
Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery


 , and
, and 
Abstract
:1. Introduction
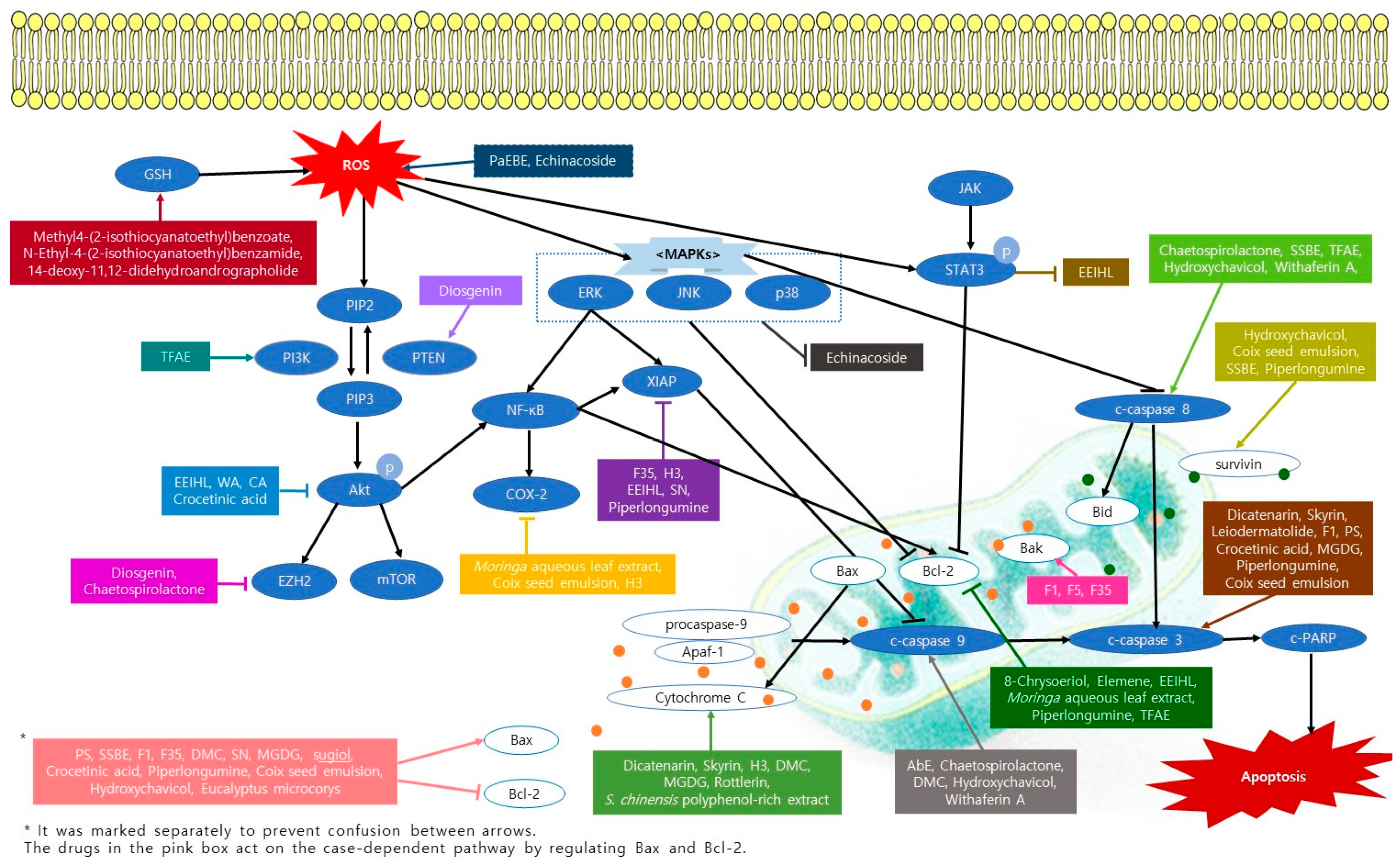
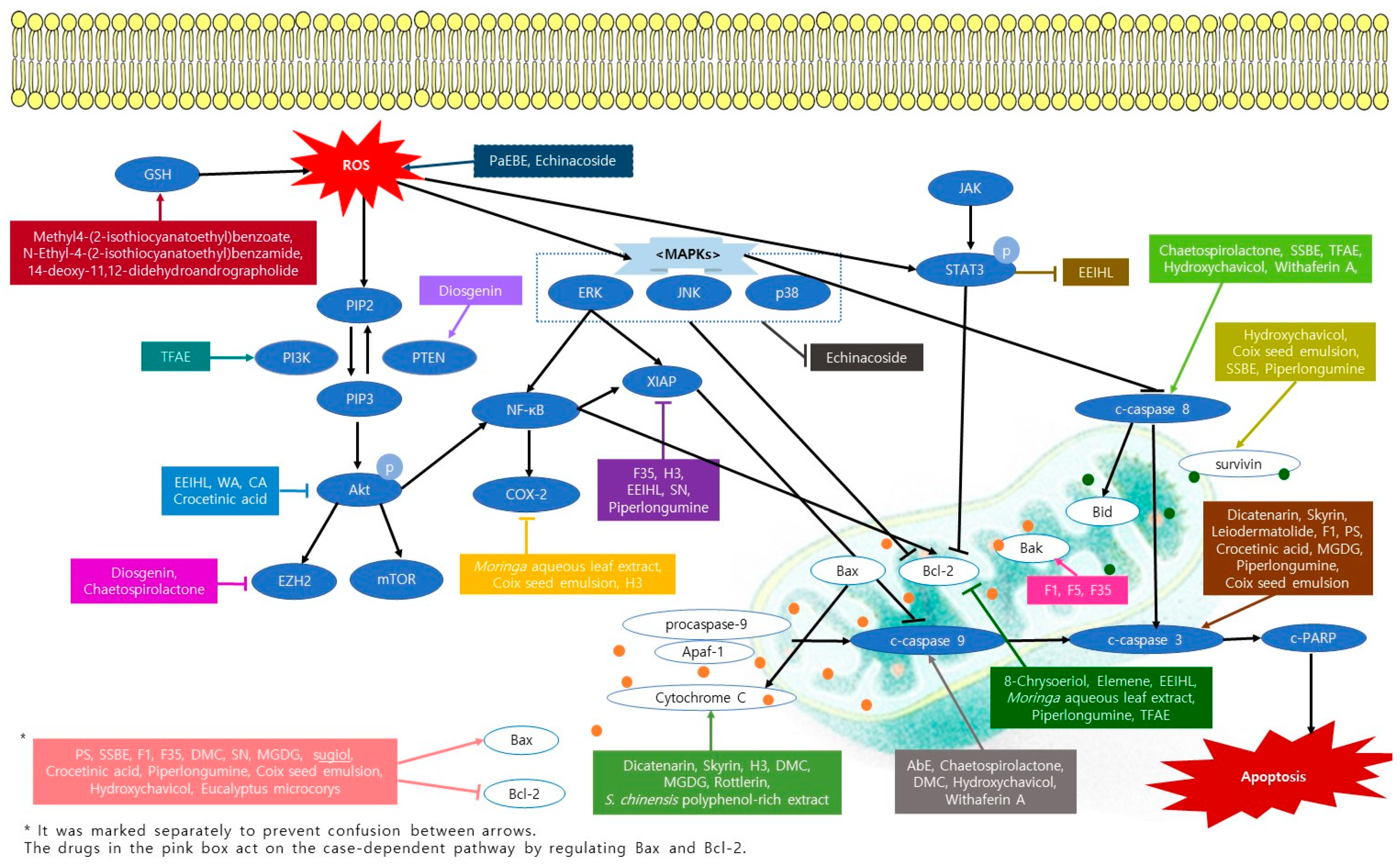
2. Apoptosis Inducing Natural Products
2.1. Apoptosis Inducing Fungi
2.2. Apoptosis Inducing Marine Sponge
2.3. Apoptosis Inducing Plants
2.3.1. Natural Compounds from Plants
2.3.2. Plants Extracts
2.3.3. Formulations

{kind=link}
{kind=link}
{kind=link}
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Plant | 14-deoxy-11,12-didehydroandrographolide | Andrographis paniculata | PANC-1, PSN-1 | 6.25, 12.5, 25, 50, 100 µM; 12, 16, 20, 24 h | Induction of apoptosis | ↓GSH | [28] |
| Plant | 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone (DMC) | Cleistocalyx operculatus buds | PANC-1 | 3, 10, 30 μM; 48 h | Induction of apoptosis | ↑Bax, cytochrome c, c-caspase-3, -9, c-PARP ↓Bcl-2 | [29] |
| Plant | 3,5-dihydroxy-6,7,8-trimethoxyflavone (flavone B) | Achyrocline bogotensis | Mia PaCa-2 | 40 μM; 6 h | Induction of apoptosis and cell cycle arrest | ↑p-ERK, p-c-JUN ↓pS6 | [30] |
| 5,7-dihydroxy-3,6,8-trimethoxyflavone (flavone A) | Gnaphalium elegans | PANC-28 | 40 μM; 6 h | ↓p-ERK, pS6, p-Bad, Bcl-xL, Bcl-2 | |||
| Plant | 8-Chrysoeriol | SW1990 | 50, 100 μM; 24 h | Induction of apoptosis | ↓Bcl-2 | [31] | |
| Plant | Cardiac glycosides | seed of Thevetia peruviana | SW1990 | Inhibition of proliferation of tumor cell lines | [32] | ||
| Plant | Carnosol | Rosmarinus officinali, Salvia carnosa, Origanum vulgare | AsPC-1 | 1 μM; 48 h | Induction of apoptosis | [49] | |
| Plant | Crocetinic acid | Crocus sativus | MIA PaCa-2 | 1, 10, 25, 50 µM; 72 h | Induction of apoptosis Inhibition of proliferation | ↑c-caspase 3, Bax ↓CD133, DCLK-1, Shh, PCNA, p-EGFR, p-AKTa, Bcl-2 | [33] |
| MIA PaCa-2 bearing athymic nude-mice | 0.5 mg/kg; 30 days | Inhibition of pancreatic cancer growth | ↑c-caspase 3 ↓PCNA, p-EGFR, p-AKT, Bcl-2 | ||||
| Plant | Diosgenin | Solanum, Dioscorea, Costus species | Patu8988, PANC-1 | 50, 75 μM; 48 h | Induction of apoptosis | ↑PTEN ↓EZH2, vimentin | [34] |
| PANC-1 bearing mice | 20 mg/kg; 4 weeks | Inhibition of tumor growth | ↑PTEN ↓EZH2, vimentin | ||||
| Plant | Echinacoside | Stems of Cistanchessalsa | SW1990 | 20, 50, 100 µM; 5 days | Induction of apoptosis | ↑ROS, Bax, p38 ↓MMP, JNK, ERK1/2 | [35] |
| Plant | Elemene | Zingiberaceae | BxPC-3, PANC-1 | 15, 30, 60 μg/mL; 12 h | Inhibition of cell proliferation, Induction of cell cycle arrest | [36] | |
| BxPC-3 bearing BALB/c mice | 20, 40, 60 mg/kg; 18 days | Inhibition of cell proliferation, Induction of cell cycle arrest | ↑p53 ↓Bcl-2 | ||||
| Plant | Grape seed proanthocyanidins (GSPs) | Vitis vinifera | PANC-1 | 20 µg/mL; 3, 12, 24 h | Induction of apoptosis | ↓miRNA-SS3, SS12, SS24 | [37] |
| ↓CDK6, EGFR, MSH6, DNMT1 | |||||||
| Plant | Hydroxychavicol | Piper betle | MIA PaCa-2 | 100 µM; 48 h | Induction of apoptosis | ↑c-caspase -8, -9, c-Bid | [38] |
| MIA PaCa-2, PANC-1 | 50, 100 µM; 48 h | ↑c-caspase-3, -8, -9, c-Bid, c-PARP, Bax ↓Bcl-2, survivin | |||||
| Plant | Hyperoside | MIA PaCa-2 | 50 μM; 48 h | Induction of caspase-dependent apoptosis | ↑c-caspase-3 | [39] | |
| Hypoxoside | INS-1 | 25 μM; 48 h | ↑c-caspase-3 | ||||
| Plant | Icariin | Herba Epimedii | PANC-2 | 100, 150, 200 μM; 48 h | Induction of apoptosis | [40] | |
| PANC-2 bearing C57BL/6 mice | 120 mg/kg; 11 days | Inhibition of pancreatic tumor progression | |||||
| Plant | Methyl4-(2-isothiocyanatoethyl)benzoate | Cruciferous vegetables | PANC-1 | 10 µM; 72 h | Induction of apoptosis | ↑ROS ↓GSH | [41] |
| N-Ethyl-4-(2-isothiocyanatoethyl)benzamide | |||||||
| Plant | Mastic gum resin | Pistacia atlantica | PANC-1 | 20, 40, 60, 80, 100 µg/mL; 72 h | Induction of cytotoxicity | [42] | |
| Plant | Monogalactosyl diacylglycerol | Spinach | MIA PaCa-2 | 25, 50, 75 μM; 24 h | Induction of apoptosis | ↑cytochrome c, c-PARP, Bax, c-caspase-3 ↓Bcl-2 | [43] |
| MIA PaCa-2 bearing BALB/cAJcl-nu/nu mice | 2 mg; 23 days | Inhibition of tumor growth | |||||
| Plant | Piperlongumine | Piper longum | BxPC-3, PANC-1, AsPC-1 | 5, 10, 20, 30, 40 μmol/L; 24, 48, 72 h | Induction of apoptosis Enhancement of gemcitabine-induced apoptosis | ↑procaspase-3, c-PARP ↓Bcl-2, Bcl-xL, survivin, XIAP | [44] |
| BxPC-3 bearing BALB/c mice | 10 mg/kg; 21 days | Inhibition of tumor growth Enhancement of gemcitabine-induced apoptosis | |||||
| Plant | Piperlongumine | Piper longum | PANC-1 | 5, 10, 15 μmol/mL; 24, 48 h | Induction of apoptosis, Inhibition of cell proliferation | ↓miR-27a, miR-17/miR-20a | [45] |
| ↑c-PARP, ROS | |||||||
| L3.6pL bearing athymic nu/nu mice | 30 mg/kg; 21 days | Inhibition of tumor growth | ↓miR-27a, miR-17/miR-20a | ||||
| ↓Sp1, Sp3, Sp4 | |||||||
| Plant | RN1 | Flower of Panax notoginseng | AsPC-1, BxPC-3 | 62.5, 125, 250, 500, 1000 μg/mL; 48 h | Inhibition of PDAC cell growth | ↓Galectin-3, EGFR, ERK, Runx1 | [46] |
| BxPC-3 bearing BALB/c nude mice | 0.5, 20 mg/kg; 46 days | ↓Galectin-3, Ki-67, EGFR, ERK, Runx1 | |||||
| Plant | Rottlerin | Mallotus phillippinensis | Patu8988 | 4 μM; 48 h | Induction of apoptosis | ↑cytochrome c ↓Skp2 | [47] |
| PANC-1 | 3 μM; 48 h | ||||||
| Plant | Sugiol | MIA PaCa-2 | 7.5, 15, 30 µM; 48 h | Induction of apoptosis and cell cycle arrest Increase of ROS production | ↑Bax ↓Bcl-2, MMP | [48] | |
| Plant | Withaferin A | Withania somnifora | AsPC-1 | 1 μM; 48 h | Induction of apoptosis | [49] | |
| Plant | Withaferin A | Withania somnifera | PANC-1, MIA PaCa-2 | 0.5, 1, 2.5, 5 μM; 24 h | Induction of apoptosis | ↑c-caspase-3, -8, -9, c-PARP1 | [50] |
| PANC-1 bearing BALB/c mice | 4 mg/kg; 24 days | Enhancement of the therapeutic efficacy of ER stress aggravators | |||||
| Plant | Bitter apricot ethanolic extract | Prunus armeniaca L. | PANC-1 | 704 μg/mL; 72 h | Induction of apoptosis | ↑Bax, caspase-3 ↓Bcl-2 | [51] |
| Plant | Clinacanthus nutans non-polar stem extracts (SN) and gemcitabine combination | Clinacanthus nutans | BxPC-3, SW1990 | 5 μg/mL (and/or 5 μg/mL of gemcitabine); 48 h | Induction of apoptosis Enhancement of gemcitabine-induced apoptosis | ↑Bax ↓Bcl-2, cIAP-2, XIAP | [52] |
| Plant | Coix seed emulsion | Coix lacryma-jobi | BxPC-3 | 1.50–10 mg/mL; 48 h | Induction of apoptosis | ↑caspase-3, c-PARP, Bax ↓Bcl-2, survivin, COX-2 | [53] |
| PANC-1 | 1.75–10 mg/mL; 48 h | ||||||
