
The Controversial Role of Adiponectin in Appetite Regulation of Animals


{kind=link}
{kind=link}
{kind=link}
{kind=link}
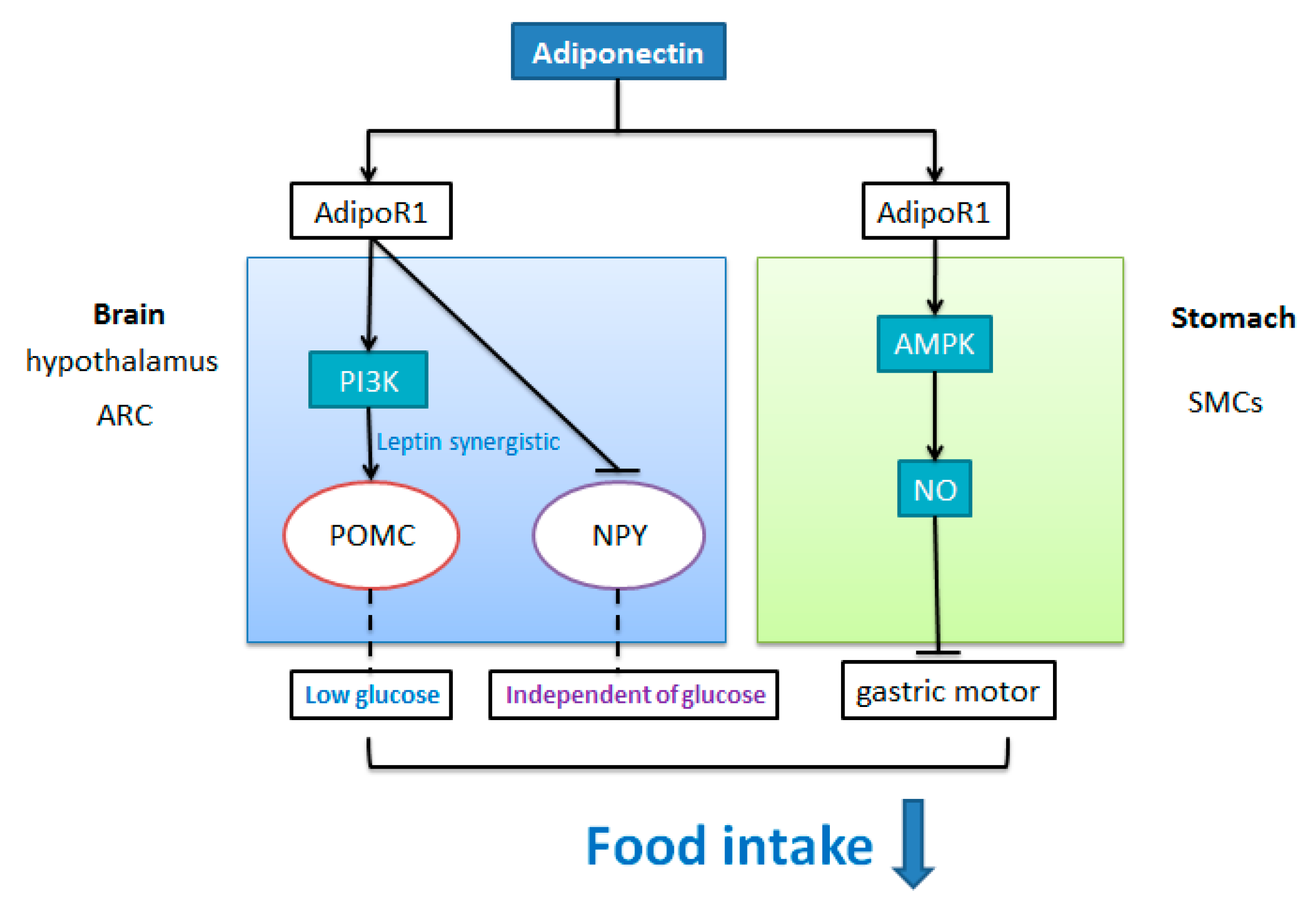
Abstract
1. Introduction
2. Discovery
3. Structure
4. Tissue Distribution
5. Receptors
6. The Effect of Feeding and Fasting on AdipoQ Expression
7. The Effect of Macronutrients on AdipoQ Expression
8. The Effect of AdipoQ on the Regulation of Appetite
8.1. The Anorexigenic Role of AdipoQ
8.1.1. Central Effect
8.1.2. Peripheral Effect
8.2. The Orexigenic Role of AdipoQ
8.2.1. Central Effect
8.2.2. Peripheral Effect
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agüera, Z.; Lozano-Madrid, M.; Mallorquí-Bagué, N.; Jiménez-Murcia, S.; Menchón, J.M.; Fernández-Aranda, F. A review of binge eating disorder and obesity. Neuropsychiatrie 2021, 35, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Hay, P.; Mitchison, D. Eating Disorders and Obesity: The Challenge for Our Times. Nutrients 2019, 11, 1055. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Klockars, A.; Levine, A.S.; Olszewski, P.K. Hypothalamic Integration of the Endocrine Signaling Related to Food Intake. Curr. Top. Behav. Neurosci. 2018, 43, 239–269. [Google Scholar] [CrossRef]
- Thomas, M.A.; Xue, B. Mechanisms for AgRP neuron-mediated regulation of appetitive behaviors in rodents. Physiol. Behav. 2018, 190, 34–42. [Google Scholar] [CrossRef]
- Lenard, N.R.; Berthoud, H.-R. Central and Peripheral Regulation of Food Intake and Physical Activity: Pathways and Genes. Obesity 2008, 16, S11–S22. [Google Scholar] [CrossRef] [PubMed]
- Lehr, S.; Hartwig, S.; Lamers, D.; Famulla, S.; Müller, S.; Hanisch, F.-G.; Cuvelier, C.; Ruige, J.; Eckardt, K.; Ouwens, M.; et al. Identification and Validation of Novel Adipokines Released from Primary Human Adipocytes. Mol. Cell. Proteom. 2012, 11, 111 010504. [Google Scholar] [CrossRef]
- Berg, A.H.; Combs, T.P.; Du, X.; Brownlee, M.; Scherer, P.E. The adipocyte-secreted protein Acrp30 enhances hepatic insulin action. Nat. Med. 2001, 7, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Shimomura, I.; Kishida, K.; Nishizawa, H.; Matsuda, M.; Nagaretani, H.; Furuyama, N.; Kondo, H.; Takahashi, M.; Arita, Y.; et al. Diet-induced insulin resistance in mice lacking adiponectin/ACRP30. Nat. Med. 2002, 8, 731–737. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Waki, H.; Terauchi, Y.; Kubota, N.; Hara, K.; Mori, Y.; Ide, T.; Murakami, K.; Tsuboyama-Kasaoka, N.; et al. The Fat-Derived Hormone Adiponectin Reverses Insulin Resistance As-sociated with Both Lipoatrophy and Obesity. Nat. Med. 2001, 7, 941–946. [Google Scholar] [CrossRef]
- Matsuda, M.; Shimomura, I.; Sata, M.; Arita, Y.; Nishida, M.; Maeda, N.; Kumada, M.; Okamoto, Y.; Nagaretani, H.; Nishizawa, H. Role of Adiponectin in Preventing Vascular Stenosis the Missing Link of Adipo-Vascular Axis. J. Biol. Chem. 2002, 277, 37487–37491. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef]
- Xiangdong, W.; Motoshima, H.; Mahadev, K.; Stalker, T.J.; Scalia, R.; Goldstein, B.J. In-volvement of Amp-Activated Protein Kinase in Glucose Uptake Stimulated by the Globular Domain of Adiponectin in Primary Rat Adipocytes. Diabetes 2003, 52, 1355–1363. [Google Scholar]
