A Review of the Potential Interaction of Selenium and Iodine on Placental and Child Health



Abstract
1. Introduction
2. Oxidative Stress and the Importance of Antioxidants
Oxidative Stress and Pregnancy Complications
3. Micronutrients and Pregnancy
3.1. Selenium
3.2. Selenium and Pregnancy Complications
3.3. Iodine
3.4. Iodine and Pregnancy Complications
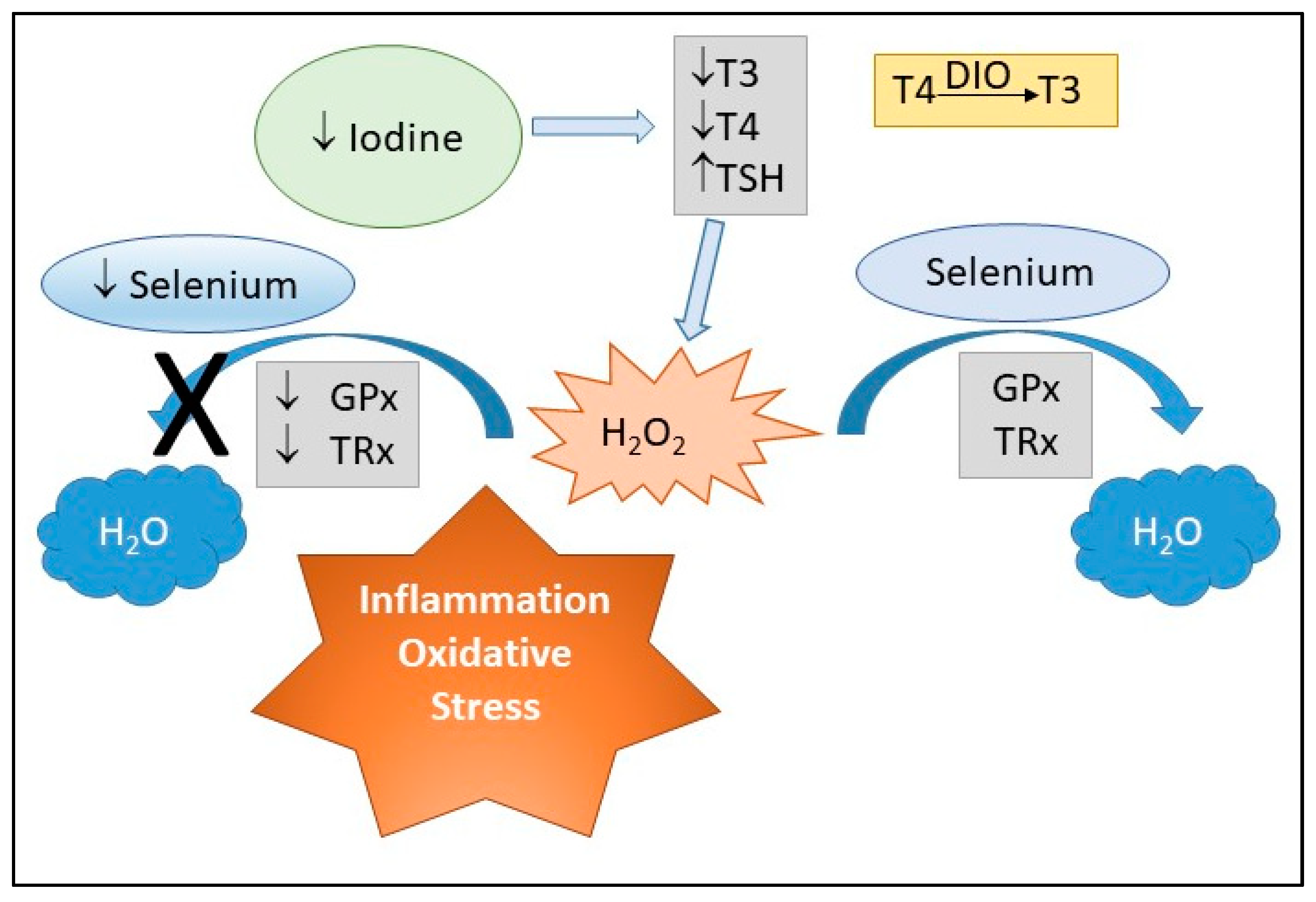
3.5. Potential Synergistic Effects of Selenium and Iodine
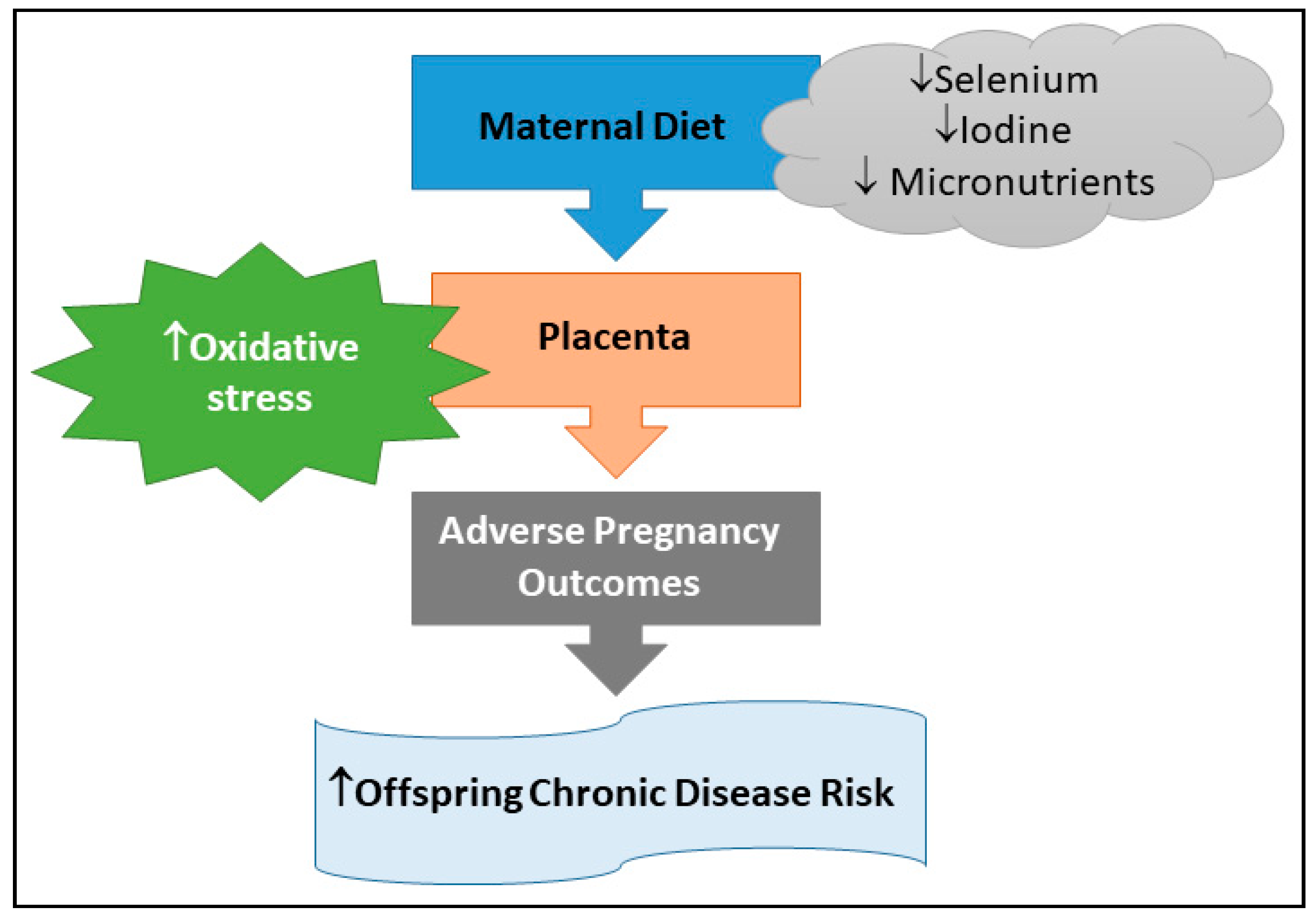
4. Impact of Maternal Selenium and Iodine, via Oxidative Stress, on Child Health
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Denison, F.C.; Roberts, K.A.; Barr, S.M.; Norman, J.E. Obesity, pregnancy, inflammation, and vascular function. Reproduction 2010, 140, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Williams, D. Pregnancy: A stress test for life. Curr. Opin. Obstet. Gynecol. 2003, 15, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Lockitch, G. Clinical biochemistry of pregnancy. Crit. Rev. Clin. Lab. Sci. 1997, 34, 67–139. [Google Scholar] [CrossRef] [PubMed]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Myatt, L.; Cui, X. Oxidative stress in the placenta. Histochem. Cell Biol. 2004, 122, 369–382. [Google Scholar] [CrossRef]
- Ji, L.; Brkic, J.; Liu, M.; Fu, G.; Peng, C.; Wang, Y.L. Placental trophoblast cell differentiation: Physiological regulation and pathological relevance to preeclampsia. Mol. Asp. Med. 2013, 34, 981–1023. [Google Scholar] [CrossRef]
- Wong, F.; Cox, B.J. Cellular analysis of trophoblast and placenta. Placenta 2017, 59 (Suppl. 1), S2–S7. [Google Scholar] [CrossRef]
- Raghavan, R.; Dreibelbis, C.; Kingshipp, B.L.; Wong, Y.P.; Abrams, B.; Gernand, A.D.; Rasmussen, K.M.; Siega-Riz, A.M.; Stang, J.; Casavale, K.O.; et al. Dietary patterns before and during pregnancy and maternal outcomes: A systematic review. Am. J. Clin. Nutr. 2019, 109, 705S–728S. [Google Scholar] [CrossRef]
- Chen, Q.; Feng, Y.; Yang, H.; Wu, W.; Zhang, P.; Wang, K.; Wang, Y.; Ko, J.; Shen, J.; Guo, L.; et al. A Vitamin Pattern Diet Is Associated with Decreased Risk of Gestational Diabetes Mellitus in Chinese Women: Results from a Case Control Study in Taiyuan, China. J. Diabetes Res. 2019, 2019, 5232308. [Google Scholar] [CrossRef]
- Grieger, J.A.; Clifton, V.L. A review of the impact of dietary intakes in human pregnancy on infant birthweight. Nutrients 2014, 7, 153–178. [Google Scholar] [CrossRef]
- Ramakrishnan, U.; Grant, F.; Goldenberg, T.; Zongrone, A.; Martorell, R. Effect of women’s nutrition before and during early pregnancy on maternal and infant outcomes: A systematic review. Paediatr. Perinat. Epidemiol. 2012, 26 (Suppl. 1), 285–301. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, J.; Heslehurst, N.; Hall, J.; Schoenaker, D.; Hutchinson, J.; Cade, J.E.; Poston, L.; Barrett, G.; Crozier, S.R.; Barker, M.; et al. Before the beginning: Nutrition and lifestyle in the preconception period and its importance for future health. Lancet 2018, 391, 1830–1841. [Google Scholar] [CrossRef]
- Grieger, J.A.; Grzeskowiak, L.E.; Clifton, V.L. Preconception dietary patterns in human pregnancies are associated with preterm delivery. J. Nutr. 2014, 144, 1075–1080. [Google Scholar] [CrossRef] [PubMed]
- Emond, J.A.; Karagas, M.R.; Baker, E.R.; Gilbert-Diamond, D. Better Diet Quality during Pregnancy Is Associated with a Reduced Likelihood of an Infant Born Small for Gestational Age: An Analysis of the Prospective New Hampshire Birth Cohort Study. J. Nutr. 2018, 148, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, U.; Grant, F.K.; Goldenberg, T.; Bui, V.; Imdad, A.; Bhutta, Z.A. Effect of multiple micronutrient supplementation on pregnancy and infant outcomes: A systematic review. Paediatr. Perinat. Epidemiol. 2012, 26 (Suppl. 1), 153–167. [Google Scholar] [CrossRef]
- Glinoer, D. The importance of iodine nutrition during pregnancy. Public Health Nutr. 2007, 10, 1542–1546. [Google Scholar] [CrossRef]
- Bizerea, T.O.; Dezsi, S.G.; Marginean, O.; Stroescu, R.; Rogobete, A.; Bizerea-Spiridon, O.; Ilie, C. The Link between Selenium, Oxidative Stress and Pregnancy Induced Hypertensive Disorders. Clin. Lab. 2018, 64, 1593–1610. [Google Scholar] [CrossRef]
- Omeljaniuk, W.J.; Socha, K.; Borawska, M.H.; Charkiewicz, A.E.; Laudanski, T.; Kulikowski, M.; Kobylec, E. Antioxidant status in women who have had a miscarriage. Adv. Med. Sci 2015, 60, 329–334. [Google Scholar] [CrossRef]
