Z. Bioactive S-alk(en)yl cysteine sulfoxide metabolites in the genus Allium: The chemistry of potential therapeutic agents. Nat. Prod. Rep. 2005, 22, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Arnault, I.; Auger, J. Seleno-compounds in garlic and onion. J. Chromatogr. A 2006, 1112, 23–30. [Google Scholar] [CrossRef]
- Chung, L.Y. The antioxidant properties of garlic compounds: Allyl cysteine, alliin, allicin, and allyl disulfide. J. Med. Food 2006, 9, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Jastrzębski, Z.; Leontowicz, H.; Leontowicz, M.; Namiesnik, J.; Zachwieja, Z.; Barton, H. The bioactivity of processed garlic (Allium sativum L.) as shown in vitro and in vivo studies on rats. Food Chem. Toxicol. 2007, 45, 1626–1633. [Google Scholar] [CrossRef]
- Pedraza-Chaverri, J.; Medina-Campos, O.N.; Segoviano-Murillo, S. Effect of heating on peroxynitrite scavenging capacity of garlic. Food Chem. Toxicol. 2007, 45, 622–627. [Google Scholar] [CrossRef]
- Bozin, B.; Dukic, N.M.; Samojlik, I.; Goran, A.; Igic, R. Phenolics as antioxidants in garlic (Allium sativum L.). Food Chem. 2008, 111, 925–929. [Google Scholar] [CrossRef]
- Chowdhury, R.; Dutta, A.; Chaudhuri, S.R.; Sharma, N.; Giri, A.K.; Chaudhuri, K. In vitro and in vivo reduction of sodium arsenite induced toxicity by aqueous garlic extracts. Food Chem. Toxicol. 2008, 46, 740–751. [Google Scholar] [CrossRef]
- Chen, S.; Shen, X.; Cheng, S.; Li, P.; Du, J.; Chang, Y.; Meng, H. Evaluation of garlic cultivars for polyphenolic content and antioxidant properties. PLoS ONE 2013, 8, e79730. [Google Scholar] [CrossRef] [PubMed]
- Santhosha, S.G.; Prakash, J.; Prabhavathi, S.N. Bioactive components of garlic and their physiological role in health maintenance: A review. Food Biosci. 2013, 3, 59–74. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Rybczyńska-Tkaczyk, K.; Gaweł-Bęben, K.; Świeca, M.; Karaś, M.; Jakubczyk, A.; Matysiak, M.; Binduga, U.E.; Gmiński, J. Characterization of active compounds of different garlic (Allium sativum L.) cultivars. Polish J. Food Nutr. Sci. 2018, 68, 73–81. [Google Scholar] [CrossRef]
- Ramirez, D.A.; Locatelli, D.A.; Gonzálezc, R.E.; Cavagnaro, P.F.; Alejandra, B.; Camargo, A.B. Analytical methods for bioactive sulfur compounds in Allium: An integrated review and future directions. J. Food Compost. Anal. 2017, 61, 4–19. [Google Scholar] [CrossRef]
- Ryu, J.H.; Kang, D. Physicochemical properties, biological activity, health benefits, and general limitations of aged black garlic: A review. Molecules 2017, 22, 919. [Google Scholar] [CrossRef]
- Najman, K.; Sadowska, A.; Hallmann, E. Influence of thermal processing on the bioactive, antioxidant, and physicochemical properties of conventional and organic agriculture black garlic (Allium sativum L.). Appl. Sci. 2020, 10, 8638. [Google Scholar] [CrossRef]
- Najman, K.; Sadowska, A.; Hallmann, E. Evaluation of bioactive and physicochemical properties of white and black garlic (Allium sativum L.) from conventional and organic cultivation. Appl. Sci. 2021, 11, 874. [Google Scholar] [CrossRef]
- Elmahdi, B.; Maha, M.K.; Afaf, I.K.A. The effect of fresh crushed garlic bulbs (Allium sativum) on plasma lipids in hypercholesterolemic rats. Am. J. Anim. Vet. 2008, 3, 15–19. [Google Scholar]
