Sex-Dependent Wheel Running Effects on High Fat Diet Preference, Metabolic Outcomes, and Performance on the Barnes Maze in Rats


Abstract
1. Introduction
2. Materials and Methods
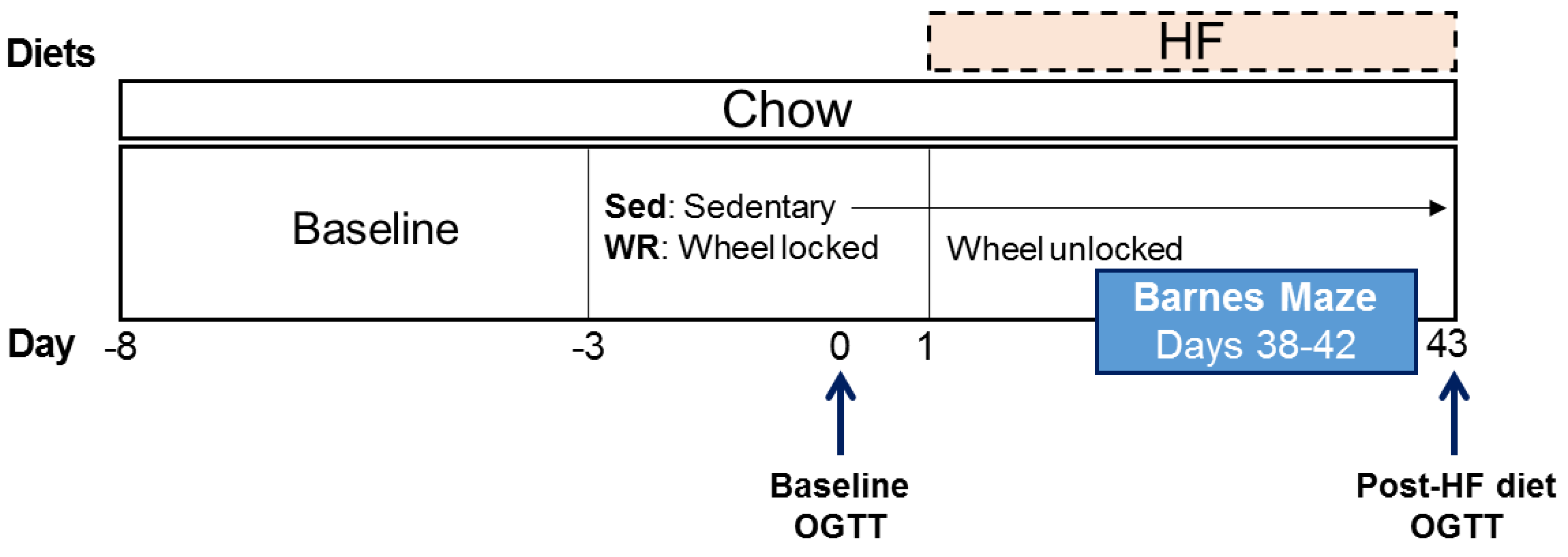
2.1. Subjects
2.2. Procedures
2.2.1. Wheel Running and Two-Diet Choice
2.2.2. Oral Glucose Tolerance Test (OGTT)
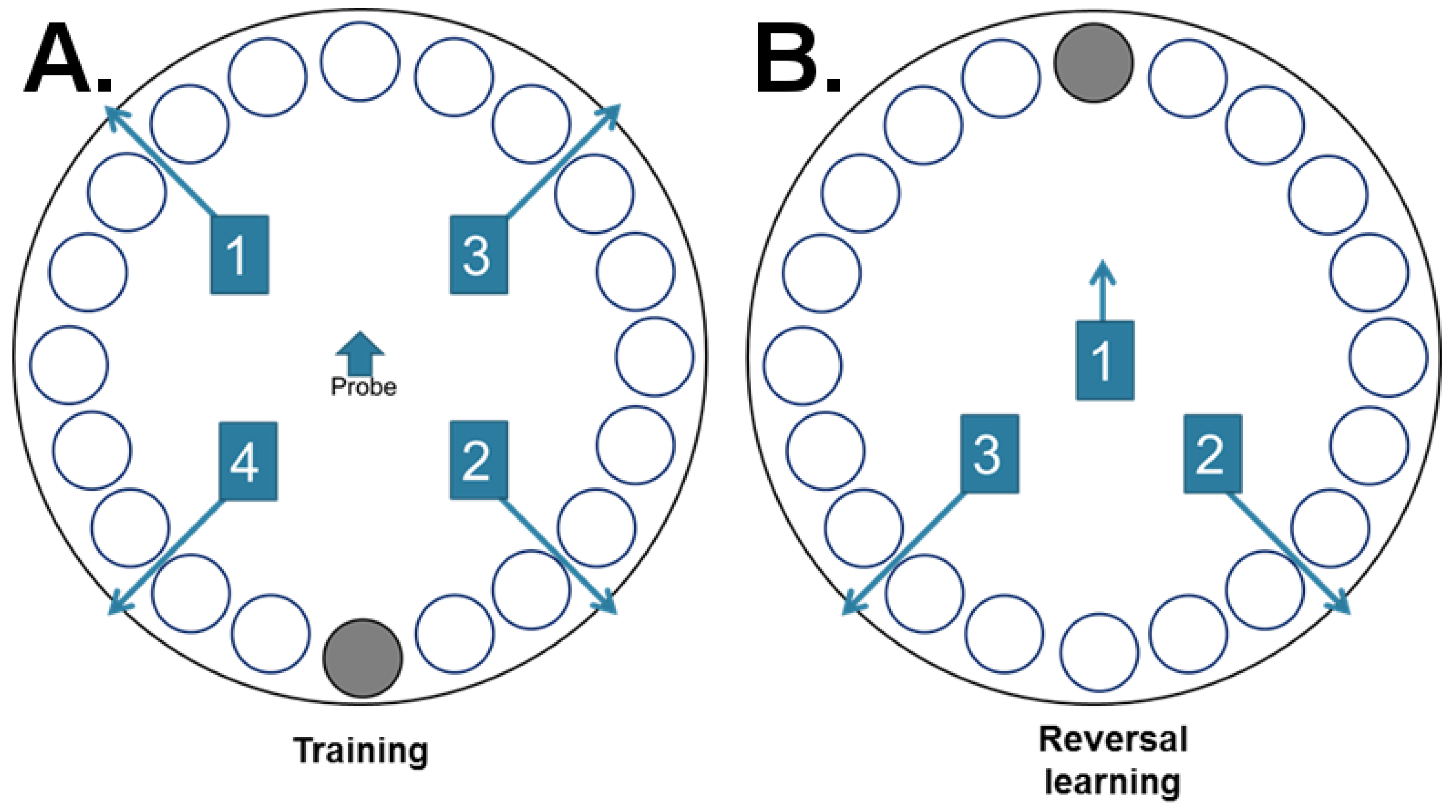
2.2.3. Barnes Maze
2.3. Statistical Analysis
3. Results
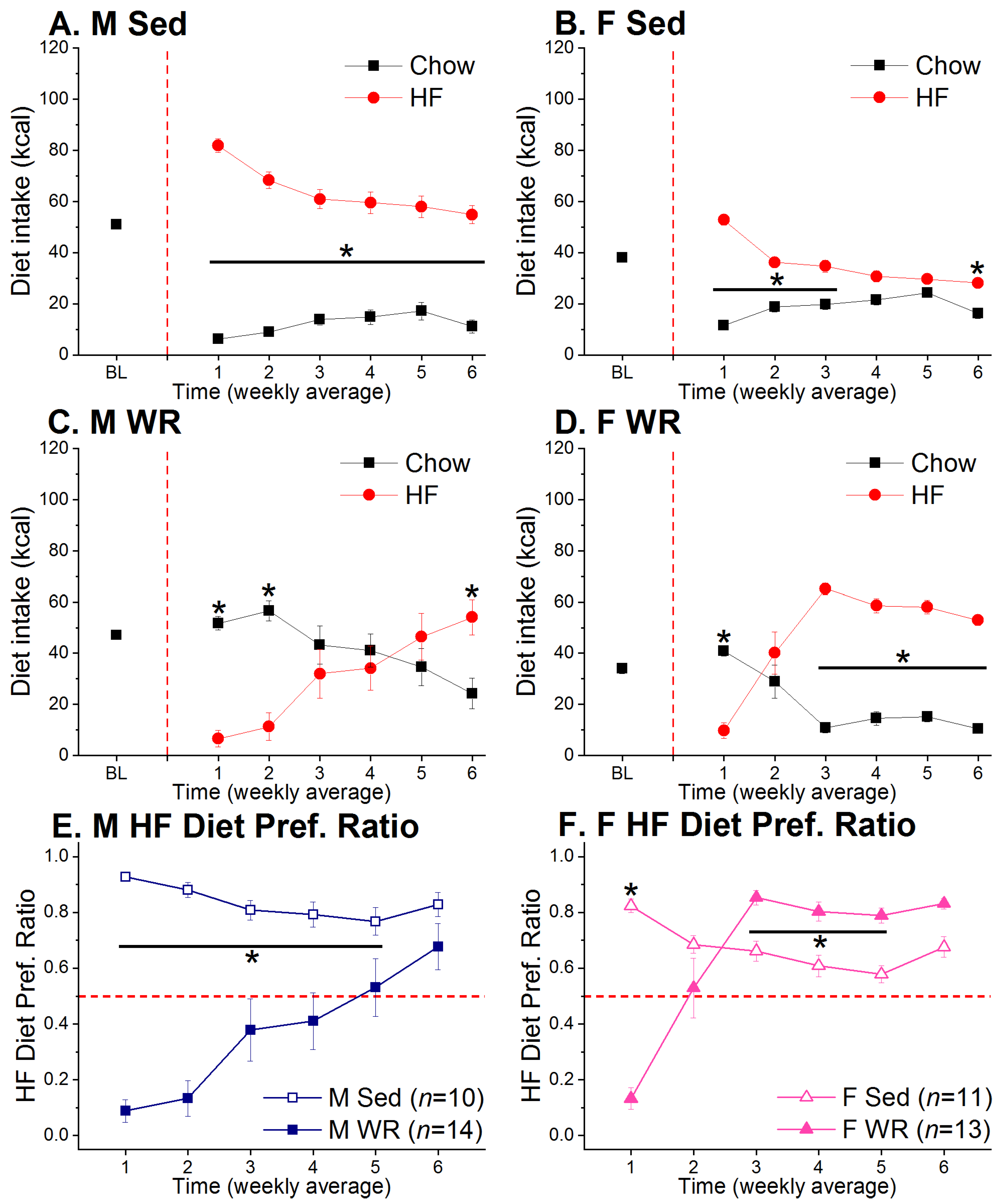
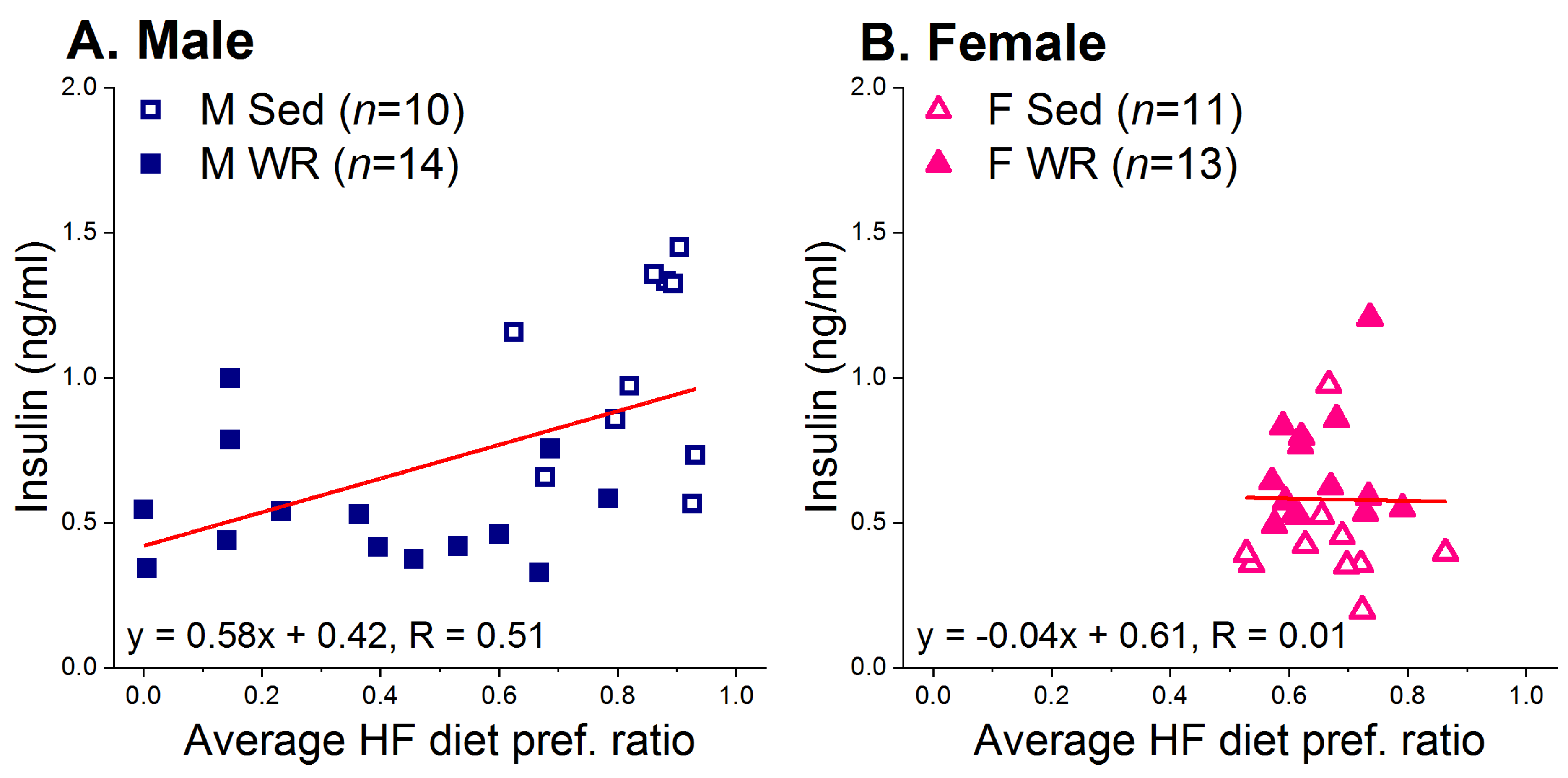
3.1. Wheel Running and Two-Diet Choice
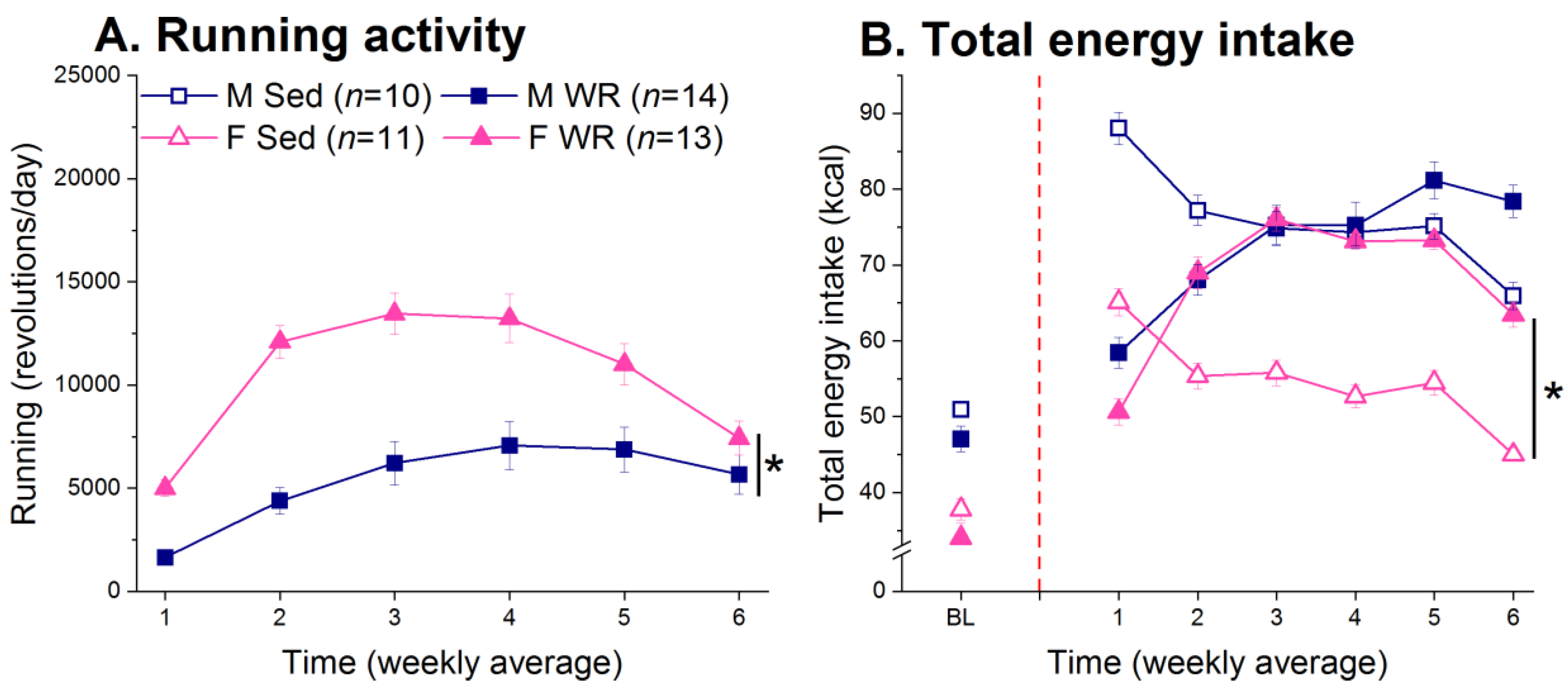
3.2. Running Activity and Energy Intake
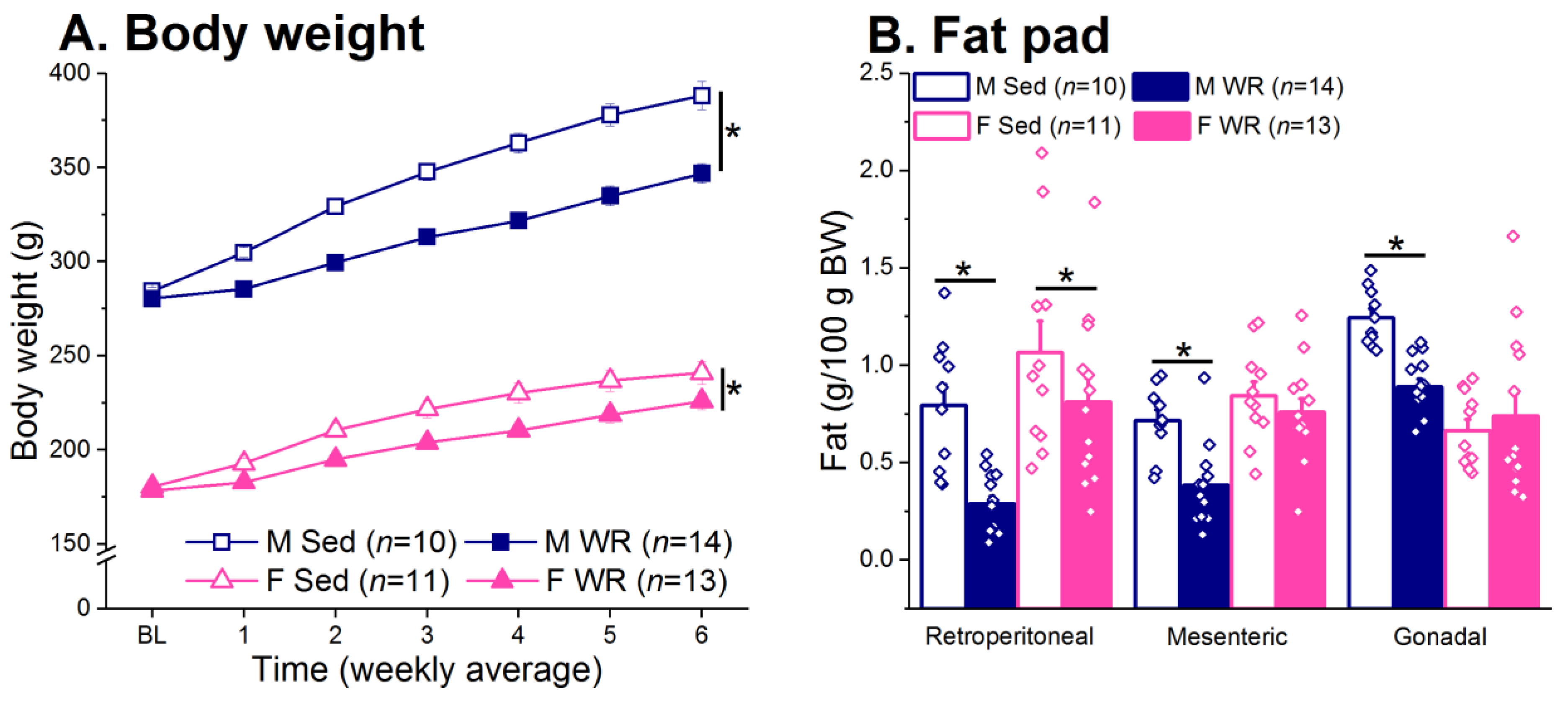
3.3. Body Weight and Adiposity
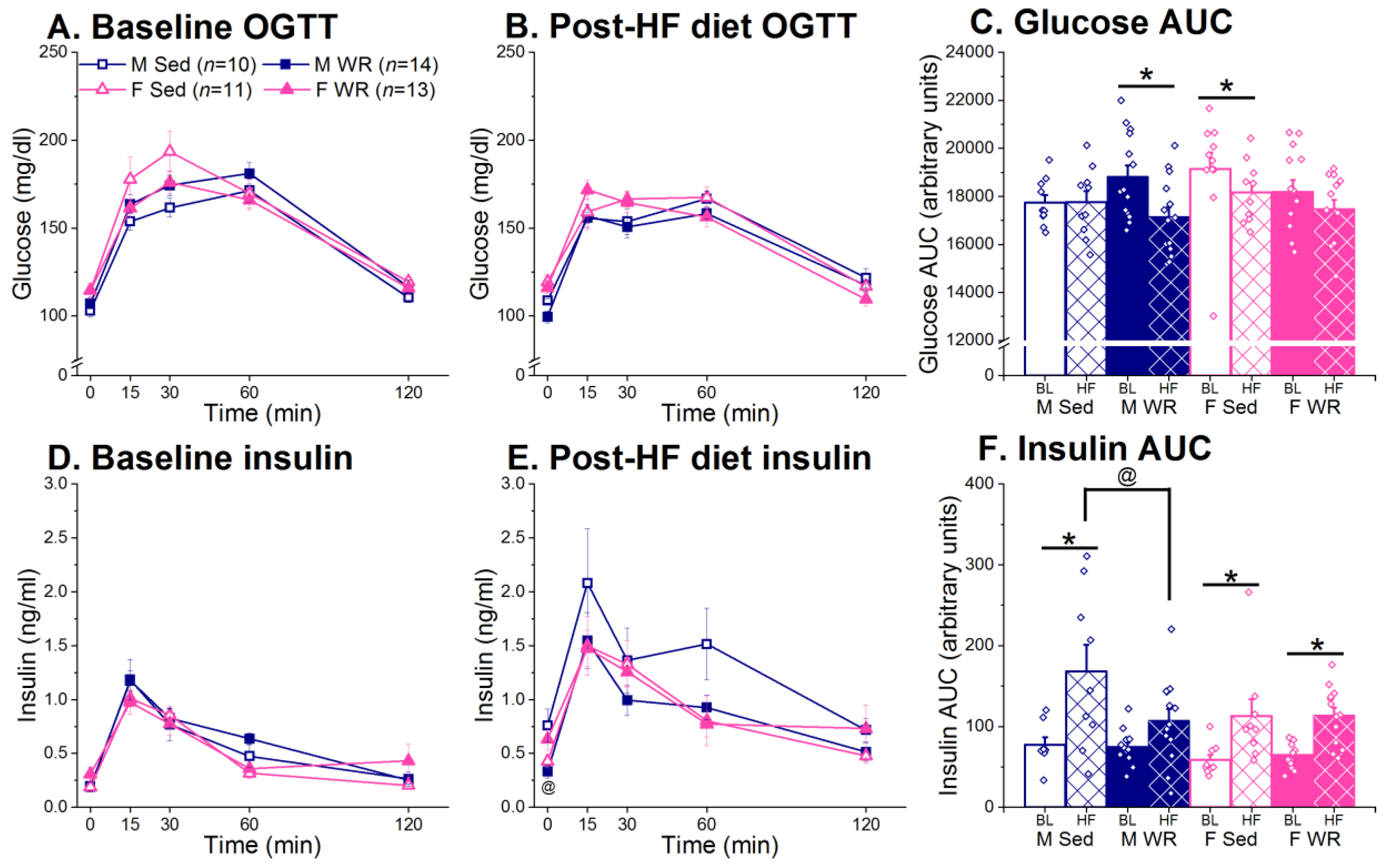
3.4. Oral Glucose Tolerance Test (OGTT)
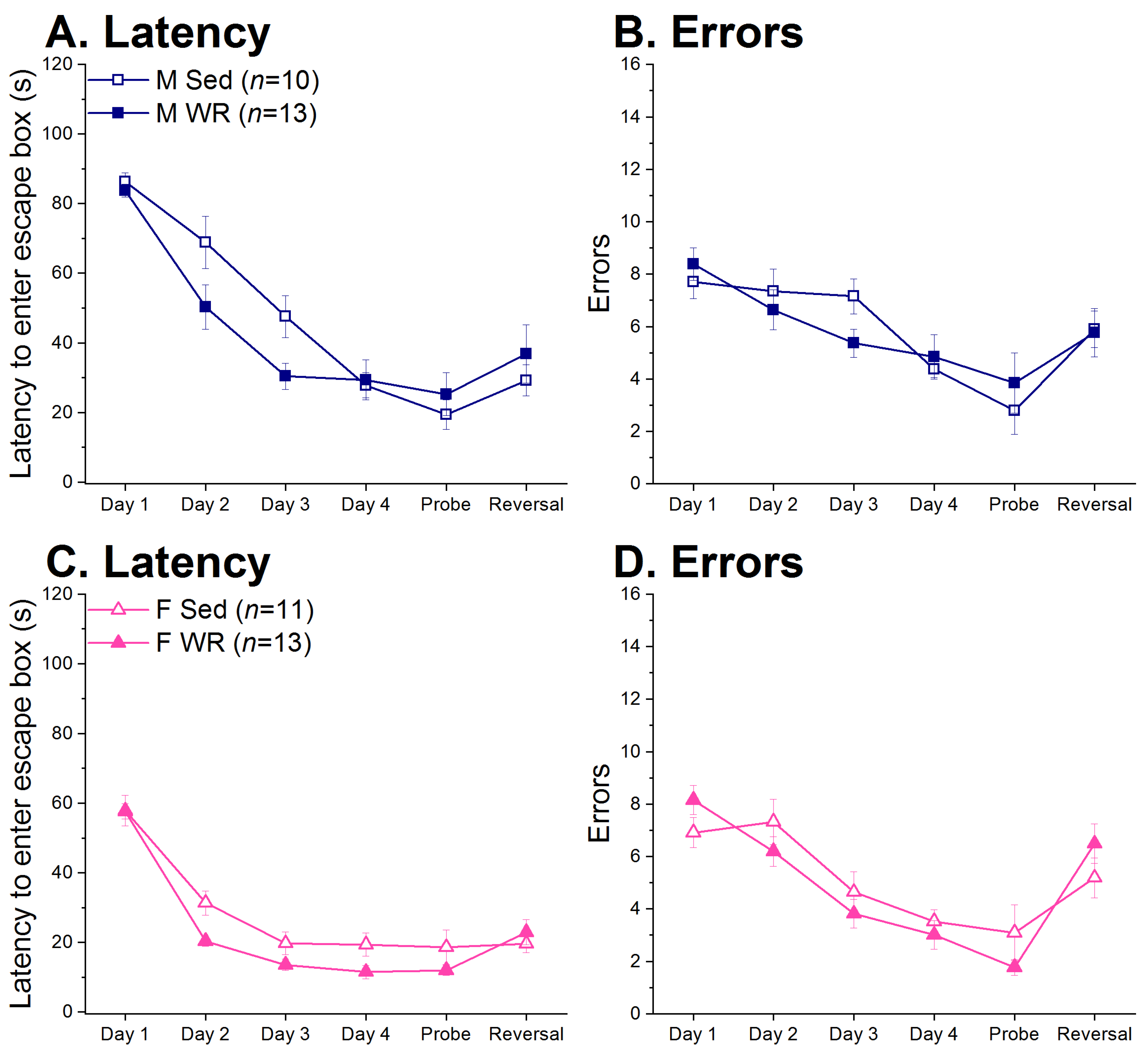
3.5. Barnes Maze
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hedley, A.A.; Ogden, C.L.; Johnson, C.L.; Carroll, M.D.; Curtin, L.R.; Flegal, K.M. Prevalence of overweight and obesity among US children, adolescents, and adults, 1999–2002. JAMA 2004, 291, 2847–2850. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, A.N.; Schnorr, S.L. Current views on hunter-gatherer nutrition and the evolution of the human diet. Am. J. Phys. Anthropol. 2017, 162 (Suppl. 63), 84–109. [Google Scholar] [CrossRef]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, E. Secular changes in the stature of adults and the secular trend of the modern sugar consumption. Z. Kinderheilkd 1967, 99, 146–166. [Google Scholar] [CrossRef] [PubMed]
- Shan, Z.; Rehm, C.D.; Rogers, G.; Ruan, M.; Wang, D.D.; Hu, F.B.; Mozaffarian, D.; Zhang, F.F.; Bhupathiraju, S.N. Trends in Dietary Carbohydrate, Protein, and Fat Intake and Diet Quality Among US Adults, 1999–2016. JAMA 2019, 322, 1178–1187. [Google Scholar] [CrossRef]
- Vadiveloo, M.; Scott, M.; Quatromoni, P.; Jacques, P.; Parekh, N. Trends in dietary fat and high-fat food intakes from 1991 to 2008 in the Framingham Heart Study participants. Br. J. Nutr. 2014, 111, 724–734. [Google Scholar] [CrossRef]
- Okreglicka, K. Health effects of changes in the structure of dietary macronutrients intake in western societies. Rocz. Panstw. Zakl. Hig. 2015, 66, 97–105. [Google Scholar]
- Foster-Powell, K.; Holt, S.H.; Brand-Miller, J.C. International table of glycemic index and glycemic load values: 2002. Am. J. Clin. Nutr. 2002, 76, 5–56. [Google Scholar] [CrossRef]
- Steenbergen, L.; Colzato, L.S. Overweight and Cognitive Performance: High Body Mass Index Is Associated with Impairment in Reactive Control during Task Switching. Front. Nutr. 2017, 4, 51. [Google Scholar] [CrossRef]
- Yang, Y.; Shields, G.S.; Guo, C.; Liu, Y. Executive function performance in obesity and overweight individuals: A meta-analysis and review. Neurosci. BioBehav. Rev. 2018, 84, 225–244. [Google Scholar] [CrossRef]
- Smith, E.; Hay, P.; Campbell, L.; Trollor, J.N. A review of the association between obesity and cognitive function across the lifespan: Implications for novel approaches to prevention and treatment. Obes. Rev. 2011, 12, 740–755. [Google Scholar] [CrossRef]
- Chen, F.T.; Chen, S.R.; Chu, I.H.; Liu, J.H.; Chang, Y.K. Multicomponent Exercise Intervention and Metacognition in Obese Preadolescents: A Randomized Controlled Study. J. Sport Exerc. Psychol. 2017, 39, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Pearson, M.; Kebejian, L.; Golden, E.; Keselman, A.; Bender, M.; Carlson, O.; Egan, J.; Ladenheim, B.; Cadet, J.L.; et al. Sex-dependent metabolic, neuroendocrine, and cognitive responses to dietary energy restriction and excess. Endocrinology 2007, 148, 4318–4333. [Google Scholar] [CrossRef] [PubMed]
- Winocur, G.; Greenwood, C.E.; Piroli, G.G.; Grillo, C.A.; Reznikov, L.R.; Reagan, L.P.; McEwen, B.S. Memory impairment in obese Zucker rats: An investigation of cognitive function in an animal model of insulin resistance and obesity. Behav. Neurosci. 2005, 119, 1389–1395. [Google Scholar] [CrossRef]