- Rayman, M.P.; Wijnen, H.; Vader, H.; Kooistra, L.; Pop, V. Maternal selenium status during early gestation and risk for preterm birth. CMAJ 2011, 183, 549–555. [Google Scholar] [CrossRef]
- Molnar, J.; Garamvolgyi, Z.; Herold, M.; Adanyi, N.; Somogyi, A.; Rigo, J., Jr. Serum selenium concentrations correlate significantly with inflammatory biomarker high-sensitive CRP levels in Hungarian gestational diabetic and healthy pregnant women at mid-pregnancy. Biol. Trace Elem. Res. 2008, 121, 16–22. [Google Scholar] [CrossRef]
- Kilinc, M.; Guven, M.A.; Ezer, M.; Ertas, I.E.; Coskun, A. Evaluation of serum selenium levels in Turkish women with gestational diabetes mellitus, glucose intolerants, and normal controls. Biol. Trace Elem. Res. 2008, 123, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Biagi, C.; Nunzio, M.D.; Bordoni, A.; Gori, D.; Lanari, M. Effect of Adherence to Mediterranean Diet during Pregnancy on Children’s Health: A Systematic Review. Nutrients 2019, 11, 997. [Google Scholar] [CrossRef] [PubMed]
- Schoots, M.H.; Gordijn, S.J.; Scherjon, S.A.; van Goor, H.; Hillebrands, J.L. Oxidative stress in placental pathology. Placenta 2018, 69, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Habibi, N.; Jankovic-Karasoulos, T.; Leemaqz, S.Y.; Francois, M.; Zhou, S.J.; Leifert, W.R.; Perkins, A.V.; Roberts, C.T.; Bianco-Miotto, T. Effect of Iodine and Selenium on Proliferation, Viability, and Oxidative Stress in HTR-8/SVneo Placental Cells. Biol. Trace Elem. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Smyth, P.P. Role of iodine in antioxidant defence in thyroid and breast disease. Biofactors 2003, 19, 121–130. [Google Scholar] [CrossRef]
- Shackelford, R.E.; Kaufmann, W.K.; Paules, R.S. Oxidative stress and cell cycle checkpoint function. Free Radic. Biol. Med. 2000, 28, 1387–1404. [Google Scholar] [CrossRef]
- Droge, W. Free radicals in the physiological control of cell function. Physiol Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharm. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Fehér, J.; Csomós, G.; Vereckei, A. The Chemistry of Free Radical Reactions. In Free Radical Reactions in Medicine; Springer: Berlin/Heidelberg, Germany, 1987; pp. 2–10. [Google Scholar] [CrossRef]
- Zaragoza, A.; Diez-Fernandez, C.; Alvarez, A.M.; Andres, D.; Cascales, M. Effect of N-acetylcysteine and deferoxamine on endogenous antioxidant defense system gene expression in a rat hepatocyte model of cocaine cytotoxicity. Biochim. Biophys. Acta 2000, 1496, 183–195. [Google Scholar] [CrossRef]
- Augustin, W.; Wiswedel, I.; Noack, H.; Reinheckel, T.; Reichelt, O. Role of endogenous and exogenous antioxidants in the defence against functional damage and lipid peroxidation in rat liver mitochondria. Mol. Cell Biochem. 1997, 174, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Jeeva, J.S.; Sunitha, J.; Ananthalakshmi, R.; Rajkumari, S.; Ramesh, M.; Krishnan, R. Enzymatic antioxidants and its role in oral diseases. J. Pharm. Bioallied Sci. 2015, 7, S331–S333. [Google Scholar] [CrossRef]
- Mistry, H.D.; Williams, P.J. The importance of antioxidant micronutrients in pregnancy. Oxidative Med. Cell Longev. 2011, 2011, 841749. [Google Scholar] [CrossRef] [PubMed]
- Paiva, S.A.; Russell, R.M. Beta-carotene and other carotenoids as antioxidants. J. Am. Coll. Nutr. 1999, 18, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Padayatty, S.J.; Katz, A.; Wang, Y.; Eck, P.; Kwon, O.; Lee, J.H.; Chen, S.; Corpe, C.; Dutta, A.; Dutta, S.K.; et al. Vitamin C as an antioxidant: Evaluation of its role in disease prevention. J. Am. Coll. Nutr. 2003, 22, 18–35. [Google Scholar] [CrossRef]
- Neiger, R. Long-Term Effects of Pregnancy Complications on Maternal Health: A Review. J. Clin. Med. 2017, 6, 76. [Google Scholar] [CrossRef]
- Leeson, P. Long term cardiovascular outcomes for mother and child. Pregnancy Hypertens. 2013, 3, 60–61. [Google Scholar] [CrossRef]
- Pisaneschi, S.; Boldrini, A.; Genazzani, A.R.; Coceani, F.; Simoncini, T. Feto-placental vascular dysfunction as a prenatal determinant of adult cardiovascular disease. Intern. Emerg. Med. 2013, 8 (Suppl. 1), S41–S45. [Google Scholar] [CrossRef]
- Tranquilli, A.L.; Landi, B.; Giannubilo, S.R.; Sibai, B.M. Preeclampsia: No longer solely a pregnancy disease. Pregnancy Hypertens. 2012, 2, 350–357. [Google Scholar] [CrossRef]