- Dillon, S.A.; Burmi, R.S.; Lowe, G.M.; Billington, D.; Rahman, K. Antioxidant properties of aged garlic extract: An in vitro study incorporating human low density lipoprotein. Life Sci. 2003, 72, 1583–1994. [Google Scholar] [CrossRef]
- Lau, B.H.S. Suppression of LDL oxidation by garlic compounds is a possible mechanism of cardiovascular health benefit. J Nutr. 2006, 136, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Bordia, T.; Mustafa, T. Effect of raw versus boiled aqueous extract of garlic and onion on platelet aggregation. Prostaglandins Leukot. Essent. Fatty Acids 1999, 60, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Thomson, M.; Afzal, M. Garlic and onions: Their effect on eicosanoid metabolism and its clinical relevance. Prostaglandins Leukot. Essent. Fatty Acids 2000, 62, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.A.; Demmig-Adams, B. Antioxidant, anti-inflammatory, and antimicrobial properties of garlic and onions. Nutr. Food Sci. 2007, 37, 178–183. [Google Scholar] [CrossRef]
- Osmont, K.S.; Arnt, C.R.; Goldman, I.L. Temporal aspects of onion-induced antiplatelet activity. Plant Foods Hum. Nutr. 2003, 58, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Im, M.H.; Park, Y.S.; Leontowicz, H.; Leontowicz, M.; Namiesnik, J.; Ham, K.S.; Kang, S.G.; Najman, K.; Gorinstein, S. The thermostability, bioactive compounds and antioxidant activity of some vegetables subjected to different duration of boiling: Investigation in vito. LWT—Food Sci. Technol. 2011, 44, 92–99. [Google Scholar] [CrossRef]
- Im, M.H.; Park, Y.S.; Ham, K.S.; Kang, S.G.; Leontowicz, H.; Leontowicz, M.; Namiesnik, J.; Najman, K.; Gorinstein, S. Effects of cooking on the bioactivity of lotus roots and white onions. Int. J. Food Prop. 2012, 15, 49–59. [Google Scholar] [CrossRef]
- Gabler, N.K.; Osrowska, E. Dietary onion intake as part of a typical high fat diet improves indices of cardiovascular health using the mixed pig model. Plant Foods Hum. Nutr. 2003, 61, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Gabler, N.K.; Ostrowska, E.; Sterling, S.J.; Jones, R.B.; Tatham, B.G.; Eagling, D.R.; Jois, M.; Dunshea, F.R. Consumption of raw brown onions variably modulate plasma lipid profile and lipoprotein oxidation in pigs fed a high-fat diet. J. Sci. Food Agric. 2005, 85, 154–160. [Google Scholar] [CrossRef]
- Glasser, G.; Graefe, E.U.; Struck, F.; Veit, M.; Gebhardt, R. Comparison of antioxidative capacities and inhibitory effects of cholesterol biosynthesis of quercetin and potential metabolites. Phytomedicine 2002, 9, 22–40. [Google Scholar] [CrossRef] [PubMed]
- Ostrowska, E.; Gabler, N.K.; Sterling, S.J.; Tatham, B.G.; Jones, R.B.; Eagling, D.R.; Jois, M.; Dunshea, F.R. Consumption of brown onions (Allium cepa var Cavalier and var Destiny) moderately modulates blond lipids, haematological and haemostatic variables in health pigs. Br. J. Nutr. 2004, 91, 211–218. [Google Scholar] [CrossRef]
- Hubbard, G.P.; Wolffram, S.; Gibbns, J.M.; Lovegrove, J.A. Ingestion of quercetin inhibits platelet aggregation and essential components of the collagen-stimulated platelet activation pathway in man: A pilot study. J. Thromb. 2006, 2, 2138–2145. [Google Scholar] [CrossRef]