- Pandey, G.K.; Vadivel, S.; Raghavan, S.; Mohan, V.; Balasubramanyam, M.; Gokulakrishnan, K. High molecular weight adiponectin reduces glucolipotoxicity-induced inflammation and improves lipid metabolism and insulin sensitivity via APPL1-AMPK-GLUT4 regulation in 3T3-L1 adipocytes. Atherosclerosis 2019, 288, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Choi, R.S.; Lim, J.H.; Kim, M.Y.; Kim, E.N.; Kim, Y.; Choi, B.S.; Kim, Y.S.; Kim, H.W.; Lim, K.M.; Kim, M.J.; et al. Adi-ponectin Receptor Agonist Adiporon Decreased Ceramide, and Lipotoxicity, and Ameliorated Diabetic Nephropathy. Metabolism 2018, 85, 348–360. [Google Scholar] [CrossRef]
- Tagami, T.; Satoh, N.; Usui, T.; Yamada, K.; Shimatsu, A.; Kuzuya, H. Adiponectin in Anorexia Nervosa and Bulimia Nervosa. J. Clin. Endocrinol. Metab. 2004, 89, 1833–1837. [Google Scholar] [CrossRef]
- Khalil, R.B.; El Hachem, C. Adiponectin in eating disorders. Eat. Weight. Disord. Stud. Anorexia Bulim. Obes. 2013, 19, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Housova, J.; Anderlova, K.; Krizová, J.; Haluzikova, D.; Kremen, J.; Kumstyrová, T.; Papezová, H.; Haluzik, M. Serum Adi-ponectin and Resistin Concentrations in Patients with Restrictive and Binge/Purge Form of Anorexia Nervosa and Bulimia Nervosa. J. Clin. Endocrinol. Metab. 2005, 90, 1366–1370. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Monteleone, P.; Fabrazzo, M.; Martiadis, V.; Fuschino, A.; Serritella, C.; Milici, N.; Maj, M. Opposite Changes in Circulating Adiponectin in Women With Bulimia Nervosa or Binge Eating Disorder. J. Clin. Endocrinol. Metab. 2003, 88, 5387–5391. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Shklyaev, S.; Aslanidi, G.; Tennant, M.; Prima, V.; Kohlbrenner, E.; Kroutov, V.; Camp-bell-Thompson, M.; Crawford, J.; Shek, E.W.; Scarpace, P.J. Sustained Peripheral Expression of Transgene Adi-ponectin Offsets the Development of Diet-Induced Obesity in Rats. Proc. Natl. Acad. Sci. USA 2003, 100, 14217–14222. [Google Scholar] [CrossRef]
- Suyama, S.; Maekawa, F.; Maejima, Y.; Kubota, N.; Kadowaki, T.; Yada, T. Glucose level determines excitatory or inhibitory effects of adiponectin on arcuate POMC neuron activity and feeding. Sci. Rep. 2016, 6, 30796. [Google Scholar] [CrossRef] [PubMed]
- Kubota, N.; Yano, W.; Kubota, T.; Yamauchi, T.; Itoh, S.; Kumagai, H.; Kozono, H.; Takamoto, I.; Okamoto, S.; Shiuchi, T.; et al. Adiponectin Stimulates AMP-Activated Protein Kinase in the Hypothalamus and Increases Food Intake. Cell Metab. 2007, 6, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Takahashi, N.; Hileman, S.M.; Patel, H.R.; Berg, A.H.; Pajvani, U.B.; Scherer, P.E.; Ahima, R.S. Adiponectin Acts in the Brain to Decrease Body Weight. Nat. Med. 2004, 10, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Scherer, P.E.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. A Novel Serum Protein Similar to C1q, Produced Exclusively in Adipocytes. J. Biol. Chem. 1995, 270, 26746–26749. [Google Scholar] [CrossRef]