- Tranquilli, A.L.; Dekker, G.; Magee, L.; Roberts, J.; Sibai, B.M.; Steyn, W.; Zeeman, G.G.; Brown, M.A. The classification, diagnosis and management of the hypertensive disorders of pregnancy: A revised statement from the ISSHP. Pregnancy Hypertens. 2014, 4, 97–104. [Google Scholar] [CrossRef]
- Bellamy, L.; Casas, J.P.; Hingorani, A.D.; Williams, D.J. Pre-eclampsia and risk of cardiovascular disease and cancer in later life: Systematic review and meta-analysis. BMJ 2007, 335, 974. [Google Scholar] [CrossRef] [PubMed]
- Ayansina, D.; Black, C.; Hall, S.J.; Marks, A.; Millar, C.; Prescott, G.J.; Wilde, K.; Bhattacharya, S. Long term effects of gestational hypertension and pre-eclampsia on kidney function: Record linkage study. Pregnancy Hypertens. 2016, 6, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Odegard, R.A.; Vatten, L.J.; Nilsen, S.T.; Salvesen, K.A.; Austgulen, R. Preeclampsia and fetal growth. Obstet. Gynecol. 2000, 96, 950–955. [Google Scholar] [PubMed]
- van Esch, J.J.A.; van Heijst, A.F.; de Haan, A.F.J.; van der Heijden, O.W.H. Early-onset preeclampsia is associated with perinatal mortality and severe neonatal morbidity. J. Matern.-Fetal Neonatal Med. 2017, 30, 2789–2794. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.S.; Nohr, E.A.; Bech, B.H.; Vestergaard, M.; Catov, J.M.; Olsen, J. Health of children born to mothers who had preeclampsia: A population-based cohort study. Am. J. Obstet. Gynecol. 2009, 201, 269-e1. [Google Scholar] [CrossRef]
- Gastrich, M.D.; Zinonos, S.; Bachmann, G.; Cosgrove, N.M.; Cabrera, J.; Cheng, J.Q.; Kostis, J.B. Preeclamptic Women Are at Significantly Higher Risk of Future Cardiovascular Outcomes over a 15-Year Period. J. Women’s Health 2020, 29, 74–83. [Google Scholar] [CrossRef]
- Roberts, C.T. IFPA Award in Placentology Lecture: Complicated interactions between genes and the environment in placentation, pregnancy outcome and long term health. Placenta 2010, 31, S47–S53. [Google Scholar] [CrossRef]
- Steegers, E.A.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Pre-eclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef]
- Roberts, J.M.; Escudero, C. The placenta in preeclampsia. Pregnancy Hypertens. 2012, 2, 72–83. [Google Scholar] [CrossRef]
- Roberts, J.M.; Hubel, C.A. Oxidative stress in preeclampsia. Am. J. Obstet. Gynecol. 2004, 190, 1177–1178. [Google Scholar] [CrossRef]
- Salam, R.A.; Das, J.K.; Bhutta, Z.A. Impact of intrauterine growth restriction on long-term health. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Pathophysiology of placental-derived fetal growth restriction. Am. J. Obstet. Gynecol. 2018, 218, S745–S761. [Google Scholar] [CrossRef] [PubMed]
- Gernand, A.D.; Schulze, K.J.; Stewart, C.P.; West, K.P., Jr.; Christian, P. Micronutrient deficiencies in pregnancy worldwide: Health effects and prevention. Nat. Rev. Endocrinol 2016, 12, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Parisi, F.; Laoreti, A.; Cetin, I. Multiple micronutrient needs in pregnancy in industrialized countries. Ann. Nutr. Metab. 2014, 65, 13–21. [Google Scholar] [CrossRef]
- Berti, C.; Biesalski, H.K.; Gartner, R.; Lapillonne, A.; Pietrzik, K.; Poston, L.; Redman, C.; Koletzko, B.; Cetin, I. Micronutrients in pregnancy: Current knowledge and unresolved questions. Clin. Nutr. 2011, 30, 689–701. [Google Scholar] [CrossRef]
- Perkins, A.V.; Vanderlelie, J.J. Multiple micronutrient supplementation and birth outcomes: The potential importance of selenium. Placenta 2016, 48 (Suppl. 1), S61–S65. [Google Scholar] [CrossRef]
- Cogswell, M.E.; Parvanta, I.; Ickes, L.; Yip, R.; Brittenham, G.M. Iron supplementation during pregnancy, anemia, and birth weight: A randomized controlled trial. Am. J. Clin. Nutr. 2003, 78, 773–781. [Google Scholar] [CrossRef]
- Christian, P.; Khatry, S.K.; Katz, J.; Pradhan, E.K.; LeClerq, S.C.; Shrestha, S.R.; Adhikari, R.K.; Sommer, A.; West, K.P., Jr. Effects of alternative maternal micronutrient supplements on low birth weight in rural Nepal: Double blind randomised community trial. BMJ 2003, 326, 571. [Google Scholar] [CrossRef]
- Smits, L.J.; Essed, G.G. Short interpregnancy intervals and unfavourable pregnancy outcome: Role of folate depletion. Lancet 2001, 358, 2074–2077. [Google Scholar] [CrossRef]
- Siega-Riz, A.M.; Savitz, D.A.; Zeisel, S.H.; Thorp, J.M.; Herring, A. Second trimester folate status and preterm birth. Am. J. Obstet. Gynecol. 2004, 191, 1851–1857. [Google Scholar] [CrossRef]