- Moon, J.H.; Nakata, R.; Oshima, S.; Inakuma, T.; Terao, J. Accumulation of quercetin conjugates in blood plasma after the short-term ingestion of onion by women. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, 461–467. [Google Scholar] [CrossRef]
- Olaiya, O.G.; Ailenosi, S.S.; Adelaja, A.; Eniola, K. Effects of aqueous extracts of garlic and vitamin C on the kidney of albino rats. Asian J. Exp. Biol. Sci. 2011, 2, 455–461. [Google Scholar]
- Hoshino, T.; Kashimoto, N.; Kasuga, S. Effects of garlic preparations on the gastrointestinal mucosa. J. Nutr. 2001, 131, 1109–1113. [Google Scholar] [CrossRef]
- Sood, D.R.; Vinod, C.; Shilpa. Effect of garlic (Allium sativum L.) extract on degree of hydration, fructose, sulphur and phosphorus contents of eye lens and intestinal absorption of nutrients. Indian J. Clin. Biochem. 2003, 18, 190–196. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gupta, A.; Sandhu, S. Effect of ASA110 on jejunal enzymes and nucleic acids and morphological changes in jejunum. J. Nutr. 1998, 18, 841–850. [Google Scholar]
- Brenner, S.; Wolf, R. Possible nutritional factors in induced pemphigus. Dermatology 1994, 189, 337–339. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.K.; Maulik, M.; Manchandra, S.C.; Dinda, A.K.; Das, T.K.; Maulik, S.K. Garlic-induced alteration in rat liver and Sidney morphology and associated changes in endogenous antioxidant status. Food Chem. Toxicol. 2001, 39, 793–797. [Google Scholar] [CrossRef]
- Joseph, P.K.; Ramesha, R.; Sundaresh, C.S. Toxic effects of garlic extract and garlic oil in rats. Indian J. Exp. Biol. 1989, 27, 977–979. [Google Scholar] [PubMed]
- Nakagawa, S.; Masamoto, K.; Sumiyoshi, H.; Kunihiro, K.; Fuwa, T. Effect of raw and extracted aged garlic on growth of young rats and their organ after peroral administration. Toxicology 1980, 5, 91–112. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Awazu, S.; Itakura, Y.; Fuwa, T. Alleviation by garlic of antitumor drug-induced damage to the intestine. J. Nutr. 2001, 131, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Insani, E.M.; Mignaqui, A.C.; Salomón, V.M.; Pazos, A.A.; Pighin, D.G.; Sucar, S.; Roux, M.E.; Slobodianik, N.H. Cellular immune response in intestinal villi of rats after consumption of onion (Allium cepa L.) or quercetin. Proc. Nutr. Soc. 2010, 69, E284. [Google Scholar] [CrossRef]
- Mabrouk, M.A.; Nnawodu, F.I.; Tanko, Y.; Dawud, F.; Mohammed, A. Effect of aqueous garlic (Ag) extract on aspirin induces gastric musocal lesion in albino Wistar rats. Curr. Res. J. Biol. Sci. 2009, 1, 15–19. [Google Scholar]
- Omotoso, G.O.; Muonagolu, J.; Enaibe, B.U. Histological evaluation of the jejunum and ileum of rats after administration of high dose garlic aqueous exctract. Int. J. Health Sci. 2012, 6, 111–116. [Google Scholar] [CrossRef]
- David, A.V.A.; Satyanarayana, N.; Parasuraman, S.; Bharathi, S.; Arulmoli, R. Ameliorative effect of quercetin on methotrexate induced toxicity in Sprague-Dawley rats: A histopathological study. Indian J. Pharm. Educ. 2016, 50, 200–2008. [Google Scholar] [CrossRef]