- Hu, E.; Liang, P.; Spiegelman, B.M. AdipoQ Is a Novel Adipose-specific Gene Dysregulated in Obesity. J. Biol. Chem. 1996, 271, 10697–10703. [Google Scholar] [CrossRef]
- Maeda, K.; Okubo, K.; Shimomura, I.; Funahashi, T.; Matsuzawa, Y.; Matsubara, K. cDNA Cloning and Expression of a Novel Adipose Specific Collagen-like Factor, apM1 (AdiposeMost Abundant Gene Transcript 1). Biochem. Biophys. Res. Commun. 1996, 221, 286–289. [Google Scholar] [CrossRef]
- Nakano, Y.; Tobe, T.; Choi-Miura, N.-H.; Mazda, T.; Tomita, M. Isolation and Characterization of GBP28, a Novel Gelatin-Binding Protein Purified from Human Plasma. J. Biochem. 1996, 120, 803–812. [Google Scholar] [CrossRef]
- Arita, Y.; Kihara, S.; Ouchi, N.; Takahashi, M.; Maeda, K.; Miyagawa, J.; Hotta, K.; Shimomura, I.; Nakamura, T.; Miyaoka, K. Paradoxical Decrease of an Adipose-Specific Protein, Adiponectin, in Obesity. Biochem. Biophys. Res. Commun. 1999, 425, 560–564. [Google Scholar] [CrossRef]
- Ding, T.S.; Liu, B.H.; Ko, Y.H. Cloning and Expression of Porcine Adiponectin and Adiponectin Receptor 1 and 2 Genes in Pigs. J. Anim. Sci. 2004, 82, 3162–3174. [Google Scholar] [CrossRef]
- Onay-Besikci, A.; Altarejos, J.Y.; Lopaschuk, G.D. Gad-Globular Head Domain of Adiponectin Increases Fatty Acid Oxidation in Newborn Rabbit Hearts. J. Biol. Chem. 2004, 279, 44320–44326. [Google Scholar] [CrossRef]
- Maddineni, S.; Metzger, S.; Ocón, O.; Hendricks, G., III; Ramachandran, R. Adiponectin Gene Is Expressed in Multiple Tissues in the Chicken: Food Deprivation Influences Adiponectin Messenger Ribonucleic Acid Expression. Endocrinology 2005, 146, 4250–4256. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Dong, B.; Zhang, Y.; Yu, J.; Meng, H.; Gong, D.; Gu, Z. Cloning and Prokaryotic Expression of Full Length Cdna of Duck Adiponectin Gene. J. Anim. Husb. Vet. Med. 2010, 41, 1232–1239. [Google Scholar]
- Xu, G.-Q.; Gong, D.-Q.; Chu, D.-S.; Biao, D.; He, M. Cloning and tissue expression of the adiponectin gene in geese. Chin. J. Agric. Biotechnol. 2009, 6, 91–96. [Google Scholar] [CrossRef]
- Nishio, I.S.; Gibert, Y.; Bernard, L.; Brunet, F.; Triqueneaux, G.; Laudet, V. Adiponectin and Adiponectin Receptor Genes Are Coexpressed During Zebrafish Embryogenesis and Regulated by Food Deprivation. Developmental Dynamics An. Off. Publ. Am. Assoc. Anat. 2008, 237, 1682. [Google Scholar]
- Kondo, H.; Suga, R.; Suda, S.; Nozaki, R.; Hirono, I.; Nagasaka, R.; Kaneko, G.; Ushio, H.; Watabe, S. EST analysis on adipose tissue of rainbow trout Oncorhynchus mykiss and tissue distribution of adiponectin. Gene 2011, 485, 40–45. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Xin-Jiang, L.; Jiong, C. Full-Length and a Smaller Globular Fragment of Adi-ponectin Have Opposite Roles in Regulating Monocyte/Macrophage Functions in Ayu, Plecoglossus Altivelis. Fish Shellfish. Immunol. 2018, 82, 319–329. [Google Scholar]
- Ji, R.; Xu, X.; Xiang, X.; Zhu, S.; Li, Y.; Mai, K.; Ai, Q. Regulation of adiponectin on lipid metabolism in large yellow croaker (Larimichthys crocea). Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158711. [Google Scholar] [CrossRef]
