
Does Physical Inactivity Induce Significant Changes in Human Gut Microbiota? New Answers Using the Dry Immersion Hypoactivity Model

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dry Immersion
2.2. Participants
2.3. Body Composition and Diet
2.4. Overall Study Design
2.5. Stool Collection and Metagenomic Analysis
2.5.1. DNA Extraction from Feces
2.5.2. Evaluation of Total Bacteria by Real-Time qPCR Analysis of Bacterial 16 s rRNA Genes
2.5.3. Phylum Abundance Quantification by Real-Time qPCR
2.5.4. Microbiota Composition Analysis by Sequencing
2.5.5. Metagenomic Analysis
2.6. Short-Chain Fatty Acid Analysis
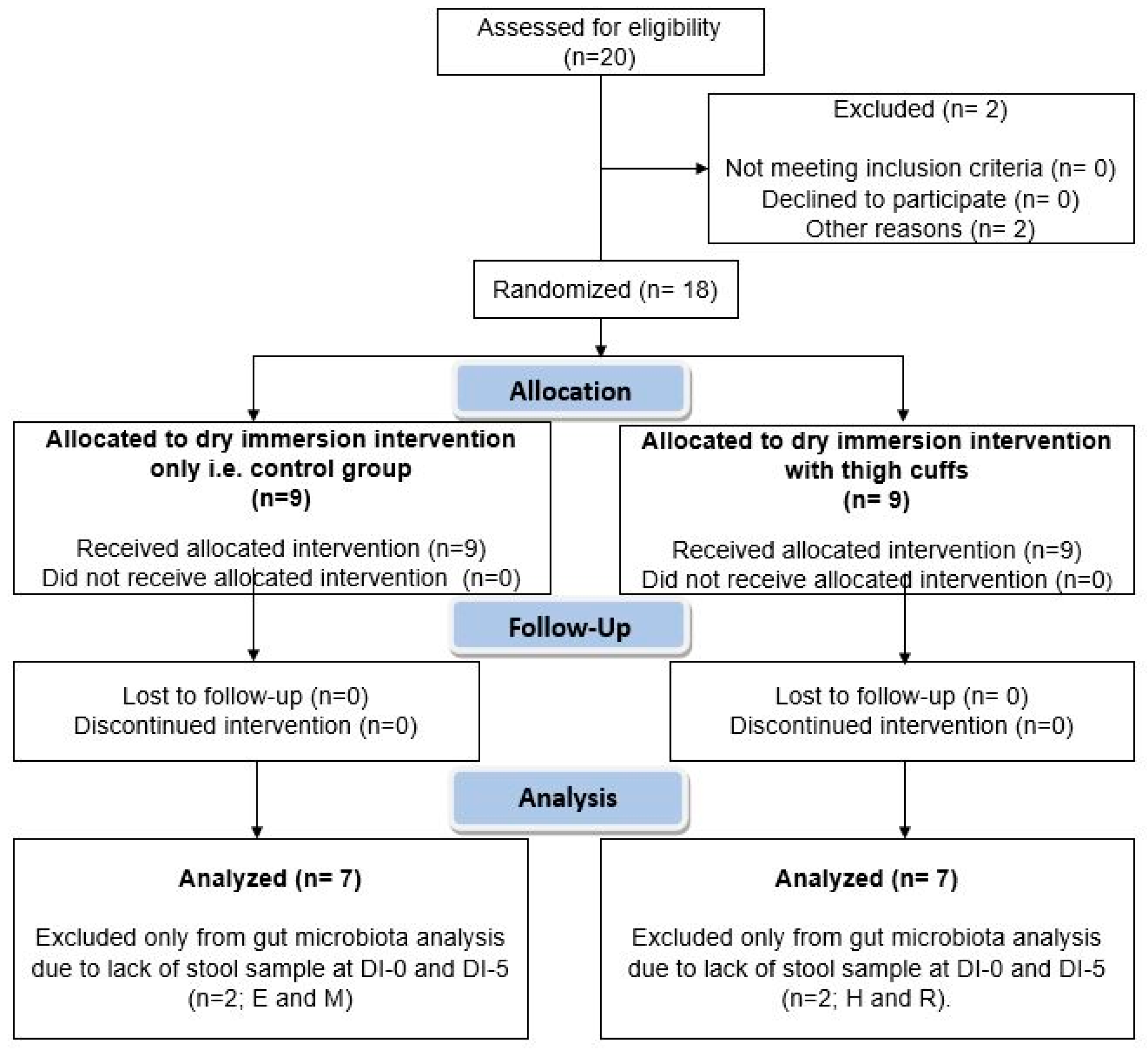
2.7. Participant Flow and Statistics
3. Results
3.1. Dry Immersion-Induced Muscle Atrophy Despite a Controlled and Preserved Nutrient Intake
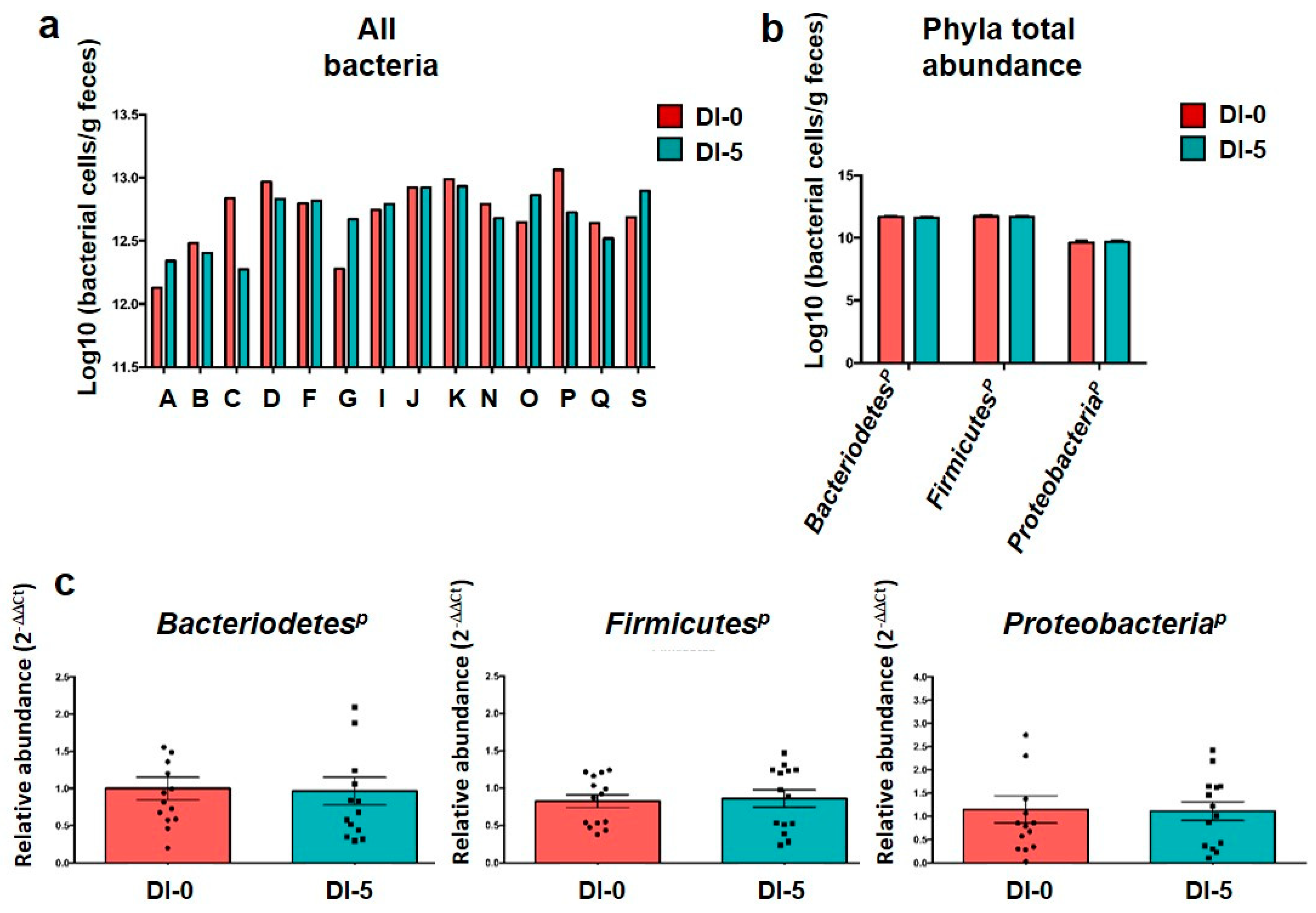
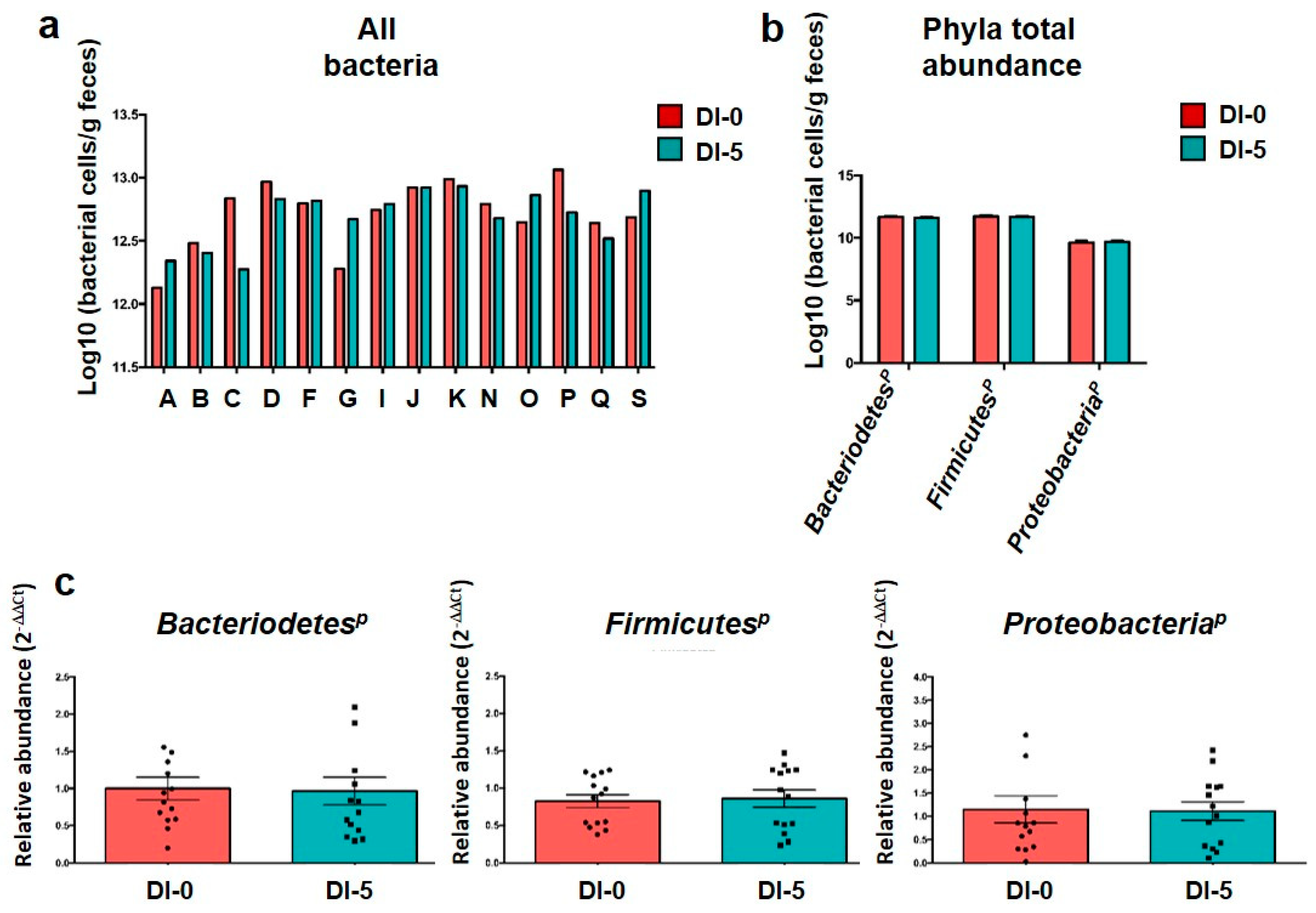
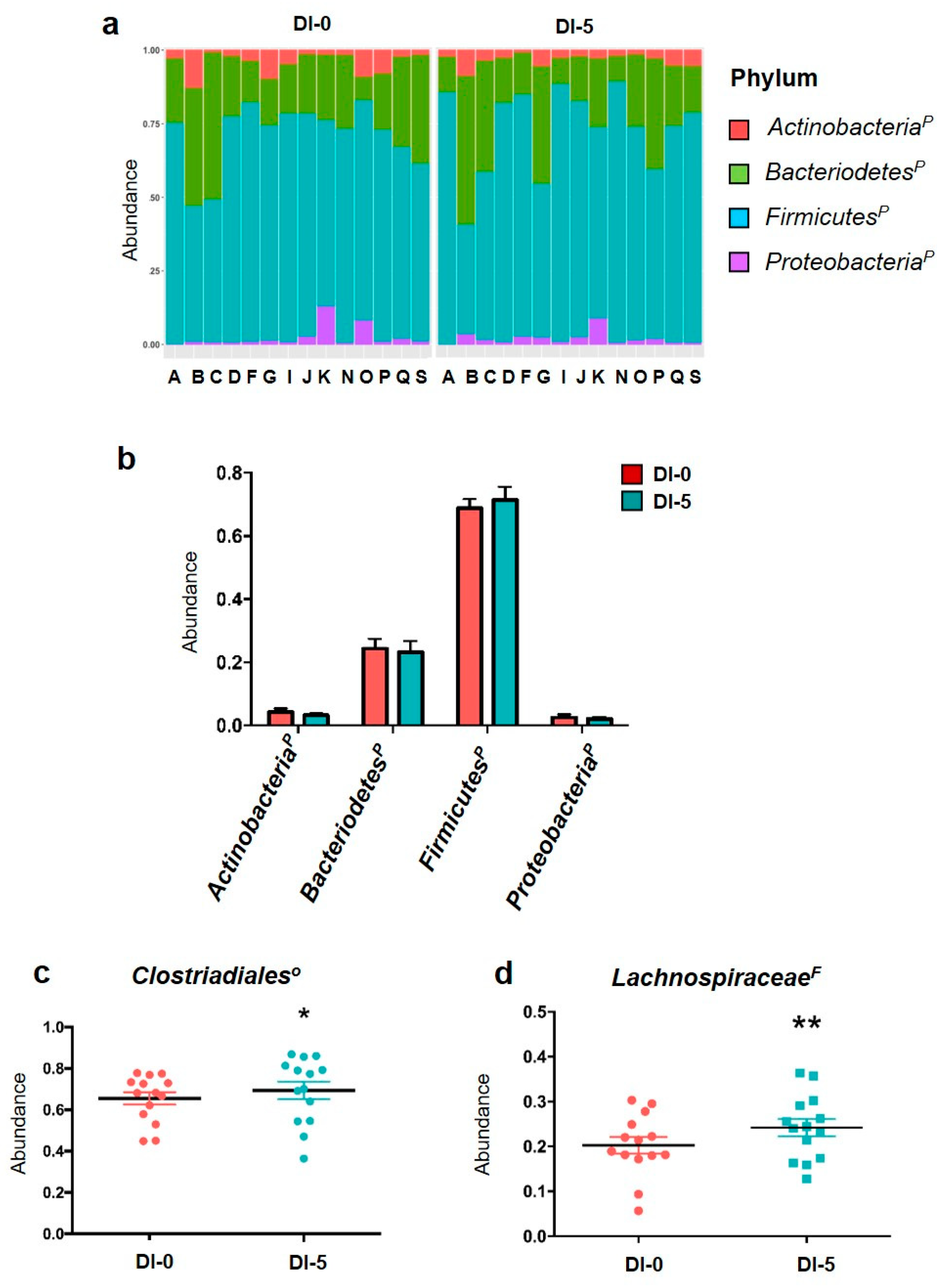
3.2. The Abundance of the Main Microbiota Phyla Is Not Affected by Dry Immersion
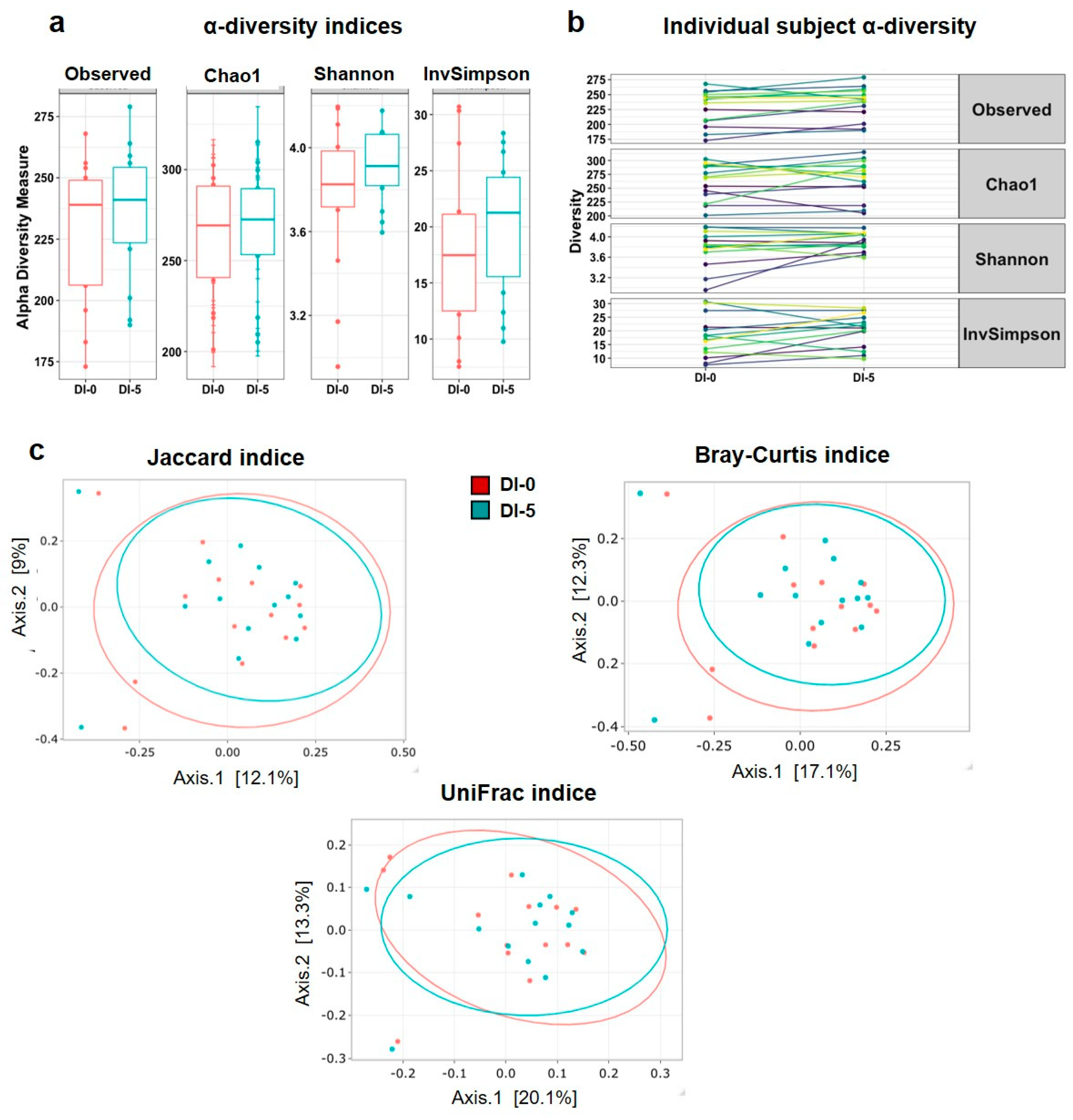
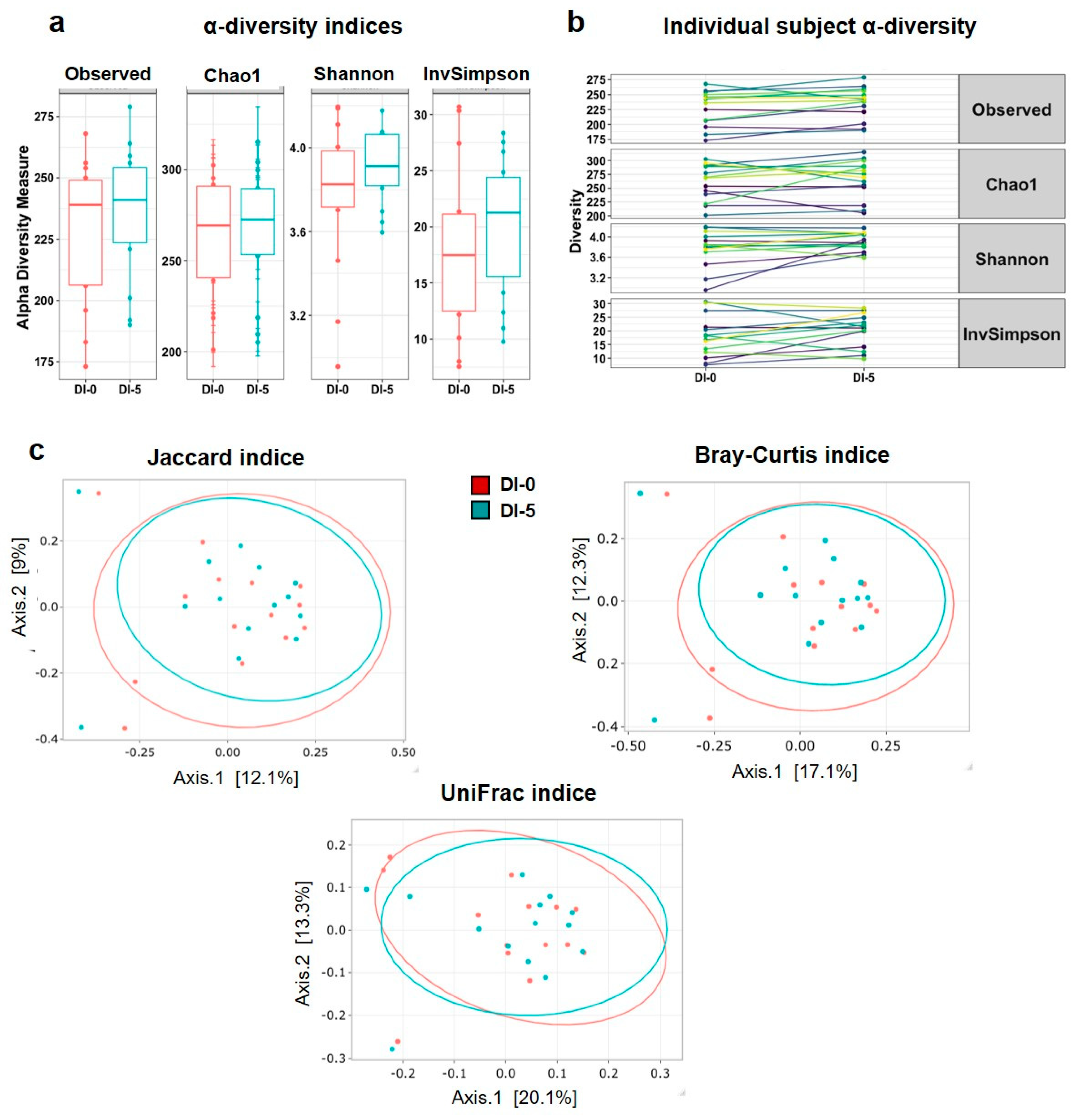
3.3. Dry Immersion Does Not Significantly Affect α and β Diversity
3.4. DI Affects OTUs Associated with the Clostridiales Order and the Lachnospiraceae Family with Impacts on Bacterial Functional Profiles Linked to Anaerobic Glycolysis
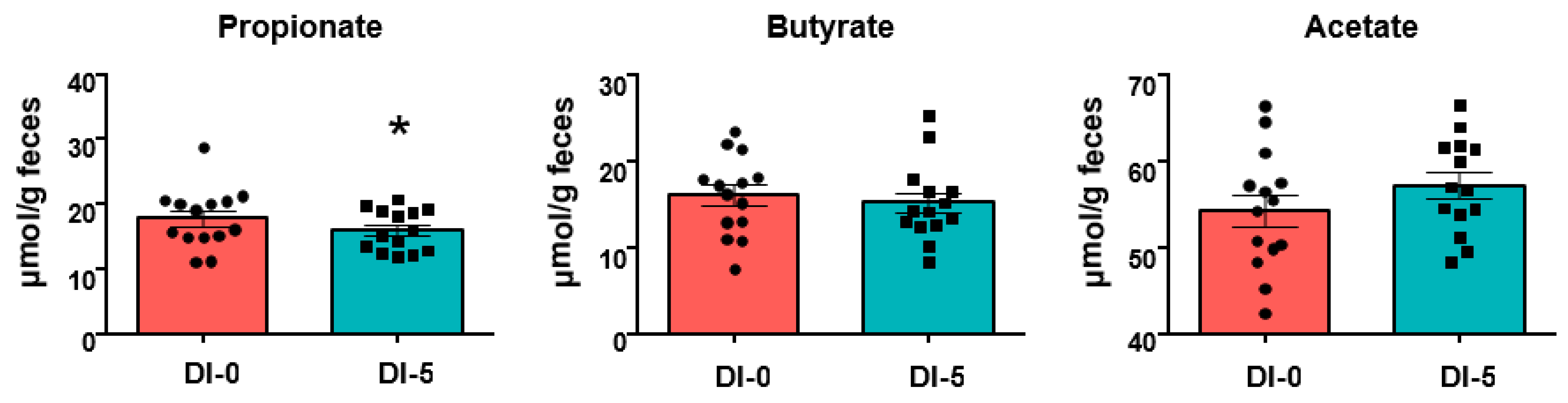
3.5. Propionate Production Is Decreased by 5 Days of Dry Immersion
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clemente, J.C.; Ursell, L.K.; Parfrey, L.W.; Knight, R. The impact of the gut microbiota on human health: An integrative view. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohr, A.E.; Jager, R.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Townsend, J.R.; West, N.P.; Black, K.; Gleeson, M.; Pyne, D.B.; et al. The athletic gut microbiota. J. Int. Soc. Sports Nutr. 2020, 17, 24. [Google Scholar] [CrossRef]
- Marttinen, M.; Ala-Jaakkola, R.; Laitila, A.; Lehtinen, M.J. Gut Microbiota, Probiotics and Physical Performance in Athletes and Physically Active Individuals. Nutrients 2020, 12, 2936. [Google Scholar] [CrossRef]
- Juarez-Fernandez, M.; Roman-Saguillo, S.; Porras, D.; Garcia-Mediavilla, M.V.; Linares, P.; Ballesteros-Pomar, M.D.; Urioste-Fondo, A.; Alvarez-Cuenllas, B.; Gonzalez-Gallego, J.; Sanchez-Campos, S.; et al. Long-Term Effects of Bariatric Surgery on Gut Microbiota Composition and Faecal Metabolome Related to Obesity Remission. Nutrients 2021, 13, 2519. [Google Scholar] [CrossRef] [PubMed]
- Makin, S. Do microbes affect athletic performance? Nature 2021, 592, S17–S19. [Google Scholar] [CrossRef] [PubMed]