- De Wit, N.J.W.; Hulst, M.; Govers, C.; van der Meulen, J.; van Hoef, A.; Stoopen, G.; Hamers, A.; Hoekman, A.; de Vos, R.; Bovee, T.F.H.; et al. Effects of digested onion extracts on intestinal gene expression: An interspecies comparison using different intestine models. PLoS ONE 2016, 11, 1–18. [Google Scholar] [CrossRef]
- Pişkin, Ö.; Baş, Y.; Karakaya, K.; Can, M.; Elmas, Ö.; Büyükuysal, M.Ç. Protective effects of quercetin on intestinal damage caused by ionizing radiation. Med. Bull. Haseki 2016, 54, 1–7. [Google Scholar] [CrossRef]
- Riad, N.H.A.; Fares, N.H.; Mostafa, O.M.S.; Mahmoud, Y.I. The effect of garlic on murine Schistosomiasis Mansoni: A histological and ultrastructural study on the ileum. Res. J. Med. Sci. 2008, 3, 188–201. [Google Scholar]
- Tatara, M.R.; Śliwa, E.; Dudek, K.; Mosiewicz, J.; Studziński, T. Effect of aged garlic extract and allicin administration to sows during pregnancy and lactation on body weight gain and gastrointestinal tract development of piglets. Part 1. Bull. Vet. Inst. Pulawy 2005, 49, 349–355. [Google Scholar]
- Arija, I.; Viveros, A.; Brenes, A.; Canales, R.; Pizarro, M.; Castaño, M. Histological alterations in the intestinal epithelium caused by the inclusion of full-fat sunflower kernels in broiler chicken diets. Poult 2000, 79, 1332–1334. [Google Scholar] [CrossRef] [PubMed]
- Jamroz, D.; Wertelecki, T.; Houszka, M.; Kamel, C. Influence of diet type on the inclusion of plant origin active substances on morphological and histochemical characteristics of the stomach and jejunum walls in chicken. J. Anim. Physiol. Anim. Nutr. 2006, 90, 255–268. [Google Scholar] [CrossRef]
- Incharoen, T.; Yamauchi, K.; Erikawa, T.; Gotoh, H. Histology of intestinal villi and epithelial cells in chickens fed low-crude protein or low-crude fat diets. Ital. J. Anim. Sci. 2010, 9, 428–434. [Google Scholar] [CrossRef]
- Kaya, H.; Kaya, A.; Gül, M.; Çelebi, S.; Timurkaan, S.; Apaydın, B. Effects of supplementation of different levels of organic acids mixture to the diet on performance, egg quality parameters, serum traits and histological criteria of laying hens. Ital. J. Anim. Sci. 2014, 78, 1–12. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means Folin-Ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Feucht, W.; Polster, J. Nuclei of plants as a sink for flavanols. J. Biosci. 2001, 56, 479–481. [Google Scholar] [CrossRef] [PubMed]
- Lo Scalzo, R.; Genna, A.; Branca, F.; Chedin, M.; Chassaigne, H. Anthocyanin composition of cauliflower (Brassica oleracea L. var. botrytis) and cabbage (B. oleracea L. var. capitata) and its stability in relation to thermal treatments. Food Chem. 2008, 107, 136–144. [Google Scholar] [CrossRef]
- Ozgen, M.; Reese, R.N.; Tulio, A.Z., Jr.; Scheerens, J.C.; Miller, A.R. Modified 2,2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS) method to measure antioxidant capacity of selected small fruits and comparison to ferric reducing antioxidant power (FRAP) and 2,2’-diphenyl-1-picrylhydrazyl (DPPH) methods. J. Agric. Food Chem. 2006, 54, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Sellappan, S.; Akoh, C.C. Flavonoids and antioxidant capacity of Georgia-grown Vidalia onions. J. Agric. Food Chem. 2002, 50, 5338–5342. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.A.; Hao, Y.; Su, X.; Zubik, L. Phenol antioxidant quantity and quality in foods: Vegetables. J. Agric. Food Chem. 1998, 46, 3630–3634. [Google Scholar] [CrossRef]