| AsPC-1 | 1.80–10 mg/mL; 48 h | ||||||
| BxPC-3 bearing nude BALB/c mice | 12.5 mL/kg; 24 days | ↓p65, Ki-67 | |||||
| Plant | Cordifoliketones A | Codonopsis cordifolioidea | AsPC-1, BxPC-3, PANC-1 | 2, 4, 6 µg/mL; 24, 48 h | Induction of apoptosis | ↑Bax, Bad, caspase-3, -8, -9 ↓Bcl-2, Bcl-xL | [54] |
| AsPC-1, BxPC-3, PANC-1 bearing BALB/c nude mice | 20, 80, 120, 240 M/kg; 27 days | ||||||
| Plant | Eucalyptus microcorys aqueous extract (F1) | Leaf of Eucalyptus microcorys | MIA PaCa-2 | 100, 150 μg/mL; 48 h | Induction of cell cycle arrest and apoptosis | ↑Bak, Bax, c-PARP, c- caspase-3 ↓Bcl-2, procaspase-3 | [55] |
| Plant | Eucalyptus microcorys leaf aqueous extract | Leaf of Eucalyptus microcorys | MIA PaCa-2, BxPC-3, CFPAC-1 | 50, 100 μg/mL; 24 h | Induction of apoptosis | [56] | |
| Eucalyptus microcorys leaf ethanolic extract | |||||||
| Eucalyptus microcorys fruit aqueous extract | Fruit of Eucalyptus microcorys | ||||||
| Eucalyptus saligna ethanolic extract | Eucalyptus saligna | ||||||
| Plant | Herbal mixture ethanol extract (H3) | Meliae Fructus, bark of Cinnamomum cassia, Sparganium rhizome | PANC-1 | 0.05 mg/mL; 72 h | Induction of apoptosis Induction of cell cycle arrest | ↑cytochrome c | [57] |
| ↓COX-2, CXCR4, JAK2, XIAP | |||||||
| PANC-1 bearing BALB/c nude mice | 200 mg/kg; 31 days | Inhibition of tumor growth | ↑cytochrome c ↓COX-2 | ||||
| Plant | Inula helenium L. ethyl acetate extract (EEIHL) | Inula helenium L. | CFPAC-1 | 2, 4, 6 µg/mL; 24 h | Inhibition of proliferation | ↑p-AKT, p-STAT3 | [58] |
| Inhibition of cell migration | ↑E-cadherin, c-PARP ↓Snail, XIAP, | ||||||
| Mitochondrial-dependent apoptosis | ↑Bim ↓Bcl-2, Mcl-1 | ||||||
| Plant | Moringa aqueous leaf extract | Moringa oleifera (Moringa) | PANC-1 | 1.8 mg/mL; 24 h | Induction of apoptosis | ↓Bcl-2, COX-2 | [59] |
| PANC-1 bearing CD-1 mice | 0.5, 1.0, 1.5 mg/g; 6 weeks | Inhibition of tumor growth | |||||
| Plant | Paeonia suffruticosa aqueous extracts (PS) | Paeonia suffruticosa | PANC-1, AsPC-1, BxPC-3 | 750 µg/mL; 72 h | Induction of autophagy | ↑caspase-3, -8, -9, c-caspase-3, DAPK3 | [60] |
| AsPC-1 bearing mice | 0.9, 1.8 g/kg; 21 days | Inhibition of cell cycle progression and cell migration | ↓Cyclin, CDK | ||||
| Plant | Pterospermum acerifolium ethanolic bark extract (PaEBE) | Bark of Pterospermum acerifolium | PANC-1 | 50, 75 µg/mL; 24 h | Induction of apoptosis | ↑ROS | [61] |
| Induction of mitochondrial-mediated cell death | ↓MMP | ||||||
| 50 µg/mL; 24 h | Induction of cell cycle arrest | ||||||
| Plant | Salvia chinensis polyphenol-rich extract | Salvia chinensis | MIA PaCa-2 | 20, 40, 60, 80 µg/mL; 48 h | Induction of apoptosis | ↑cytochrome c ↓MMP | [62] |
| Plant | Sedum sarmentosum Bunge extract (SSBE) | Sedum sarmentosum Bunge | PANC-1 | 100 µg/mL; 24 h | Induction of apoptosis | ↑Bax, Bad, caspase-3, -8, p53 | [63] |
| ↓Bcl-2, c-Myc, survivin | |||||||
| PANC-1 bearing BALB/c mice | 10, 100 mg/kg; 30 days | Inhibition of tumor growth | |||||
| Plant | Total flavonoid aglycones extract | Radix Scutellariae | BxPC-3 | 3.2, 6.4, 12.8 μg/mL; 24 h | Induction of apoptosis | ↑c-caspase-3, -8, c-PARP, c-Bid | [65] |
| BxPC-3-bearing BALB/c nu/nu mice | 50, 100, 150 mg/kg; 56 days | Induction of apoptosis and autophagy | ↑c-caspase-3, c-PARP, LC3-II ↓p62 | ||||
| Plant | F35 (alloalantolactone, alantolacton, isoalantolactone [1:5:4]) | Inula helenium L. | PANC-1, SW1990 | 8 µg/mL; 48 h | Inhibition of proliferation | [66] | |
| 6 µg/mL; 24 h | Induction of mitochondrion-related apoptosis | ↑Bak ↓Bcl-2, Mcl-1, XIAP | |||||
| 2, 4 µg/mL; 24 h | Inhibition of colony-formation and migration |
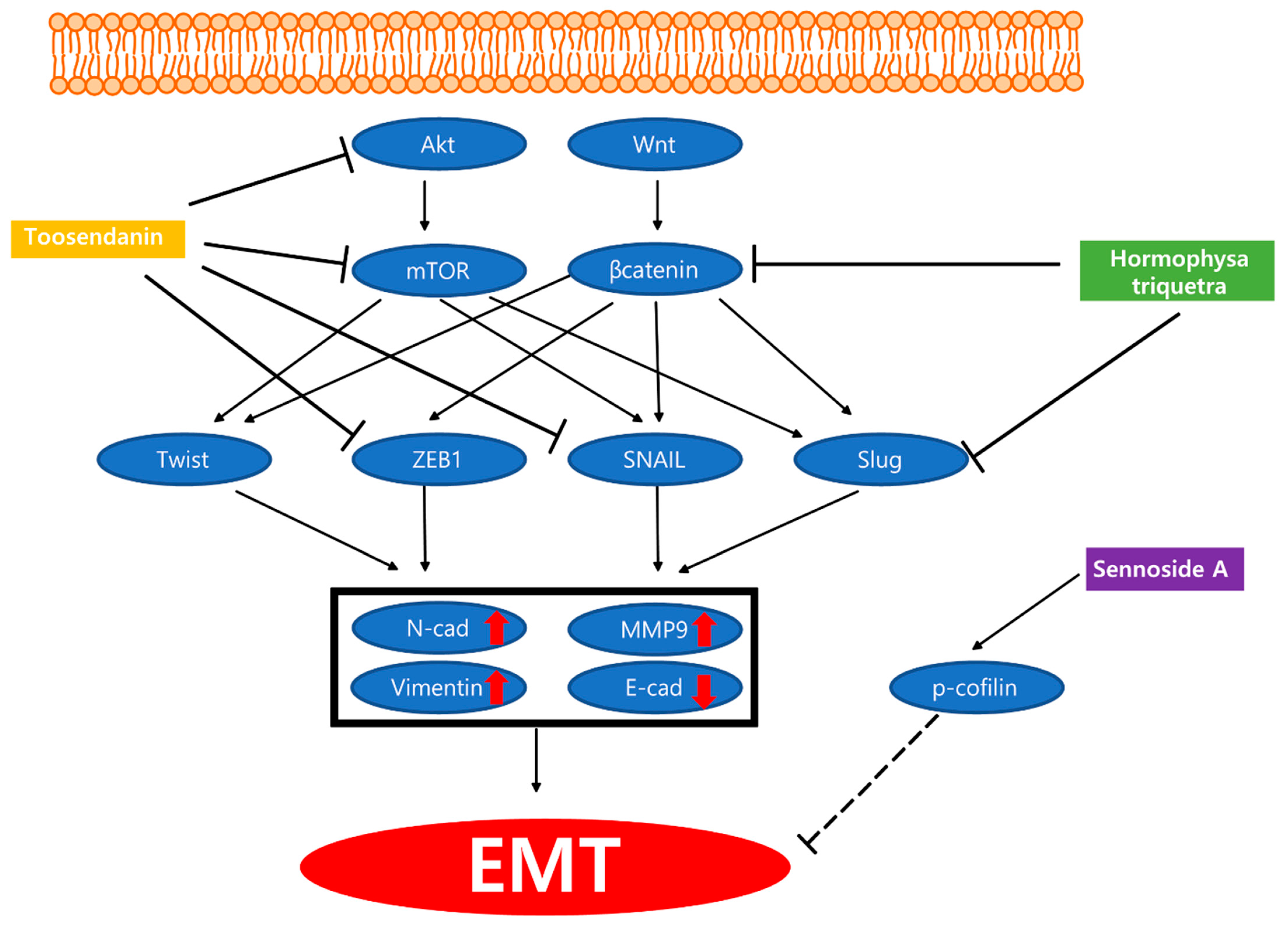
3. Metastasis Inhibiting Natural Products
4. Angiogenesis Inhibiting Natural Products
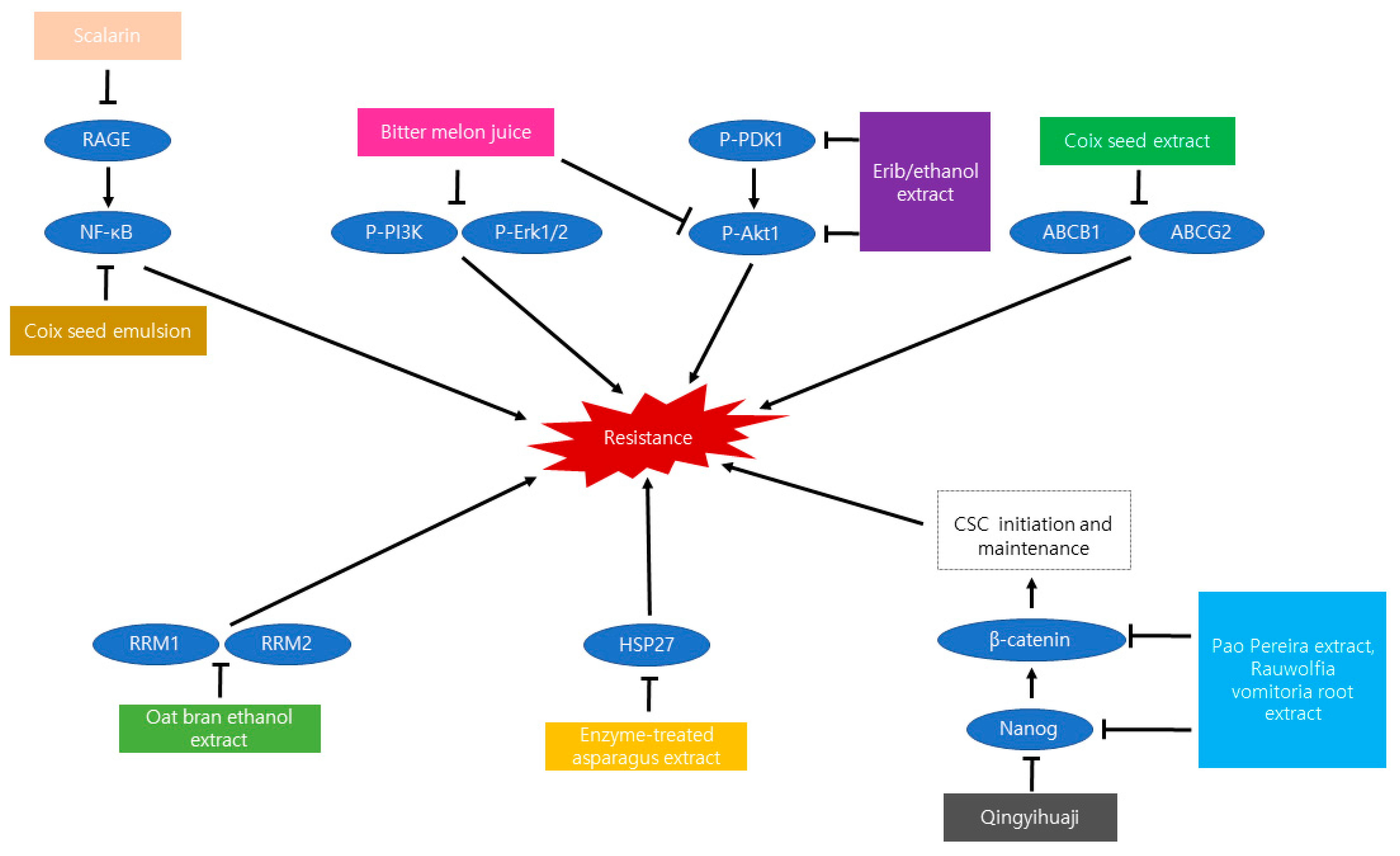
5. Resistance Inhibiting Natural Products
6. Clinical Trials
| Compound/Extract | Source | Phase | Patients | Status | Results | Registry Number | Reference |
|---|---|---|---|---|---|---|---|
| GBS-01 | Fruit of Arctium lappa | Phase 1 | Pancreatic Cancer (≥20 yrs) 15 participants | Completed | Safe up to 12 g per day Favorable response | UMIN000005787 | [86] |
| Curcumin | Curcuma longa Linn | Phase 2 | Pancreatic Cancer (≥18 yrs) 17 participants | Completed | Partial response 1 Stable disease 4 Tumor progression 6 Discontinued 5 Dose reduced 2 | NCT00192842 | [88] |
| Curcumin | Curcuma longa Linn | Phase 2 | Pancreatic Cancer (≥18 yrs) 50 participants | Completed | The six-month survival rate was 15.9% (7/44) | NCT00094445 | [89] |
| Huachansu/Bufo toad skin water extract | Bufo gargarizans | Phase 2 | Pancreatic Cancer (≥18 yrs) 80 participants | Completed | Median overall survival Experimental: 160 days Control: 156 days Not statistically significant (p = 0.339) | NCT00837239 | [90] |
| Kanglaite/oil extract | Coicis Semen | Phase 2 | Pancreatic Cancer (≥18 yrs) 85 participants | Completed | Progression-free survival Experimental: 112 days Control: 58 days | NCT00733850 | [92] |
| Iscador Qu Spzial | Viscum album | Phase 3 | Pancreatic Cancer (≥18 yrs) 434 patients | Completed | Mistletoe treatment improve the global health status significantly | ISRCTN70760582 | [94] |
| Iscador Qu/Mistletoe extract | Viscum album L. | Phase 3 | Pancreatic Cancer (≥18 yrs) 290 participants | Recruiting | N/A | NCT02948309 | [96] |
7. Discussion
7.1. Anti-Cancer Mechanisms of Natural Products
7.2. Clinical Trials of Natural Products against Pancreatic Cancer
7.3. Multi-Functional Natural Products
7.4. Anti-Inflammatory and Anti-Tumor Effects of Natural Products
7.5. miRNA Regulating Natural Products
7.6. Strengths and Limitations
7.7. Future Research Directions for Natural Product Treatment against Pancreatic Cancer
8. Methods
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| c-caspase | cleaved caspase |
| c-PARP | cleaved poly adenosine diphosphate ribose polymerase |
| Bax | Bcl-2-associated X protein |
| Bcl-family | B cell lymphoma-family |
| miRNA | microRNA |
| BRCA1/2 | breast cancer susceptibility gene 1/2 |
| PALB2 | partner and localizer of BRCA2 |
| TCM | Traditional Chinese Medicine |
| TTM | Traditional Thai Medicine |
| TKM | Tracitional Korean Medicine |
| CAM | Complementary and Alternative Medicine |
| PCD | programmed cell death |
| AbE | Agaricus blazei Murrill water extract |
| DAPK3 | death-associated protein kinase 3 |