- Watanabe, H.; Fukuoka, H.; Sugiyama, T.; Nagai, Y.; Ogasawara, K.; Yoshiike, N. Dietary folate intake during pregnancy and birth weight in Japan. Eur. J. Nutr. 2008, 47, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Lindblad, B.; Zaman, S.; Malik, A.; Martin, H.; Ekstrom, A.M.; Amu, S.; Holmgren, A.; Norman, M. Folate, vitamin B12, and homocysteine levels in South Asian women with growth-retarded fetuses. Acta Obstet. Gynecol. Scand. 2005, 84, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Charoenratana, C.; Leelapat, P.; Traisrisilp, K.; Tongsong, T. Maternal iodine insufficiency and adverse pregnancy outcomes. Matern. Child. Nutr. 2016, 12, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Gulaboglu, M.; Borekci, B.; Halici, Z. Placental tissue iodine level and blood magnesium concentration in pre-eclamptic and normal pregnancy. Int. J. Gynaecol. Obstet. 2007, 98, 100–104. [Google Scholar] [CrossRef]
- Cuellar-Rufino, S.; Navarro-Meza, M.; Garcia-Solis, P.; Xochihua-Rosas, I.; Arroyo-Helguera, O. Iodine levels are associated with oxidative stress and antioxidant status in pregnant women with hypertensive disease. Nutr. Hosp. 2017, 34, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Bo, S.; Lezo, A.; Menato, G.; Gallo, M.L.; Bardelli, C.; Signorile, A.; Berutti, C.; Massobrio, M.; Pagano, G.F. Gestational hyperglycemia, zinc, selenium, and antioxidant vitamins. Nutrition 2005, 21, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Kharb, S. Lipid peroxidation in pregnancy with preeclampsia and diabetes. Gynecol. Obstet. Investig. 2000, 50, 113–116. [Google Scholar] [CrossRef]
- Xiao, Y.; Sun, H.; Li, C.; Li, Y.; Peng, S.; Fan, C.; Teng, W.; Shan, Z. Effect of Iodine Nutrition on Pregnancy Outcomes in an Iodine-Sufficient Area in China. Biol. Trace Elem. Res. 2018, 182, 231–237. [Google Scholar] [CrossRef]
- Nafee, T.M.; Farrell, W.E.; Carroll, W.D.; Fryer, A.A.; Ismail, K.M. Epigenetic control of fetal gene expression. Br. J. Obstet. Gynaecol. 2008, 115, 158–168. [Google Scholar] [CrossRef]
- Waterland, R.A.; Jirtle, R.L. Early nutrition, epigenetic changes at transposons and imprinted genes, and enhanced susceptibility to adult chronic diseases. Nutrition 2004, 20, 63–68. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Shchedrina, V.A.; Zhang, Y.; Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Structure-function relations, physiological roles, and evolution of mammalian ER-resident selenoproteins. Antioxidants Redox Signal. 2010, 12, 839–849. [Google Scholar] [CrossRef]
- Grumolato, L.; Ghzili, H.; Montero-Hadjadje, M.; Gasman, S.; Lesage, J.; Tanguy, Y.; Galas, L.; Ait-Ali, D.; Leprince, J.; Guerineau, N.C.; et al. Selenoprotein T is a PACAP-regulated gene involved in intracellular Ca2+ mobilization and neuroendocrine secretion. FASEB J. 2008, 22, 1756–1768. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Speckmann, B.; Klotz, L.O. Selenoproteins: Antioxidant selenoenzymes and beyond. Arch. Biochem. Biophys. 2016, 595, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Arteel, G.E.; Mostert, V.; Oubrahim, H.; Briviba, K.; Abel, J.; Sies, H. Protection by selenoprotein P in human plasma against peroxynitrite-mediated oxidation and nitration. Biol. Chem. 1998, 379, 1201–1205. [Google Scholar] [PubMed]
- Saito, Y.; Hayashi, T.; Tanaka, A.; Watanabe, Y.; Suzuki, M.; Saito, E.; Takahashi, K. Selenoprotein P in human plasma as an extracellular phospholipid hydroperoxide glutathione peroxidase. Isolation and enzymatic characterization of human selenoprotein p. J. Biol. Chem. 1999, 274, 2866–2871. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. The Sep15 protein family: Roles in disulfide bond formation and quality control in the endoplasmic reticulum. IUBMB Life 2007, 59, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H. Selenium as an essential micronutrient: Roles in cell cycle and apoptosis. Molecules 2009, 14, 1263–1278. [Google Scholar] [CrossRef]
- Thomson, C.D. Assessment of requirements for selenium and adequacy of selenium status: A review. Eur. J. Clin. Nutr. 2004, 58, 391–402. [Google Scholar] [CrossRef]
- Kipp, A.P.; Strohm, D.; Brigelius-Flohe, R.; Schomburg, L.; Bechthold, A.; Leschik-Bonnet, E.; Heseker, H.; German Nutrition, S. Revised reference values for selenium intake. J. Trace Elem. Med. Biol 2015, 32, 195–199. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. Nutrient Reference Values for Australia and New Zealand. Available online: https://www.nrv.gov.au/nutrients (accessed on 14 April 2020).
