Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats

 , , ,
, , ,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Protein-Rich Flours from Quinoa and Buckwheat
2.2. Chemical Composition of Protein-Rich Flours
2.3. In Vivo Experiment
2.4. Sample Collection and Basic Analyses
2.5. RNA Isolation and Quantitative RT-PCR
2.6. Statistical Analysis
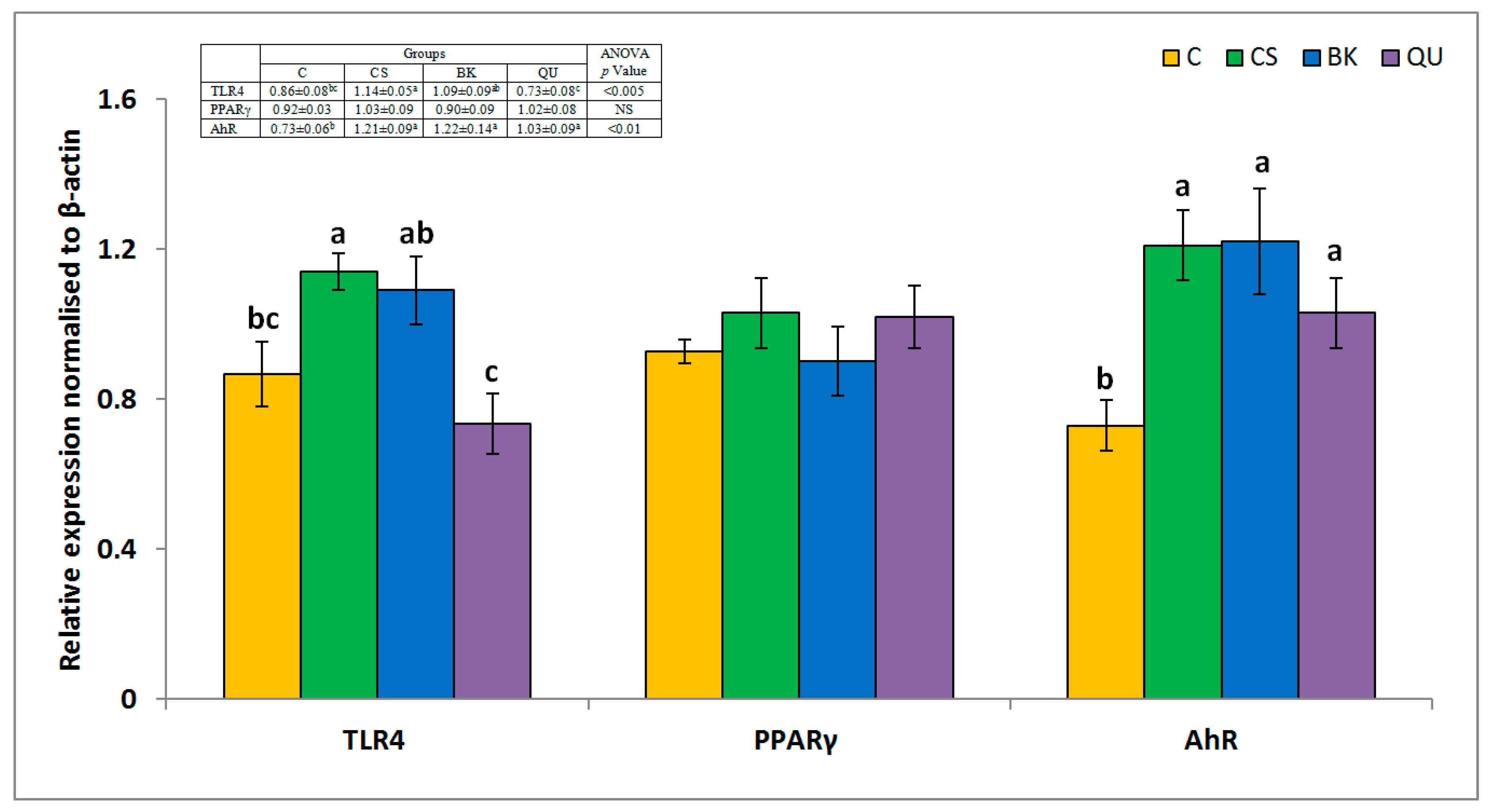
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Filho, A.M.; Pirozi, M.R.; Borges, J.T.; Pinheiro-Sant’Ana, H.M.; Chaves, J.B.; Coimbra, J.S. Quinoa: Nutritional, functional, and antinutritional aspects. Crit. Rev. Food Sci. Nutr. 2017, 8, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Sytar, O.; Brestic, M.; Zivcak, M.; Tran, L.S. The contribution of buckwheat genetic resources to health and dietary diversity. Curr. Genom. 2016, 3, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Comino, I.; Moreno-Mde, L.; Real, A.; Rodríguez-Herrera, A.; Barro, F.; Sousa, C. The gluten-free diet: Testing alternative cereals tolerated by celiac patients. Nutrients 2013, 10, 4250–4268. [Google Scholar] [CrossRef] [PubMed]
- Blaut, M. Relationship of prebiotics and food to intestinal microflora. Eur. J. Nutr. 2002, 41, 11–16. [Google Scholar] [CrossRef]
- Bilić-Šobot, D.; Kubale, V.; Škrlep, M.; Čandek-Potokar, M.; Prevolnik Povše, M.; Fazarinc, G.; Škorjanc, D. Effect of hydrolysable tannins on intestinal morphology, proliferation and apoptosis in entire male pigs. Arch. Anim. Nutr. 2016, 5, 378–388. [Google Scholar] [CrossRef]
- Zevallos, V.F.; Raker, V.; Tenzer, S.; Jimenez-Calvente, C.; Ashfaq-Khan, M.; Rüssel, N.; Pickert, G.; Schild, H.; Steinbrink, K.; Schuppan, D. Nutritional wheat amylase-trypsin inhibitors promote intestinal inflammation via activation of myeloid cells. Gastroenterology 2017, 5, 1100–1113. [Google Scholar] [CrossRef]
- Guo, G.; Lv, D.; Yan, X.; Subburaj, S.; Ge, P.; Li, X.; Hu, Y.; Yan, Y. Proteome characterization of developing grains in bread wheat cultivars (Triticum aestivum L.). BMC Plant Biol. 2012, 12, 147. [Google Scholar] [CrossRef]
- Salazar, F.; Awuah, D.; Negm, O.H.; Shakib, F.; Ghaemmaghami, A.M. The role of indoleamine 2,3-dioxygenase-aryl hydrocarbon receptor pathway in the TLR4-induced tolerogenic phenotype in human DCs. Sci. Rep. 2017, 7, 43337. [Google Scholar] [CrossRef]
- Tanos, R.; Patel, R.D.; Murray, I.A.; Smith, P.B.; Patterson, A.D.; Perdew, G.H. Aryl hydrocarbon receptor regulates the cholesterol biosynthetic pathway in a dioxin response element-independent manner. Hepatology 2012, 6, 1994–2004. [Google Scholar] [CrossRef]
- Laparra, J.; Fotschki, B.; Haros, C. Immunonutritional consequences of different serine-type protease inhibitors in a C57BL/6 hepatocarcinoma model. Oncotarget 2019, 7, 760–772. [Google Scholar] [CrossRef]