| NLRP1 | NACHT, LRR, and PYD domains-containing protein 1 |
| TRAIL | tumor necrosis factor-related apoptosis inducing ligand |
| ROS | reactive oxygen species |
| MMP | mitochondrial membrane potential |
| compound 17 | 14-deoxy-11,12-didehydroandrographolide |
| DMC | 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone |
| flavone A | 5,7-dihydroxy-3,6,8-trimethoxyflavone |
| flavone B | 3,5-dihydroxy-6,7,8-trimethoxyflavone |
| PCNA | proliferating cell nuclear antigen |
| EGFR | epidermal growth factor receptor |
| AKT | protein kinase B (PKB) |
| EZH2 | enhancer of Zeste homolog 2 |
| ECH | Echinacoside |
| MAPK | mitogen-activated protein kinase |
| GSPs | grape seed proanthocyanidins |
| CDK6 | cell division protein kinase 6 |
| MSH6 | MutS homolog 6 |
| DNMT1 | DNA methyltransferase 1 |
| JNK | c-Jun N-terminal kinase |
| c-Bid | cleaved-Bcl-2 homology 3 interacting domain death agonist |
| GSH | Glutathione |
| MGR | mastic gum resin |
| MGDG | monogalactosyl diacylglycerol |
| XIAP | X-linked inhibitor of apoptosis |
| COX-2 | cyclooxygenase 2 |
| VEGF | vascular endothelial growth factor |
| MMP-9 | matrix metalloproteinase 9 |
| NF-κB | nuclear factor kappa B |
| RN1 | a polysaccharide from the flower of Panax notoginseng |
| Gal-3 | Galectin-3 |
| ERK | extracellular signal-regulated kinase |
| Runx1 | Runt-related transcription factor 1 |
| Skp2 | S-phase kinase associated protein 2 |
| WA | Withaferin A |
| CA | Carnosol |
| HGF | hepatocyte growth factor |
| c-Met | mesenchymal-epithelial transition factor |
| BAEE | Bitter apricot ethanolic extract |
| SN | non-polar stem extracts |
| cIAP-2 | cellular inhibitor of apoptosis 2 |
| CSE | coix seed emulsion |
| F1 | Eucalyptus microcorys aqueous extract |
| H3 | herbal mixture ethanol extract |
| CXCR-4 | C-X-C motif chemokine receptor 4 |
| JAK2 | Janus kinase 2 |
| RBC | red blood cell |
| EEIHL | ethyl acetate extract of Inula helenium L. |
| STAT3 | signal transducers and activators of transcription 3 |
| PS | Paeonia suffruticosa aqueous extracts |
| SSBE | the extract of Sedum sarmentosum Bunge |
| TFAE | total flavonoid aglycones extract |
| F35 | alloalantolactone, alantolacton, isoalantolactone [1:5:4] |
| PaEBE | Pterospermum acerifolium ethanolic bark extract |
| p-c-JUN | phospho-c-Jun |
| pS6 | phospho-S6 |
| DCLK-1 | doublecortin-like kinase 1 |
| Shh | sonic hedgehog signaling molecule |
| PTEN | phosphatase and tensin homolog |
| E-cadherin | epithelial cadherin |
| Mcl-1 | myeloid cell leukemia 1 |
| EMT | epithelial mesenchymal transition |
| CDC20 | cell division cycle protein 20 |
| CXCR-4 | C-X-C chemokine receptor type 4 |
| HT-EA | Hormophysa triquertra polyphenol |
| SSH | slingshot homologs |
| ZEB | zinc finger E-box-binding homeobox |
| mTOR | mammalian target of rapamycin |
| PRAS | proline rich protein |
| BAPX | bagpipe homeobox homolog |
| PhPT-1 | phosphohistidine phosphatase 1 |
| MEGF10 | multiple EGF-like domains 10 |
| DSGOST | Danggui-Sayuk-Ga-Osuyu-Saenggang-Tang |
| HUVECs | human umbilical vascular endothelial cells |
| HDMECs | human dermal microvascular endothelial cells |
| VEGFR2 | vascular endothelial growth factor 2 |
| p-FAK | phosphorylated focal adhesion kinase |
| p-IKKα/β | phosphorylated inhibitor of nuclear factor kappa-B kinaseα/β |
| RAGE | receptor for advanced glycation end products |
| ABC | ATP-binding cassette |
| ABCB1 | ATP-binding cassette B1 |
| ABCG2 | ATP-binding cassette G2 |
| ETAS | enzyme-treated asparagus extract |
| HSP | heat-shock protein |
| EriB | Eriocalyxin B |
| PDK1 | pyruvate dehydrogenase kinase 1 |
| OBE | Oat bran ethanol extract |
| RRM1 | ribonucleotide reductase subunit M1 |
| RRM2 | ribonucleotide reductase subunit M2 |
| AMPK | AMP-activated protein kinase |
| CSCs | cancer stem-like cells |
| EpCam+ | epithelial cell adhesion molecule+ |
| BCL2L2 | Bcl-2 like protein 2 |
| lncRNA | long non-coding RNAs |
| HPNE | human pancreatic epithelial nestin-expressing |
| CDk4 | cyclin-dependent kinase 4 |
| DLTs | dose-limiting toxicities |
| IQuS | Iscador Qu Spezial |
References
- Oberstein, P.E.; Olive, K.P. Pancreatic cancer: Why is it so hard to treat? Ther. Adv. Gastroenterol. 2013, 6, 321–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tempero, M.A. NCCN Guidelines Updates: Pancreatic Cancer. J. Natl. Compr. Canc. Netw. 2019, 17, 603–605. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic cancer: A review of clinical diagnosis, epidemiology, treatment and outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Lumlerdkij, N.; Tantiwongse, J.; Booranasubkajorn, S.; Boonrak, R.; Akarasereenont, P.; Laohapand, T.; Heinrich, M. Understanding cancer and its treatment in Thai traditional medicine: An ethnopharmacological-anthropological investigation. J. Ethnopharmacol. 2018, 216, 259–273. [Google Scholar] [CrossRef] [Green Version]
- Gokani, T. Ayurveda--the science of healing. Headache 2014, 54, 1103–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leem, K.-H.; Park, H.-K. Traditional Korean medicine: Now and the future. Neurol. Res. 2007, 29, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.M.; Komakech, R.; Karigar, C.S.; Saqib, A. Traditional Indian medicine (TIM) and traditional Korean medicine (TKM): Aconstitutional-based concept and comparison. Integr. Med. Res. 2017, 6, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Guo, Z.; Zhu, P.; Chen, J.; Huang, Y. Traditional Chinese medicine as a cancer treatment: Modern perspectives of ancient but advanced science. Cancer Med. 2019, 8, 1958–1975. [Google Scholar] [CrossRef] [PubMed]
- Owen, H.C.; Appiah, S.; Hasan, N.; Ghali, L.; Elayat, G.; Bell, C. Phytochemical Modulation of Apoptosis and Autophagy: Strategies to Overcome Chemoresistance in Leukemic Stem Cells in the Bone Marrow Microenvironment. Int. Rev. Neurobiol. 2017, 135, 249–278. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Li, T.; Fong, C.M.; Chen, X.; Chen, X.J.; Wang, Y.T.; Huang, M.Q.; Lu, J.J. Saponins from Chinese Medicines as Anticancer Agents. Molecules 2016, 21, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural compounds for cancer treatment and prevention. Pharm. Res 2009, 59, 365–378. [Google Scholar] [CrossRef]