- Food and Nutrition Board, Institute of Medicine. Dietary Reference Intakes for Vitamin C, Vitamin E, Selenium, and Carotenoids a Report of the Panel on Dietary Antioxidants and Related Compounds, Subcommittees on Upper Reference Levels of Nutrients and of Interpretation and Use of Dietary Reference Intakes, and the Standing Committee on the Scientific Evaluation of Dietary Reference Intakes; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; McGrath, S.P.; Seneviratne, S.I.; Smith, P.; Winkel, L.H. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2848–2853. [Google Scholar] [CrossRef] [PubMed]
- Shreenath, A.P.; Ameer, M.A.; Dooley, J. Selenium Deficiency. In Treasure Island (FL); StatPearls: Petersburg, FL, USA, 2020. [Google Scholar]
- Li, S.; Banuelos, G.S.; Wu, L.; Shi, W. The changing selenium nutritional status of Chinese residents. Nutrients 2014, 6, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Mariath, A.B.; Bergamaschi, D.P.; Rondo, P.H.; Tanaka, A.C.; Hinnig Pde, F.; Abbade, J.F.; Diniz, S.G. The possible role of selenium status in adverse pregnancy outcomes. Br. J. Nutr. 2011, 105, 1418–1428. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Hill, K.E.; Li, P.; Xu, J.; Zhou, D.; Motley, A.K.; Wang, L.; Byrne, D.W.; Burk, R.F. Optimization of selenoprotein P and other plasma selenium biomarkers for the assessment of the selenium nutritional requirement: A placebo-controlled, double-blind study of selenomethionine supplementation in selenium-deficient Chinese subjects. Am. J. Clin. Nutr. 2010, 92, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Ambroziak, U.; Hybsier, S.; Shahnazaryan, U.; Krasnodebska-Kiljanska, M.; Rijntjes, E.; Bartoszewicz, Z.; Bednarczuk, T.; Schomburg, L. Severe selenium deficits in pregnant women irrespective of autoimmune thyroid disease in an area with marginal selenium intake. J. Trace Elem. Med. Biol. 2017, 44, 186–191. [Google Scholar] [CrossRef]
- Irwinda, R.; Wibowo, N.; Putri, A.S. The Concentration of Micronutrients and Heavy Metals in Maternal Serum, Placenta, and Cord Blood: A Cross-Sectional Study in Preterm Birth. J. Pregnancy 2019, 2019, 5062365. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, E.; Derici, M.K.; Demir, E.; Apaydin, H.; Kocak, O.; Kan, O.; Gorkem, U. Is the Concentration of Cadmium, Lead, Mercury, and Selenium Related to Preterm Birth? Biol. Trace Elem. Res. 2019, 191, 306–312. [Google Scholar] [CrossRef]
- Wilson, R.L.; Bianco-Miotto, T.; Leemaqz, S.Y.; Grzeskowiak, L.E.; Dekker, G.A.; Roberts, C.T. Early pregnancy maternal trace mineral status and the association with adverse pregnancy outcome in a cohort of Australian women. J. Trace Elem. Med. Biol. 2018, 46, 103–109. [Google Scholar] [CrossRef]
- Hamdan, H.Z.; Elbashir, L.M.; Hamdan, S.Z.; Elhassan, E.M.; Adam, I. Zinc and selenium levels in women with gestational diabetes mellitus at Medani Hospital, Sudan. J. Obstet. Gynaecol. 2014, 34, 567–570. [Google Scholar] [CrossRef]
- Maleki, A.; Fard, M.K.; Zadeh, D.H.; Mamegani, M.A.; Abasaizadeh, S.; Mazloomzadeh, S. The relationship between plasma level of Se and preeclampsia. Hypertens. Pregnancy 2011, 30, 180–187. [Google Scholar] [CrossRef]
- Mistry, H.D.; Wilson, V.; Ramsay, M.M.; Symonds, M.E.; Broughton Pipkin, F. Reduced selenium concentrations and glutathione peroxidase activity in preeclamptic pregnancies. Hypertension 2008, 52, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Atamer, Y.; Kocyigit, Y.; Yokus, B.; Atamer, A.; Erden, A.C. Lipid peroxidation, antioxidant defense, status of trace metals and leptin levels in preeclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 2005, 119, 60–66. [Google Scholar] [CrossRef]
- Al-Saleh, E.; Nandakumaran, M.; Al-Shammari, M.; Al-Harouny, A. Maternal-fetal status of copper, iron, molybdenum, selenium and zinc in patients with gestational diabetes. J. Matern.-Fetal Neonatal Med. 2004, 16, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, W.C.; Alkan, Z.; Lang, K.; King, J.C. Plasma selenium decrease during pregnancy is associated with glucose intolerance. Biol. Trace Elem. Res. 2004, 100, 19–29. [Google Scholar] [CrossRef]
- Zachara, B.A.; Dobrzynski, W.; Trafikowska, U.; Szymanski, W. Blood selenium and glutathione peroxidases in miscarriage. Br. J. Obstet. Gynaecol. 2001, 108, 244–247. [Google Scholar] [CrossRef]
- Ceko, M.J.; O’Leary, S.; Harris, H.H.; Hummitzsch, K.; Rodgers, R.J. Trace Elements in Ovaries: Measurement and Physiology. Biol. Reprod. 2016, 94, 86. [Google Scholar] [CrossRef]
- Grieger, J.A.; Grzeskowiak, L.E.; Wilson, R.L.; Bianco-Miotto, T.; Leemaqz, S.Y.; Jankovic-Karasoulos, T.; Perkins, A.V.; Norman, R.J.; Dekker, G.A.; Roberts, C.T. Maternal Selenium, Copper and Zinc Concentrations in Early Pregnancy, and the Association with Fertility. Nutrients 2019, 11. [Google Scholar] [CrossRef]
- Rayman, M.P.; Bode, P.; Redman, C.W. Low selenium status is associated with the occurrence of the pregnancy disease preeclampsia in women from the United Kingdom. Am. J. Obstet. Gynecol. 2003, 189, 1343–1349. [Google Scholar] [CrossRef]
- Tan, M.; Sheng, L.; Qian, Y.; Ge, Y.; Wang, Y.; Zhang, H.; Jiang, M.; Zhang, G. Changes of serum selenium in pregnant women with gestational diabetes mellitus. Biol. Trace Elem. Res. 2001, 83, 231–237. [Google Scholar] [CrossRef]
- Rayman, M.P.; Bath, S.C.; Westaway, J.; Williams, P.; Mao, J.; Vanderlelie, J.J.; Perkins, A.V.; Redman, C.W. Selenium status in U.K. pregnant women and its relationship with hypertensive conditions of pregnancy. Br. J. Nutr. 2015, 113, 249–258. [Google Scholar] [CrossRef]
- Tara, F.; Rayman, M.P.; Boskabadi, H.; Ghayour-Mobarhan, M.; Sahebkar, A.; Yazarlu, O.; Ouladan, S.; Tavallaie, S.; Azimi-Nezhad, M.; Shakeri, M.T.; et al. Selenium supplementation and premature (pre-labour) rupture of membranes: A randomised double-blind placebo-controlled trial. J. Obstet. Gynaecol. 2010, 30, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Searle, E.; Kelly, L.; Johnsen, S.; Bodman-Smith, K.; Bath, S.C.; Mao, J.; Redman, C.W. Effect of selenium on markers of risk of pre-eclampsia in UK pregnant women: A randomised, controlled pilot trial. Br. J. Nutr. 2014, 112, 99–111. [Google Scholar] [CrossRef]
- Dobrzynski, W.; Trafikowska, U.; Trafikowska, A.; Pilecki, A.; Szymanski, W.; Zachara, B.A. Decreased selenium concentration in maternal and cord blood in preterm compared with term delivery. Analyst 1998, 123, 93–97. [Google Scholar] [CrossRef]
- Orhan, H.; Onderoglu, L.; Yucel, A.; Sahin, G. Circulating biomarkers of oxidative stress in complicated pregnancies. Arch. Gynecol. Obstet. 2003, 267, 189–195. [Google Scholar] [CrossRef]
- Morreale de Escobar, G.; Obregon, M.J.; Escobar del Rey, F. Role of thyroid hormone during early brain development. Eur J. Endocrinol 2004, 151 (Suppl. 3), U25–U37. [Google Scholar] [CrossRef]
- Bernal, J. Thyroid hormones and brain development. Vitam. Horm. 2005, 71, 95–122. [Google Scholar] [CrossRef]
- Abel, M.H.; Brandlistuen, R.E.; Caspersen, I.H.; Aase, H.; Torheim, L.E.; Meltzer, H.M.; Brantsaeter, A.L. Language delay and poorer school performance in children of mothers with inadequate iodine intake in pregnancy: Results from follow-up at 8 years in the Norwegian Mother and Child Cohort Study. Eur. J. Nutr. 2019, 58, 3047–3058. [Google Scholar] [CrossRef]
- Abel, M.H.; Caspersen, I.H.; Meltzer, H.M.; Haugen, M.; Brandlistuen, R.E.; Aase, H.; Alexander, J.; Torheim, L.E.; Brantsaeter, A.L. Suboptimal Maternal Iodine Intake Is Associated with Impaired Child Neurodevelopment at 3 Years of Age in the Norwegian Mother and Child Cohort Study. J. Nutr. 2017, 147, 1314–1324. [Google Scholar] [CrossRef]
- World Health Organization. Assessment of Iodine Deficiency Disorders and Monitoring Their Elimination: A Guide for Programme Managers; World Health Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Azizi, F.; Sheikholeslam, R.; Hedayati, M.; Mirmiran, P.; Malekafzali, H.; Kimiagar, M.; Pajouhi, M. Sustainable control of iodinedeficiency in Iran: Beneficial results of the implementation of the mandatory law on salt iodization. J. Endocrinol Investig. 2002, 25, 409–413. [Google Scholar] [CrossRef]
- Charlton, K.; Probst, Y.; Kiene, G. Dietary Iodine Intake of the Australian Population after Introduction of a Mandatory Iodine Fortification Programme. Nutrients 2016, 8, 701. [Google Scholar] [CrossRef]
- Pandav, C.S.; Yadav, K.; Srivastava, R.; Pandav, R.; Karmarkar, M.G. Iodine deficiency disorders (IDD) control in India. Indian J. Med. Res. 2013, 138, 418–433. [Google Scholar] [PubMed]
- Pearce, E.N.; Andersson, M.; Zimmermann, M.B. Global iodine nutrition: Where do we stand in 2013? Thyroid 2013, 23, 523–528. [Google Scholar] [CrossRef]
- Australian Institute of Health and Welfare. Monitoring the Health Impacts of Mandatory Folic Acid and Iodine Fortification. Available online: www.aihw.gov.au (accessed on 14 April 2020).