- Laparra, J.M.; Haros, C.M. Plant seed protease inhibitors differentially affect innate immunity in a tumor microenvironment to control hepatocarcinoma. Food Funct. 2019, 7, 4210–4219. [Google Scholar] [CrossRef]
- Bai, C.Z.; Feng, M.L.; Hao, X.L.; Zhao, Z.J.; Li, Y.Y.; Wang, Z.H. Anti-tumoral effects of a trypsin inhibitor derived from buckwheat in vitro and in vivo. Mol. Med. Rep. 2015, 2, 1777–1782. [Google Scholar] [CrossRef] [PubMed]
- Mao, K.; Baptista, A.P.; Tamoutounour, S.; Zhuang, L.; Bouladoux, N.; Martins, A.J.; Huang, Y.; Gerner, M.Y.; Belkaid, Y.; Germain, R.N. Innate and adaptive lymphocytes sequentially shape the gut microbiota and lipid metabolism. Nature 2018, 554, 255–259. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Horwitz, W., Latimer, G.W., Eds.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Valcárcel-Yamani, B.; Lannes, S.C.S. Application of quinoa (Chenopodium Quinoa Willd.) and amaranth (Amaranthus Spp.) and their influence in the nutritional value of cereal based food. Food Pub. Health 2012, 2, 265–275. [Google Scholar]
- Wronkowska, M.; Piskuła, K.M.; Zieliński, H. Effect of roasting time of buckwheat groats on the formation of Maillard reaction products and antioxidant capacity. Food Chem. 2016, 196, 355–358. [Google Scholar]
- Pendl, R.; Bauer, M.; Caviezel, R.; Schulthess, P. Determination of total fat in foods and feeds by the caviezel method, based on a gas chromatographic technique. J. AOAC Int. 1998, 81, 907–917. [Google Scholar] [CrossRef]
- Amarowicz, R.; Raab, B. Antioxidative activity of leguminous seed extracts evaluated by chemiluminescence methods. Z. Nat. C 1997, 52, 709–712. [Google Scholar] [CrossRef]
- Amarowicz, R.; Wanasundara, U.N.; Karamać, M.; Shahidi, F. Antioxidant activity of ethanolic extract of mustard seed. Nahr. Food 1996, 40, 261–263. [Google Scholar] [CrossRef]
- Price, M.L.; van Scoyoc, S.; Butler, L.G. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Kakade, M.D.; Rackis, J.J.; McGhee, J.E.; Puski, G. Determination of trypsin inhibitor activity of soy products: A collaborative analysis of an improved procedure. Cereal Chem. 1974, 51, 376–382. [Google Scholar]
- Reeves, P.G. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr. 1997, 127, 838–841. [Google Scholar] [CrossRef] [PubMed]
- Fotschki, B.; Jurgoński, A.; Fotschki, J.; Majewski, M.; Ognik, K.; Juśkiewicz, J. Dietary chicory inulin-rich meal exerts greater healing effects than fructooligosaccharide preparation in rats with trinitrobenzenesulfonic acid-induced necrotic colitis. Pol. J. Food Nutr. Sci. 2019, 69, 147–155. [Google Scholar] [CrossRef]