- Dai, M.; Xia, T.; Zhang, G.; Chen, X.; Gan, L.; Feng, S.; Qiu, H.; Peng, Y.; Yang, Z. Cloning, expression and chromosome localization of porcine adiponectin and adiponectin receptors genes. Domest. Anim. Endocrinol. 2006, 30, 117–125. [Google Scholar] [CrossRef]
- Fruebis, J.; Tsao, T.S.; Javorschi, S.; Ebbets-Reed, D.; Erickson, M.R.; Yen, F.T.; Bihain, B.E.; Lodish, H.F. Proteolytic Cleavage Product of 30-Kda Adipocyte Complement-Related Protein Increases Fatty Acid Oxidation in Muscle and Causes Weight Loss in Mice. Proc. Natl Acad. Sci. USA 2001, 98, 2005–2010. [Google Scholar] [CrossRef]
- Waki, H.; Yamauchi, T.; Kamon, J.; Ito, Y.; Uchida, S.; Kita, S.; Hara, K.; Hada, Y.; Vasseur, F.; Froguel, P.; et al. Impaired Multimerization of Human Adiponectin Mutants Associated with Diabetes. Molecular Structure and Multimer Formation of Adiponectin. J. Biol. Chem. 2003, 278, 40352–40363. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, A.; Knight, C.; Xu, L.Y.; Cooper, G. Hydroxylation and Glycosylation of the Four Conserved Lysine Residues in the Collagenous Domain of Adiponectin. Potential Role in the Modulation of Its Insulin-Sensitizing Activity. J. Biol. Chem. 2002, 277, 19521–19529. [Google Scholar] [CrossRef]
- Wang, Y.; Lam, K.S.L.; Yau, M.-H.; Xu, A. Post-translational modifications of adiponectin: Mechanisms and functional implications. Biochem. J. 2008, 409, 623–633. [Google Scholar] [CrossRef]
- Liu, M.; Liu, F. Regulation of adiponectin multimerization, signaling and function. Best Pr. Res. Clin. Endocrinol. Metab. 2014, 28, 25–31. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Zhang, Z.; Huang, B.; Cheng, X.; Wang, D.; la Gahu, Z.; Xue, Z.; Da, Y.; Li, D. Adiponectin-Derived Active Peptide Adp355 Exerts Anti-Inflammatory and Anti-Fibrotic Activities in Thioacetamide-Induced Liver Injury. Sci. Rep. 2016, 6, 19445. [Google Scholar] [CrossRef]
- Kumar, P.; Smith, T.; Rahman, K.; Thorn, N.E.; Anania, F.A. Adiponectin Agonist Adp355 Attenuates Ccl4-Induced Liver Fibrosis in Mice. PLoS ONE 2014, 9, e110405. [Google Scholar] [CrossRef]
- Lyzogubov, V.V.; Tytarenko, R.G.; Bora, N.S.; Bora, P.S. Inhibitory role of adiponectin peptide I on rat choroidal neovascularization. Biochim. Biophys. Acta Bioenerg. 2012, 1823, 1264–1272. [Google Scholar] [CrossRef]
- Otvos, L., Jr.; Haspinger, E.; La Russa, F.; Maspero, F.; Graziano, P.; Kovalszky, I.; Lovas, S.; Nama, K.; Hoffmann, R.; Knappe, D.; et al. Design and development of a peptide-based adiponectin receptor agonist for cancer treatment. BMC Biotechnol. 2011, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Saxena, N.K.; Anania, F.A. Adipocytokines and hepatic fibrosis. Trends Endocrinol. Metab. 2015, 26, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tan, B.; Karteris, E.; Zervou, S.; Digby, J.; Hillhouse, E.W.; Vatish, M.; Randeva, H.S. Secretion of adiponectin by human placenta: Differential modulation of adiponectin and its receptors by cytokines. Diabetologia 2006, 49, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-Y.; Hong, X.; Cao, Q.-Q.; So, K.-F. Adiponectin, exercise and eye diseases. Int. Rev. Neurobiol. 2019, 147, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Liu, W.; Liu, Z.; Li, N. cDNA cloning, genomic structure, chromosomal mapping and expression analysis of ADIPOQ (adiponectin) in chicken. Cytogenet. Genome Res. 2005, 112, 148–151. [Google Scholar] [CrossRef]
