
Study of the Potential Hepatoprotective Effect of Myo-Inositol and Its Influence on Zebrafish Development

 , , ,
, , ,
Abstract
:1. Introduction
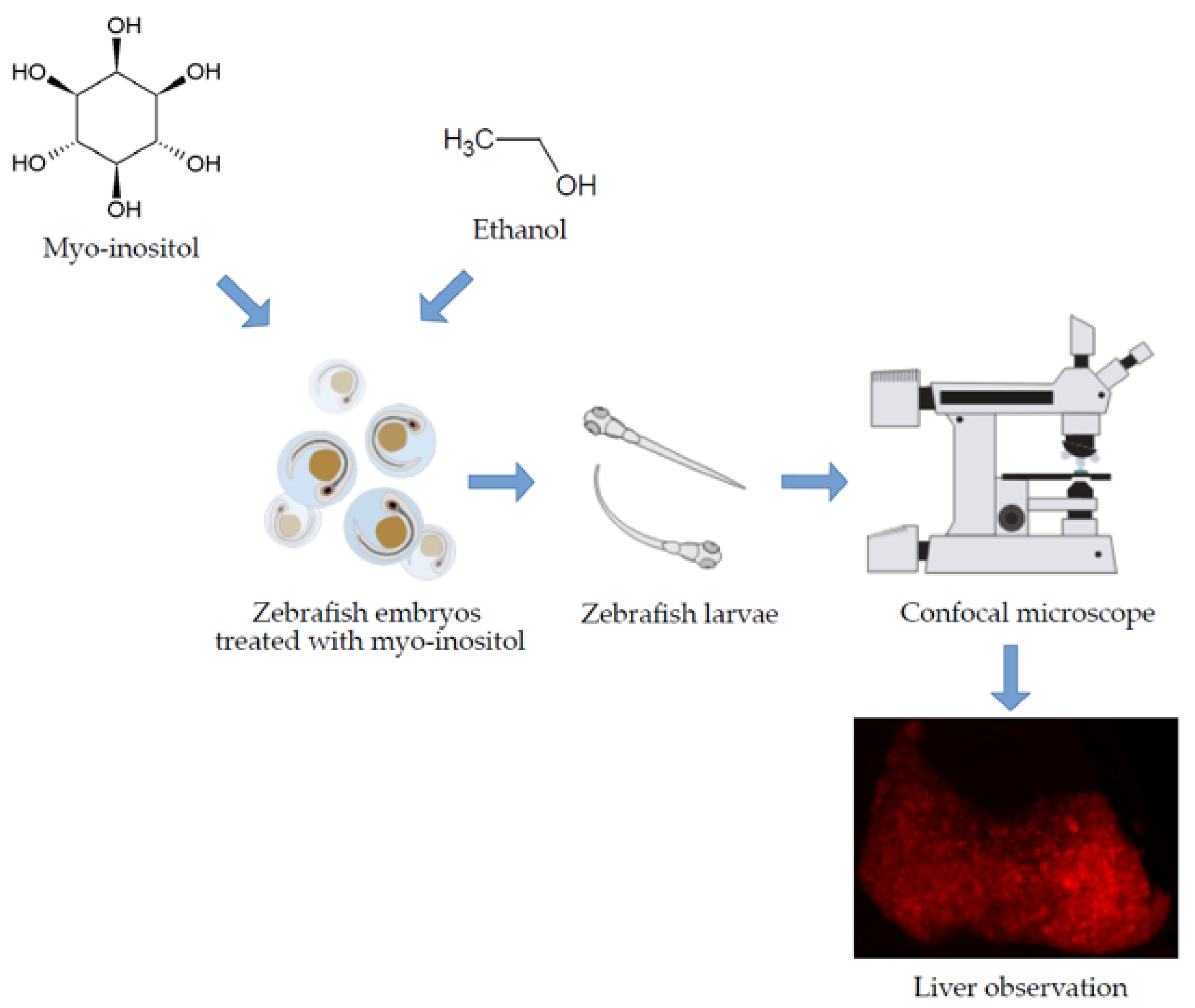
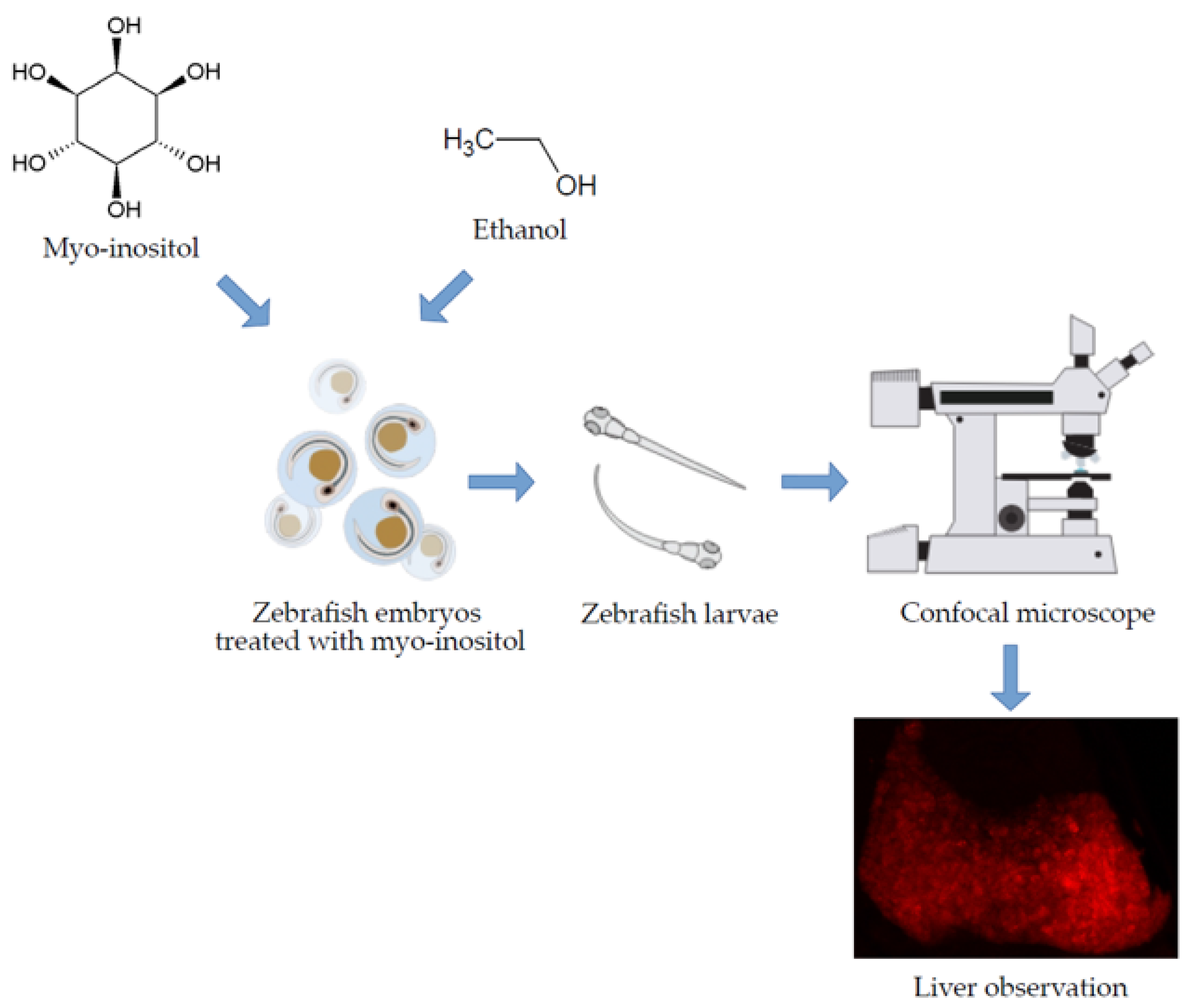
2. Materials and Methods
2.1. MI Toxicity Studies on Zebrafish Larvae
2.2. Hepatoprotective Influence of MI in Acute Liver Damage Zebrafish Larvae
2.3. Imaging and Data Analysis
3. Results
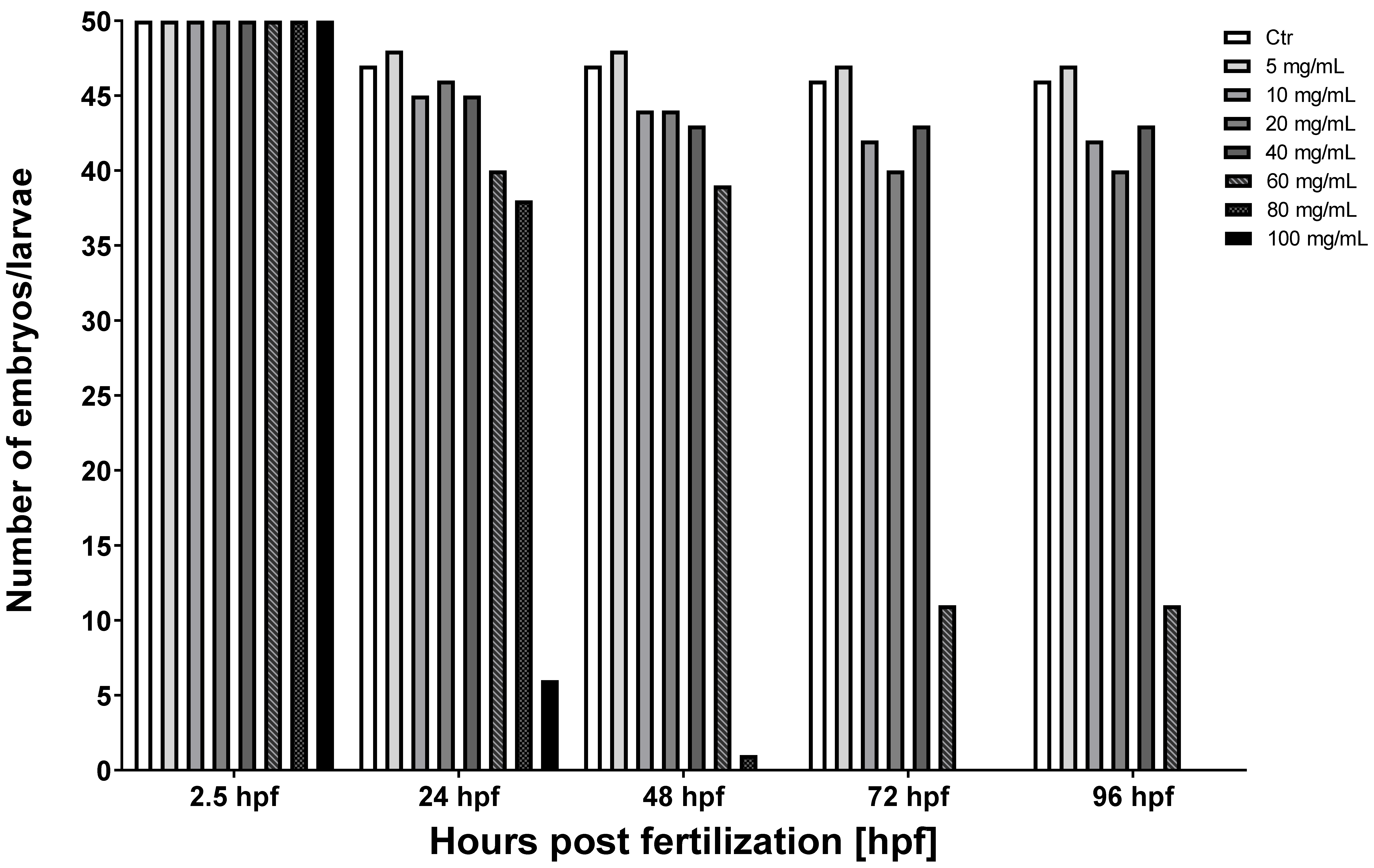
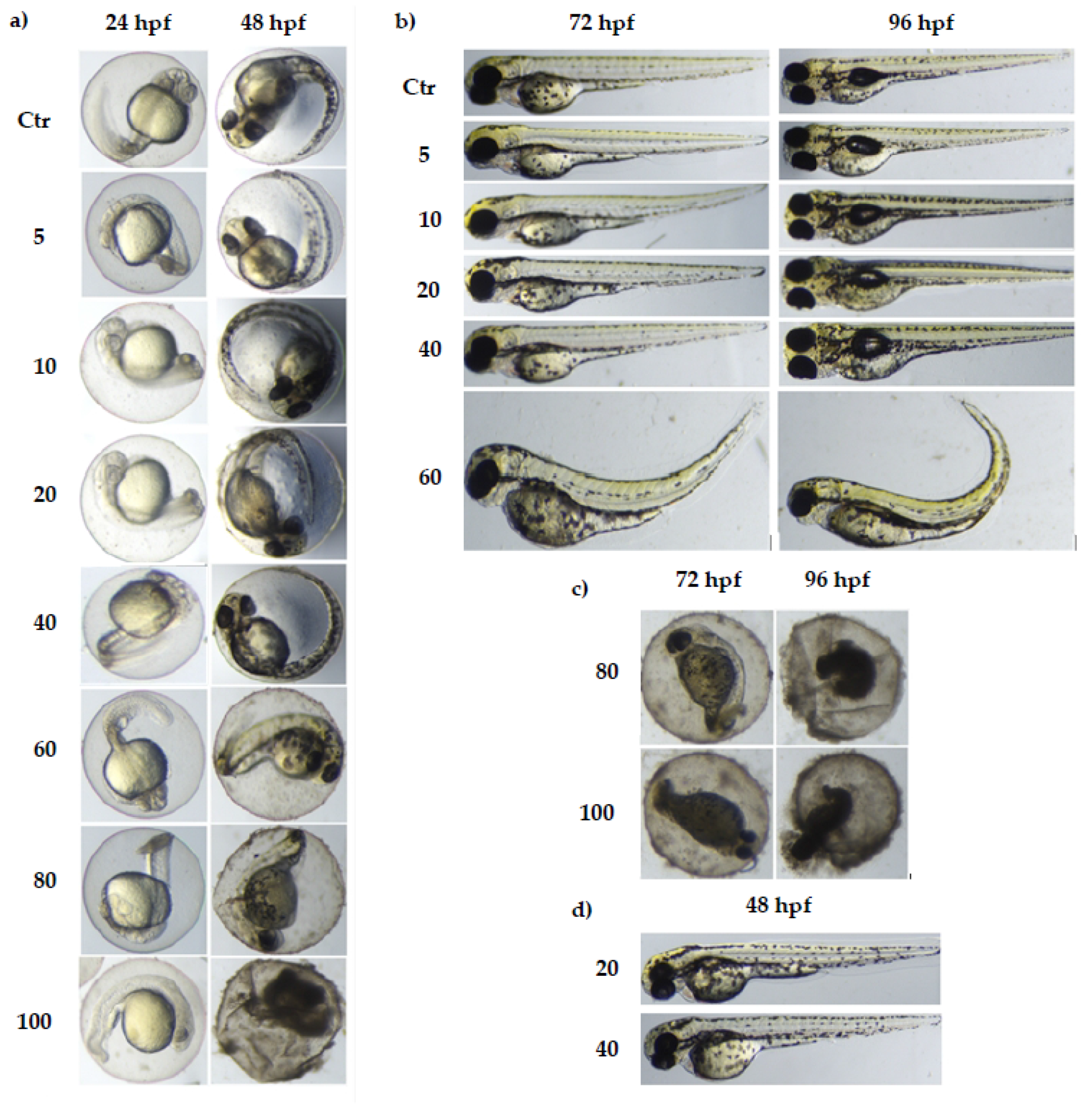
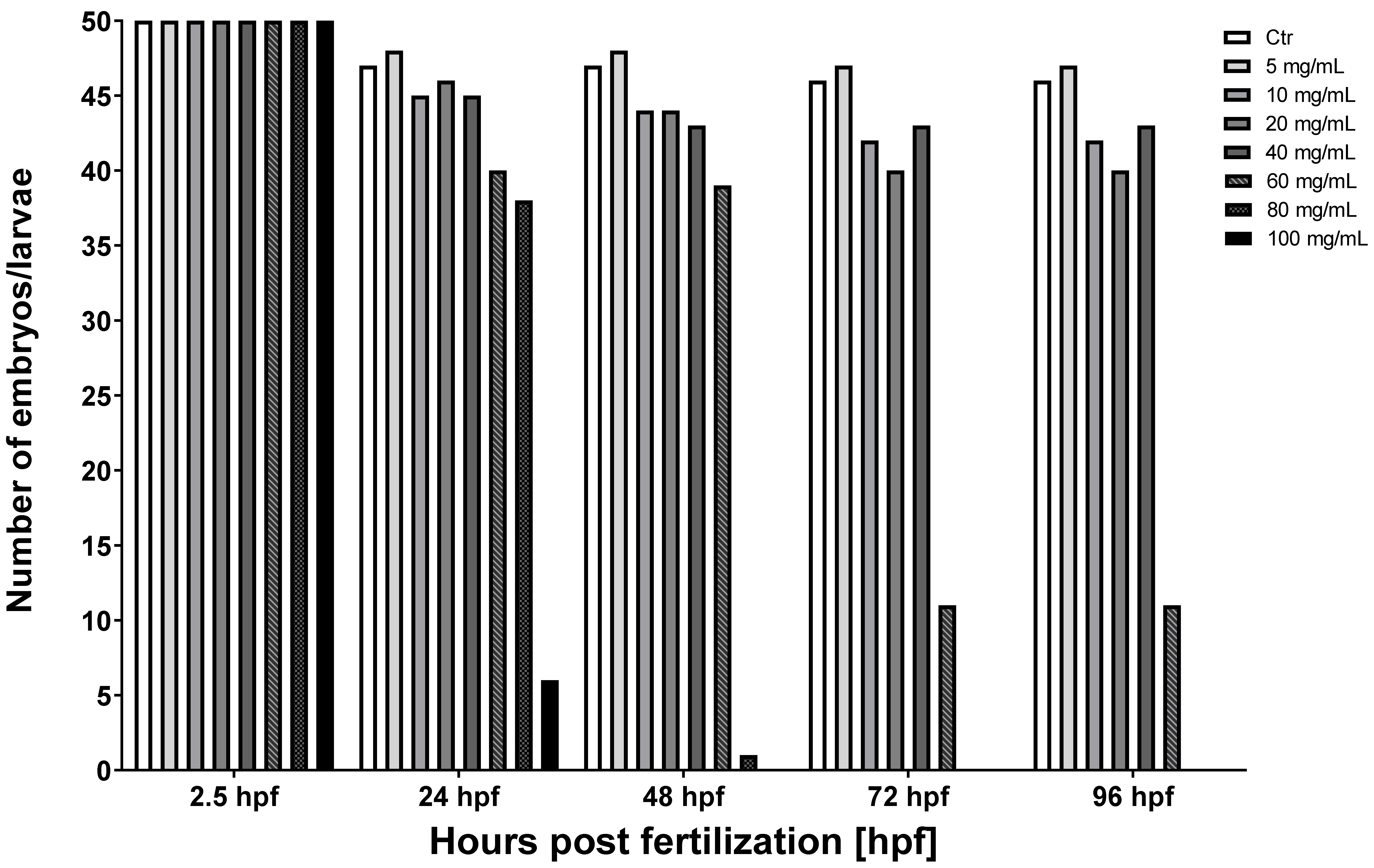
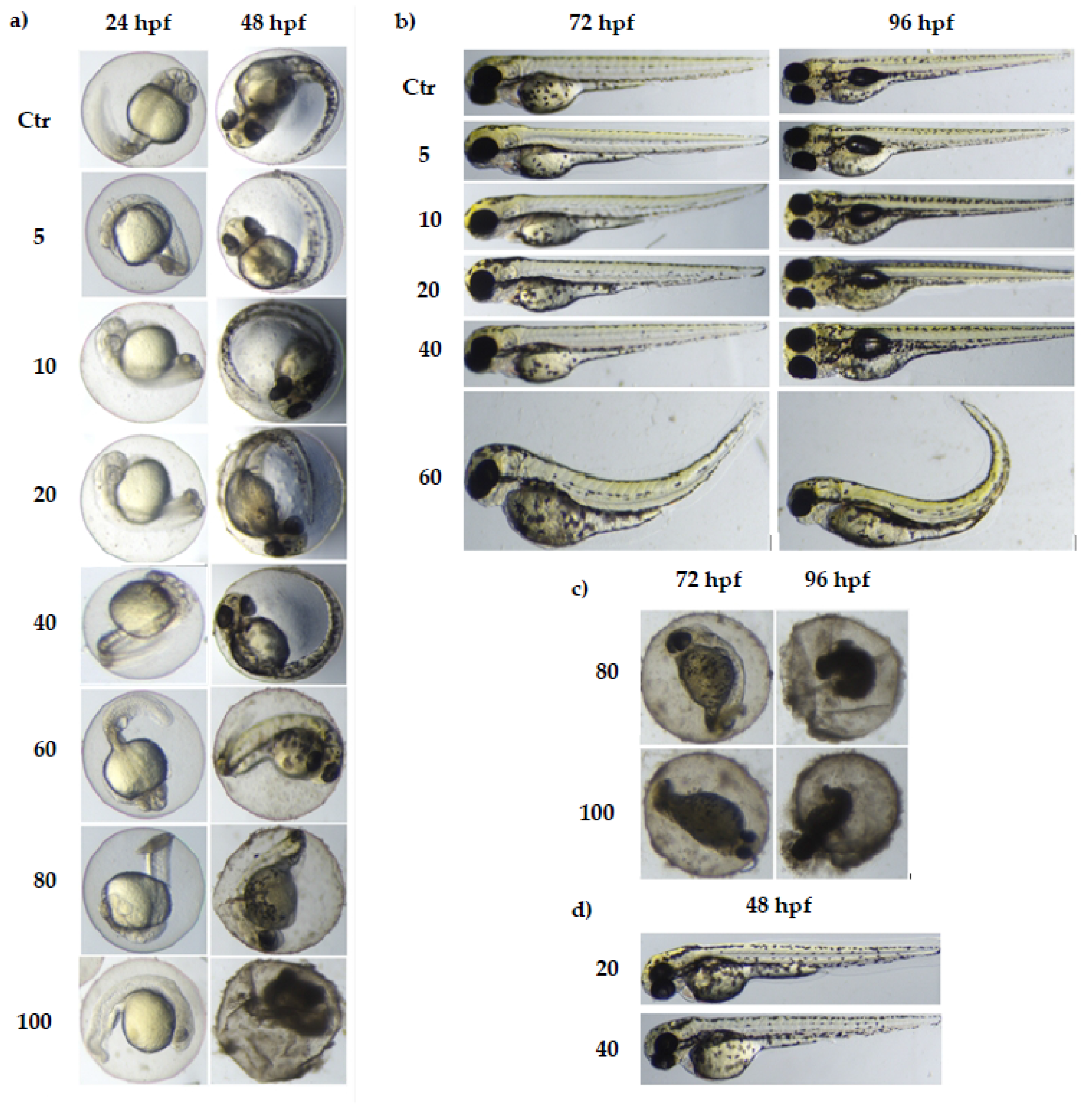
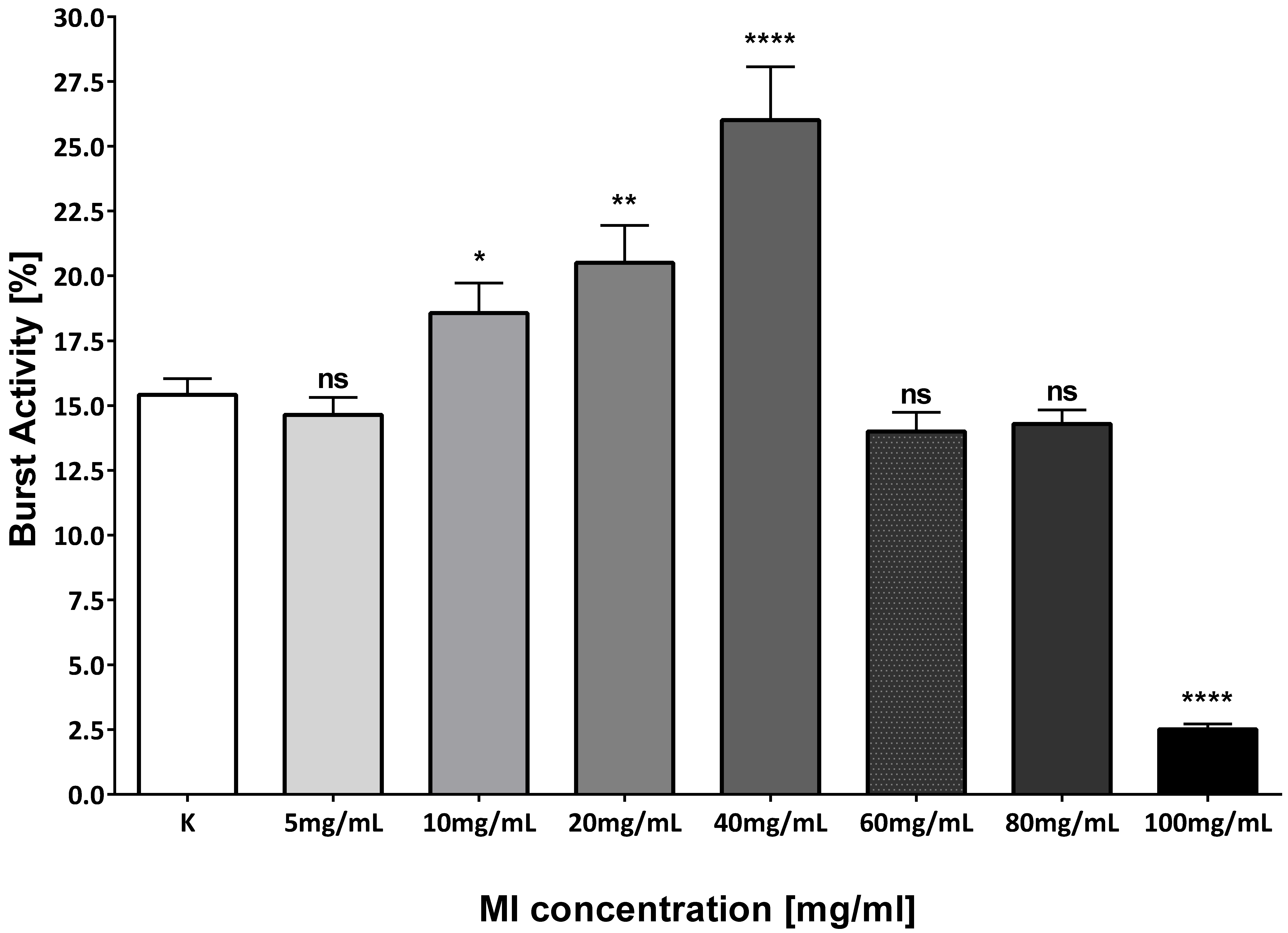
3.1. The Toxicity of Various Concentrations of MI on Zebrafish Model
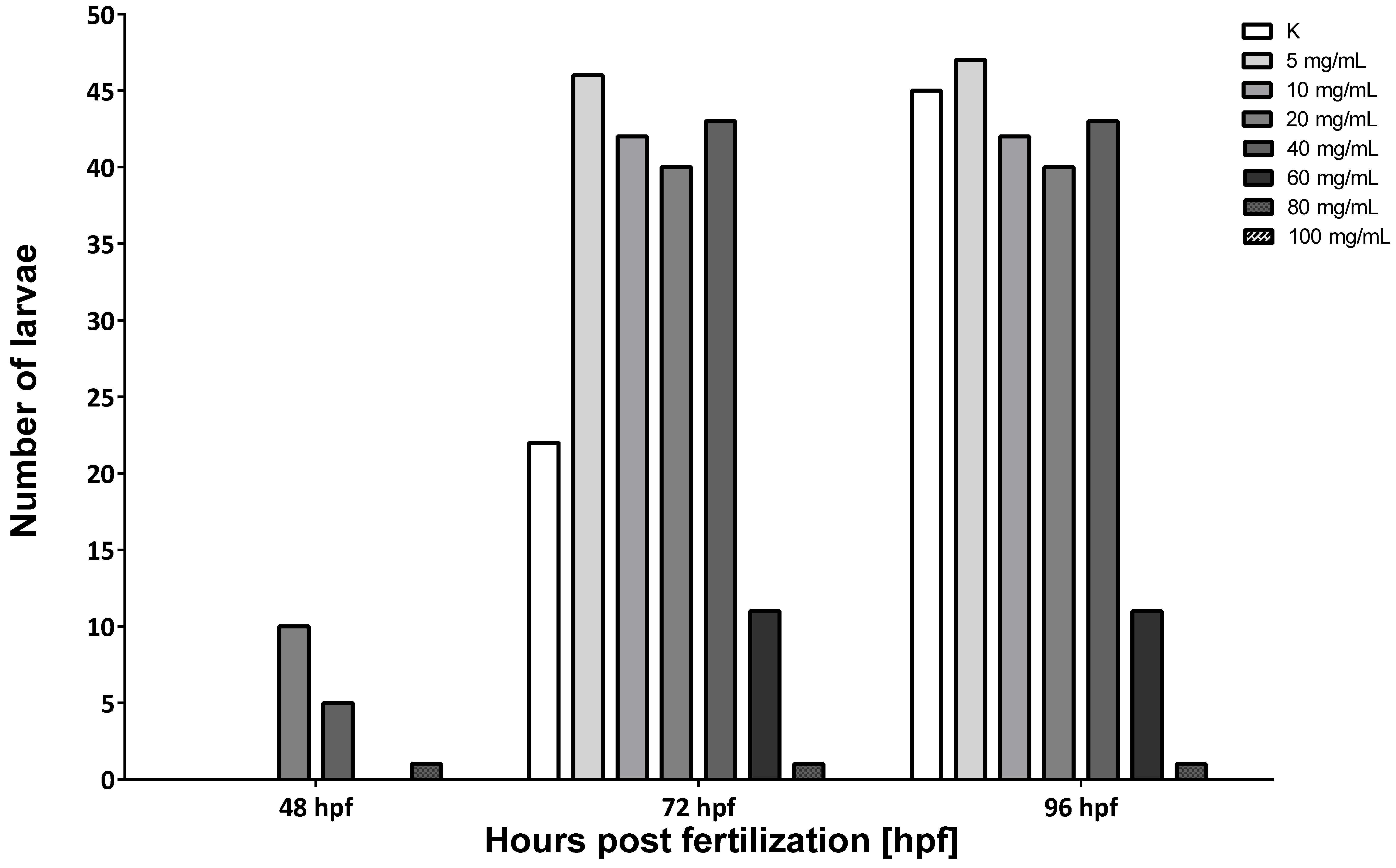
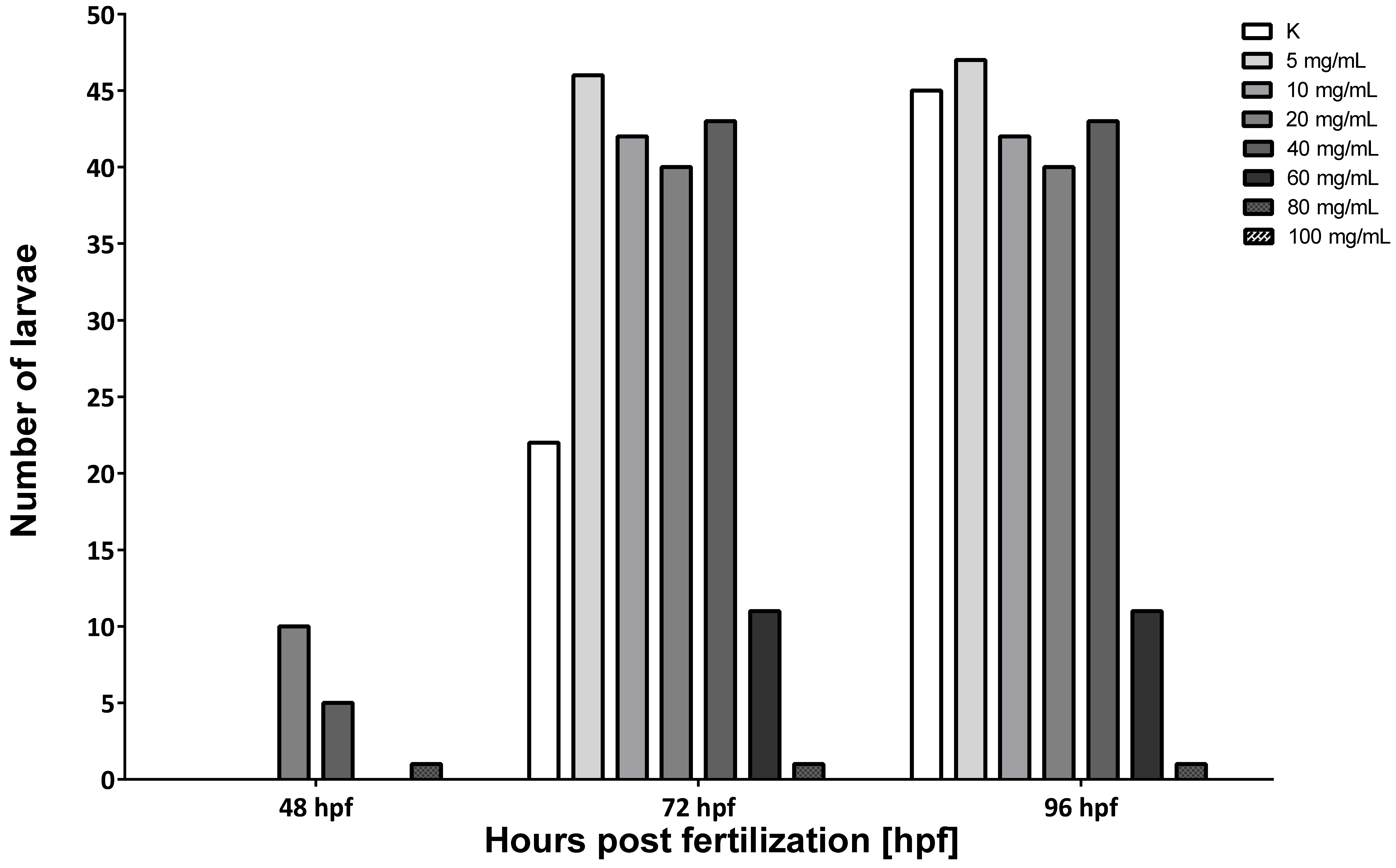
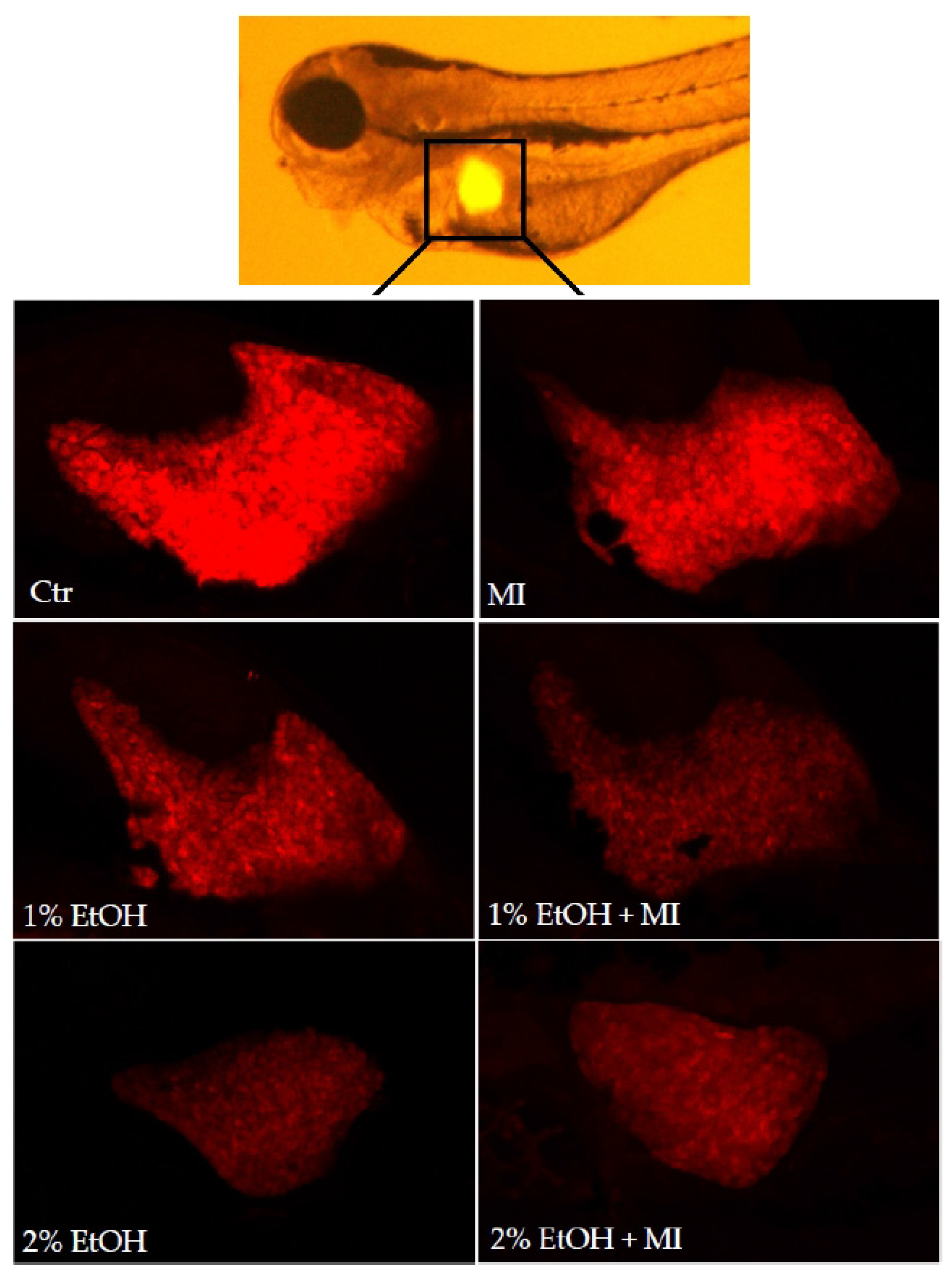
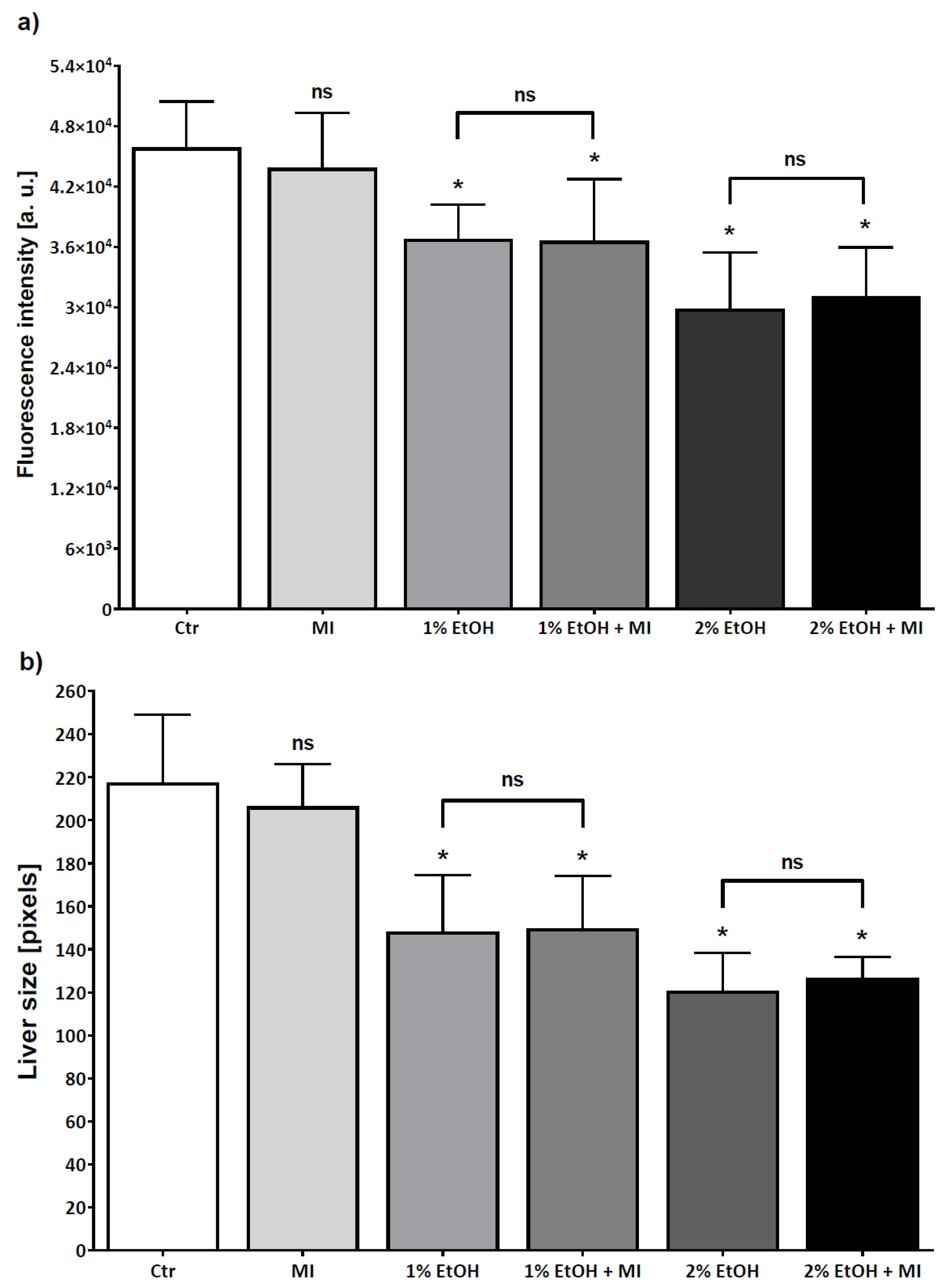
3.2. Hepatoprotective Potential of MI in Alcohol-Induced Acute Liver Damage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MI | myo-inositol |
| EtOH | ethanol |
| hpf | hour post-fertilization |
| hpe | hour post-exposition |
| XAP | Xiaoaiping |
| SPEE | ethanol extracts of Salvia plebeia R. Br. |
| TAA | thioacetamide |
| RCT | randomized controlled trial |
| ALD | alcoholic liver disease |
References
- Wilkins, B.J.; Pack, M. Zebrafish Models of Human Liver Development and Disease. Compr. Physiol. 2013, 3, 1213–1230. [Google Scholar] [CrossRef] [Green Version]