- Abdel-Hafez, S.M.; Hathout, R.M.; Sammour, O.A. Attempts to enhance the anti-cancer activity of curcumin as a magical oncological agent using transdermal delivery. Adv. Tradit. Med. 2021, 21, 15–29. [Google Scholar] [CrossRef]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Shayan, P.; Goel, A.; Aggarwal, B.B.; Shakibaei, M. Resveratrol Chemosensitizes TNF-β-Induced Survival of 5-FU-Treated Colorectal Cancer Cells. Nutrients 2018, 10, 888. [Google Scholar] [CrossRef] [Green Version]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Kunnumakkara, A.B.; Aggarwal, B.B.; Shakibaei, M. Induction of the Epithelial-to-Mesenchymal Transition of Human Colorectal Cancer by Human TNF-β (Lymphotoxin) and its Reversal by Resveratrol. Nutrients 2019, 11, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toden, S.; Okugawa, Y.; Jascur, T.; Wodarz, D.; Komarova, N.L.; Buhrmann, C.; Shakibaei, M.; Boland, C.R.; Goel, A. Curcumin mediates chemosensitization to 5-fluorouracil through miRNA-induced suppression of epithelial-to-mesenchymal transition in chemoresistant colorectal cancer. Carcinogenesis 2015, 36, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Kim, B. Anti-Cancer Natural Products and Their Bioactive Compounds Inducing ER Stress-Mediated Apoptosis: A Review. Nutrients 2018, 10, 1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczanowski, S. Apoptosis: Its origin, history, maintenance and the medical implications for cancer and aging. Phys. Biol. 2016, 13, 031001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, R.S.Y. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. CR 2011, 30, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massagué, J.; Batlle, E.; Gomis, R.R. Understanding the molecular mechanisms driving metastasis. Mol. Oncol. 2017, 11, 3–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, N.; Bivona, T.G. Polytherapy and Targeted Cancer Drug Resistance. Trends Cancer 2019, 5, 170–182. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Matsushita, Y.; Furutani, Y.; Matsuoka, R.; Furukawa, T. Hot water extract of Agaricus blazei Murrill specifically inhibits growth and induces apoptosis in human pancreatic cancer cells. BMC Complement. Altern. Med. 2018, 18, 319. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Jia, X.; Gao, Y.; Zhang, Q. Chaetospirolactone reverses the apoptotic resistance towards TRAIL in pancreatic cancer. Biochem. Biophys. Res. Commun. 2018, 495, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Koul, M.; Meena, S.; Kumar, A.; Sharma, P.R.; Singamaneni, V.; Riyaz-Ul-Hassan, S.; Hamid, A.; Chaubey, A.; Prabhakar, A.; Gupta, P.; et al. Secondary Metabolites from Endophytic Fungus Penicillium pinophilum Induce ROS-Mediated Apoptosis through Mitochondrial Pathway in Pancreatic Cancer Cells. Planta Med. 2016, 82, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, D.; Sharma, N.; Singamaneni, V.; Sharma, V.; Kushwaha, M.; Abrol, V.; Guru, S.; Sharma, S.; Gupta, A.P.; Bhushan, S.; et al. Isolation and characterization of bioactive metabolites from Xylaria psidii, an endophytic fungus of the medicinal plant Aegle marmelos and their role in mitochondrial dependent apoptosis against pancreatic cancer cells. Phytomedicine 2016, 23, 1312–1320. [Google Scholar] [CrossRef]
- Guzman, E.A.; Xu, Q.; Pitts, T.P.; Mitsuhashi, K.O.; Baker, C.; Linley, P.A.; Oestreicher, J.; Tendyke, K.; Winder, P.L.; Suh, E.M.; et al. Leiodermatolide, a novel marine natural product, has potent cytotoxic and antimitotic activity against cancer cells, appears to affect microtubule dynamics, and exhibits antitumor activity. Int. J. Cancer 2016, 139, 2116–2126. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Morita, H.; Tezuka, Y. Preferentially cytotoxic constituents of Andrographis paniculata and their preferential cytotoxicity against human pancreatic cancer cell lines. Nat. Prod. Commun. 2015, 10, 1934578X1501000704. [Google Scholar] [CrossRef] [Green Version]
- Tuan, H.N.; Minh, B.H.; Tran, P.T.; Lee, J.H.; Oanh, H.V.; Ngo, Q.M.T.; Nguyen, Y.N.; Lien, P.T.K.; Tran, M.H. The Effects of 2′,4′-Dihydroxy-6′-methoxy-3′,5′-dimethylchalcone from Cleistocalyx operculatus Buds on Human Pancreatic Cancer Cell Lines. Molecules 2019, 24, 2538. [Google Scholar] [CrossRef] [Green Version]
- LeJeune, T.M.; Tsui, H.Y.; Parsons, L.B.; Miller, G.E.; Whitted, C.; Lynch, K.E.; Ramsauer, R.E.; Patel, J.U.; Wyatt, J.E.; Street, D.S. Mechanism of action of two flavone isomers targeting cancer cells with varying cell differentiation status. PLoS ONE 2015, 10, e0142928. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Min, Q.; Palida, A.; Zhang, Y.; Tang, R.; Chen, L.; Li, H. 8-Chrysoeriol, as a potential BCL-2 inhibitor triggers apoptosis of SW1990 pancreatic cancer cells. Bioorg. Chem. 2018, 77, 478–484. [Google Scholar] [CrossRef]
- Tian, D.M.; Cheng, H.Y.; Jiang, M.M.; Shen, W.Z.; Tang, J.S.; Yao, X.S. Cardiac Glycosides from the Seeds of Thevetia peruviana. J. Nat. Prod. 2016, 79, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Rangarajan, P.; Dharmalingam Subramaniam, S.P.; Kwatra, D.; Palaniyandi, K.; Islam, S.; Harihar, S.; Ramalingam, S.; Gutheil, W.; Putty, S.; Pradhan, R. Crocetinic acid inhibits hedgehog signaling to inhibit pancreatic cancer stem cells. Oncotarget 2015, 6, 27661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Chen, Y.; Gao, J.; Zhong, K.; Wei, H.; Li, K.; Tang, M.; Zhao, X.; Liu, X.; Nie, C.; et al. Diosgenin exhibits tumor suppressive function via down-regulation of EZH2 in pancreatic cancer cells. Cell Cycle 2019, 18, 1745–1758. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Luo, J.; Liang, Y.; Li, X. Echinacoside suppresses pancreatic adenocarcinoma cell growth by inducing apoptosis via the mitogen-activated protein kinase pathway. Mol. Med. Rep. 2016, 13, 2613–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.; Liu, Z.; Hui, L. Anti-tumor effect and mechanistic study of elemene on pancreatic carcinoma. BMC Complement. Altern. Med. 2019, 19, 133. [Google Scholar] [CrossRef]
- Wang, W.; Zhan, L.; Guo, D.; Xiang, Y.; Tian, M.; Zhang, Y.; Wu, H.; Wei, Y.; Ma, G.; Han, Z. Grape seed proanthocyanidins inhibit proliferation of pancreatic cancer cells by modulating microRNA expression. Oncol. Lett. 2019, 17, 2777–2787. [Google Scholar] [CrossRef] [Green Version]
- Guha Majumdar, A.; Subramanian, M. Hydroxychavicol from Piper betle induces apoptosis, cell cycle arrest, and inhibits epithelial-mesenchymal transition in pancreatic cancer cells. Biochem. Pharm. 2019, 166, 274–291. [Google Scholar] [CrossRef]