- Chabrolle, C.; Tosca, L.; Crochet, S.; Tesseraud, S.; Dupont, J. Expression of adiponectin and its receptors (AdipoR1 and AdipoR2) in chicken ovary: Potential role in ovarian steroidogenesis. Domest. Anim. Endocrinol. 2007, 33, 480–487. [Google Scholar] [CrossRef]
- A Hadley, J.; Ocon-Grove, O.M.; Ramachandran, R. Adiponectin is secreted by theca layer cells isolated from chicken ovarian follicles. Reproduction 2020, 159, 275–288. [Google Scholar] [CrossRef]
- Sánchez-Gurmaches, J.; Cruz-Garcia, L.; Gutiérrez, J.; Navarro, I. Adiponectin effects and gene expression in rainbow trout: An in vivo and in vitro approach. J. Exp. Biol. 2012, 215, 1373–1383. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Ito, Y.; Tsuchida, A.; Yokomizo, T.; Kita, S.; Sugiyama, T.; Miyagishi, M.; Hara, K.; Tsunoda, M.; et al. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 2003, 423, 762–769. [Google Scholar] [CrossRef]
- Vasiliauskaite-Brooks, I.; Sounier, R.; Rochaix, P.; Bellot, G.; Fortier, M.; Hoh, F.; De Colibus, L.; Bechara, C.; Saied, E.M.; Arenz, C.; et al. Structural insights into adiponectin receptors suggest ceramidase activity. Nat. Cell Biol. 2017, 544, 120–123. [Google Scholar] [CrossRef]
- Wang, L.; Xue, K.; Wang, Y.; Niu, L.; Zhang, H. Molecular and Functional Characterization of the Adiponectin (Adipoq) Gene in Goat Skeletal Muscle Satellite Cells. Asian Australas. J. Anim. Sci. 2018, 31, 1088–1097. [Google Scholar] [CrossRef]
- Guillod-Maximin, E.; Roy, A.F.; Vacher, C.M.; Aubourg, A.; Bailleux, V.; Lorsignol, A.; Pénicaud, L.; Parquet, M.; Taouis, M. Adiponectin receptors are expressed in hypothalamus and colocalized with proopiomelanocortin and neuropeptide Y in rodent arcuate neurons. J. Endocrinol. 2008, 200, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Hug, C.; Wang, J.; Ahmad, N.S.; Bogan, J.S.; Tsao, T.-S.; Lodish, H.F. T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc. Natl. Acad. Sci. USA 2004, 101, 10308–10313. [Google Scholar] [CrossRef] [PubMed]
- Denzel, S.M.; Scimia, M.C.; Zumstein, P.M.; Walsh, K.; Ruiz-Lozano, P.; Ranscht, B. T-Cadherin Is Critical for Adiponectin-Mediated Cardioprotection in Mice. J. Clin. Investig. 2010, 120, 4342–4352. [Google Scholar] [CrossRef] [PubMed]
- Fujishima, Y.; Maeda, N.; Matsuda, K.; Masuda, S.; Mori, T.; Fukuda, S.; Sekimoto, R.; Yamaoka, M.; Obata, Y.; Kita, S.; et al. Adiponectin association with T-cadherin protects against neointima proliferation and atherosclerosis. FASEB J. 2017, 31, 1571–1583. [Google Scholar] [CrossRef]
- Akins, M.R.; Benson, D.L.; Greer, C.A. Cadherin expression in the developing mouse olfactory system. J. Comp. Neurol. 2007, 501, 483–497. [Google Scholar] [CrossRef]