- Stratil, P.; Klejdus, B.; Kuban, V. Determination of total content of phenolics compounds and their antioxidant activity in vegetables—Evaluation of spectrophotometric methods. J. Agric. Food Chem. 2006, 54, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Tang, C.Y. Determination of total phenolic and flavonoid contents in selected fruits and vegetables, as well as their stimulatory effects on mouse splenocyte proliferation. Food Chem. 2007, 101, 140–147. [Google Scholar] [CrossRef]
- Prakash, D.; Singh, B.; Upadhyay, G. Antioxidant and free radical scavenging activities of phenols from onion (Allium cepa). Food Chem. 2007, 102, 1389–1393. [Google Scholar] [CrossRef]
- Nuutila, A.M.; Puupponen-Pimia, R.; Aarni, M.; Oksman-Caldentey, K.M. Comparison of antioxidant activities of onion and garlic extracts by inhibition of lipid peroxidation and radical scavenging activity. Food Chem. 2003, 81, 485–493. [Google Scholar] [CrossRef]
- Yang, J.; Meyers, K.J.; Van der Heide, J.; Liu, R.H. Varietal differences in phenolic content and antioxidant and antiproliferative activities of onions. J. Agric. Food Chem. 2004, 52, 6787–6793. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Vagen, I.M. Onions: A source of unique dietary flavonoids. J. Agric. Food Chem. 2007, 55, 10067–10080. [Google Scholar] [CrossRef] [PubMed]
- Bonaccorsi, P.; Caristi, C.; Gargiulli, C.; Leuzzi, U. Flavonol glucosides in Allium species: A comparative study by means of HPLC-DAD-ESI-MS-MS. Food Chem. 2008, 107, 1668–1673. [Google Scholar] [CrossRef]
- Ninfali, P.; Mea, G.; Giorgini, S.; Rocchi, M.; Bacchiocca, M. Antioxidant capacity of vegetables, spices and dressing relevant to nutrition. Br. J. Nutr. 2005, 93, 257–266. [Google Scholar] [CrossRef] [PubMed]
- De Pascual-Teresa, S.; Santos-Buelga, C.; Rivas-Gonzalo, J.C. Quantitative analysis of flavon-3-ols in Spanish foodstuffs and beverages. J. Agric. Food Chem. 2000, 48, 5331–5337. [Google Scholar] [CrossRef] [PubMed]
- Geetha, M.; Ponmozhi, P.; Saravanakumar, M.; Suganyadevi, P. Extraction of anthocyanin and analyzing its antioxidant properties from different onion (Allium cepa L.) varieties. Int. J. Pharm 2011, 2, 497–506. [Google Scholar]
- Santas, J.; Carbo, R.; Gordon, M.H.; Almajano, M.P. Comparison of the antioxidant activity of two Spanish onion varieties. Food Chem. 2008, 107, 1210–1216. [Google Scholar] [CrossRef]
- Bahorun, T.; Luximon-Ramma, A.; Crozier, A.; Aruoma, O.I. Total phenol, flavonoid, proanthocyanidin and vitamin C levels and antioxidant activities of Mauritian vegetables. J. Agric. Food Chem. 2004, 84, 1553–1561. [Google Scholar] [CrossRef]
- Kevers, C.; Falkowski, M.; Tabert, J.; Defraigne, J.O.; Dommes, J.; Pincemail, J. Evaluation of antioxidant capacity during storage of selected fruits and vegetables. J. Agric. Food Chem. 2007, 55, 8596–8603. [Google Scholar] [CrossRef] [PubMed]