- Iodine Global Network. Global Scorecard of Iodine Nutrition in 2016. Available online: https://www.ign.org/cm_data/Scorecard_2016_SAC_PW.pdf (accessed on 20 April 2020).
- Stagnaro-Green, A.; Pearce, E.N. Iodine and pregnancy: A call to action. Lancet 2013, 382, 292–293. [Google Scholar] [CrossRef]
- World Health Organization. Iodine Supplementation in Pregnant and Lactating Women. Available online: https://www.who.int/elena/titles/guidance_summaries/iodine_pregnancy/en/ (accessed on 20 April 2020).
- Burns, R.; Azizi, F.; Hedayati, M.; Mirmiran, P.; O’Herlihy, C.; Smyth, P.P. Is placental iodine content related to dietary iodine intake? Clin. Endocrinol. 2011, 75, 261–264. [Google Scholar] [CrossRef]
- Burns, R.; O’Herlihy, C.; Smyth, P.P. The placenta as a compensatory iodine storage organ. Thyroid 2011, 21, 541–546. [Google Scholar] [CrossRef]
- Schomburg, L.; Kohrle, J. On the importance of selenium and iodine metabolism for thyroid hormone biosynthesis and human health. Mol. Nutr. Food Res. 2008, 52, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.B.; Boelaert, K. Iodine deficiency and thyroid disorders. Lancet Diabetes Endocrinol. 2015, 3, 286–295. [Google Scholar] [CrossRef]
- Song, Y.; Driessens, N.; Costa, M.; De Deken, X.; Detours, V.; Corvilain, B.; Maenhaut, C.; Miot, F.; Van Sande, J.; Many, M.C.; et al. Roles of hydrogen peroxide in thyroid physiology and disease. J. Clin. Endocrinol. Metab. 2007, 92, 3764–3773. [Google Scholar] [CrossRef] [PubMed]
- Venturi, S. Is there a role for iodine in breast diseases? Breast 2001, 10, 379–382. [Google Scholar] [CrossRef]
- Barman, M.; Brantsaeter, A.L.; Nilsson, S.; Haugen, M.; Lundh, T.; Combs, G.F.; Zhang, G.; Muglia, L.J.; Meltzer, H.M.; Jacobsson, B.; et al. Maternal dietary selenium intake is associated with increased gestational length and decreased risk of preterm delivery. Br. J. Nutr. 2020, 123, 209–219. [Google Scholar] [CrossRef]
- Mesdaghinia, E.; Rahavi, A.; Bahmani, F.; Sharifi, N.; Asemi, Z. Clinical and Metabolic Response to Selenium Supplementation in Pregnant Women at Risk for Intrauterine Growth Restriction: Randomized, Double-Blind, Placebo-Controlled Trial. Biol. Trace Elem. Res. 2017, 178, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Maduray, K.; Moodley, J.; Soobramoney, C.; Moodley, R.; Naicker, T. Elemental analysis of serum and hair from pre-eclamptic South African women. J. Trace Elem. Med. Biol. 2017, 43, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.M.; Moghal, M.M.; Sarwar, M.S.; Anonna, S.N.; Akter, M.; Karmakar, P.; Ahmed, S.; Sattar, M.A.; Islam, M.S. Low serum selenium concentration is associated with preeclampsia in pregnant women from Bangladesh. J. Trace Elem. Med. Biol. 2016, 33, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Asemi, Z.; Jamilian, M.; Mesdaghinia, E.; Esmaillzadeh, A. Effects of selenium supplementation on glucose homeostasis, inflammation, and oxidative stress in gestational diabetes: Randomized, double-blind, placebo-controlled trial. Nutrition 2015, 31, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Mistry, H.D.; Gill, C.A.; Kurlak, L.O.; Seed, P.T.; Hesketh, J.E.; Meplan, C.; Schomburg, L.; Chappell, L.C.; Morgan, L.; Poston, L.; et al. Association between maternal micronutrient status, oxidative stress, and common genetic variants in antioxidant enzymes at 15 weeks gestation in nulliparous women who subsequently develop preeclampsia. Free Radic. Biol. Med. 2015, 78, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Ghaemi, S.Z.; Forouhari, S.; Dabbaghmanesh, M.H.; Sayadi, M.; Bakhshayeshkaram, M.; Vaziri, F.; Tavana, Z. A prospective study of selenium concentration and risk of preeclampsia in pregnant Iranian women: A nested case-control study. Biol. Trace Elem. Res. 2013, 152, 174–179. [Google Scholar] [CrossRef]
- Kurlak, L.O.; Mistry, H.D.; Kaptein, E.; Visser, T.J.; Broughton Pipkin, F. Thyroid hormones and their placental deiodination in normal and pre-eclamptic pregnancy. Placenta 2013, 34, 395–400. [Google Scholar] [CrossRef]
- Abdulah, R.; Noerjasin, H.; Septiani, L.; Defi, I.R.; Suradji, E.W.; Puspitasari, I.M.; Barliana, M.I.; Yamazaki, C.; Nakazawa, M.; Koyama, H. Reduced serum selenium concentration in miscarriage incidence of Indonesian subjects. Biol. Trace Elem. Res. 2013, 154, 1–6. [Google Scholar] [CrossRef]
- Al-Saleh, E.; Nandakumaran, M.; Al-Rashdan, I.; Al-Harmi, J.; Al-Shammari, M. Maternal-foetal status of copper, iron, molybdenum, selenium and zinc in obese gestational diabetic pregnancies. Acta Diabetol. 2007, 44, 106–113. [Google Scholar] [CrossRef]
- Vidal, Z.E.; Rufino, S.C.; Tlaxcalteco, E.H.; Trejo, C.H.; Campos, R.M.; Meza, M.N.; Rodriguez, R.C.; Arroyo-Helguera, O. Oxidative stress increased in pregnant women with iodine deficiency. Biol. Trace Elem. Res. 2014, 157, 211–217. [Google Scholar] [CrossRef]
- Olivo-Vidal, Z.E.; Rodriguez, R.C.; Arroyo-Helguera, O. Iodine Affects Differentiation and Migration Process in Trophoblastic Cells. Biol. Trace Elem. Res. 2016, 169, 180–188. [Google Scholar] [CrossRef]
- Elstner, E.F.; Adamczyk, R.; Kroner, R.; Furch, A. The uptake of potassium iodide and its effect as an antioxidant in isolated rabbit eyes. Ophthalmologica 1985, 191, 122–126. [Google Scholar] [CrossRef]
- Rieger, G.; Winkler, R.; Buchberger, W.; Moser, M. Iodine distribution in a porcine eye model following iontophoresis. Ophthalmologica 1995, 209, 84–87. [Google Scholar] [CrossRef]
- Venturi, S.; Donati, F.M.; Venturi, A.; Venturi, M.; Grossi, L.; Guidi, A. Role of iodine in evolution and carcinogenesis of thyroid, breast and stomach. Adv. Clin. Pathol. 2000, 4, 11–17. [Google Scholar]