- Charreire, H.; Kesse-Guyot, E.; Bertrais, S.; Simon, C.; Chaix, B.; Weber, C.; Touvier, M.; Galan, P.; Hercberg, S.; Oppert, J.M. Associations between dietary patterns, physical activity (leisure-time and occupational) and television viewing in middle-aged French adults. Br. J. Nutr. 2011, 105, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Son, J.; Jang, L.G.; Kim, B.Y.; Lee, S.; Park, H. The Effect of Athletes’ Probiotic Intake May Depend on Protein and Dietary Fiber Intake. Nutrients 2020, 12, 2947. [Google Scholar] [CrossRef]
- Sheflin, A.M.; Melby, C.L.; Carbonero, F.; Weir, T.L. Linking dietary patterns with gut microbial composition and function. Gut Microbes 2017, 8, 113–129. [Google Scholar] [CrossRef]
- Monda, V.; Villano, I.; Messina, A.; Valenzano, A.; Esposito, T.; Moscatelli, F.; Viggiano, A.; Cibelli, G.; Chieffi, S.; Monda, M.; et al. Exercise Modifies the Gut Microbiota with Positive Health Effects. Oxid. Med. Cell. Longev. 2017, 2017, 3831972. [Google Scholar] [CrossRef]
- De Sire, A.; de Sire, R.; Petito, V.; Masi, L.; Cisari, C.; Gasbarrini, A.; Scaldaferri, F.; Invernizzi, M. Gut-Joint Axis: The Role of Physical Exercise on Gut Microbiota Modulation in Older People with Osteoarthritis. Nutrients 2020, 12, 574. [Google Scholar] [CrossRef] [Green Version]
- Harmsen, H.J.; de Goffau, M.C. The Human Gut Microbiota. Adv. Exp. Med. Biol. 2016, 902, 95–108. [Google Scholar] [CrossRef]
- Hooper, L.V.; Macpherson, A.J. Immune adaptations that maintain homeostasis with the intestinal microbiota. Nat. Rev. Immunol. 2010, 10, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Nay, K.; Jollet, M.; Goustard, B.; Baati, N.; Vernus, B.; Pontones, M.; Lefeuvre-Orfila, L.; Bendavid, C.; Rue, O.; Mariadassou, M.; et al. Gut bacteria are critical for optimal muscle function: A potential link with glucose homeostasis. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E158–E171. [Google Scholar] [CrossRef]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Garrett-Bakelman, F.E.; Darshi, M.; Green, S.J.; Gur, R.C.; Lin, L.; Macias, B.R.; McKenna, M.J.; Meydan, C.; Mishra, T.; Nasrini, J.; et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science 2019, 364. [Google Scholar] [CrossRef]
- Voorhies, A.A.; Mark Ott, C.; Mehta, S.; Pierson, D.L.; Crucian, B.E.; Feiveson, A.; Oubre, C.M.; Torralba, M.; Moncera, K.; Zhang, Y.; et al. Study of the impact of long-duration space missions at the International Space Station on the astronaut microbiome. Sci. Rep. 2019, 9, 9911. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.E.; Chouker, A.; Simpson, R.J.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A.; et al. Immune System Dysregulation During Spaceflight: Potential Countermeasures for Deep Space Exploration Missions. Front. Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef]