- Crosson, M.S.; Marques, A.; Dib, P.; Dotson, C.D.; Munger, S.D.; Zolotukhin, S. Taste Receptor Cells in Mice Express Receptors for the Hormone Adiponectin. Chem. Senses 2019, 44, 409–422. [Google Scholar] [CrossRef]
- Michalak, M.; Groenendyk, J.; Szabo, E.; Gold, L.I.; Opas, M. Calreticulin, a multi-process calcium-buffering chaperone of the endoplasmic reticulum. Biochem. J. 2009, 417, 651–666. [Google Scholar] [CrossRef]
- Takemura, Y.; Ouchi, N.; Shibata, R.; Aprahamian, T.; Kirber, M.T.; Summer, R.S.; Kihara, S.; Walsh, K. Adiponectin Modulates Inflammatory Reactions Via Calreticulin Receptor-Dependent Clearance of Early Apoptotic Bodies. J. Clin. Investig. 2007, 117, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.G.; Zahradka, P.; Clark, J. Exploring the Cardio-metabolic Relevance of T-cadherin: A Pleiotropic Adiponectin Receptor. Endocrine Metab. Immune Disord. Drug Targets 2017, 17, 200–206. [Google Scholar] [CrossRef]
- Tanaka, Y.; Kita, S.; Nishizawa, H.; Fukuda, S.; Fujishima, Y.; Obata, Y.; Nagao, H.; Masuda, S.; Nakamura, Y.; Shimizu, Y.; et al. Adiponectin Promotes Muscle Regeneration through Binding to T-Cadherin. Sci. Rep. 2019, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Kita, S.; Fukuda, S.; Maeda, N.; Shimomura, I. Native adiponectin in serum binds to mammalian cells expressing T-cadherin, but not AdipoRs or calreticulin. eLife 2019, 8, 48675. [Google Scholar] [CrossRef] [PubMed]
- Rohde, K.; Keller, M.; Horstmann, A.; Liu, X.; Eichelmann, F.; Stumvoll, M.; Villringer, A.; Kovacs, P.; Tönjes, A.; Böttcher, Y. Role of genetic variants in ADIPOQ in human eating behavior. Genes Nutr. 2014, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Dastani, Z.; Hivert, M.-F.; Timpson, N.; Perry, J.R.B.; Yuan, X.; Scott, R.A.; Henneman, P.; Heid, I.M.; Kizer, J.R.; Lyytikäinen, L.-P.; et al. Novel Loci for Adiponectin Levels and Their Influence on Type 2 Diabetes and Metabolic Traits: A Multi-Ethnic Meta-Analysis of 45,891 Individuals. PLoS Genet. 2012, 8, e1002607. [Google Scholar] [CrossRef]
- Ma, W.; Huang, T.; Heianza, Y.; Wang, T.; Sun, D.; Tong, J.; Williamson, D.A.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic variations of circulating adiponectin levels modulate changes in appetite in response to weight-loss diets. J. Clin. Endocrinol. Metab. 2016, 102, 316–325. [Google Scholar] [CrossRef]
- Turyn, J.; Korczynska, J.; Presler, M.; Stelmanska, E.; Goyke, E.; Swierczynski, J. Up-regulation of rat adipose tissue adiponectin gene expression by long-term but not by short-term food restriction. Mol. Cell. Biochem. 2008, 312, 185–191. [Google Scholar] [CrossRef]
- Polito, R.; Di Meo, I.; Barbieri, M.; Daniele, A.; Paolisso, G.; Rizzo, M.R. Adiponectin Role in Neurodegenerative Diseases: Focus on Nutrition Review. Int. J. Mol. Sci. 2020, 21, 9255. [Google Scholar] [CrossRef]
- Janiszewska, J.; Ostrowska, J.; Szostak-Węgierek, D. The Influence of Nutrition on Adiponectin—A Narrative Review. Nutrients 2021, 13, 1394. [Google Scholar] [CrossRef]