- Kanoski, S.E.; Meisel, R.L.; Mullins, A.J.; Davidson, T.L. The effects of energy-rich diets on discrimination reversal learning and on BDNF in the hippocampus and prefrontal cortex of the rat. Behav. Brain Res. 2007, 182, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Maesako, M.; Uemura, K.; Kubota, M.; Kuzuya, A.; Sasaki, K.; Hayashida, N.; Asada-Utsugi, M.; Watanabe, K.; Uemura, M.; Kihara, T.; et al. Exercise is more effective than diet control in preventing high fat diet-induced beta-amyloid deposition and memory deficit in amyloid precursor protein transgenic mice. J. Biol. Chem. 2012, 287, 23024–23033. [Google Scholar] [CrossRef] [PubMed]
- Klem, M.L.; Wing, R.R.; McGuire, M.T.; Seagle, H.M.; Hill, J.O. A descriptive study of individuals successful at long-term maintenance of substantial weight loss. Am. J. Clin. Nutr. 1997, 66, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Shick, S.M.; Wing, R.R.; Klem, M.L.; McGuire, M.T.; Hill, J.O.; Seagle, H. Persons successful at long-term weight loss and maintenance continue to consume a low-energy, low-fat diet. J. Am. Diet. Assoc. 1998, 98, 408–413. [Google Scholar] [CrossRef]
- Looy, H.; Eikelboom, R. Wheel running, food intake, and body weight in male rats. Physiol. Behav. 1989, 45, 403–405. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Scott, K.A.; Moran, T.H.; Bi, S. Dorsomedial hypothalamic corticotropin-releasing factor mediation of exercise-induced anorexia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1800–R1805. [Google Scholar] [CrossRef]
- Tokuyama, K.; Saito, M.; Okuda, H. Effects of wheel running on food intake and weight gain of male and female rats. Physiol. Behav. 1982, 28, 899–903. [Google Scholar] [CrossRef]
- Carrera, O.; Cerrato, M.; Vazquez, R.; Sineiro, C.; Gutierrez, E. Gender dimorphic effects of voluntary running in laboratory rats depends on maturational status. Q. J. Exp. Psychol. (Hove) 2011, 64, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Eckel, L.A.; Moore, S.R. Diet-induced hyperphagia in the rat is influenced by sex and exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R1080–R1085. [Google Scholar] [CrossRef] [PubMed]
- Blundell, J.E.; King, N.A. Physical activity and regulation of food intake: Current evidence. Med. Sci. Sports Exerc. 1999, 31, S573–S583. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, J.E.; Herrmann, S.D.; Lambourne, K.; Szabo, A.N.; Honas, J.J.; Washburn, R.A. Does increased exercise or physical activity alter ad-libitum daily energy intake or macronutrient composition in healthy adults? A systematic review. PLoS ONE 2014, 9, e83498. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Rahmani-Nia, F.; Damirchi, A.; Mirzaie, B.; Asghar Pur, S. Effect of Short-term Exercise on Appetite, Energy Intake and Energy-regulating Hormones. Iran. J. Basic Med. Sci. 2013, 16, 829–834. [Google Scholar]
- King, N.A.; Hopkins, M.; Caudwell, P.; Stubbs, R.J.; Blundell, J.E. Beneficial effects of exercise: Shifting the focus from body weight to other markers of health. Br. J. Sports Med. 2009, 43, 924–927. [Google Scholar] [CrossRef]
- Schubert, M.M.; Desbrow, B.; Sabapathy, S.; Leveritt, M. Acute exercise and subsequent energy intake. A meta-analysis. Appetite 2013, 63, 92–104. [Google Scholar] [CrossRef]
- Castro, E.A.; Carraca, E.V.; Cupeiro, R.; Lopez-Plaza, B.; Teixeira, P.J.; Gonzalez-Lamuno, D.; Peinado, A.B. The Effects of the Type of Exercise and Physical Activity on Eating Behavior and Body Composition in Overweight and Obese Subjects. Nutrients 2020, 12, 557. [Google Scholar] [CrossRef]
- Donnelly, J.E.; Smith, B.K. Is exercise effective for weight loss with ad libitum diet? Energy balance, compensation, and gender differences. Exerc. Sport Sci. Rev. 2005, 33, 169–174. [Google Scholar] [CrossRef]
- Anderson, J.W.; Konz, E.C.; Frederich, R.C.; Wood, C.L. Long-term weight-loss maintenance: A meta-analysis of US studies. Am. J. Clin. Nutr. 2001, 74, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Despres, J.P.; Bouchard, C.; Savard, R.; Tremblay, A.; Marcotte, M.; Theriault, G. The effect of a 20-week endurance training program on adipose-tissue morphology and lipolysis in men and women. Metabolism 1984, 33, 235–239. [Google Scholar] [CrossRef]