- Guthold, R.; Stevens, G.A.; Riley, L.M.; Bull, F.C. Worldwide trends in insufficient physical activity from 2001 to 2016: A pooled analysis of 358 population-based surveys with 1.9 million participants. Lancet Glob. Health 2018, 6, e1077–e1086. [Google Scholar] [CrossRef] [Green Version]
- Demangel, R.; Treffel, L.; Py, G.; Brioche, T.; Pagano, A.F.; Bareille, M.P.; Beck, A.; Pessemesse, L.; Candau, R.; Gharib, C.; et al. Early structural and functional signature of 3-day human skeletal muscle disuse using the dry immersion model. J. Physiol. 2017, 595, 4301–4315. [Google Scholar] [CrossRef]
- Tomilovskaya, E.; Shigueva, T.; Sayenko, D.; Rukavishnikov, I.; Kozlovskaya, I. Dry Immersion as a Ground-Based Model of Microgravity Physiological Effects. Front. Physiol. 2019, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Navasiolava, N.; Yuan, M.; Murphy, R.; Robin, A.; Coupe, M.; Wang, L.; Alameddine, A.; Gauquelin-Koch, G.; Gharib, C.; Li, Y.; et al. Vascular and Microvascular Dysfunction Induced by Microgravity and Its Analogs in Humans: Mechanisms and Countermeasures. Front. Physiol. 2020, 11, 952. [Google Scholar] [CrossRef]
- De Abreu, S.; Amirova, L.; Murphy, R.; Wallace, R.; Twomey, L.; Gauquelin-Koch, G.; Raverot, V.; Larcher, F.; Custaud, M.A.; Navasiolava, N. Multi-System Deconditioning in 3-Day Dry Immersion without Daily Raise. Front. Physiol. 2017, 8, 799. [Google Scholar] [CrossRef] [Green Version]
- Pavy-Le Traon, A.; Heer, M.; Narici, M.V.; Rittweger, J.; Vernikos, J. From space to Earth: Advances in human physiology from 20 years of bed rest studies (1986–2006). Eur. J. Appl. Physiol. 2007, 101, 143–194. [Google Scholar] [CrossRef]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimaraes, V.; Sokol, H.; Dore, J.; Corthier, G.; Furet, J.P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef]
- Nakanishi, Y.; Murashima, K.; Ohara, H.; Suzuki, T.; Hayashi, H.; Sakamoto, M.; Fukasawa, T.; Kubota, H.; Hosono, A.; Kono, T.; et al. Increase in terminal restriction fragments of Bacteroidetes-derived 16S rRNA genes after administration of short-chain fructooligosaccharides. Appl. Environ. Microbiol. 2006, 72, 6271–6276. [Google Scholar] [CrossRef] [Green Version]