- Maher, J.J. Exploring alcohol’s effects on liver function. Alcohol Health Res. World 1997, 21, 1–12. [Google Scholar]
- Liu, Y.; Liu, X.; Wang, Y.; Yi, C.; Tian, J.; Liu, K.; Chu, J. Protective effect of lactobacillus plantarum on alcoholic liver injury and regulating of keap-Nrf2-ARE signaling pathway in zebrafish larvae. PLoS ONE 2019, 14, e0222339. [Google Scholar] [CrossRef]
- Osna, N.A.; Donohue, T.M., Jr.; Kharbanda, K.K. Alcoholic Liver Disease: Pathogenesis and Current Management. Alcohol Res. Curr. Rev. 2017, 38, 147–161. [Google Scholar]
- Tarantino, G.; Citro, V.; Capone, D. Nonalcoholic Fatty Liver Disease: A Challenge from Mechanisms to Therapy. J. Clin. Med. 2019, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Osowski, A.; Kasparek, A.; Wieczorek, Z.; Amarowicz, R.; Szabelski, M. Evaluation of the characteristics of some plant polyphenols as molecules intercepting mitoxantrone. Food Chem. 2017, 227, 142–148. [Google Scholar] [CrossRef]
- Carlomagno, G.; Unfer, V. Inositol safety: Clinical evidences. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 931–936. [Google Scholar] [PubMed]
- Pak, Y.; Hong, Y.; Kim, S.; Piccariello, T.; Farse, R.; Larner, J. In vivo chiro-inositol metabolism in the rat: A defect in chiro-inositol synthesis from myo-inositol and an increased incorporation of chiro-[3H] inositol into phospolipid in the Goto-Kakizaki (G.K.) rat. Mol. Cells 1998, 8, 301–309. [Google Scholar] [PubMed]