- Williams, R.L.; Wood, L.G.; Collins, C.E.; Callister, R. Effectiveness of weight loss interventions--is there a difference between men and women: A systematic review. Obes. Rev. 2015, 16, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Washburn, R.A.; Honas, J.J.; Ptomey, L.T.; Mayo, M.S.; Lee, J.; Sullivan, D.K.; Lambourne, K.; Willis, E.A.; Donnelly, J.E. Energy and Macronutrient Intake in the Midwest Exercise Trial 2 (MET-2). Med. Sci. Sports Exerc. 2015, 47, 1941–1949. [Google Scholar] [CrossRef] [PubMed]
- Westerterp, K.R.; Meijer, G.A.; Janssen, E.M.; Saris, W.H.; Ten Hoor, F. Long-term effect of physical activity on energy balance and body composition. Br. J. Nutr. 1992, 68, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Coker, R.H.; Williams, R.H.; Yeo, S.E.; Kortebein, P.M.; Bodenner, D.L.; Kern, P.A.; Evans, W.J. The impact of exercise training compared to caloric restriction on hepatic and peripheral insulin resistance in obesity. J. Clin. Endocrinol. Metab. 2009, 94, 4258–4266. [Google Scholar] [CrossRef] [PubMed]
- Nara, M.; Takahashi, M.; Kanda, T.; Shimomura, Y.; Kobayashi, I. Running exercise improves metabolic abnormalities and fat accumulation in sucrose-induced insulin-resistant rats. Obes. Res. 1997, 5, 348–353. [Google Scholar] [CrossRef]
- Greenwood, C.E.; Winocur, G. High-fat diets, insulin resistance and declining cognitive function. Neurobiol. Aging 2005, 26 (Suppl. 1), 42–45. [Google Scholar] [CrossRef]
- Winocur, G.; Greenwood, C.E. Studies of the effects of high fat diets on cognitive function in a rat model. Neurobiol. Aging 2005, 26 (Suppl. 1), 46–49. [Google Scholar] [CrossRef]
- Greenwood, C.E.; Winocur, G. Cognitive impairment in rats fed high-fat diets: A specific effect of saturated fatty-acid intake. Behav. Neurosci. 1996, 110, 451–459. [Google Scholar] [CrossRef]
- Winocur, G.; Greenwood, C.E. The effects of high fat diets and environmental influences on cognitive performance in rats. Behav. Brain Res. 1999, 101, 153–161. [Google Scholar] [CrossRef]
- Stranahan, A.M.; Norman, E.D.; Lee, K.; Cutler, R.G.; Telljohann, R.S.; Egan, J.M.; Mattson, M.P. Diet-induced insulin resistance impairs hippocampal synaptic plasticity and cognition in middle-aged rats. Hippocampus 2008, 18, 1085–1088. [Google Scholar] [CrossRef] [PubMed]
- Valladolid-Acebes, I.; Stucchi, P.; Cano, V.; Fernandez-Alfonso, M.S.; Merino, B.; Gil-Ortega, M.; Fole, A.; Morales, L.; Ruiz-Gayo, M.; Del Olmo, N. High-fat diets impair spatial learning in the radial-arm maze in mice. Neurobiol. Learn. Mem. 2011, 95, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Kanoski, S.E.; Davidson, T.L. Different patterns of memory impairments accompany short- and longer-term maintenance on a high-energy diet. J. Exp. Psychol. Anim. Behav. Process. 2010, 36, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Brockett, A.T.; LaMarca, E.A.; Gould, E. Physical exercise enhances cognitive flexibility as well as astrocytic and synaptic markers in the medial prefrontal cortex. PLoS ONE 2015, 10, e0124859. [Google Scholar] [CrossRef] [PubMed]
- Fordyce, D.E.; Farrar, R.P. Enhancement of spatial learning in F344 rats by physical activity and related learning-associated alterations in hippocampal and cortical cholinergic functioning. Behav. Brain Res. 1991, 46, 123–133. [Google Scholar] [CrossRef]
- Voss, M.W.; Vivar, C.; Kramer, A.F.; van Praag, H. Bridging animal and human models of exercise-induced brain plasticity. Trends Cogn. Sci. 2013, 17, 525–544. [Google Scholar] [CrossRef]
- Molteni, R.; Wu, A.; Vaynman, S.; Ying, Z.; Barnard, R.J.; Gomez-Pinilla, F. Exercise reverses the harmful effects of consumption of a high-fat diet on synaptic and behavioral plasticity associated to the action of brain-derived neurotrophic factor. Neuroscience 2004, 123, 429–440. [Google Scholar] [CrossRef]
- Klein, C.; Jonas, W.; Iggena, D.; Empl, L.; Rivalan, M.; Wiedmer, P.; Spranger, J.; Hellweg, R.; Winter, Y.; Steiner, B. Exercise prevents high-fat diet-induced impairment of flexible memory expression in the water maze and modulates adult hippocampal neurogenesis in mice. Neurobiol. Learn. Mem. 2016, 131, 26–35. [Google Scholar] [CrossRef]
- Noble, E.E.; Mavanji, V.; Little, M.R.; Billington, C.J.; Kotz, C.M.; Wang, C. Exercise reduces diet-induced cognitive decline and increases hippocampal brain-derived neurotrophic factor in CA3 neurons. Neurobiol. Learn. Mem. 2014, 114, 40–50. [Google Scholar] [CrossRef]