- Furet, J.P.; Firmesse, O.; Gourmelon, M.; Bridonneau, C.; Tap, J.; Mondot, S.; Dore, J.; Corthier, G. Comparative assessment of human and farm animal faecal microbiota using real-time quantitative PCR. FEMS Microbiol. Ecol. 2009, 68, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Escudie, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegants Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Lan, A.; Bruneau, A.; Philippe, C.; Rochet, V.; Rouault, A.; Herve, C.; Roland, N.; Rabot, S.; Jan, G. Survival and metabolic activity of selected strains of Propionibacterium freudenreichii in the gastrointestinal tract of human microbiota-associated rats. Br. J. Nutr. 2007, 97, 714–724. [Google Scholar] [CrossRef] [Green Version]
- Adams, G.R.; Caiozzo, V.J.; Baldwin, K.M. Skeletal muscle unweighting: Spaceflight and ground-based models. J. Appl. Physiol. 2003, 95, 2185–2201. [Google Scholar] [CrossRef] [Green Version]
- Chopard, A.; Hillock, S.; Jasmin, B.J. Molecular events and signalling pathways involved in skeletal muscle disuse-induced atrophy and the impact of countermeasures. J. Cell. Mol. Med. 2009, 13, 3032–3050. [Google Scholar] [CrossRef] [Green Version]
- Navasiolava, N.M.; Custaud, M.A.; Tomilovskaya, E.S.; Larina, I.M.; Mano, T.; Gauquelin-Koch, G.; Gharib, C.; Kozlovskaya, I.B. Long-term dry immersion: Review and prospects. Eur. J. Appl. Physiol. 2011, 111, 1235–1260. [Google Scholar] [CrossRef] [Green Version]
- Treffel, L.; Mkhitaryan, K.; Gellee, S.; Gauquelin-Koch, G.; Gharib, C.; Blanc, S.; Millet, C. Intervertebral Disc Swelling Demonstrated by 3D and Water Content Magnetic Resonance Analyses after a 3-Day Dry Immersion Simulating Microgravity. Front. Physiol. 2016, 7, 605. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, K.M.; Haddad, F.; Pandorf, C.E.; Roy, R.R.; Edgerton, V.R. Alterations in muscle mass and contractile phenotype in response to unloading models: Role of transcriptional/pretranslational mechanisms. Front. Physiol. 2013, 4, 284. [Google Scholar] [CrossRef] [Green Version]
- Cros, N.; Muller, J.; Bouju, S.; Pietu, G.; Jacquet, C.; Leger, J.J.; Marini, J.F.; Dechesne, C.A. Upregulation of M-creatine kinase and glyceraldehyde3-phosphate dehydrogenase: Two markers of muscle disuse. Am. J. Physiol. 1999, 276, R308–R316. [Google Scholar] [CrossRef]
- Bodine, S.C.; Latres, E.; Baumhueter, S.; Lai, V.K.; Nunez, L.; Clarke, B.A.; Poueymirou, W.T.; Panaro, F.J.; Na, E.; Dharmarajan, K.; et al. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 2001, 294, 1704–1708. [Google Scholar] [CrossRef]
- Kawashima, S.; Akima, H.; Kuno, S.Y.; Gunji, A.; Fukunaga, T. Human adductor muscles atrophy after short duration of unweighting. Eur. J. Appl. Physiol. 2004, 92, 602–605. [Google Scholar] [CrossRef]