- Michell, R.H. Inositol and its derivatives: Their evolution and functions. Adv. Enzym. Regul. 2011, 51, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Kornienko, A.; D’Alarcao, M. Synthesis of cyclitols via ring-closing metathesis. Tetrahedron Asymmetry 1999, 10, 827–829. [Google Scholar] [CrossRef]
- Wassink, A.M.J.; Olijhoek, J.K.; Visseren, F.L.J. The metabolic syndrome: Metabolic changes with vascular consequences. Eur. J. Clin. Investig. 2007, 37, 8–17. [Google Scholar] [CrossRef]
- Unfer, V.; Carlomango, G.; Rizzo, P.; Raone, E.; Rose, S. Myo-inositol rather than D-chiro-inositol is able to improve oocyte quality in intracytoplasmic sperm injection cycles. A prospective, controlled, randomized trial. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 452–457. [Google Scholar]
- Facchinetti, F.; Bizzarri, M.; Benvenga, S.; D’Anna, R.; Lanzone, A.; Soulage, C.; Di Renzo, G.C.; Hod, M.; Cavalli, P.; Chiu, T.T.; et al. Results from the International Consensus Conference on Myo-inositol and d-chiro-inositol in Obstetrics and Gynecology: The link between metabolic syndrome and PCOS. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 195, 72–76. [Google Scholar] [CrossRef]