- Boukes, G.J.; van de Venter, M. The apoptotic and autophagic properties of two natural occurring prodrugs, hyperoside and hypoxoside, against pancreatic cancer cell lines. Biomed. Pharm. 2016, 83, 617–626. [Google Scholar] [CrossRef]
- Zheng, X.; Li, D.; Li, J.; Wang, B.; Zhang, L.; Yuan, X.; Li, C.; Cui, L.; Zhang, Q.; Yang, L.; et al. Optimization of the process for purifying icariin from Herba Epimedii by macroporous resin and the regulatory role of icariin in the tumor immune microenvironment. Biomed. Pharm. 2019, 118, 109275. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Wang, J.; Li, X.; Lu, W.; Yang, J.; Hu, Y.; Huang, P.; Wen, S. New mild and simple approach to isothiocyanates: A class of potent anticancer agents. Molecules 2017, 22, 773. [Google Scholar] [CrossRef] [Green Version]
- Rahman, H.S. Phytochemical analysis and antioxidant and anticancer activities of mastic gum resin from Pistacia atlantica subspecies kurdica. OncoTargets Ther. 2018, 11, 4559. [Google Scholar] [CrossRef] [Green Version]
- Akasaka, H.; Mizushina, Y.; Yoshida, K.; Ejima, Y.; Mukumoto, N.; Wang, T.; Inubushi, S.; Nakayama, M.; Wakahara, Y.; Sasaki, R. MGDG extracted from spinach enhances the cytotoxicity of radiation in pancreatic cancer cells. Radiat. Oncol. 2016, 11, 153. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, X.; Zhou, Y.; Jiang, H.; Pan, S.; Sun, B. Piperlongumine Suppresses Growth and Sensitizes Pancreatic Tumors to Gemcitabine in a Xenograft Mouse Model by Modulating the NF-kappa B Pathway. Cancer Prev. Res. 2016, 9, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Karki, K.; Hedrick, E.; Kasiappan, R.; Jin, U.-H.; Safe, S. Piperlongumine induces reactive oxygen species (ROS)-dependent downregulation of specificity protein transcription factors. Cancer Prev. Res. 2017, 10, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, P.; Qin, Y.; Cong, Q.; Shao, C.; Du, Z.; Ni, X.; Li, P.; Ding, K. RN1, a novel galectin-3 inhibitor, inhibits pancreatic cancer cell growth in vitro and in vivo via blocking galectin-3 associated signaling pathways. Oncogene 2017, 36, 1297–1308. [Google Scholar] [CrossRef]
- Su, J.; Wang, L.; Yin, X.; Zhao, Z.; Hou, Y.; Ye, X.; Zhou, X.; Wang, Z. Rottlerin exhibits anti-cancer effect through inactivation of S phase kinase-associated protein 2 in pancreatic cancer cells. Am. J. Cancer Res. 2016, 6, 2178–2191. [Google Scholar] [PubMed]
- Hao, C.; Zhang, X.; Zhang, H.; Shang, H.; Bao, J.; Wang, H.; Li, Z. Sugiol (127horbar;hydroxyabieta-8,11,13-trien-7-one) targets human pancreatic carcinoma cells (Mia-PaCa2) by inducing apoptosis, G2/M cell cycle arrest, ROS production and inhibition of cancer cell migration. J. BUON 2018, 23, 205–210. [Google Scholar] [PubMed]
- Aliebrahimi, S.; Kouhsari, S.M.; Arab, S.S.; Shadboorestan, A.; Ostad, S.N. Phytochemicals, withaferin A and carnosol, overcome pancreatic cancer stem cells as c-Met inhibitors. Biomed. Pharm. 2018, 106, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhu, F.; Jiang, J.; Sun, C.; Zhong, Q.; Shen, M.; Wang, X.; Tian, R.; Shi, C.; Xu, M.; et al. Simultaneous inhibition of the ubiquitin-proteasome system and autophagy enhances apoptosis induced by ER stress aggravators in human pancreatic cancer cells. Autophagy 2016, 12, 1521–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aamazadeh, F.; Ostadrahimi, A.; Rahbar Saadat, Y.; Barar, J. Bitter apricot ethanolic extract induces apoptosis through increasing expression of Bax/Bcl-2 ratio and caspase-3 in PANC-1 pancreatic cancer cells. Mol. Biol. Rep. 2020, 47, 1895–1904. [Google Scholar] [CrossRef]
- Hii, L.W.; Lim, S.E.; Leong, C.O.; Chin, S.Y.; Tan, N.P.; Lai, K.S.; Mai, C.W. The synergism of Clinacanthus nutans Lindau extracts with gemcitabine: Downregulation of anti-apoptotic markers in squamous pancreatic ductal adenocarcinoma. BMC Complement. Altern. Med. 2019, 19, 257. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Yang, B.; Xiong, Y.; Gu, M. Coix seed emulsion synergistically enhances the antitumor activity of gemcitabine in pancreatic cancer through abrogation of NF-κB signaling. Oncol. Rep. 2016, 36, 1517–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, Y.P.; Li, Q.F.; Wu, S.G.; Mao, D.C.; Deng, Y.Y.; Chen, R.W. Tsoong induces apoptosis and inhibits proliferation, migration and invasion of pancreatic ductal adenocarcinoma cells. Mol. Med. Rep. 2018, 17, 3527–3536. [Google Scholar] [CrossRef] [PubMed]
- Bhuyan, D.J.; Vuong, Q.V.; Bond, D.R.; Chalmers, A.C.; Bowyer, M.C.; Scarlett, C.J. Eucalyptus microcorys leaf extract derived HPLC-fraction reduces the viability of MIA PaCa-2 cells by inducing apoptosis and arresting cell cycle. Biomed. Pharm. 2018, 105, 449–460. [Google Scholar] [CrossRef]
- Bhuyan, D.J.; Sakoff, J.; Bond, D.R.; Predebon, M.; Vuong, Q.V.; Chalmers, A.C.; van Altena, I.A.; Bowyer, M.C.; Scarlett, C.J. In vitro anticancer properties of selected Eucalyptus species. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Pak, P.J.; Kang, B.H.; Park, S.H.; Sung, J.H.; Joo, Y.H.; Jung, S.H.; Chung, N. Antitumor effects of herbal mixture extract in the pancreatic adenocarcinoma cell line PANC1. Oncol. Rep. 2016, 36, 2875–2883. [Google Scholar] [CrossRef]
- Zhang, B.; Zeng, J.; Yan, Y.; Yang, B.; Huang, M.; Wang, L.; Zhang, Q.; Lin, N. Ethyl acetate extract from Inula helenium L. inhibits the proliferation of pancreatic cancer cells by regulating the STAT3/AKT pathway. Mol. Med. Rep. 2018, 17, 5440–5448. [Google Scholar] [CrossRef] [PubMed]