- Basciano, H.; Federico, L.; Adeli, K. Fructose, insulin resistance, and metabolic dyslipidemia. Nutr. Metab. 2005, 2, 5–15. [Google Scholar] [CrossRef]
- Mahfouz, M.M.; Kummerow, F.A. Cholesterol-rich diets have different effects on lipid peroxidation, cholesterol oxides, and antioxidant enzymes in rats and rabbits. J. Nutr. Biochem. 2000, 11, 293–302. [Google Scholar] [CrossRef]
- Lawson, L.D.; Gardner, C.D. Composition, stability and bioavailability of garlic products used in a clinical trial. J. Agric. Food Chem. 2005, 53, 6254–6261. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Guo, F.f.; Zhang, C.I.; Zhao, S.Z.; Dou, D.D.; Gao, X.C.; Xie, K.Q.; Yu, L.; Xie, K. The anti-fatty liver effects of garlic oil on acute ethanol-exposed mice. Chem. Biol. Interact. 2008, 176, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Mathew, B.C.; Daniel, R.S. Hypolipidemic effect of garlic protein substituted for caseinin diet of rats compared to those of garlic oil. Indian J. Exp. Biol. 1996, 34, 337–340. [Google Scholar]
- Rajasree, C.R.; Rajmoha, T.; Agusti, K.T. Biochemical effects of garlic on lipid metabolism in alcohol fed rats. Indian J. Exp. Biol. 1999, 37, 234–247. [Google Scholar]
- Gardner, C.D.; Chatterjee, L.M.; Carlson, J.J. The effect of a garlic preparation on plasma lipid levels in moderately hypercholesterolemic adults. Atherosclerosis 2001, 154, 213–220. [Google Scholar] [CrossRef]
- Koseoglu, M.; Isleten, F.; Atay, A.; Kaplan, Y.C. Effect of acute and subacute garlic supplement administration on serum total antioxidant capacity and lipid parameters in healthy volunteers. Phytother. Res. 2010, 24, 374–380. [Google Scholar] [CrossRef]
- Aouadi, R.; Aouidet, A.; Elkadhi, A.; Rayana, C.B.; Jaafoura, H.; Tritar, B.; Nagati, K. Effect of fresh garlic (Allium sativum L.) on lipid metabolizm in male rats. Nutr. Res. 2000, 20, 273–280. [Google Scholar] [CrossRef]
- Kumari, K.; Methew, B.C.; Augusti, K.T. Antidiabetic and hypolipidemic effects of S-methyl cysteine sulfoxide isolated from Allium cepa L. Indian J. Biochem. Biophys. 1995, 32, 49–54. [Google Scholar]
- Liu, Y.L.; Shieh, M.S.; Hsin, C.; Shieh, M.J. Garlic presents better effect than onion on lowering plasma lipids and antioxidative function in hamsters. Atherosclerosis 1998, 136, S80. [Google Scholar] [CrossRef]
- Ros, E. Intestinal absorption of triglyceride and cholesterol. Dietary and pharmacological inhibition to reduce cardiovascular risk. Atherosclerosis 2000, 151, 357–379. [Google Scholar] [CrossRef]
- Yeh, Y.Y.; Yeh, S.M. Garlic reduces plasma lipids by inhibiting hepatic cholesterol and triacylglycerols synthesis. Lipids 1994, 29, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, A.A.; Crenshaw, T.D.; Abuirmeileh, N.; Peterson, D.M.; Elson, C.E. Influence of minor plant constituents on porcine hepatic lipid metabolism. Impact on serum lipids. Atherosclerosis 1987, 64, 109–115. [Google Scholar] [CrossRef]
- Bok, S.H.; Park, S.Y.; Lee, M.K.; Jeon, S.M.; Jeong, T.S.; Choi, M.S. Quercetin dehydrate and gallate supplements lower plasma and hepatic lipids and changes activities of hepatic antioxidant enzymes in cholesterol-fed rats. Int. J. Vitam. Nutr. Res. 2002, 72, 161–169. [Google Scholar] [CrossRef]