- Mancini, M.; Andreassi, A.; Salvioni, M.; Pelliccione, F.; Mantellassi, G.; Banderali, G. Myoinositol and D-Chiro Inositol in Improving Insulin Resistance in Obese Male Children: Preliminary Data. Int. J. Endocrinol. 2016, 2016, 8720342. [Google Scholar] [CrossRef] [Green Version]
- Martin, K.A.; Mani, M.V.; Mani, A. New targets to treat obesity and the metabolic syndrome. Eur. J. Pharmacol. 2015, 763, 64–74. [Google Scholar] [CrossRef] [Green Version]
- Merchant, A.; Arndt, S.K.; Rowell, D.M.; Posch, S.; Callister, A.; Tausz, M.; Adams, M. Seasonal changes in carbohydrates, cyclitols, and water relations of 3 field grown Eucalyptus species from contrasting taxonomy on a common site. Ann. For. Sci. 2010, 67, 104. [Google Scholar] [CrossRef]
- Unfer, V.; Facchinetti, F.; Orrù, B.; Giordani, B.; Nestler, J. Myo-inositol effects in women with PCOS: A meta-analysis of randomized controlled trials. Endocr. Connect. 2017, 6, 647–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastegar, S.; Soltani, S.; Roohipoor, A.; Ebrahimi, E. Study of plants with D-chiro-inositol and its derivatives on diabetes. Int. J. Pharmacol. 2017, 4, 43–53. [Google Scholar]
- Croze, M.L.; Soulage, C.O. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochemie 2013, 95, 1811–1827. [Google Scholar] [CrossRef] [PubMed]