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.; Bragazzi, N. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Goto, A.; Morita, A.; Deura, K.; Sasaki, S.; Aiba, N.; Shimbo, T.; Terauchi, Y.; Miyachi, M.; Noda, M.; et al. Low-molecular-weight adiponectin and high-molecular-weight adiponectin levels in relation to diabetes. Obesity 2013, 22, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Kestin, M.; Schwarz, Y.; Yang, P.; Hu, X.; Lampe, J.W.; Kratz, M. A Low-Fat High-Carbohydrate Diet Reduces Plasma Total Adiponectin Concentrations Compared to a Moderate-Fat Diet with No Impact on Biomarkers of Systemic Inflammation in a Randomized Controlled Feeding Study. Eur. J. Nutr. 2016, 55, 237–246. [Google Scholar] [CrossRef]
- Zhang, D.-M.; Jiao, R.-Q.; Kong, L.-D. High Dietary Fructose: Direct or Indirect Dangerous Factors Disturbing Tissue and Organ Functions. Nutrition 2017, 9, 335. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, R.; Cianflone, K.; McGahan, J.P.; Berglund, L.; Bremer, A.A.; Keim, N.L.; Griffen, S.C.; Havel, P.J.; Stanhope, K.L. Effects of sugar-sweetened beverages on plasma acylation stimulating protein, leptin and adiponectin: Relationships with Metabolic Outcomes. Obesity 2013, 21, 2471–2480. [Google Scholar] [CrossRef]
- Magalhães, C.G.; Feitoza, F.M.; Moreira, S.B.; Carmo, A.V.; Souto, F.J.; Reis, S.R.; Martins, M.S.; da Silva, M.H.G. Hy-poadiponectinaemia in Nonalcoholic Fatty Liver Disease Obese Women Is Associated with Infrequent Intake of Dietary Sucrose and Fatty Foods. J. Hum. Nutr. Diet. 2014, 27, 301–312. [Google Scholar] [CrossRef]
- Shih, P.-H.; Shiue, S.-J.; Chen, C.-N.; Cheng, S.-W.; Lin, H.-Y.; Wu, L.-W.; Wu, M.-S. Fucoidan and Fucoxanthin Attenuate Hepatic Steatosis and Inflammation of NAFLD through Modulation of Leptin/Adiponectin Axis. Mar. Drugs 2021, 19, 148. [Google Scholar] [CrossRef]
- Sánchez, D.; Miguel, M.; Aleixandre, A. Dietary Fiber, Gut Peptides, and Adipocytokines. J. Med. Food 2012, 15, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Alessa, H.B.; Ley, S.; Rosner, B.; Malik, V.S.; Willett, W.C.; Campos, H.; Hu, F.B. High Fiber and Low Starch Intakes Are Associated with Circulating Intermediate Biomarkers of Type 2 Diabetes among Women. J. Nutr. 2016, 146, 306–317. [Google Scholar] [CrossRef]
- E Kasim-Karakas, S.; Tsodikov, A.; Singh, U.; Jialal, I. Responses of inflammatory markers to a low-fat, high-carbohydrate diet: Effects of energy intake. Am. J. Clin. Nutr. 2006, 83, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; You, G.-P.; Cui, F.; Chen, L.-F.; Yang, X.-J.; Chen, L.-G.; Lu, H.-D.; Zhang, W.-Q. Effects of a low-fat diet on the hepatic expression of adiponectin and its receptors in rats with NAFLD. Ann. Hepatol. 2015, 14, 108–117. [Google Scholar] [CrossRef]
- Barbosa, M.M.; Melo, A.L.; Damasce, N.R. The Benefits of Ω-3 Supplementation Depend on Adiponectin Basal Level and Adiponectin Increase after the Supplementation: A Randomized Clinical Trial. Nutrition 2017, 34, 7–13. [Google Scholar] [CrossRef]
- Prates, R.E.; Beretta, M.V.; Nascimento, F.V.; Bernaud, F.R.; de Almeira, J.C.; Rodrigues, T.C. Saturated fatty acid intake decreases serum adiponectin levels in subjects with type 1 diabetes. Diabetes Res. Clin. Pract. 2016, 116, 205–211. [Google Scholar] [CrossRef]