- Gebhardt, R.; Beck, H. Differential inhibitory effects of garlic-derived organosulfur compounds on cholesterol biosynthesis in primary rat hepatocyte culture. Lipids 1996, 31, 1269–1276. [Google Scholar] [CrossRef]
- Siegel, G.; Klussendorf, D. The anti-atheroslerotic effect of Allium sativum: Statistics re-evaluated. Atherosclerosis 2000, 150, 437–438. [Google Scholar] [CrossRef]
- Chi, M.S.; Koh, E.T.; Stewart, T.J. Effect of garlic on lipid metabolism in rats fed cholesterol or lard. J. Nutr. 1982, 112, 241–248. [Google Scholar] [CrossRef]
- Oliveira, T.V.; Maniero, F.; Santos, M.H.H.; Bydlowski, S.P.; Moranhăo, R.C. Impact of high cholesterol intake on tissue cholesterol content and lipid transfers to high-density lipoprotein. Nutrition 2011, 27, 713–718. [Google Scholar] [CrossRef]
- Steinberg, D. Low density lipoprotein oxidation and its pathobiological significance. Int. J. Biol. Chem. 1997, 272, 20963–20966. [Google Scholar] [CrossRef]
- Banerjee, S.K.; Maulik, M.; Mancahanda, S.C.; Dinda, A.K.; Gupta, S.K.; Maulik, S.K. Dose-dependend induction of endogenous antioxidants in rat heart by chronic administration of garlic. Life Sci. 2002, 70, 1509–1518. [Google Scholar] [CrossRef]
- Imai, J.; Ide, N.; Nagae, S.; Moriguchi, T.; Matsuura, H.; Itakura, Y. Antioxidant and radical scavenging effects of aged garlic extract and its constituents. Planta Med. 1994, 60, 417–420. [Google Scholar] [CrossRef]
- Geng, Z.; Lau, B.H.S. Aged garlic extract modulates glutathione redox cycle and superoxide dismutase activity in vascular endothelial cells. Phytother. Res. 1997, 11, 54–60. [Google Scholar] [CrossRef]
- Ide, N.; Lau, B.H.S. Garlic compounds minimise intracellular oxidative stress and inhibit nuclear factor-KB activation. J. Nutr. 2001, 131, 1020–1026. [Google Scholar] [CrossRef]
- Leake, D.S. Flavonoids and oxidation of low-density lipoprotein. Nutrition 2001, 17, 63–66. [Google Scholar] [CrossRef]
- Ibarra, M.; Perez-Vizcaino, F.; Cogolludo, A.; Duarte, J.; Zaragoza-Arnaez, F.; Lopez-Lopez, J.G. Cardiovascular effects of isorhamnetin and quercetin in isolated rat and porcine vascular smooth muscle and isolated rat atria. Planta Med. 2002, 68, 307–310. [Google Scholar] [CrossRef] [PubMed]
- El-Sheshtawy, S.M.; Abeer, E.K.M.S.; Eltalawy, M.F. Antioxidant potential and toxicity of garlic (Allium sativum). Egypt. J. Chem. Environ. Health 2016, 2, 56–65. [Google Scholar]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955–962. [Google Scholar] [CrossRef]
- Kodera, Y. Dietary Tolerance/Absorption/Metabolism of Garlic. In Nutraceuticals: Designer Foods III Garlic, Soy and Licorice; Trumbell, C.T., Lanchance, P., Eds.; Food and Nutrition Press: Thousand Oaks, CA, USA, 1997; pp. 95–103. [Google Scholar]
- Bajaj-Elliott, M.; Poulsom, R.; Pender, S.L.F.; Wathen, N.C.; MacDonald, T.T. Interactions between stromal cell-derived keratinocyte growth factor and epithelial transforming growth factor in immune-mediated crypt cell hyperplasia. J. Clin. Investig. 1998, 102, 1473–1480. [Google Scholar] [CrossRef]