- Woo, J.; Shin, K.O.; Park, S.Y.; Jang, K.S.; Kang, S. Effects of exercise and diet change on cognition function and synaptic plasticity in high fat diet induced obese rats. Lipids Health Dis. 2013, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, T.E.; Pence, B.D.; Petr, G.; Ossyra, J.M.; Mach, H.C.; Bhattacharya, T.K.; Perez, S.; Martin, S.A.; McCusker, R.H.; Kelley, K.W.; et al. Voluntary wheel running, but not a diet containing (-)-epigallocatechin-3-gallate and beta-alanine, improves learning, memory and hippocampal neurogenesis in aged mice. Behav. Brain Res. 2014, 272, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Aslani, S.; Vieira, N.; Marques, F.; Costa, P.S.; Sousa, N.; Palha, J.A. The effect of high-fat diet on rat’s mood, feeding behavior and response to stress. Transl. Psychiatry 2015, 5, e684. [Google Scholar] [CrossRef] [PubMed]
- McNeilly, A.D.; Williamson, R.; Sutherland, C.; Balfour, D.J.; Stewart, C.A. High fat feeding promotes simultaneous decline in insulin sensitivity and cognitive performance in a delayed matching and non-matching to position task. Behav. Brain Res. 2011, 217, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Bocarsly, M.E.; Fasolino, M.; Kane, G.A.; LaMarca, E.A.; Kirschen, G.W.; Karatsoreos, I.N.; McEwen, B.S.; Gould, E. Obesity diminishes synaptic markers, alters microglial morphology, and impairs cognitive function. Proc. Natl. Acad. Sci. USA 2015, 112, 15731–15736. [Google Scholar] [CrossRef]
- Edwards, C.G.; Walk, A.M.; Thompson, S.V.; Mullen, S.P.; Holscher, H.D.; Khan, N.A. Disordered Eating Attitudes and Behavioral and Neuroelectric Indices of Cognitive Flexibility in Individuals with Overweight and Obesity. Nutrients 2018, 10, 1902. [Google Scholar] [CrossRef]
- Perpina, C.; Segura, M.; Sanchez-Reales, S. Cognitive flexibility and decision-making in eating disorders and obesity. Eat. Weight Disord. 2017, 22, 435–444. [Google Scholar] [CrossRef]
- McNeilly, A.D.; Gao, A.; Hill, A.Y.; Gomersall, T.; Balfour, D.J.K.; Sutherland, C.; Stewart, C.A. The effect of dietary intervention on the metabolic and behavioural impairments generated by short term high fat feeding in the rat. Physiol. Behav. 2016, 167, 100–109. [Google Scholar] [CrossRef][Green Version]
- Boitard, C.; Parkes, S.L.; Cavaroc, A.; Tantot, F.; Castanon, N.; Laye, S.; Tronel, S.; Pacheco-Lopez, G.; Coutureau, E.; Ferreira, G. Switching Adolescent High-Fat Diet to Adult Control Diet Restores Neurocognitive Alterations. Front. Behav. Neurosci. 2016, 10, 225. [Google Scholar] [CrossRef]
- McNeilly, A.D.; Williamson, R.; Balfour, D.J.; Stewart, C.A.; Sutherland, C. A high-fat-diet-induced cognitive deficit in rats that is not prevented by improving insulin sensitivity with metformin. Diabetologia 2012, 55, 3061–3070. [Google Scholar] [CrossRef]
- Amengual-Cladera, E.; Llado, I.; Gianotti, M.; Proenza, A.M. Sex differences in the effect of high-fat diet feeding on rat white adipose tissue mitochondrial function and insulin sensitivity. Metabolism 2012, 61, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Barron, A.M.; Rosario, E.R.; Elteriefi, R.; Pike, C.J. Sex-specific effects of high fat diet on indices of metabolic syndrome in 3xTg-AD mice: Implications for Alzheimer’s disease. PLoS ONE 2013, 8, e78554. [Google Scholar] [CrossRef] [PubMed]
- Elias, M.F.; Elias, P.K.; Sullivan, L.M.; Wolf, P.A.; D’Agostino, R.B. Lower cognitive function in the presence of obesity and hypertension: The Framingham heart study. Int. J. Obes. Relat. Metab. Disord. 2003, 27, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Underwood, E.L.; Thompson, L.T. High-fat diet impairs spatial memory and hippocampal intrinsic excitability and sex-dependently alters circulating insulin and hippocampal insulin sensitivity. Biol. Sex Differ. 2016, 7, 9. [Google Scholar] [CrossRef]
- Shields, G.S.; Trainor, B.C.; Lam, J.C.; Yonelinas, A.P. Acute stress impairs cognitive flexibility in men, not women. Stress 2016, 19, 542–546. [Google Scholar] [CrossRef]
- Laredo, S.A.; Steinman, M.Q.; Robles, C.F.; Ferrer, E.; Ragen, B.J.; Trainor, B.C. Effects of defeat stress on behavioral flexibility in males and females: Modulation by the mu-opioid receptor. Eur. J. Neurosci. 2015, 41, 434–441. [Google Scholar] [CrossRef]