- Ostlund, R.; McGill, J.; Herskowitz, I.; Kipnis, D.; Santiago, J.; Sherman, W. D-chiro-Inositol metabolism in diabetes mellitus. Proc. Natl. Acad. Sci. USA 1993, 90, 9988–9992. [Google Scholar] [CrossRef] [Green Version]
- Nordio, M.; Proietti, E. The combined therapy with myo-inositol and D-chiro-inositol reduces the risk of metabolic disease in PCOS overweight patients compared to myo-inositol supplementation alone. Eur. Rev. Med. Pharmcol. Sci. 2012, 16, 575–581. [Google Scholar]
- Dinicola, S.; Minini, M.; Unfer, V.; Verna, R.; Cucina, A.; Bizzarii, M. Nutritional and acquired deficiencies in inositol bioavailability. Correlations with metabolic disorders. Int. J. Mol. Sci. 2017, 18, 2187. [Google Scholar] [CrossRef]
- Davis, A.; Christiansen, M.; Horowitz, J.F.; Klein, S.; Hellerstein, M.K.; Ostlund, R.E. Effect of pinitol treatment on insulin action in subjects with insulin resistance. Diabetes Care 2000, 23, 1000–1005. [Google Scholar] [CrossRef] [Green Version]
- Pani, A.; Giossi, R.; Menichelli, D.; Fittipaldo, V.; Agnelli, F.; Inglese, E.; Romandini, A.; Roncato, R.; Pintaudi, B.; Del Sole, F.; et al. Inositol and Non-Alcoholic Fatty Liver Disease: A Systematic Review on Deficiencies and Supplementation. Nutrition 2020, 12, 3379. [Google Scholar] [CrossRef]
- Shwartz, A.; Goessling, W.; Yin, C. Macrophages in Zebrafish Models of Liver Diseases. Front. Immunol. 2019, 10, 2840. [Google Scholar] [CrossRef]