- Garcia-Macedo, R.; Sanchez-Muñoz, F.; Almanza-Perez, J.C.; Duran-Reyes, G.; Alarcon-Aguilar, F.; Cruz, M. Glycine In-creases Mrna Adiponectin and Diminishes Pro-Inflammatory Adipokines Expression in 3t3-L1 Cells. Eur. J. Pharmacol. 2008, 587, 317–321. [Google Scholar] [CrossRef]
- Srinivasan, V.; Radhakrishnan, S.; Angayarkanni, N.; Sulochana, K.N. Antidiabetic Effect of Free Amino Acids Supple-mentation in Human Visceral Adipocytes through Adiponectin-Dependent Mechanism. Indian J. Med. Res. 2019, 149, 41–46. [Google Scholar]
- Coope, A.; Milanski, M.; Araújo, E.P.; Tambascia, M.; Saad, M.J.; Geloneze, B.; Velloso, L.A. Adipor1 Mediates the Anorexigenic and Insulin/Leptin-Like Actions of Adiponectin in the Hypothalamus. FEBS Lett. 2008, 582, 1471–1476. [Google Scholar] [CrossRef]
- Sun, J.; Gao, Y.; Yao, T.; Huang, Y.; He, Z.; Kong, X.; Yu, K.J.; Wang, R.T.; Guo, H.; Yan, J. Adiponectin Potentiates the Acute Effects of Leptin in Arcuatepomcneurons. Mol. Metab. 2016, 5, 882–891. [Google Scholar] [CrossRef]
- Idrizaj, E.; Garella, R.; Castellini, G.; Mohr, H.; Pellegata, N.S.; Francini, F.; Ricca, V.; Squecco, R.; Baccari, M.C. Adiponectin affects the mechanical responses in strips from the mouse gastric fundus. World J. Gastroenterol. 2018, 24, 4028–4035. [Google Scholar] [CrossRef]
- Min, Y.W.; Hong, Y.S.; Ko, E.-J.; Lee, J.-Y.; Ahn, K.D.; Bae, J.M.; Rhee, P.-L. Nitrergic Pathway Is the Main Contributing Mechanism in the Human Gastric Fundus Relaxation: An In Vitro Study. PLoS ONE 2016, 11, e0162146. [Google Scholar] [CrossRef] [PubMed]
- Idrizaj, E.; Garella, R.; Castellini, G.; Francini, F.; Ricca, V.; Baccari, M.C.; Squecco, R. Adiponectin Decreases Gastric Smooth Muscle Cell Excitability in Mice. Front. Physiol. 2019, 10, 1000. [Google Scholar] [CrossRef] [PubMed]
- Idrizaj, E.; Garella, R.; Nistri, S.; Dell’Accio, A.; Cassioli, E.; Rossi, E.; Castellini, G.; Ricca, V.; Squecco, R.; Baccari, M.C. Adiponectin Exerts Peripheral Inhibitory Effects on the Mouse Gastric Smooth Muscle through the Ampk Pathway. Int. J. Mol. Sci. 2020, 21, 9617. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Cheng, K.K.-Y. Hypothalamic AMPK as a Mediator of Hormonal Regulation of Energy Balance. Int. J. Mol. Sci. 2018, 19, 3552. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, N.; Zhang, X.; Chen, D.; Li, Z. The Controversial Role of Adiponectin in Appetite Regulation of Animals. Nutrients 2021, 13, 3387. https://doi.org/10.3390/nu13103387
Tang N, Zhang X, Chen D, Li Z. The Controversial Role of Adiponectin in Appetite Regulation of Animals. Nutrients. 2021; 13(10):3387. https://doi.org/10.3390/nu13103387
Chicago/Turabian StyleTang, Ni, Xin Zhang, Defang Chen, and Zhiqiong Li. 2021. "The Controversial Role of Adiponectin in Appetite Regulation of Animals" Nutrients 13, no. 10: 3387. https://doi.org/10.3390/nu13103387
APA StyleTang, N., Zhang, X., Chen, D., & Li, Z. (2021). The Controversial Role of Adiponectin in Appetite Regulation of Animals. Nutrients, 13(10), 3387. https://doi.org/10.3390/nu13103387