- Żary-Sikorska, E.; Fotschki, B.; Fotschki, J.; Wiczkowski, W.; Juśkiewicz, J. Preparations from purple carrots containing anthocyanins improved intestine microbial activity, serum lipid profile and antioxidant status in rats. J. Funct. Foods 2019, 60, 103442. [Google Scholar] [CrossRef]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef]
- Bäckhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef]
- Vega-Gálvez, A.V.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martínez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa Willd.), an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar] [CrossRef]
- Mota, C.; Santos, M.; Mauro, R.; Samman, N.; Matos, A.S.; Torres, D.; Castanheira, I. Protein content and amino acids profile of pseudocereals. Food Chem. 2016, 15, 55–61. [Google Scholar] [CrossRef]
- Cirkovic Velickovicand, T.D.; Stanic-Vucinic, D.J. The role of dietary phenolic compounds in protein digestion and processing technologiesto improve their antinutritive properties. Compr. Rev. Food Sci. Food Saf. 2018, 17, 82–103. [Google Scholar] [CrossRef]
- Kumar Singh, A.; Cabral, C.; Kumar, R.; Ganguly, R.; Kumar Rana, H.; Gupta, A.; Rosaria Lauro, M.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Qiu, B.; Fan, S.; Ding, H.; Liu, Z. Quinoa whole grain diet compromises the changes of gut microbiota and colonic colitis induced by dextran Sulfate sodium in C57BL/6 mice. Sci. Rep. 2018, 8, 14916. [Google Scholar] [CrossRef]
- Bifari, F.; Ruocco, C.; Decimo, I.; Fumagalli, G.; Valerio, A.; Nisoli, E. Amino acid supplements and metabolic health: A potential interplay between intestinal microbiota and systems control. Genes Nutr. 2017, 12, 27. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Priebe, M.G.; Vonk, R.J.; Welling, G.W. Identification of bacteria with β-galactosidase activity in faeces from lactase non-persistent subjects. FEMS Microbiol. Ecol. 2005, 54, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, S.; Watkins, C.; Stanton, C.; Ross, R.P. Gut bifidobacteria populations in human health and aging. Front. Microbiol. 2016, 7, 1204. [Google Scholar] [CrossRef]
- Neis, E.P.; Dejong, C.H.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 4, 2930–2946. [Google Scholar] [CrossRef] [PubMed]
- Davila, A.M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.H.; Sanz, Y.; Tomé, D. Re-print of “Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host”. Pharmacol. Res. 2013, 1, 114–126. [Google Scholar] [CrossRef]
- Barker, H.A. Amino acid degradation by anaerobic bacteria. Annu. Rev. Biochem. 1981, 50, 23–40. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Giménez-Bastida, J.A.; Zieliński, H. Buckwheat as a functional food and its effects on health. J. Agric. Food Chem. 2015, 63, 7896–7913. [Google Scholar] [CrossRef]