- Hwang, L.L.; Wang, C.H.; Li, T.L.; Chang, S.D.; Lin, L.C.; Chen, C.P.; Chen, C.T.; Liang, K.C.; Ho, I.K.; Yang, W.S.; et al. Sex differences in high-fat diet-induced obesity, metabolic alterations and learning, and synaptic plasticity deficits in mice. Obesity (Silver Spring) 2010, 18, 463–469. [Google Scholar] [CrossRef]
- Li, W.; Qiu, Q.; Sun, L.; Yue, L.; Wang, T.; Li, X.; Xiao, S. Sex differences in obesity and cognitive function in a cognitively normal aging Chinese Han population. Neuropsychiatr. Dis. Treat. 2017, 13, 2405–2410. [Google Scholar] [CrossRef]
- Gollisch, K.S.; Brandauer, J.; Jessen, N.; Toyoda, T.; Nayer, A.; Hirshman, M.F.; Goodyear, L.J. Effects of exercise training on subcutaneous and visceral adipose tissue in normal- and high-fat diet-fed rats. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E495–E504. [Google Scholar] [CrossRef]
- Tokuyama, K.; Suzuki, M. Intravenous glucose tolerance test-derived glucose effectiveness in endurance-trained rats. Metabolism 1998, 47, 190–194. [Google Scholar] [CrossRef]
- Goodyear, L.J.; Hirshman, M.F.; Knutson, S.M.; Horton, E.D.; Horton, E.S. Effect of exercise training on glucose homeostasis in normal and insulin-deficient diabetic rats. J. Appl. Physiol. (1985) 1988, 65, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, J.L.; Dunford, E.C.; Leclair, E.; Mandel, E.R.; Peckett, A.J.; Haas, T.L.; Riddell, M.C. Voluntary exercise improves metabolic profile in high-fat fed glucocorticoid-treated rats. J. Appl. Physiol. (1985) 2015, 118, 1331–1343. [Google Scholar] [CrossRef] [PubMed]
- Bongbele, J.; Gutierrez, A.; Cardin, S.; Lavoie, J.M. Effect of physical training on insulin response to intravenous glucose in male peripubertal rats. J. Appl. Physiol. (1985) 1992, 73, 1227–1231. [Google Scholar] [CrossRef] [PubMed]
- Caudwell, P.; Gibbons, C.; Finlayson, G.; Naslund, E.; Blundell, J. Exercise and weight loss: No sex differences in body weight response to exercise. Exerc. Sport Sci. Rev. 2014, 42, 92–101. [Google Scholar] [CrossRef]
- Donnelly, J.E.; Smith, B.; Jacobsen, D.J.; Kirk, E.; Dubose, K.; Hyder, M.; Bailey, B.; Washburn, R. The role of exercise for weight loss and maintenance. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 1009–1029. [Google Scholar] [CrossRef]
- Hill, J.O.; Thiel, J.; Heller, P.A.; Markon, C.; Fletcher, G.; DiGirolamo, M. Differences in effects of aerobic exercise training on blood lipids in men and women. Am. J. Cardiol. 1989, 63, 254–256. [Google Scholar] [CrossRef]
- Buettner, R.; Scholmerich, J.; Bollheimer, L.C. High-fat diets: Modeling the metabolic disorders of human obesity in rodents. Obesity (Silver Spring) 2007, 15, 798–808. [Google Scholar] [CrossRef]
- Gawel, K.; Gibula, E.; Marszalek-Grabska, M.; Filarowska, J.; Kotlinska, J.H. Assessment of spatial learning and memory in the Barnes maze task in rodents-methodological consideration. Naunyn. Schmiedebergs Arch. Pharmacol. 2019, 392, 1–18. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Research Council: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Yang, T.Y.; Liang, N.C. Ovarian hormones mediate running-induced changes in high fat diet choice patterns in female rats. Horm. Behav. 2018, 100, 81–93. [Google Scholar] [CrossRef]
- Bowe, J.E.; Franklin, Z.J.; Hauge-Evans, A.C.; King, A.J.; Persaud, S.J.; Jones, P.M. Metabolic phenotyping guidelines: Assessing glucose homeostasis in rodent models. J. Endocrinol. 2014, 222, G13–G25. [Google Scholar] [CrossRef]
- Ayala, J.E.; Bracy, D.P.; McGuinness, O.P.; Wasserman, D.H. Considerations in the design of hyperinsulinemic-euglycemic clamps in the conscious mouse. Diabetes 2006, 55, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.; Gupta, N.; Goh, T.; Naigamwalla, D.; Chia, M.C.; Koohestani, N.; Mehrotra, S.; McKeown-Eyssen, G.; Giacca, A.; Bruce, W.R. Direct measure of insulin sensitivity with the hyperinsulinemic-euglycemic clamp and surrogate measures of insulin sensitivity with the oral glucose tolerance test: Correlations with aberrant crypt foci promotion in rats. Cancer Epidemiol. Biomarkers Prev. 2003, 12, 47–56. [Google Scholar] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Gutt, M.; Davis, C.L.; Spitzer, S.B.; Llabre, M.M.; Kumar, M.; Czarnecki, E.M.; Schneiderman, N.; Skyler, J.S.; Marks, J.B. Validation of the insulin sensitivity index (ISI(0,120)): Comparison with other measures. Diabetes Res. Clin. Pract. 2000, 47, 177–184. [Google Scholar] [CrossRef]