- Maier, J.; van Steeg, H.; van Oostrom, C.; Paschke, R.; Weiss, R.E.; Krohn, K. Iodine deficiency activates antioxidant genes and causes DNA damage in the thyroid gland of rats and mice. Biochim. Biophys. Acta 2007, 1773, 990–999. [Google Scholar] [CrossRef]
- Borekci, B.; Gulaboglu, M.; Gul, M. Iodine and magnesium levels in maternal and umbilical cord blood of preeclamptic and normal pregnant women. Biol. Trace Elem. Res. 2009, 129, 1–8. [Google Scholar] [CrossRef]
- Dillon, J.C.; Milliez, J. Reproductive failure in women living in iodine deficient areas of West Africa. Br. J. Obstet. Gynaecol. 2000, 107, 631–636. [Google Scholar] [CrossRef]
- Vejbjerg, P.; Knudsen, N.; Perrild, H.; Laurberg, P.; Andersen, S.; Rasmussen, L.B.; Ovesen, L.; Jorgensen, T. Estimation of iodine intake from various urinary iodine measurements in population studies. Thyroid 2009, 19, 1281–1286. [Google Scholar] [CrossRef]
- Eastman, C.J.; Ma, G.; Li, M. Optimal Assessment and Quantification of Iodine Nutrition in Pregnancy and Lactation: Laboratory and Clinical Methods, Controversies and Future Directions. Nutrients 2019, 11, 2378. [Google Scholar] [CrossRef]
- Kohrle, J. The deiodinase family: Selenoenzymes regulating thyroid hormone availability and action. Cell Mol. Life Sci. 2000, 57, 1853–1863. [Google Scholar] [CrossRef]
- Glinoer, D. The regulation of thyroid function in pregnancy: Pathways of endocrine adaptation from physiology to pathology. Endocr. Rev. 1997, 18, 404–433. [Google Scholar] [CrossRef]
- Sharlin, D.S. Chapter 8—Thyroid-Disrupting Chemicals as Developmental Neurotoxicants. In Environmental Factors in Neurodevelopmental and Neurodegenerative Disorders; Aschner, M., Costa, L.G., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 167–192. [Google Scholar] [CrossRef]
- Huang, S.A.; Dorfman, D.M.; Genest, D.R.; Salvatore, D.; Larsen, P.R. Type 3 iodothyronine deiodinase is highly expressed in the human uteroplacental unit and in fetal epithelium. J. Clin. Endocrinol. Metab. 2003, 88, 1384–1388. [Google Scholar] [CrossRef]
- Beckett, G.J.; Arthur, J.R. Selenium and endocrine systems. J. Endocrinol. 2005, 184, 455–465. [Google Scholar] [CrossRef]
- Torlinska, B.; Bath, S.C.; Janjua, A.; Boelaert, K.; Chan, S.Y. Iodine Status during Pregnancy in a Region of Mild-to-Moderate Iodine Deficiency is not Associated with Adverse Obstetric Outcomes; Results from the Avon Longitudinal Study of Parents and Children (ALSPAC). Nutrients 2018, 10, 291. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Y.; Liu, H.; Zheng, H.; Li, X.; Zhu, L.; Wang, Z. Associations of maternal iodine status and thyroid function with adverse pregnancy outcomes in Henan Province of China. J. Trace Elem. Med. Biol. 2018, 47, 104–110. [Google Scholar] [CrossRef]
- Olivares, J.L.; Olivi, G.I.; Verdasco, C.; Ortiz, V.A.; Mayer, M.A.; Cresto, J.C. Low iodine intake during pregnancy: Relationship to placental development and head circumference in newborn. Endocrinol. Nutr. 2012, 59, 326–330. [Google Scholar] [CrossRef]
- Gulaboglu, M.; Borekci, B.; Delibas, I. Urine iodine levels in preeclamptic and normal pregnant women. Biol. Trace Elem. Res. 2010, 136, 249–257. [Google Scholar] [CrossRef]
- DeLong, G.R.; Leslie, P.W.; Wang, S.H.; Jiang, X.M.; Zhang, M.L.; Rakeman, M.; Jiang, J.Y.; Ma, T.; Cao, X.Y. Effect on infant mortality of iodination of irrigation water in a severely iodine-deficient area of China. Lancet 1997, 350, 771–773. [Google Scholar] [CrossRef]
- Barker, D.J.; Gluckman, P.D.; Godfrey, K.M.; Harding, J.E.; Owens, J.A.; Robinson, J.S. Fetal nutrition and cardiovascular disease in adult life. Lancet 1993, 341, 938–941. [Google Scholar] [CrossRef]
- Hofstee, P.; McKeating, D.R.; Perkins, A.V.; Cuffe, J.S. Placental adaptations to micronutrient dysregulation in the programming of chronic disease. Clin. Exp. Pharmacol. Physiol. 2018, 45, 871–884. [Google Scholar] [CrossRef]
- Hofstee, P.; Bartho, L.A.; McKeating, D.R.; Radenkovic, F.; McEnroe, G.; Fisher, J.J.; Holland, O.J.; Vanderlelie, J.J.; Perkins, A.V.; Cuffe, J.S.M. Maternal selenium deficiency during pregnancy in mice increases thyroid hormone concentrations, alters placental function and reduces fetal growth. J. Physiol. 2019, 597, 5597–5617. [Google Scholar] [CrossRef] [PubMed]
- Hofstee, P.; McKeating, D.R.; Bartho, L.A.; Anderson, S.T.; Perkins, A.V.; Cuffe, J.S.M. Maternal Selenium Deficiency in Mice Alters Offspring Glucose Metabolism and Thyroid Status in a Sexually Dimorphic Manner. Nutrients 2020, 12, 267. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. The Good, the Bad, and the Ugly of Pregnancy Nutrients and Developmental Programming of Adult Disease. Nutrients 2019, 11, 894. [Google Scholar] [CrossRef] [PubMed]
- Chavatte-Palmer, P.; Tarrade, A.; Rousseau-Ralliard, D. Diet before and during Pregnancy and Offspring Health: The Importance of Animal Models and What Can Be Learned from Them. Int. J. Environ. Res. Public Health 2016, 13, 586. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Cindrova-Davies, T.; Yung, H.W.; Jauniaux, E. Oxygen and development of the human placenta. Reproduction 2020. [Google Scholar] [CrossRef]
- Aljunaidy, M.M.; Morton, J.S.; Cooke, C.M.; Davidge, S.T. Prenatal hypoxia and placental oxidative stress: Linkages to developmental origins of cardiovascular disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 313, R395–R399. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.S.; Cooke, C.L.; Davidge, S.T. In Utero Origins of Hypertension: Mechanisms and Targets for Therapy. Physiol. Rev. 2016, 96, 549–603. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| References (Ref.) | Country Year | Sample Size (Women) | Time of Sampling | Sample Type | Selenium Concentration (µg/L) |