- Ticinesi, A.; Lauretani, F.; Milani, C.; Nouvenne, A.; Tana, C.; Del Rio, D.; Maggio, M.; Ventura, M.; Meschi, T. Aging Gut Microbiota at the Cross-Road between Nutrition, Physical Frailty, and Sarcopenia: Is There a Gut-Muscle Axis? Nutrients 2017, 9, 1303. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, L.E.; Taddeo, S.S.; Weeks, B.R.; Lima, F.; Bloomfield, S.A.; Azcarate-Peril, M.A.; Zwart, S.R.; Smith, S.M.; Turner, N.D. Space Environmental Factor Impacts upon Murine Colon Microbiota and Mucosal Homeostasis. PLoS ONE 2015, 10, e0125792. [Google Scholar] [CrossRef]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef] [Green Version]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Rainey, F.A. The Firmicutes. In Bergey’s Manual of Systematic Bacteriology; De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.H., Whitman, W.B., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2009; Volume 3, p. 921. [Google Scholar]
- Buffie, C.G.; Bucci, V.; Stein, R.R.; McKenney, P.T.; Ling, L.; Gobourne, A.; No, D.; Liu, H.; Kinnebrew, M.; Viale, A.; et al. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature 2015, 517, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Wang, Y.; He, J.; Li, P.; Jin, R.; Wang, K.; Xu, X.; Hao, J.; Zhang, Y.; Liu, H.; et al. Intestinal microbiota contributes to colonic epithelial changes in simulated microgravity mouse model. FASEB J. 2017, 31, 3695–3709. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.; Mach, N. The Crosstalk between the Gut Microbiota and Mitochondria during Exercise. Front. Physiol. 2017, 8, 319. [Google Scholar] [CrossRef]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nat. Metab. 2020, 2, 840–848. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTL (n = 9) | CUFFS (n = 9) | ALL (n = 18) | |
|---|---|---|---|
| Age (years) | 33.4 ± 7.1 | 33.8 ± 3.7 | 33.6 ± 5.5 |
| Height (cm) | 176 ± 6 | 180 ± 4 | 178 ± 6 |
| Weight (kg) | 73.9 ± 7.5 | 74.3 ± 8.8 | 74.4 ± 8.0 |
| BMI (kg/m2) | 23.9 ± 1.7 | 22.7 ± 1.8 | 23.5 ± 1.9 |
| VO2max (ml/min/kg) | 46.5 ± 8.1 | 46.9 ± 5.8 | 46.7 ± 6.9 |
| Morning HR (bpm) | 57 ± 6 | 58 ± 8 | 58 ± 7 |
| Morning T (°C) | 36.4 ± 0.3 | 36.4 ± 0.5 | 36.4 ± 0.4 |
| Morning SBP (mmHg) | 115 ± 11 | 117 ± 10 | 116 ± 10 |
| Morning DBP (mmHg) | 68 ± 5 | 68 ± 9 | 68 ± 7 |
| CTL (n = 9) | CUFFS (n = 9) | ALL (n = 18) | |
|---|---|---|---|
| Whole body lean mass (kg) | |||
| BDC-4 | 55.5 ± 4.7 | 55.8 ± 6.7 | 55.6 ± 5.6 |
| DI-5 | 54.2 ± 4.5 *** | 54.3 ± 6.5 *** | 54.2 ± 5.4 *** |
| Leg lean mass (kg) | |||
| BDC-4 | 18.3 ± 2.5 | 18.4 ± 1.9 | 18.3 ± 2.2 |