- Hagoel, L.; Vexler, A.; Kalich-Philosoph, L.; Earon, G.; Ron, I.; Shtabsky, A.; Marmor, S.; Lev-Ari, S. Combined Effect of Moringa oleifera and Ionizing Radiation on Survival and Metastatic Activity of Pancreatic Cancer Cells. Integr. Cancer Ther. 2019, 18, 1534735419828829. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-H.; Weng, Y.-P.; Tsai, H.-Y.; Chen, C.-J.; Lee, D.-Y.; Hsieh, C.-L.; Wu, Y.-C.; Lin, J.-Y. Aqueous extracts of Paeonia suffruticosa modulates mitochondrial proteostasis by reactive oxygen species-induced endoplasmic reticulum stress in pancreatic cancer cells. Phytomedicine 2018, 46, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.K.; Biswal, B.K. Pterospermum acerifolium (L.) wild bark extract induces anticarcinogenic effect in human cancer cells through mitochondrial-mediated ROS generation. Mol. Biol. Rep. 2018, 45, 2283–2294. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Huo, X.C.; Sun, F.D.; Dong, R.Q. Polyphenol-rich extract of Salvia chinensis exhibits anticancer activity in different cancer cell lines, and induces cell cycle arrest at the G(0)/G(1)-phase, apoptosis and loss of mitochondrial membrane potential in pancreatic cancer cells. Mol. Med. Rep. 2015, 12, 4843–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Chen, B.; Hong, W.; Liang, Y.; Zhou, M.; Zhou, L. Sedum sarmentosum Bunge extract induces apoptosis and inhibits proliferation in pancreatic cancer cells via the hedgehog signaling pathway. Oncol. Rep. 2016, 35, 2775–2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Cao, H.J.; Sun, S.J.; Dai, J.Y.; Fang, J.W.; Li, Q.H.; Yan, C.; Mao, W.W.; Zhang, Y.Y. Total flavonoid aglycones extract in Radix scutellariae inhibits lung carcinoma and lung metastasis by affecting cell cycle and DNA synthesis. J. Ethnopharmacol. 2016, 194, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, H.; Wang, J.; Chang, Q.; Hu, Z.; Shen, X.; Feng, J.; Zhang, Z.; Wu, X. Total flavonoid aglycones extract in Radix Scutellariae induces cross-regulation between autophagy and apoptosis in pancreatic cancer cells. J. Ethnopharmacol. 2019, 235, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhang, Q.; Zhang, B.; Yang, B.; Lin, N. Active ingredients of Inula helenium L. exhibits similar anti-cancer effects as isoalantolactone in pancreatic cancer cells. Nat. Prod. Res. 2019, 34, 2539–2544. [Google Scholar] [CrossRef]
- Steeg, P.S. Tumor metastasis: Mechanistic insights and clinical challenges. Nat. Med. 2006, 12, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Castillo, V.; Sliva, D. CDC20 associated with cancer metastasis and novel mushroom-derived CDC20 inhibitors with antimetastatic activity. Int. J. Oncol. 2019, 54, 2250–2256. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, W.; Zhou, Y.; Yang, J.; Xu, J.; Xu, Z.; Xu, B.; Yan, L.; Cheng, X.D.; Li, M.; et al. Terphenyllin Suppresses Orthotopic Pancreatic Tumor Growth and Prevents Metastasis in Mice. Front. Pharm. 2020, 11, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravindan, S.; Ramraj, S.; Kandasamy, K.; Thirugnanasambandan, S.S.; Somasundaram, D.B.; Herman, T.S.; Aravindan, N. Hormophysa triquerta polyphenol, an elixir that deters CXCR4- and COX2-dependent dissemination destiny of treatment-resistant pancreatic cancer cells. Oncotarget 2017, 8, 5717–5734. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Kim, W.; Lee, Y.G.; Kang, H.J.; Lee, S.H.; Park, S.Y.; Min, J.K.; Lee, S.R.; Chung, S.J. Identification of sennoside A as a novel inhibitor of the slingshot (SSH) family proteins related to cancer metastasis. Pharm. Res. 2017, 119, 422–430. [Google Scholar] [CrossRef]
- Pei, Z.; Fu, W.; Wang, G. A natural product toosendanin inhibits epithelial-mesenchymal transition and tumor growth in pancreatic cancer via deactivating Akt/mTOR signaling. Biochem. Biophys. Res. Commun. 2017, 493, 455–460. [Google Scholar] [CrossRef]
- Choi, H.S.; Lee, K.; Kim, M.K.; Lee, K.M.; Shin, Y.C.; Cho, S.G.; Ko, S.G. DSGOST inhibits tumor growth by blocking VEGF/VEGFR2-activated angiogenesis. Oncotarget 2016, 7, 21775–21785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.S.; Kim, M.K.; Lee, K.; Lee, K.M.; Choi, Y.K.; Shin, Y.C.; Cho, S.G.; Ko, S.G. SH003 represses tumor angiogenesis by blocking VEGF binding to VEGFR2. Oncotarget 2016, 7, 32969–32979. [Google Scholar] [CrossRef] [Green Version]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Qian, Y.; Xiong, Y.; Feng, D.; Wu, Y.; Zhang, X.; Chen, L.; Gu, M. Coix Seed Extract Enhances the Anti-Pancreatic Cancer Efficacy of Gemcitabine through Regulating ABCB1- and ABCG2-Mediated Drug Efflux: A Bioluminescent Pharmacokinetic and Pharmacodynamic Study. Int. J. Mol. Sci. 2019, 20, 5250. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Nanimoto, Y.; Baron, B.; Kitagawa, T.; Tokuda, K.; Kuramitsu, Y. Enzyme-treated Asparagus Extract Down-regulates Heat Shock Protein 27 of Pancreatic Cancer Cells. Vivo 2018, 32, 759–763. [Google Scholar] [CrossRef]
- Kim, M.; Mun, J.G.; Lee, H.J.; Son, S.R.; Lee, M.J.; Kee, J.Y. Inhibitory Effect of Oat Bran Ethanol Extract on Survival and Gemcitabine Resistance of Pancreatic Cancer Cells. Molecules 2019, 24, 3829. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhao, S.L.; Yue, G.G.L.; Wong, T.P.; Pu, J.X.; Sun, H.D.; Fung, K.P.; Leung, P.C.; Han, Q.B.; Lau, C.B.S.; et al. Isodon eriocalyx and its bioactive component Eriocalyxin B enhance cytotoxic and apoptotic effects of gemcitabine in pancreatic cancer. Phytomedicine 2018, 44, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Shapira, S.; Pleban, S.; Kazanov, D.; Tirosh, P.; Arber, N. Terpinen-4-ol: A Novel and Promising Therapeutic Agent for Human Gastrointestinal Cancers. PLoS ONE 2016, 11, e0156540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán, E.A.; Pitts, T.P.; Diaz, M.C.; Wright, A.E. The marine natural product Scalarin inhibits the receptor for advanced glycation end products (RAGE) and autophagy in the PANC-1 and MIA PaCa-2 pancreatic cancer cell lines. Investig. New Drugs 2019, 37, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Somasagara, R.R.; Deep, G.; Shrotriya, S.; Patel, M.; Agarwal, C.; Agarwal, R. Bitter melon juice targets molecular mechanisms underlying gemcitabine resistance in pancreatic cancer cells. Int. J. Oncol. 2015, 46, 1849–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, R.; Chen, P.; Chen, Q. Extract of the Medicinal Plant Pao Pereira Inhibits Pancreatic Cancer Stem-Like Cell In Vitro and In Vivo. Integr Cancer Ther. 2018, 17, 1204–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Wang, M.; Wang, C. Qingyihuaji formula reverses gemcitabine resistant human pancreatic cancer through regulate lncRNA AB209630/miR-373/EphB2-NANOG signals. Biosci. Rep. 2019, 39, BSR20190610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, R.; Chen, P.; Chen, Q. Inhibition of pancreatic cancer stem cells by Rauwolfia vomitoria extract. Oncol. Rep. 2018, 40, 3144–3154. [Google Scholar] [CrossRef] [PubMed]
- UMIN-CTR. Phase 1/2 Study of gbs-01 in Patients with Gemcitabine-Refractory Advanced Pancreatic Cancer. Available online: https://upload.umin.ac.jp/cgi-open-bin/ctr/ctr.cgi?function=brows&action=brows&recptno=R000006430&type=summary&language=J (accessed on 16 August 2020).