- Gullón, B.; Gullón, P.; Tavaria, F.K.; Yáñez, R. Assessment of the prebiotic effect of quinoa and amaranth in the human intestinal ecosystem. Food Funct. 2016, 7, 3782–3788. [Google Scholar] [CrossRef]
- Hara, H.; Haga, S.; Aoyama, Y.; Kiriyama, S. Short-chain fatty acids suppress cholesterol synthesis in rat liver and intestine. J. Nutr. 1999, 5, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, L.; Martin-Gallausiaux, C.; Bourhis, J.M.; Béguet-Crespel, F.; Blottière, H.M.; Lapaque, N. Identification of the novel role of butyrate as AhR ligand in human intestinal epithelial cells. Sci. Rep. 2019, 1, 643. [Google Scholar] [CrossRef] [PubMed]
- Singhal, R.; Badger, T.M.; Ronis, M.J. Rats fed soy protein isolate (SPI) have impaired hepatic CYP1A1 induction by polycyclic aromatic hydrocarbons as a result of interference with aryl hydrocarbon receptor signaling. Toxicol. Appl. Pharmacol. 2008, 2, 275–283. [Google Scholar] [CrossRef]
- Seth, R.K.; Kimono, D.; Alhasson, F.; Sarkar, S.; Albadrani, M.; Lasley, S.K.; Horner, R.; Janulewicz, P.; Nagarkatti, M.; Nagarkatti, P.; et al. Increased butyrate priming in the gut stalls microbiome associated-gastrointestinal inflammation and hepatic metabolic reprogramming in a mouse model of Gulf War Illness. Toxicol. Appl. Pharmacol. 2018, 350, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Park, K.T.; Yun, C.H.; Bae, C.S.; Ahn, T. Decreased level of albumin in peripheral blood mononuclear cells of streptozotocin-induced diabetic rats. J. Vet. Med. Sci. 2014, 8, 1087–1092. [Google Scholar] [CrossRef]
- Pinno, J.; Bongartz, H.; Klepsch, O.; Wundrack, N.; Poli, V.; Schaper, F.; Dittrich, A. Interleukin-6 influences stress-signalling by reducing the expression of the mTOR-inhibitor REDD1 in a STAT3-dependent manner. Cell. Signal. 2016, 8, 907–916. [Google Scholar] [CrossRef]


{kind=link}
| Ingredient (%) | Groups | |||
|---|---|---|---|---|
| C | CS | BK | QU | |
| Casein | 11.15 | |||
| D,L-methionine | 0.20 | |||
| Soya protein isolate | 10.8 | |||
| Buckwheat protein-rich flour | 41.32 | |||
| Quinoa protein-rich flour | 28.01 | |||
| Cellulose 1 | 8 | 8 | 8 | 8 |
| Soya oil | 8 | 8 | 8 | 8 |
| Mineral mix 2 | 3.5 | 3.5 | 3.5 | 3.5 |
| Vitamin mix 2 | 1 | 1 | 1 | 1 |
| Choline chloride | 0.2 | 0.2 | 0.2 | 0.2 |
| Cholesterol | 0.3 | 0.3 | 0.3 | 0.3 |
| Sucrose | 5 | 5 | 5 | 5 |
| Corn starch | 62.65 | 63.2 | 32.68 | 45.99 |
| Calculated calorie per kg of diet | 3880 | 3880 | 3776 | 3986 |
| Buckwheat (BK) | Quinoa (QU) | |
|---|---|---|
| DM (%) | 92.5 | 94.9 |
| Ash (%) | 3.05 | 3.6 |
| Protein (%) | 24.2 | 35.7 |
| Fat (%) | 3.4 | 14.7 |
| Carbohydrates a (%) | 61.85 | 40.9 |
| Antinutrients | ||
| Tannins (mg/g) | 13.9 | 0.0 |
| Phenolics (mg/g) | 7.41 | 2.05 |