- Harris, J.C.; Plummer, S.; Turner, M.P.; Lloyd, D. The microaerophillic flagellate Giardia intestinalis: Allium sativum (garlic) is an effective antigiardial. Microbiology 2000, 146, 3119–3127. [Google Scholar] [CrossRef] [PubMed]
- Augusti, K.T. Therapeutic values of onion (Allium cepa L.) and garlic (Allium sativum L.). Indian J. Exp. Biol. 1996, 34, 634–640. [Google Scholar] [PubMed]
- Hemmaid, K.Z.; Rahmy, T.R. Does preliminary oral administration of garlic diminish histopathological effects of cobra venom in gastric and hepatic tissues? J. Egypt. Ger. Soc. Zool. C Histol. Histochem. 1999, 29, 29–47. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Polyphenols [mg GAE/g d.m.] | Flavonoids [mg CE/g d.m.] | Flavanols [µg CE/g d.m.] | Anthocyanins [mg CGE/kg d.m.] | ABTS [µM TE/g d.m.] |
|---|---|---|---|---|---|
| G | 19.4 ± 1.1 a | 3.4 ± 0.3 a | 6.7 ± 0.4 c | LQQ | 47.7 ± 1.7 a |
| OW | 24.5 ± 1.2 b | 4.0 ± 0.3 b | 4.8 ± 0.4 a | 28.3 ± 1.3 a | 50.9 ± 3.0 b |
| OR | 29.7 ± 1.3 c | 3.8 ± 0.3 b | 5.9 ± 0.4 b | 460.2 ± 10.9 b | 69.9 ± 3.1 c |
| Group | Feed Intake [g] | Cholesterol Intake [g] | Lyophilized Vegetables Intake [g] |
|---|---|---|---|
| C | 589.8 ± 68.4 b | - | - |
| CH | 501.3 ± 36.3 a | 5.0 ± 0.4 a | - |
| CHG | 480.9 ± 42.0 a | 4.8 ± 0.4 a | 578.6 ± 50.5 a |
| CHOW | 514.0 ± 16.2 a | 5.1 ± 0.2 a | 621.0 ± 22.4 a |
| CHOR | 578.9 ± 16.3 b | 5.8 ± 0.2 b | 700.1 ± 23.2 b |
| Group | Body Weight Gain [g BW] | FER [g/g BW] | SI-L [% BW] |
|---|---|---|---|
| C | 178.2 ± 27.0 b | 3.3 ± 0.3 a | 3.6 ± 0.4 a |
| CH | 134.9 ± 26.2 a | 3.8 ± 0.6 ab | 3.7 ± 0.4 a |
| CHG | 113.9 ± 27.0 a | 4.3 ± 0.5 b | 3.3 ± 0.3 a |
| CHOW | 134.3 ± 12.1 a | 3.8 ± 0.3 ab | 3.3 ± 0.3 a |
| CHOR | 175.4 ± 17.8 b | 3.3 ± 0.3 a | 3.7 ± 0.4 a |
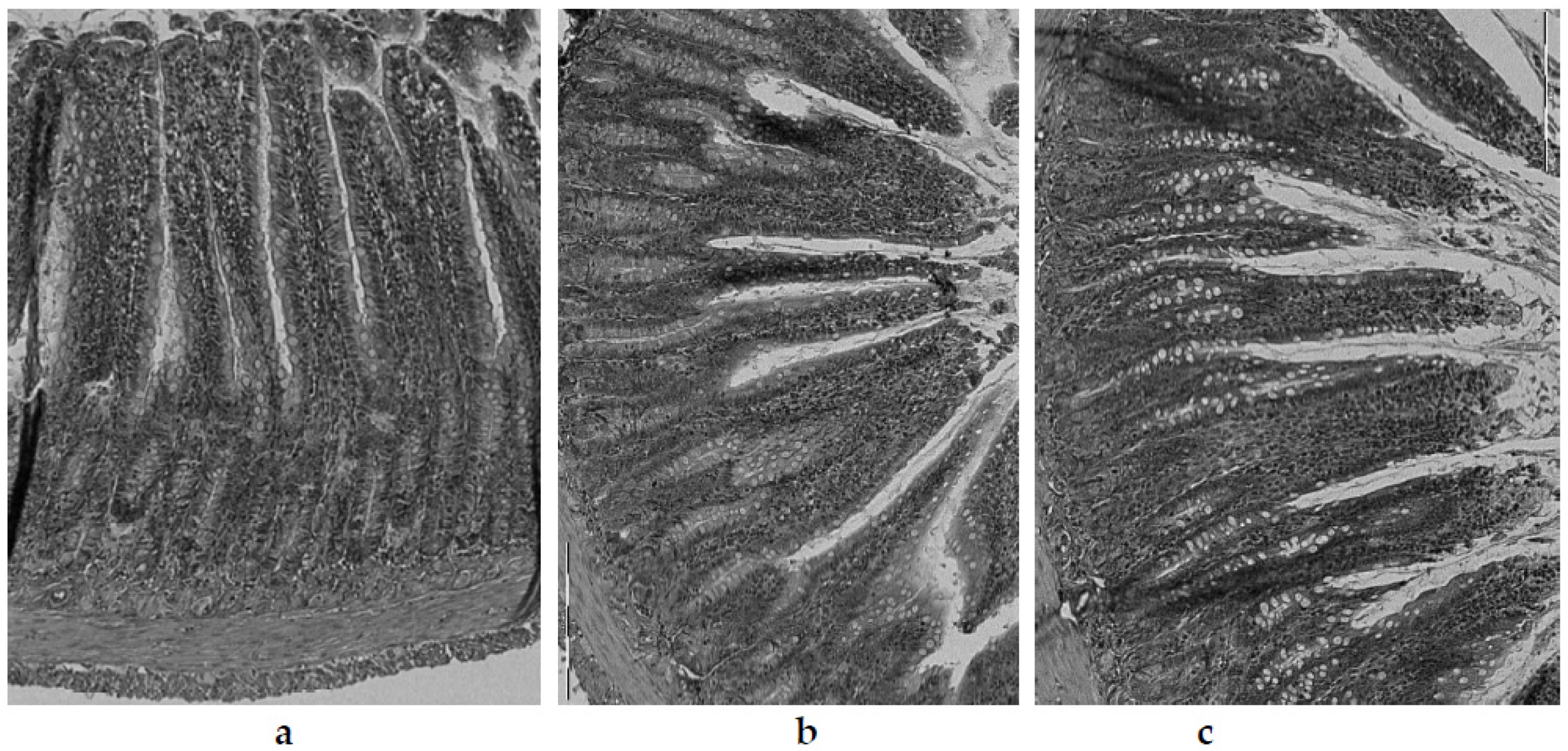
| Group | Villi Length [µm] | Crypt Depth [µm] | Thickness of Tunica Mucosa [µm] | Thickness of Tunica Muscle [µm] |
|---|---|---|---|---|
| C | 468.6 ± 6.4 e | 130.5 ± 1.8 a | 624.5 ± 7.0 d | 60.2 ± 0.2 a |
| CH | 363.6 ± 4.9 c | 135.2 ± 5.2 ab | 527.7 ± 5.9 b | 67.0 ± 0.5 bc |
| CHG | 305.7 ± 2.1 a | 136.1 ± 4.4 b | 465.2 ± 7.4 a | 69.1 ± 2.4 c |
| CHOW | 435.4 ± 16.1 d | 131.2 ± 3.0 a | 581.2 ± 14.9 c | 65.9 ± 2.5 b |
| CHOR | 330.2 ± 25.9 b | 133.5 ± 2.9 a | 544.3 ± 16.1 b | 66.3 ± 1.7 bc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Najman, K.; Leontowicz, H.; Leontowicz, M. The Influence of Plants from the Alliaceae Family on Morphological Parameters of the Intestine in Atherogenic Rats. Nutrients 2021, 13, 3876. https://doi.org/10.3390/nu13113876
Najman K, Leontowicz H, Leontowicz M. The Influence of Plants from the Alliaceae Family on Morphological Parameters of the Intestine in Atherogenic Rats. Nutrients. 2021; 13(11):3876. https://doi.org/10.3390/nu13113876
Chicago/Turabian StyleNajman, Katarzyna, Hanna Leontowicz, and Maria Leontowicz. 2021. "The Influence of Plants from the Alliaceae Family on Morphological Parameters of the Intestine in Atherogenic Rats" Nutrients 13, no. 11: 3876. https://doi.org/10.3390/nu13113876
APA StyleNajman, K., Leontowicz, H., & Leontowicz, M. (2021). The Influence of Plants from the Alliaceae Family on Morphological Parameters of the Intestine in Atherogenic Rats. Nutrients, 13(11), 3876. https://doi.org/10.3390/nu13113876