- O’Leary, T.P.; Savoie, V.; Brown, R.E. Learning, memory and search strategies of inbred mouse strains with different visual abilities in the Barnes maze. Behav. Brain Res. 2011, 216, 531–542. [Google Scholar] [CrossRef]
- Rosenfeld, C.S.; Ferguson, S.A. Barnes maze testing strategies with small and large rodent models. J. Vis. Exp. 2014, e51194. [Google Scholar] [CrossRef]
- Brooks, G.A. Importance of the ’crossover’ concept in exercise metabolism. Clin. Exp. Pharmacol. Physiol. 1997, 24, 889–895. [Google Scholar] [CrossRef]
- Horowitz, J.F. Fatty acid mobilization from adipose tissue during exercise. Trends Endocrinol. Metab. 2003, 14, 386–392. [Google Scholar] [CrossRef]
- Ruby, B.C.; Robergs, R.A. Gender differences in substrate utilisation during exercise. Sports Med. 1994, 17, 393–410. [Google Scholar] [CrossRef]
- Horton, T.J.; Pagliassotti, M.J.; Hobbs, K.; Hill, J.O. Fuel metabolism in men and women during and after long-duration exercise. J. Appl. Physiol. (1985) 1998, 85, 1823–1832. [Google Scholar] [CrossRef]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover” concept. J. Appl. Physiol. (1985) 1994, 76, 2253–2261. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.C.; Bello, N.T.; Moran, T.H. Wheel running reduces high-fat diet intake, preference and mu-opioid agonist stimulated intake. Behav. Brain Res. 2015, 284, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Moody, L.; Liang, J.; Choi, P.P.; Moran, T.H.; Liang, N.C. Wheel running decreases palatable diet preference in Sprague-Dawley rats. Physiol. Behav. 2015, 150, 53–63. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, T.Y.; Gardner, J.C.; Gao, Z.; Pan, Y.X.; Liang, N.C. Role of glucocorticoid signaling in exercise-associated changes in high-fat diet preference in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R515–R528. [Google Scholar] [CrossRef]
- Ellis, G.S.; Lanza-Jacoby, S.; Gow, A.; Kendrick, Z.V. Effects of estradiol on lipoprotein lipase activity and lipid availability in exercised male rats. J. Appl. Physiol (1985) 1994, 77, 209–215. [Google Scholar] [CrossRef]
- Roepstorff, C.; Steffensen, C.H.; Madsen, M.; Stallknecht, B.; Kanstrup, I.L.; Richter, E.A.; Kiens, B. Gender differences in substrate utilization during submaximal exercise in endurance-trained subjects. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E435–E447. [Google Scholar] [CrossRef]
- Carter, S.L.; Rennie, C.; Tarnopolsky, M.A. Substrate utilization during endurance exercise in men and women after endurance training. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E898–E907. [Google Scholar] [CrossRef]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. 1993, 265, E380–E391. [Google Scholar] [CrossRef]
- Hawley, J.A.; Hopkins, W.G. Aerobic glycolytic and aerobic lipolytic power systems. A new paradigm with implications for endurance and ultraendurance events. Sports Med. 1995, 19, 240–250. [Google Scholar] [CrossRef]
- Hudson, J.I.; Hiripi, E.; Pope, H.G., Jr.; Kessler, R.C. The prevalence and correlates of eating disorders in the National Comorbidity Survey Replication. Biol. Psychiatry 2007, 61, 348–358. [Google Scholar] [CrossRef]
- Klump, K.L.; Racine, S.; Hildebrandt, B.; Sisk, C.L. Sex differences in binge eating patterns in male and female adult rats. Int. J. Eat. Disord. 2013, 46, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Tapia, M.A.; Lee, J.R.; Weise, V.N.; Tamasi, A.M.; Will, M.J. Sex differences in hedonic and homeostatic aspects of palatable food motivation. Behav. Brain Res. 2019, 359, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Buczek, L.; Migliaccio, J.; Petrovich, G.D. Hedonic Eating: Sex Differences and Characterization of Orexin Activation and Signaling. Neuroscience 2020, 436, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980-2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Lynch, W.J. Sex differences in vulnerability to drug self-administration. Exp. Clin. Psychopharmacol. 2006, 14, 34–41. [Google Scholar] [CrossRef]