- Marrs, J.A.; Clendenon, S.; Ratcliffe, D.R.; Fielding, S.M.; Liu, Q.; Bosron, W.F. Zebrafish fetal alcohol syndrome model: Effects of ethanol are rescued by retinoic acid supplement. Alcohol 2010, 44, 707–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passeri, M.; Cinaroglu, A.; Gao, C.; Sadler, K.C. Hepatic Steatosis in Response to Acute Alcohol Exposure in Zebrafish re-quires Srebp Activation. Hepatology 2009, 49, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Selderslaghs, I.W.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef]
- Osowski, A.; Pietrzak, M.; Wieczorek, Z.; Wieczorek, J. Natural Compounds in the Human Diet and their Ability to Bind Mutagens Prevents DNA–Mutagen Intercalation. J. Toxicol. Environ. Health Part A 2010, 73, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Anita Singh, A.; Tiwari, A. Phytopharmacological Overview on Controversial Drug: Murva. Tradit. Folk Herb. Med. Recent Res. 2018, 2, 475–526. [Google Scholar]
- Maldonado, E.; Juárez-Jaimes, V. Chemical constituents from Marsdenia callosa. Biochem. Syst. Ecol. 2013, 48, 219–221. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Liu, K.; He, Q.; Sun, C.; Han, J.; Han, L.; Tian, Q. Xiaoaiping Induces Developmental Toxicity in Zebrafish Embryos Through Activation of ER Stress, Apoptosis and the Wnt Pathway. Front. Pharmacol. 2018, 9, 1250. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Selkoe, D.J. Systematic analysis of time-dependent neural effects of soluble amyloid β oligomers in culture and in vivo: Prevention by scyllo-inositol. Neurobiol. Dis. 2015, 82, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Howarth, D.L.; Passeri, M.; Sadler, K.C. Drinks Like a Fish: Using Zebrafish to Understand Alcoholic Liver Disease. Alcohol. Clin. Exp. Res. 2011, 35, 826–829. [Google Scholar] [CrossRef] [Green Version]
- Shigeta, T.; Hsu, H.-C.; Enosawa, S.; Matsuno, N.; Kasahara, M.; Matsunari, H.; Umeyama, K.; Watanabe, M.; Nagashima, H. Transgenic Pig Expressing the Red Fluorescent Protein Kusabira-Orange as a Novel Tool for Preclinical Studies on Hepatocyte Transplantation. Transplant. Proc. 2013, 45, 1808–1810. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.-S.; Lee, M.-K.; Jung, U.J.; Kim, H.-J.; Do, G.-M.; Park, Y.B.; Jeon, S.-M. Metabolic response of soy pinitol on lipid-lowering, antioxidant and hepatoprotective action in hamsters fed-high fat and high cholesterol diet. Mol. Nutr. Food Res. 2009, 53, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Deng, Y.; Cao, Z.; Liao, X.; Zhang, J.; Lu, H. The hepatoprotective effects of Salvia plebeia R. Br. extract in zebrafish (Danio rerio). Fish Shellfish Immunol. 2019, 95, 399–410. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MI Dose [mg/mL] | 2.5 hpf | 24 hpf | 48 hpf | 72 hpf | 96 hpf |