- Ikeda, M.; Sato, A.; Mochizuki, N.; Toyosaki, K.; Miyoshi, C.; Fujioka, R.; Mitsunaga, S.; Ohno, I.; Hashimoto, Y.; Takahashi, H. Phase I trial of GBS-01 for advanced pancreatic cancer refractory to gemcitabine. Cancer Sci. 2016, 107, 1818–1824. [Google Scholar] [CrossRef] [PubMed]
- clinicalTrials.gov. Gemcitabine with Curcumin for Pancreatic Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT00192842?term=NCT00192842&draw=2&rank=1 (accessed on 1 June 2020).
- ClinicalTrials.gov. Trial of Curcumin in Advanced Pancreatic Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT00094445?term=NCT00094445&draw=2&rank=1 (accessed on 1 June 2020).
- ClinicalTrials.gov. Huachansu & Gemcitabine in Pancreatic Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT00837239?term=NCT00837239&draw=2&rank=1 (accessed on 15 May 2020).
- Meng, Z.; Garrett, C.; Shen, Y.; Liu, L.; Yang, P.; Huo, Y.; Zhao, Q.; Spelman, A.; Ng, C.; Chang, D. Prospective randomised evaluation of traditional Chinese medicine combined with chemotherapy: A randomised phase II study of wild toad extract plus gemcitabine in patients with advanced pancreatic adenocarcinomas. Br. J. Cancer 2012, 107, 411–416. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Safety and Exploratory Efficacy of Kanglaite Injection in Pancreatic Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT00733850?term=NCT00733850&draw=2&rank=1 (accessed on 15 May 2020).
- Schwartzberg, L.S.; Arena, F.P.; Bienvenu, B.J.; Kaplan, E.H.; Camacho, L.H.; Campos, L.T.; Waymack, J.P.; Tagliaferri, M.A.; Chen, M.M.; Li, D. A randomized, open-label, safety and exploratory efficacy study of kanglaite injection (KLTi) plus gemcitabine versus gemcitabine in patients with advanced pancreatic cancer. J. Cancer 2017, 8, 1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISRCTNregistry. Mistletoe Therapy for Advanced Pancreatic Cancer. Available online: http://www.isrctn.com/ISRCTN70760582 (accessed on 1 June 2020).
- Tröger, W.; Galun, D.; Reif, M.; Schumann, A.; Stankovic, N.; Milicevic, M. Quality of life of patients with advanced pancreatic cancer during treatment with mistletoe: A randomized controlled trial. Dtsch. Ärzteblatt Int. 2014, 111, 493. [Google Scholar]
- clinicalTrials.gov. Mistletoe Therapy in Primary and Recurrent Inoperable Pancreatic Cancer (Mistral). Available online: https://clinicaltrials.gov/ct2/show/NCT02948309?term=Viscum+album&draw=3&rank=1 (accessed on 16 August 2020).
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 1–17. [Google Scholar]
- Qi, F.; Li, A.; Inagaki, Y.; Kokudo, N.; Tamura, S.; Nakata, M.; Tang, W. Antitumor activity of extracts and compounds from the skin of the toad Bufo bufo gargarizans Cantor. Int. Immunopharmacol. 2011, 11, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Enesel, M.B.; Acalovschi, I.; Grosu, V.; Sbarcea, A.; Rusu, C.; Dobre, A.; Weiss, T.; Zarkovic, N. Perioperative application of the Viscum album extract Isorel in digestive tract cancer patients. Anticancer Res. 2005, 25, 4583–4590. [Google Scholar]
- Tezuka, Y.; Yamamoto, K.; Awale, S.; Li, F.; Yomoda, S.; Kadota, S. Anti-austeric activity of phenolic constituents of seeds of Arctium lappa. Nat. Prod. Commun. 2013, 8, 1934578X1300800414. [Google Scholar] [CrossRef] [Green Version]
- Dhillon, N.; Aggarwal, B.B.; Newman, R.A.; Wolff, R.A.; Kunnumakkara, A.B.; Abbruzzese, J.L.; Ng, C.S.; Badmaev, V.; Kurzrock, R. Phase II trial of curcumin in patients with advanced pancreatic cancer. Clin. Cancer Res. 2008, 14, 4491–4499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udenigwe, C.C.; Ramprasath, V.R.; Aluko, R.E.; Jones, P.J. Potential of resveratrol in anticancer and anti-inflammatory therapy. Nutr. Rev. 2008, 66, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-L.; Lin, J.-K. Curcumin: A potential cancer chemopreventive agent through suppressing NF-κB signaling. J. Cancer Mol. 2008, 4, 11–16. [Google Scholar]
- Niu, Y.; Jin, Y.; Deng, S.-C.; Deng, S.-J.; Zhu, S.; Liu, Y.; Li, X.; He, C.; Liu, M.-L.; Zeng, Z. MiRNA-646-mediated reciprocal repression between HIF-1α and MIIP contributes to tumorigenesis of pancreatic cancer. Oncogene 2018, 37, 1743–1758. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Gao, G.; Zou, G.; Yu, H.; Zheng, X. Natural products as adjunctive treatment for pancreatic cancer: Recent trends and advancements. BioMed Res. Int. 2017, 2017, 8412508. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Classification | Compound/ Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Fungus | Agaricus blazei Murrill water extract | Agaricus blazei Murrill | MIA PaCa-2, PCI-35, PK-8 | 0.005, 0.015, 0.045%(w/v); 48 h | Induction of apoptosis | ↑c-caspase-3, -9, c-PARP | [23] |
| Fungus | Chaetospirolactone | Chaetomium sp. NF00754 | HPDE6c-7, AsPC-1, PANC-1 | 100 nM; 18 h | Induction of apoptosis | ↑c-caspase-3, -8, -9 ↓EZH2 | [24] |
| AsPC-1-bearing BALB/c mice | 0.075 mg/kg; 28 days | ↑c-caspase-3 | |||||
| Fungus | Dicatenarin | Penicillium pinophilum | MIA PaCa-2 | 20 µg/mL; 48 h | Induction of apoptosis | ↑cytochrome c, caspase-3 | [25] |
| Fungus | Skyrin | Penicillium pinophilum | MIA PaCa-2 | 50 µg/mL; 48 h | Induction of apoptosis | ↑cytochrome c, caspase-3 | |
| Fungus | Xylarione A | Xylaria psidii | MIA PaCa-2 | 10, 30, 50 µm; 24 h | Induction of apoptosis | ↓MMP | [26] |
| (-) 5-methylmellein |
| Classification | Compound/ Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Marine sponge | Leiodermatolide | Leiodermatium | AsPC-1, BxPC-3, MIA PaCa-2, PANC-1 | 10 nM; 24 h | Induction of apoptosis | ↑c-caspase-3 | [27] |
| L3.6pl cells bearing mice | 10 mg/kg; 3 weeks | Reduction of tumor weight |
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Fungus | Polyporenic acid | Poria cocos | PANC-1 | 30, 60 μM; 24 h | Inhibition of metastasis | ↓CDC20 | [68] |
| Fungus | Poria cocos EtOH extract | 30, 60 μg/mL; 24 h | |||||
| Fungus | Terphenyllin | PANC-1, HPAC | 25 μM; 24 h | Inhibition of invasion and migration | [69] | ||