| DI-5 | 17.6 ± 2.1 *** | 17.7 ± 1.7 *** | 17.7 ± 1.9 *** |
| Time | Precribed Energy (Kcal) | Energy (kcal) | Carbohydrates (g) | Proteins (g) | Total Fat (g) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| BDC-4 | 2624.8 | 241.7 | 2625.1 | 241.3 | 304.0 | 28.4 | 81.5 | 8.0 | 101.2 | 9.3 | ||
| BDC-3 | 2624.8 | 241.7 | 2624.8 | 241.6 | 322.6 | 30.9 | 85.1 | 8.4 | 103.9 | 9.8 | ||
| BDC-2 | 2624.8 | 241.7 | 2624.6 | 241.5 | 324.5 | 32.3 | 86.7 | 8.4 | 102.7 | 9.3 | ||
| BDC-1 | 2624.8 | 241.7 | 2624.6 | 241.2 | 322.8 | 34.4 | 87.4 | 8.9 | 101.3 | 8.8 | ||
| DI-1 | 2160.1 | 205.7 | 2152.0 | 210. | 252.8 | 27.6 | 80.6 | 6.3 | 82.3 | 8.7 | ||
| DI-2 | 2160.1 | 205.7 | 2160.5 | 205.2 | 250.9 | 24.3 | 78.3 | 7.4 | 86.3 | 9.0 | ||
| DI-3 | 2160.1 | 205.7 | 2160.4 | 205.7 | 255.5 | 23.0 | 85.7 | 9.7 | 83.5 | 8.9 | ||
| DI-4 | 2160.1 | 205.7 | 2160.2 | 205.5 | 252.7 | 24.6 | 82.4 | 7.8 | 83.6 | 8.1 | ||
| DI-5 | 2160.1 | 205.7 | 2160.5 | 205.5 | 237.5 | 25.1 | 75.3 | 7.9 | 81.6 | 8.1 | ||
| R+0 | 2658.8 | 253.1 | 2658.6 | 253.0 | 328.0 | 31.5 | 84.8 | 7.6 | 105.3 | 10.7 | ||
| R+1 | 2658.8 | 253.1 | 2658.9 | 253.0 | 325.9 | 30.7 | 85.8 | 8.0 | 102.8 | 10.7 | ||
| Time | Saturated fatty acids (g) | Monounsaturated fatty acids (g) | Polyunsaturated fatty acids (g) | Total water (g) | Fibers (g) | |||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| BDC-4 | 36.1 | 3.6 | 34.7 | 3.9 | 22.9 | 2.2 | 3422.6 | 530.2 | 39.2 | 3.3 | ||
| BDC-3 | 35.6 | 3.3 | 31.1 | 3.4 | 24.5 | 2.5 | 3462.5 | 485.0 | 35.0 | 2.8 | ||
| BDC-2 | 28.1 | 2.5 | 41.8 | 5.1 | 22.6 | 1.5 | 3585.2 | 450.0 | 35.7 | 3.4 | ||
| BDC-1 | 23.6 | 2.3 | 38.1 | 3.8 | 27.2 | 3.3 | 3490.4 | 532.8 | 40.1 | 2.8 | ||
| DI-1 | 17.8 | 2.1 | 31.1 | 4.5 | 24.8 | 2.6 | 3043.0 | 608.6 | 36.1 | 3.1 | ||
| DI-2 | 25.5 | 3.1 | 33.5 | 3.6 | 17.2 | 2.0 | 3307.3 | 492.8 | 32.1 | 2.2 | ||
| DI-3 | 24.0 | 2.7 | 31.7 | 3.8 | 19.1 | 1.9 | 3199.1 | 504.4 | 28.5 | 2.2 | ||
| DI-4 | 21.9 | 2.2 | 29.4 | 3.7 | 20.3 | 2.1 | 3287.9 | 465.1 | 33.3 | 2.9 | ||
| DI-5 | 30.3 | 2.9 | 27.2 | 3.9 | 18.0 | 1.8 | 3111.8 | 552.1 | 33.1 | 3.1 | ||
| R+0 | 35.0 | 4.0 | 32.0 | 3.6 | 25.5 | 2.9 | 4032.4 | 616.5 | 35.1 | 2.7 | ||
| R+1 | 22.4 | 2.3 | 41.3 | 5.8 | 29.3 | 3.0 | 3793.3 | 548.3 | 46.2 | 3.5 | ||
| Time | Sodium (mg) | Chloride (mg) | Potassium (mg) | Calcium (mg) | Magnesium (mg) | Phosphorus (mg) | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| BDC-4 | 3632.3 | 373.6 | 6656.0 | 674.7 | 3548.1 | 286.3 | 1360.5 | 112.5 | 353.3 | 33.1 | 1319.2 | 120.7 |
| BDC-3 | 3726.3 | 349.3 | 7027.0 | 670.3 | 4974.4 | 427.3 | 1399.9 | 150.6 | 392.5 | 36.0 | 1620.3 | 169.3 |