|---|---|---|---|---|---|
| Ctr | 50 | 47 | 47 | 46 | 46 |
| 5 | 50 | 48 | 48 | 47 | 47 |
| 10 | 50 | 45 | 44 | 42 | 42 |
| 20 | 50 | 46 | 44 | 40 | 40 |
| 40 | 50 | 45 | 43 | 43 | 43 |
| 60 | 50 | 40 | 39 | 11 | 11 |
| 80 | 50 | 38 | 1 | 0 | 0 |
| 100 | 50 | 6 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antonowski, T.; Wiśniewski, K.; Podlasz, P.; Osowski, A.; Wojtkiewicz, J. Study of the Potential Hepatoprotective Effect of Myo-Inositol and Its Influence on Zebrafish Development. Nutrients 2021, 13, 3346. https://doi.org/10.3390/nu13103346
Antonowski T, Wiśniewski K, Podlasz P, Osowski A, Wojtkiewicz J. Study of the Potential Hepatoprotective Effect of Myo-Inositol and Its Influence on Zebrafish Development. Nutrients. 2021; 13(10):3346. https://doi.org/10.3390/nu13103346
Chicago/Turabian StyleAntonowski, Tomasz, Karol Wiśniewski, Piotr Podlasz, Adam Osowski, and Joanna Wojtkiewicz. 2021. "Study of the Potential Hepatoprotective Effect of Myo-Inositol and Its Influence on Zebrafish Development" Nutrients 13, no. 10: 3346. https://doi.org/10.3390/nu13103346
APA StyleAntonowski, T., Wiśniewski, K., Podlasz, P., Osowski, A., & Wojtkiewicz, J. (2021). Study of the Potential Hepatoprotective Effect of Myo-Inositol and Its Influence on Zebrafish Development. Nutrients, 13(10), 3346. https://doi.org/10.3390/nu13103346