| Phytic acid (mg/g) | 15.9 | 8.3 |
| TIAs (IU/mg) | 13.3 | 0.4 |
| Parameters | Groups | ANOVA p Value | |||
|---|---|---|---|---|---|
| C | CS | BK | QU | ||
| Diet intake (g/14 days) | 258 ± 2.90 a | 231 ± 11.730 b | 265 ± 6.48 a | 242 ± 9.84 a,b | <0.05 |
| Calorie intake (kcal/14 days) | 1001 ± 11 a | 896 ± 45 b | 999 ± 24 a | 964 ± 39 a,b | <0.05 |
| Feed efficiency ratio (g/g) * | 0.32 ± 0.02 a | 0.15 ± 0.02 b | 0.26 ± 0.02 a | 0.25 ± 0.02 a | <0.001 |
| BV (%) | 75.3 ± 2.64 a | 44.2 ± 2.20 c | 61.8 ± 0.538 a,b | 55.6 ± 0.901 b,c | <0.001 |
| Initial body weight (g) | 170 ± 11.6 | 170 ± 11.9 | 169 ± 11.6 | 170 ± 11.9 | NS |
| Final body weight (g) | 253 ± 8.21 a | 206 ± 10.3 b | 238 ± 7.97 a | 232 ± 9.97 a,b | 0.01 |
| Body weight gain (g) | 83.5 ± 8.23 a | 35.4 ± 6.30 c | 68.7 ± 6.17 a,b | 61.8 ± 6.21 b | <0.001 |
| Liver (g/100 g BW) | 4.79 ± 0.175 a | 3.68 ± 0.150 b | 4.00 ± 0.071 b | 3.79 ± 0.085 b | <0.001 |
| Hepatosomatic index (%) ** | 1.98 ± 0.11 a | 1.90 ± 0.08 a | 1.69 ± 0.06 b | 1.64 ± 0.07 b | <0.05 |
| Kidneys (g/100 g BW) | 0.699 ± 0.019 | 0.746 ± 0.025 | 0.758 ± 0.015 | 0.741 ± 0.017 | NS |
| Caecum | |||||
| Tissue (g/100 g BW) | 0.195 ± 0.004 b | 0.262 ± 0.014 a | 0.261 ± 0.017 a | 0.314 ± 0.021 a | 0.001 |
| Digesta (g/100 g BW) | 0.614 ± 0.036 b | 0.828 ± 0.032 a | 0.861 ± 0.037 a | 0.777 ± 0.059 a | 0.005 |
| Ammonia (mg/g) | 0.244 ± 0.017 | 0.228 ± 0.028 | 0.216 ± 0.011 | 0.185 ± 0.008 | NS |
| pH | 7.69 ± 0.032 a | 7.56 ± 0.039 a | 7.38 ± 0.055 b | 7.28 ± 0.059 b | <0.001 |
| Colon | |||||
| Tissue (g/100 g BW) | 0.376 ± 0.025 | 0.412 ± 0.010 | 0.373 ± 0.014 | 0.430 ± 0.017 | 0.072 |
| Digesta (g/100 g BW) | 0.443 ± 0.055 b | 0.671 ± 0.041 a | 0.490 ± 0.037 b | 0.470 ± 0.065 b | <0.05 |
| pH | 7.80 ± 0.150 | 7.71 ± 0.134 | 7.41 ± 0.181 | 7.39 ± 0.118 | NS |
| Parameters | Groups | ANOVA p Value | |||
|---|---|---|---|---|---|
| C | CS | BK | QU | ||
| Microbial enzymatic activity (µmol/h/g digesta) | |||||
| α-Glucosidase | 10.7 ± 0.620 b | 12.3 ± 1.32 b | 20.6 ± 1.72 a | 21.0 ± 2.36 a | <0.001 |
| β-Glucosidase | 1.95 ± 0.253 d | 6.98 ± 0.749 c | 10.4 ± 0.926 b | 14.31.92 a | <0.001 |
| α-Galactosidase | 7.61 ± 0.749 b | 10.9 ± 1.73 b | 24.9 ± 2.22 a | 29.6 ± 2.72 a | <0.001 |
| β-Galactosidase | 21.2 ± 2.21 c | 29.6 ± 2.54 b,c | 39.7 ± 4.27 a,b | 60.0 ± 4.29 a | <0.001 |
| β-Glu curonidase | 8.97 ± 1.05 c | 39.6 ± 5.87 a | 22.0 ± 2.54 b | 42.0 ± 3.82 a | <0.001 |
| SCFA, (μmol/g digesta) | |||||
| Acetate | 37.9 ± 2.55 b | 43.0 ± 2.17 b | 58.0 ± 4.81 a | 47.7 ± 4.00 b | <0.005 |
| Propionate | 8.42 ± 0.651 | 9.51 ± 0.562 | 10.3 ± 0.530 | 9.48 ± 0.217 | NS |
| Isobutyrate | 0.811 ± 0.059 | 0.941 ± 0.078 | 0.947 ± 0.070 | 0.890 ± 0.060 | NS |