- Carroll, M.E.; Anker, J.J. Sex differences and ovarian hormones in animal models of drug dependence. Horm. Behav. 2010, 58, 44–56. [Google Scholar] [CrossRef]
- Day, C.J.; McHale, S.; Francis, J. Individual differences and preference for dietary fat using the Fat Preference Questionnaire((c)) in a UK sample. Appetite 2012, 58, 679–686. [Google Scholar] [CrossRef]
- Belke, T.W.; Wagner, J.P. The reinforcing property and the rewarding aftereffect of wheel running in rats: A combination of two paradigms. Behav. Process. 2005, 68, 165–172. [Google Scholar] [CrossRef]
- Spierling, S.R.; Kreisler, A.D.; Williams, C.A.; Fang, S.Y.; Pucci, S.N.; Kines, K.T.; Zorrilla, E.P. Intermittent, extended access to preferred food leads to escalated food reinforcement and cyclic whole-body metabolism in rats: Sex differences and individual vulnerability. Physiol. Behav. 2018, 192, 3–16. [Google Scholar] [CrossRef]
- Peterson, A.B.; Hivick, D.P.; Lynch, W.J. Dose-dependent effectiveness of wheel running to attenuate cocaine-seeking: Impact of sex and estrous cycle in rats. Psychopharmacology (Berl) 2014, 231, 2661–2670. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, M.; Zhou, C.; Li, R. Sex differences in drug addiction and response to exercise intervention: From human to animal studies. Front. Neuroendocrinol. 2016, 40, 24–41. [Google Scholar] [CrossRef]
- Pitts, G.C. Body composition in the rat: Interactions of exercise, age, sex, and diet. Am. J. Physiol. 1984, 246, R495–R501. [Google Scholar] [CrossRef] [PubMed]
- Foright, R.M.; Johnson, G.C.; Kahn, D.; Charleston, C.A.; Presby, D.M.; Bouchet, C.A.; Wellberg, E.A.; Sherk, V.D.; Jackman, M.R.; Greenwood, B.N.; et al. Compensatory eating behaviors in male and female rats in response to exercise training. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Latour, M.G.; Shinoda, M.; Lavoie, J.M. Metabolic effects of physical training in ovariectomized and hyperestrogenic rats. J. Appl. Physiol. (1985) 2001, 90, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Lamontagne, J.; Jalbert-Arsenault, E.; Pepin, E.; Peyot, M.L.; Ruderman, N.B.; Nolan, C.J.; Joly, E.; Madiraju, S.R.; Poitout, V.; Prentki, M. Pioglitazone acutely reduces energy metabolism and insulin secretion in rats. Diabetes 2013, 62, 2122–2129. [Google Scholar] [CrossRef] [PubMed]
- Vallerand, A.L.; Lupien, J.; Deshaies, Y.; Bukowiecki, L.J. Intensive exercise training does not improve intravenous glucose tolerance in severely diabetic rats. Horm. Metab. Res. 1986, 18, 79–81. [Google Scholar] [CrossRef]
- Goodyear, L.J.; Kahn, B.B. Exercise, glucose transport, and insulin sensitivity. Annu. Rev. Med. 1998, 49, 235–261. [Google Scholar] [CrossRef]
- Goodyear, L.J.; Hirshman, M.F.; Horton, E.D.; Knutson, S.M.; Wardzala, L.J.; Horton, E.S. Exercise training normalizes glucose metabolism in a rat model of impaired glucose tolerance. Metabolism 1991, 40, 455–464. [Google Scholar] [CrossRef]
- LaPier, T.L.; Swislocki, A.L.; Clark, R.J.; Rodnick, K.J. Voluntary running improves glucose tolerance and insulin resistance in female spontaneously hypertensive rats. Am. J. Hypertens 2001, 14, 708–715. [Google Scholar] [CrossRef]
- Ascaso, J.F.; Pardo, S.; Real, J.T.; Lorente, R.I.; Priego, A.; Carmena, R. Diagnosing insulin resistance by simple quantitative methods in subjects with normal glucose metabolism. Diabetes Care 2003, 26, 3320–3325. [Google Scholar] [CrossRef]
- Swift, D.L.; McGee, J.E.; Earnest, C.P.; Carlisle, E.; Nygard, M.; Johannsen, N.M. The Effects of Exercise and Physical Activity on Weight Loss and Maintenance. Prog. Cardiovasc. Dis. 2018, 61, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Villanueva Espino, L.A.; Silva Gomez, A.B.; Bravo Duran, D.A. Cognitive training increases dendritic arborization in the dorsal hippocampal CA1 and CA3 neurons of female and male Long-Evans rats. Synapse 2020, 74, e22140. [Google Scholar] [CrossRef] [PubMed]