| BDC-2 | 3109.2 | 284.2 | 5308.4 | 460.8 | 3424.1 | 229.2 | 1231.7 | 74.5 | 419.5 | 31.9 | 1134.7 | 89.9 |
| BDC-1 | 4006.2 | 424.4 | 7255.6 | 757.7 | 4480.5 | 284.8 | 1327.9 | 103.8 | 479.4 | 29.5 | 1364.8 | 113.2 |
| DI-1 | 2392.5 | 208.0 | 4787.9 | 373.0 | 3440.3 | 211.0 | 1118.4 | 102.9 | 398.3 | 26.3 | 1396.3 | 107.7 |
| DI-2 | 3034.6 | 308.5 | 5364.8 | 542.1 | 2986.5 | 186.7 | 1025.9 | 101.6 | 322.9 | 23.5 | 1260.0 | 112.6 |
| DI-3 | 2837.7 | 266.5 | 4856.9 | 392.7 | 3101.4 | 132.9 | 1133.3 | 106.3 | 375.9 | 26.3 | 1071.1 | 106.0 |
| DI-4 | 3620.7 | 331.0 | 6478.1 | 607.5 | 3690.0 | 329.5 | 1233.0 | 95.8 | 406.4 | 32.8 | 1247.1 | 109.9 |
| DI-5 | 3036.4 | 280.8 | 5509.5 | 503.0 | 3286.5 | 274.2 | 1254.1 | 122.3 | 310.6 | 24.6 | 1198.9 | 102.4 |
| R+0 | 3755.6 | 299.8 | 6953.9 | 578.0 | 4953.1 | 333.9 | 1407.2 | 146.0 | 408.7 | 33.8 | 1614.4 | 148.7 |
| R+1 | 2706.0 | 197.8 | 5305.3 | 384.5 | 3934.8 | 283.6 | 1251.3 | 115.6 | 463.9 | 39.0 | 1511.9 | 142.2 |
| Time | Vitamin A (µg_RE) | Vitamin K (µg) | Vitamin C (mg) | Niacin (vit PP) (mg) | Riboflavin (vit B-2) (mg) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | Mean | SD | |
| BDC-4 | 1736.2 | 162.2 | 301.8 | 29.2 | 406.0 | 45.0 | 30.9 | 3.1 | 2.3 | 0.2 |
| BDC-3 | 911.7 | 123.8 | 520.7 | 51.5 | 191.7 | 18.8 | 43.1 | 4.6 | 2.1 | 0.2 |
| BDC-2 | 2712.2 | 149.3 | 95.6 | 10.2 | 86.64 | 10.7 | 29.0 | 3.1 | 1.7 | 0.1 |
| BDC-1 | 898.4 | 56.6 | 483.6 | 35.7 | 402.3 | 16.9 | 37.0 | 3.6 | 2.1 | 0.2 |
| DI-1 | 1924.8 | 123.0 | 496.5 | 37.1 | 179.5 | 14.9 | 46.2 | 3.8 | 1.6 | 0.1 |
| DI-2 | 826.1 | 48.4 | 487.5 | 47.6 | 170.2 | 13.0 | 31.4 | 2.8 | 2.0 | 0.1 |
| DI-3 | 2397.6 | 169.1 | 74.4 | 5.8 | 71.2 | 6.6 | 28.5 | 3.3 | 1.5 | 0.1 |
| DI-4 | 830.8 | 59.1 | 473.5 | 38.7 | 358.0 | 34.0 | 34.9 | 3.2 | 1.9 | 0.2 |
| DI-5 | 1489.1 | 111.4 | 277.5 | 27.0 | 372.5 | 43.3 | 28.3 | 2.8 | 2.0 | 0.2 |
| R+0 | 878.2 | 131.2 | 503.2 | 41.6 | 194.5 | 23.6 | 43.3 | 3.4 | 2.1 | 0.2 |
| R+1 | 2162.4 | 219.0 | 507.9 | 51.2 | 210.8 | 19.1 | 46.9 | 4.6 | 1.9 | 0.2 |
| Time | Pantothenic Acid (vit B-5) (mg) | Thiamin (vit B-1) (mg) | Vitamin D (µg) | Biotin (vit H) (µg) | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| BDC-4 | 6.7 | 0.6 | 2.0 | 0.2 | 1.5 | 0.5 | 33.7 | 2.6 | ||
| BDC-3 | 8.0 | 0.8 | 2.0 | 0.2 | 3.1 | 0.5 | 57.0 | 5.9 | ||
| BDC-2 | 5.5 | 0.4 | 1.8 | 0.1 | 2.9 | 0.2 | 42.8 | 2.9 | ||
| BDC-1 | 6.6 | 0.5 | 2.3 | 0.2 | 1.5 | 0.2 | 45.4 | 3.6 | ||
| DI-1 | 5.8 | 0.4 | 1.8 | 0.1 | 2.9 | 0.3 | 50.2 | 3.9 | ||
| DI-2 | 5.2 | 0.3 | 1.3 | 0.1 | 1.8 | 0.3 | 42.6 | 2.9 | ||