| Butyrate | 6.49 ± 0.826 b | 7.14 ± 1.17 b | 12.2 ± 0.926 a | 12.1 ± 1.80 a | <0.005 |
| Isovalerate | 0.922 ± 0.067 | 1.13 ± 0.116 | 1.06 ± 0.073 | 0.923 ± 0.091 | NS |
| Valerate | 0.887 ± 0.096 | 1.01 ± 0.061 | 0.889 ± 0.054 | 0.843 ± 0.052 | NS |
| PSCFA | 2.62 ± 0.214 | 3.08 ± 0.234 | 2.90 ± 0.180 | 2.66 ± 0.180 | NS |
| SCFA total | 55.4 ± 4.02 c | 62.7 ± 2.65 b,c | 83.4 ± 5.58 a | 71.9 ± 5.38 a,b | <0.005 |
| Parameters | Groups | ANOVA p Value | |||
|---|---|---|---|---|---|
| C | CS | BK | QU | ||
| TG (mmol/L) | 3.16 ± 0.470 a | 1.25 ± 0.178 b | 1.68 ± 0.095 a,b | 1.38 ± 0.133 b | <0.005 |
| TC (mmol/L) | 2.80 ± 0.111 a | 2.41 ± 0.106 b | 2.08 ± 0.103 c | 1.92 ± 0.097 c | <0.001 |
| HDL (mmol/L) | 0.714 ± 0.027 | 0.906 ± 0.087 | 0.859 ± 0.107 | 0.894 ± 0.047 | NS |
| LDL (mmol/L) | 0.524 ± 0.035 a | 0.349 ± 0.040 b | 0.187 ± 0.022 c | 0.217 ± 0.021 c | <0.001 |
| Parameters | Groups | ANOVA p Value | |||
|---|---|---|---|---|---|
| C | CS | BK | QU | ||
| AST (U/L) | 63.5 ± 3.27 | 62.0 ± 0.933 | 61.4 ± 2.70 | 57.2 ± 2.33 | NS |
| ALT (U/L) | 18.9 ± 1.14 c | 20.5 ± 2.23 b,c | 24.6 ± 1.49 a,b | 26.4 ± 1.95 a | <0.05 |
| ALP (U/L) | 310 ± 27.0 b | 408 ± 18.4 a | 367 ± 24.2 ab | 451 ± 40.8 a | <0.05 |
| Albumin (µmol/L) | 400 ± 5.03 a | 371 ± 5.53 b | 388 ± 6.57 a | 367 ± 3.73 b | 0.001 |
| Uric acid (µmol/L) | 16.6 ± 1.94 | 20.7 ± 2.91 | 14.9 ± 1.84 | 16.0 ± 2.23 | NS |
| Creatinine (µmol/L) | 15.9 ± 2.51 | 17.4 ± 1.82 | 11.9 ± 1.61 | 14.4 ± 1.93 | NS |
| IL-6 (pg/mL) | 201 ± 1.50 b | 216 ± 8.00 a,b | 204 ± 5.55 a,b | 230 ± 7.54 a | <0.05 |
| IL-10 (pg/mL) | 48.7 ± 2.18 | 48.5 ± 2.84 | 53.0 ± 2.71 | 54.6 ± 2.01 | NS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fotschki, B.; Juśkiewicz, J.; Jurgoński, A.; Amarowicz, R.; Opyd, P.; Bez, J.; Muranyi, I.; Lykke Petersen, I.; Laparra Llopis, M. Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats. Nutrients 2020, 12, 2781. https://doi.org/10.3390/nu12092781
Fotschki B, Juśkiewicz J, Jurgoński A, Amarowicz R, Opyd P, Bez J, Muranyi I, Lykke Petersen I, Laparra Llopis M. Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats. Nutrients. 2020; 12(9):2781. https://doi.org/10.3390/nu12092781
Chicago/Turabian StyleFotschki, Bartosz, Jerzy Juśkiewicz, Adam Jurgoński, Ryszard Amarowicz, Paulina Opyd, Jürgen Bez, Isabel Muranyi, Iben Lykke Petersen, and Moisés Laparra Llopis. 2020. "Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats" Nutrients 12, no. 9: 2781. https://doi.org/10.3390/nu12092781
APA StyleFotschki, B., Juśkiewicz, J., Jurgoński, A., Amarowicz, R., Opyd, P., Bez, J., Muranyi, I., Lykke Petersen, I., & Laparra Llopis, M. (2020). Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats. Nutrients, 12(9), 2781. https://doi.org/10.3390/nu12092781