| SCID mice bearing PANC1 orthotopic tumors | 20 mg/kg/day; 5 weeks | Inhibition of metastasis | |||||
| Plant | Cordifoliketones A | Codonopsis cordifolioidea | AsPC-1, BxPC-3, PANC-1 | 2, 4, 6 μg/mL; 12 h | Inhibition of invasion and migration | [54] | |
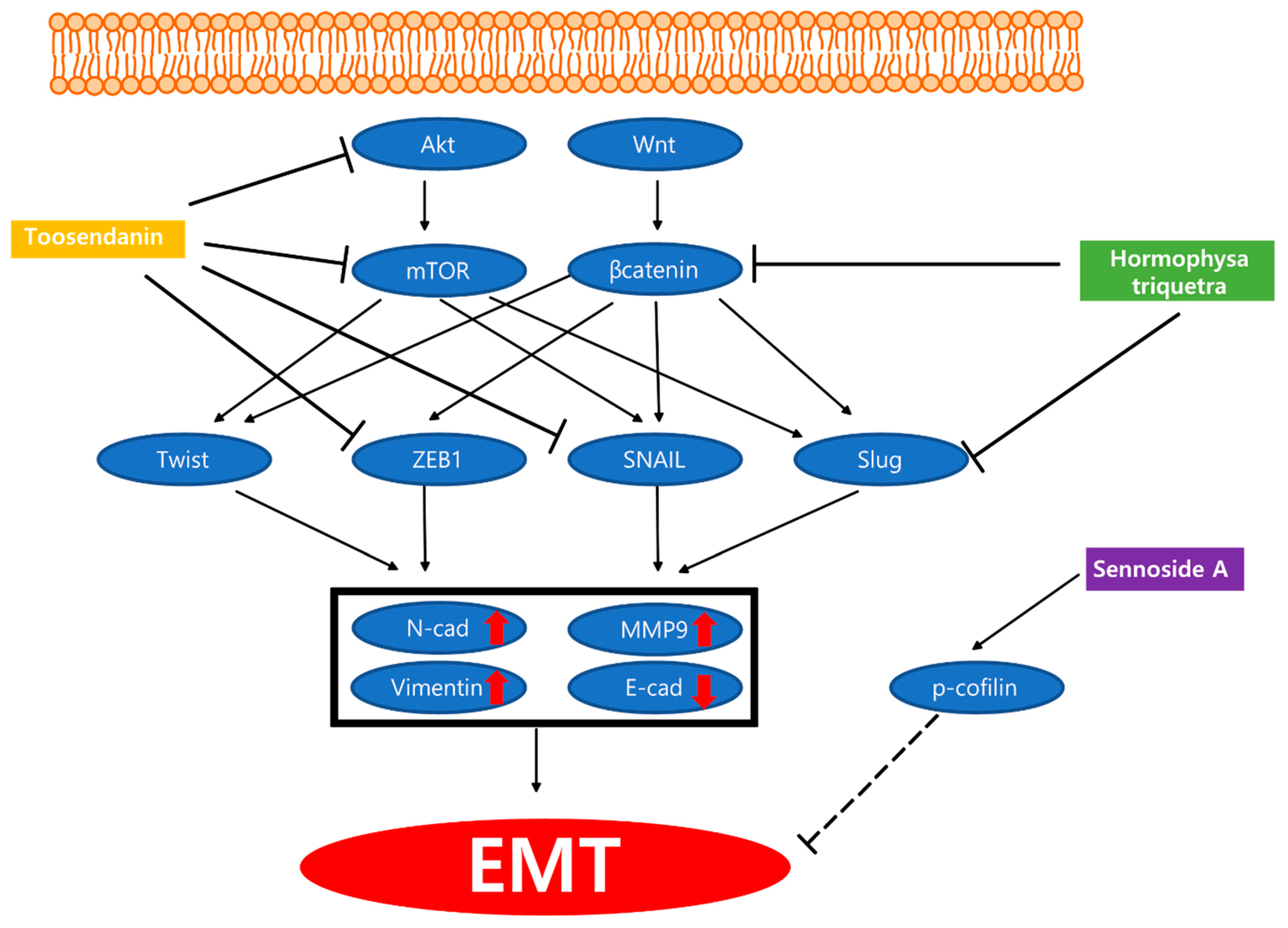
| Plant | Hormophysa triquetra polyphenol | Hormophysa triquertra | PANC-1, PANC-3.27, BxPC-3, MIA PaCa-2 | 100 µg/mL; 24 h | Inhibition of resistant cell migration/invasion | ↓CXCR4, COX-2, β-catenin, MMP-9, Ki-67, BAPX, PhPT-1, MEGF10 | [70] |
| MIA PaCa-2 bearing NCr-nu/nu nude mice | 10 mg/kg; 3 weeks | ||||||
| Plant | Moringa oleifera leaves water extract | Leaves of Moringa oleifera | PANC-1 | 0.4, 0.8, 1.8 mg/mL; 24 h, 2, 4 Gy radiation | Inhibition of metastasis | [59] | |
| Plant | Sennoside A | Rheum rhabarbarum | MIA PaCa-2, PANC-1 | 10 μmol/L; 24 h | Inhibition of invadopodia formation | ↑p-cofilin | [71] |
| PANC-1 | 10 μM; 20 m | Inhibition of invasion and migration | |||||
| PANC-1-Luc bearing BALB/c nu/nu mice | 10 mg/kg; 10 days | Inhibition of metastasis | |||||
| Plant | Toosendanin | AsPC-1, PANC-1 | 50, 100, 200 nM; 24 h | Inhibition of invasion and migration | ↑E-cadherin ↓Vimentin, ZEB1, Snail, p-AKT, p-PRAS40, p-mTOR, p-p70S6K | [72] | |
| PANC-1 bearing BALB/c mice | 0.2 mg/kg; 28 days | Inhibition of EMT | ↑E-cadherin ↓Vimentin, ZEB1, Snail |
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Plant | Danggui-Sayuk-Ga-Osuyu-Saenggang-Tang (DSGOST) | Angelica gigas,Cinnamomum cassia Blume, Paeonia lactiflora Pallas, Akebia quinata var. polyphylla Nak., Asarum sieboldii var. seoulense Nakai, Glycyrrhiza uralensis Fischer, Zizyphus jujuba var. inermis Rehder, Evodia rutaecarpa var. bodinieri Huang, Zingiber officinale Rosc. | HUVECs, HDMECs | 100 µg/mL; 72 h | Inhibition of migration Inhibition of tube formation | ↓p-VEGFR2, p-FAK, p-SRC, p-AKT, p-IKKα/β, p-IκBα, p-NF-κB, MMP-9 | [73] |
| PANC-28 bearing BALB/c nude mice | 20 mg/kg; 49 days | Inhibition of angiogenesis | ↑c-caspase-3 ↓Ki-67, p-VEGFR2, MMP-9 | ||||
| 100 µg; 0.5 h | |||||||
| Plant | SH003 | Astragalus membranaceus, Angelica gigas, Trichosanthes Kirilowii Maximowicz | HUVECs | 10, 20, 50 µg/mL; 24 h | Inhibition of angiogenesis | ↑c-caspase-3 ↓p-VEGFR2, MMP-9, p-FAK, p-SRC, p-ERK, p-AKT, p-STAT3 | [74] |
| PANC-28 bearing BALB/c nude mice | 2 mg/kg; 49 days | ↑c-caspase-3 ↓Ki-67, p-VEGFR2, MMP-9 | |||||
| 20 µg; 0.5 h |
| Classification | Compound/Extract | Source | Cell Line/Animal Model | Dose; Duration | Efficacy | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| Plant | Terpinen-4-ol | COLO357, PANC-1, MIA-PaCa | 0.005, 0.01, 0.05, 0.1%; 72 h | Inhibition of tumor growth Sensitization of gemcitabine | [80] | ||
| Animal | Scalarin | Euryspongia cf. rosea | PANC-1, MIA PaCa-2 | 10 µg/mL; 24 h | Inhibition of autophagy | ↓RAGE | [81] |
| Plant | Bitter melon juice | Momordica charantia | AsPC-1 | 1–4%; 24, 48 h | Inhibition of viability | ↓p-AKT, p-ERK1/2, p-PI3K, p-PTEN | [82] |
| Plant | Coix seed emulsion | Coix lachryma-jobi | PANC-1 | 4.0 mg/mL; 72 h | Sensitization of gemcitabine | ↑caspase-3, c-PARP, Bax ↓NF-κB, Bcl-2, survivin, COX-2 | [53] |
| BxPC-3 bearing BALB/c nude mice | 12.5 mL/kg; 24 days | ||||||
| Plant | Coix seed extract | Coix lachryma-jobi | BxPC-3, PANC-1 | 10 mg/mL; 24, 48 h | Sensitization of gemcitabine | ↓ABCB1, ABCG2 | [76] |
| BxPC-3 bearing BALB/c nude mice | 12.5 mL/kg; 3 weeks | ||||||
| Plant | Enzyme-treated asparagus extract | KLM1-R | 2 mg/mL; 120 h | Sensitization of gemcitabine | ↓HSP27, p-HSP27 | [77] | |
| Plant | EriB/ethanol extract | Isodon eriocalyx | SW1990 | 2.5 µM; 24 h | Sensitization of gemcitabine | ↑c-caspase 3, c-PARP, p-JNK ↓p-PDK1, p-AKT1 | [79] |
| Plant | Oat bran ethanol extract | Avena sativa L. | PANC-1, MIA PaCa-2 | 40 µg/mL; 72 h | Sensitization of gemcitabine | ↑p-AMPK, p21, p27 ↓p-JNK, cyclin D1, CDk4, RRM1, RRM2 | [78] |
| Plant | Pao Pereira extract | Geissospermum vellosii | PANC-1, MIA PaCa-2 | 50, 100 µg/mL; 48 h | Inhibition of tumor sphenoid formation Reduction of pancreatic CSCs | ↓ CD44, CD24, EpCam, Nanog, β-catenin, BCL2L2, COX-2 | [83] |
| PANC-1 bearing nude mice. | 20 mg/kg; 3 weeks | Reduction of pancreatic CSCs | |||||
| Plant | Qingyihuaji | Herba Scutellariae barbatae, Herba Hedyotdis, Rhizoma Arisaematis erubescentis, Herba seu Radix Gynostemmatis pentaphylli, Fructus Amomi Rotundus | CFPAC-1 | 40 μg/L; 24, 48, 72h | Sensitization of gemcitabine Inhibition of proliferation Decrease of migration | ↑lncRNA AB209630 ↓miR-373, EphB2, Nanog | [84] |
| CFPAC-1 bearing nude mice | 40 g/kg; 28 days | Sensitization of gemcitabine Inhibition of proliferation | |||||
| Plant | Rauwolfia vomitoria root extract | Rauwolfia vomitoria | PANC-1, MIA PaCa-2 | 50, 100, 200 µg/mL; 48 h | Inhibition of tumor spheroid formation Reduction of pancreatic CSCs | ↓CD24, EpCam, Nanog, β-catenin | [85] |
| PANC-1 bearing athymic NCr-nu/numice | 20 mg/kg; 5 week | Reduction of tumorigenicity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, A.; Ha, J.; Kim, J.; Cho, Y.; Ahn, J.; Cheon, C.; Kim, S.-H.; Ko, S.-G.; Kim, B. Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery. Nutrients 2021, 13, 3801. https://doi.org/10.3390/nu13113801
Kim A, Ha J, Kim J, Cho Y, Ahn J, Cheon C, Kim S-H, Ko S-G, Kim B. Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery. Nutrients. 2021; 13(11):3801. https://doi.org/10.3390/nu13113801
Chicago/Turabian StyleKim, Ahyeon, Jiwon Ha, Jeongeun Kim, Yongmin Cho, Jimyung Ahn, Chunhoo Cheon, Sung-Hoon Kim, Seong-Gyu Ko, and Bonglee Kim. 2021. "Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery" Nutrients 13, no. 11: 3801. https://doi.org/10.3390/nu13113801
APA StyleKim, A., Ha, J., Kim, J., Cho, Y., Ahn, J., Cheon, C., Kim, S.-H., Ko, S.-G., & Kim, B. (2021). Natural Products for Pancreatic Cancer Treatment: From Traditional Medicine to Modern Drug Discovery. Nutrients, 13(11), 3801. https://doi.org/10.3390/nu13113801