- Betancourt, E.; Wachtel, J.; Michaelos, M.; Haggerty, M.; Conforti, J.; Kritzer, M.F. The impact of biological sex and sex hormones on cognition in a rat model of early, pre-motor Parkinson’s disease. Neuroscience 2017, 345, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Beatty, W.W. Gonadal hormones and sex differences in nonreproductive behaviors in rodents: Organizational and activational influences. Horm. Behav. 1979, 12, 112–163. [Google Scholar] [CrossRef]
- Tropp, J.; Markus, E.J. Sex differences in the dynamics of cue utilization and exploratory behavior. Behav. Brain Res. 2001, 119, 143–154. [Google Scholar] [CrossRef]
- Jonasson, Z. Meta-analysis of sex differences in rodent models of learning and memory: A review of behavioral and biological data. Neurosci. BioBehav. Rev. 2005, 28, 811–825. [Google Scholar] [CrossRef]
- Locklear, M.N.; Kritzer, M.F. Assessment of the effects of sex and sex hormones on spatial cognition in adult rats using the Barnes maze. Horm. Behav. 2014, 66, 298–308. [Google Scholar] [CrossRef]
- Bucci, D.J.; Chiba, A.A.; Gallagher, M. Spatial learning in male and female Long-Evans rats. Behav. Neurosci. 1995, 109, 180–183. [Google Scholar] [CrossRef]
- Koss, W.A.; Frick, K.M. Sex differences in hippocampal function. J. Neurosci. Res. 2017, 95, 539–562. [Google Scholar] [CrossRef]
- Barrett, G.L.; Bennie, A.; Trieu, J.; Ping, S.; Tsafoulis, C. The chronology of age-related spatial learning impairment in two rat strains, as tested by the Barnes maze. Behav. Neurosci. 2009, 123, 533–538. [Google Scholar] [CrossRef]
- Sadeghian, A.; Fathollahi, Y.; Javan, M.; Shojaei, A.; Kosarmadar, N.; Rezaei, M.; Mirnajafi-Zadeh, J. Spatial Learning and Memory in Barnes Maze Test and Synaptic Potentiation in Schaffer Collateral-CA1 Synapses of Dorsal Hippocampus in Freely Moving Rts. Basic Clin. Neurosci. 2019, 10, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.A.; Choudhury, K.R.; Rathakrishnan, B.G.; Marks, D.M.; Petrella, J.R.; Doraiswamy, P.M.; Alzheimer’s Disease Neuroimaging, I. Marked gender differences in progression of mild cognitive impairment over 8 years. Alzheimers Dement. (NY) 2015, 1, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S.; Wolf, P.A.; Beiser, A.; Au, R.; McNulty, K.; White, R.; D’Agostino, R.B. Lifetime risk of dementia and Alzheimer’s disease. The impact of mortality on risk estimates in the Framingham Study. Neurology 1997, 49, 1498–1504. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Macronutrient | Description | Unit | Teklad 2018 (3.1 kcal/g) | 45% HF Diet (4.74 kcal/g) |
|---|---|---|---|---|
| Protein | Total | % kcal | 24 | 20 |
| Carbohydrate | Sucrose | % kcal | - | 17 |
| Other carbohydrates | % kcal | 58 | 18 | |
| Fat | Total | % kcal | 18 | 45 |
| Saturated fats | % of total fat (wt) | 0.9 | 31.4 | |
| Monounsaturated fats | % of total fat (wt) | 1.3 | 35.5 | |
| Polyunsaturated fats | % of total fat (wt) | 3.4 | 33.1 |
| Insulin Sensitivity | HOMA-IR | ISI0,120 | |||
|---|---|---|---|---|---|
| Group | Baseline | Post-HF | Baseline | Post-HF | |
| Male Sed | 1.22 ± 0.31 * | 5.01 ± 0.99 ^ | 0.79 ± 0.04 * | 0.63 ± 0.02 ^ | |
| Male WR | 1.34 ± 0.30 | 2.15 ± 0.45 | 0.74 ± 0.03 | 0.71 ± 0.03 | |
| Female Sed | 1.43 ± 0.38 * | 3.15 ± 0.47 | 0.71 ±0.03 | 0.64 ± 0.02 | |
| Female WR | 2.25 ± 0.49 * | 4.63 ± 0.81 | 0.70 ± 0.03 | 0.65 ± 0.02 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, T.Y.; Gao, Z.; Liang, N.-C. Sex-Dependent Wheel Running Effects on High Fat Diet Preference, Metabolic Outcomes, and Performance on the Barnes Maze in Rats. Nutrients 2020, 12, 2721. https://doi.org/10.3390/nu12092721
Yang TY, Gao Z, Liang N-C. Sex-Dependent Wheel Running Effects on High Fat Diet Preference, Metabolic Outcomes, and Performance on the Barnes Maze in Rats. Nutrients. 2020; 12(9):2721. https://doi.org/10.3390/nu12092721
Chicago/Turabian StyleYang, Tiffany Y., Zijun Gao, and Nu-Chu Liang. 2020. "Sex-Dependent Wheel Running Effects on High Fat Diet Preference, Metabolic Outcomes, and Performance on the Barnes Maze in Rats" Nutrients 12, no. 9: 2721. https://doi.org/10.3390/nu12092721
APA StyleYang, T. Y., Gao, Z., & Liang, N.-C. (2020). Sex-Dependent Wheel Running Effects on High Fat Diet Preference, Metabolic Outcomes, and Performance on the Barnes Maze in Rats. Nutrients, 12(9), 2721. https://doi.org/10.3390/nu12092721