| DI-3 | 5.0 | 0.4 | 1.6 | 0.1 | 2.7 | 0.3 | 37.6 | 2.8 | ||
| DI-4 | 6.0 | 0.5 | 2.1 | 0.2 | 1.4 | 0.1 | 36.1 | 3.3 | ||
| DI-5 | 6.0 | 0.5 | 1.6 | 0.1 | 1.1 | 0.1 | 28.5 | 3.0 | ||
| R+0 | 7.9 | 0.7 | 2.0 | 0.2 | 3.2 | 0.4 | 56.7 | 6.1 | ||
| R+1 | 6.4 | 0.5 | 2.2 | 0.2 | 3.0 | 0.3 | 57.4 | 4.6 | ||
| Time | Vitamin E (mg) | Vitamin B-12 (cobalamin) (µg) | Vitamin B-6 (mg) | Folate (vit B9) (µg) | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | |||
| BDC-4 | 23.9 | 2.7 | 3.9 | 0.5 | 2.1 | 0.2 | 485.8 | 44.1 | ||
| BDC-3 | 25.7 | 2.6 | 4.3 | 0.6 | 2.4 | 0.2 | 542.2 | 56.4 | ||
| BDC-2 | 25.2 | 3.1 | 3.6 | 0.3 | 2.0 | 0.2 | 315.0 | 21.4 | ||
| BDC-1 | 28.3 | 3.1 | 3.0 | 0.4 | 3.3 | 0.2 | 487.8 | 28.0 | ||
| DI-1 | 30.9 | 3.3 | 4.1 | 0.4 | 2.5 | 0.2 | 517.3 | 46.5 | ||
| DI-2 | 16.4 | 1.7 | 3.5 | 0.4 | 2.0 | 0.1 | 383.0 | 21.1 | ||
| DI-3 | 19.4 | 2.1 | 3.6 | 0.4 | 1.8 | 0.1 | 268.9 | 17.1 | ||
| DI-4 | 20.3 | 2.1 | 3.1 | 0.3 | 2.8 | 0.3 | 419.0 | 33.3 | ||
| DI-5 | 19.3 | 2.1 | 4.0 | 0.5 | 1.8 | 0.1 | 442.3 | 44.5 | ||
| R+0 | 25.7 | 2.6 | 4.3 | 0.5 | 2.4 | 0.2 | 553.5 | 63.8 | ||
| R+1 | 37.8 | 4.0 | 3.8 | 0.5 | 2.8 | 0.2 | 633.9 | 49.1 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jollet, M.; Nay, K.; Chopard, A.; Bareille, M.-P.; Beck, A.; Ollendorff, V.; Vernus, B.; Bonnieu, A.; Mariadassou, M.; Rué, O.; et al. Does Physical Inactivity Induce Significant Changes in Human Gut Microbiota? New Answers Using the Dry Immersion Hypoactivity Model. Nutrients 2021, 13, 3865. https://doi.org/10.3390/nu13113865
Jollet M, Nay K, Chopard A, Bareille M-P, Beck A, Ollendorff V, Vernus B, Bonnieu A, Mariadassou M, Rué O, et al. Does Physical Inactivity Induce Significant Changes in Human Gut Microbiota? New Answers Using the Dry Immersion Hypoactivity Model. Nutrients. 2021; 13(11):3865. https://doi.org/10.3390/nu13113865
Chicago/Turabian StyleJollet, Maxence, Kevin Nay, Angèle Chopard, Marie-Pierre Bareille, Arnaud Beck, Vincent Ollendorff, Barbara Vernus, Anne Bonnieu, Mahendra Mariadassou, Olivier Rué, and et al. 2021. "Does Physical Inactivity Induce Significant Changes in Human Gut Microbiota? New Answers Using the Dry Immersion Hypoactivity Model" Nutrients 13, no. 11: 3865. https://doi.org/10.3390/nu13113865
APA StyleJollet, M., Nay, K., Chopard, A., Bareille, M.-P., Beck, A., Ollendorff, V., Vernus, B., Bonnieu, A., Mariadassou, M., Rué, O., Derbré, F., Goustard, B., & Koechlin-Ramonatxo, C. (2021). Does Physical Inactivity Induce Significant Changes in Human Gut Microbiota? New Answers Using the Dry Immersion Hypoactivity Model. Nutrients, 13(11), 3865. https://doi.org/10.3390/nu13113865