|---|---|---|---|---|---|
| [90] | Indonesia 2019 | 25 pregnant | Delivery | Serum | 76.42 ± 16.30 |
| [91] | Turkey 2019 | 30 pregnant | Delivery | Blood, urine, amniotic fluid | Median (min-max): |
| Maternal blood: 78.98 (72.36–84.14) | |||||
| Maternal urine: 23.44 (19.66–26.69) | |||||
| Amniotic fluid: 26.00 (22.56–29.88) | |||||
| [92] | Australia 2018 | 558 pregnant | 15 ± 1 weeks’ gestation | Plasma | Mean ± SD: 71.93 ± 11.05 |
| [93] | Sudan 2014 | 31 pregnant | Not reported | Serum | Median (25–75th quartile): 204 (68–541) |
| [94] | Iran 2011 | 40 pregnant | 34–39 weeks’ gestation | Blood | Mean ± SD: 58.51 ± 11.85 |
| [95] | UK 2008 | Delivery | Blood | Mean ± SD: | |
| 27 pregnant | Pregnant: 58.4 ± 14.9 | ||||
| 22 non-pregnant | Non-pregnant: 69.8 ± 11.7 | ||||
| [20] | Hungary 2008 | 24–28 weeks’ gestation | Serum | Mean ± SD: | |
| 20 pregnant | Pregnant: 40.5 ± 8.03 | ||||
| 24 non-pregnant | Non-pregnant: 77.4 ± 14.82 | ||||
| [96] | Turkey 2005 | 28–39 weeks’ gestation | Serum | Mean ± SD: | |
| 28 pregnant | Pregnant: 87.50 ± 10.96 | ||||
| 25 non-pregnant | Non-pregnant: 109.0 ± 6.34 | ||||
| [97] | Kuwait 2004 | 15 pregnant | Delivery | Blood | Mean ± SEM: |
| Maternal vein: 102.3 ± 3.1 | |||||
| Umbilical artery: 85.4 ± 4.2 | |||||
| Umbilical vein: 82.6 ± 4.1 | |||||
| [98] | USA 2004 | 22 pregnant | 12 and 34 weeks’ gestation | Plasma | Mean ± SD: |
| 12 weeks’ gestation: 126.0 ± 15.0 | |||||
| 34 weeks’ gestation: 111.0 ± 12.0 | |||||
| [99] | Poland 2001 | Before or within 12 hours after uterine curettage | Whole blood and plasma | Mean ± SD: Whole blood: | |
| 36 pregnant | Pregnant: 74.1 ± 11.6 | ||||
| 28 non-pregnant | Non-pregnant: 90.5 ± 11.2 | ||||
| Plasma: | |||||
| 36 pregnant | Pregnant: 54.6 ± 11.1 | ||||
| 28 non-pregnant | Non-pregnant: 66.1 ± 13.1 |
| Ref. | Country Year | Sample Size (Pregnant Women) | Time of Sampling | Sample Type and Assays | Outcomes |
|---|---|---|---|---|---|
| [128] | Norway 2020 | 2638 (2558 term, 80 PTB) | 17–18 weeks | Blood Se | No association between blood Se and PTB risk with adjustment for iodine intake (population had moderate iodine deficiency) |
| [90] | Indonesia 2019 | 51 (25 term, 26 PTB) | Delivery | Serum, placental and cord blood Se | Lower Se in the placenta and cord blood from PTB compared to term deliveries (p < 0.05) |
| [91] | Turkey 2019 | 50 (30 term, 20 PTB) | Delivery | Blood, urine and amniotic fluid Se | Lower blood, urine and amniotic fluid Se in PTB compared to term (p < 0.05) |
| [92] | Australia 2018 | 1065 (480 healthy, 585 complicated) | 15 ± 1 weeks | Plasma Se, C-reactive protein | No difference between complicated and healthy pregnancies. No association between Se and C-reactive protein. |
| [129] | Iran 2017 | 60 at risk for IUGR (30 Se-supplementation, 30 placebo); RCT | Week 17 and end of intervention (week 27) | Plasma total antioxidant capacity, glutathione Serum C-reactive protein | Higher total antioxidant capacity and glutathione and lower C-reactive protein after 100 μg/d Se supplementation |
| [130] | South Africa 2017 | 66 (23 healthy, 43 PE) | Delivery | Serum and hair Se | Lower Se concentration in PE, no difference in hair Se between healthy and PE |
| [131] | Bangladesh 2015 | 74 PE (52 mild, 22 severe 118 normotensive) | ≥20 weeks | Serum Se | Lower Se concentration in mild and severe PE, lower Se in severe PE compared to mild PE |
| [132] | Iran 2015 | 65 with GDM (32 Se-supplementation 33 placebo) | Week 24 and end of intervention (week 28) | Plasma MDA and glutathione | Higher glutathione and lower MDA after 200 μg/d Se- supplementation |
| [133] | Australia 2015 | 716 (472 healthy, 244 PE) | 15 ± 1 weeks | Plasma Se | No difference in plasma Se between healthy and PE |
| [104] | UK 2015 | 230 primiparous 60 μg/d selenium yeast or placebo | (1) 12 and 35 weeks | (1) Whole-blood Se | After Se supplementation, low toenail Se was associated with decreased OR for PE/PIH (OR 0.30, 95% CI 0.09, 1.00, p = 0.049) |
| (2) 16 weeks | (2) Toenail Se | ||||
| [93] | Sudan 2014 | 62 (31 healthy,31 GDM) | Not reported | Serum Se | No difference in Se level between healthy and GDM pregnancies |
| [106] | UK 2014 | 229 primiparous (115 Se-treated, 114 placebo) 60 μg/d selenium yeast | (1) 12 and 35 weeks | (1) Whole-blood Se | Higher Se and selenoprotein P in Se-treated group at 35 weeks. Reduced PE/PIH odds in all Se-treated participants (OR 0.350, 95% CI 0.126, 0.974; p = 0.044) |
| (2) 35 weeks | (2) selenoprotein P | ||||
| [134] | Iran 2013 | 76 (38 healthy, 38 PE) | 24 weeks–2 days after delivery | Plasma Se | Lower Se in women with PE compared to healthy pregnancies |
| [135] | UK 2013 | 50 (27 healthy, 23 PE) | Delivery | Serum Se, placental deiodinase mRNA and enzyme activity | Lower Se in women with PE, correlation between placental deiodinase III mRNA expression and its enzyme activity only in PE |
| [136] | Indonesia 2013 | 71 (46 healthy, 25 miscarriage) | 8–20 weeks | Serum Se, GPx | Lower Se in women with miscarriage, no difference in GPx activity |
| [94] | Iran 2011 | 80 (40 healthy, 40 PE) | 34–39 weeks | Plasma Se | Lower Se in women with PE |
| [19] | Netherlands 2011 | 1129 (60 PTB, 21 PROM, 13 PE) | 12 weeks | Serum Se | Higher risk for PTB with lower Se (OR 2.18, 95% CI 1.25–3.77) |
| [105] | Iran 2010 | 166 primigravid (83 Se supplement, 83 placebo) 60 μg/d selenium yeast | 1st trimester and delivery | Serum Se | Increased Se (p < 0.01) and lower incidence of PROM (p < 0.001) in 100 μg/d of selenium yeast-supplemented group |
| [95] | UK 2008 | 74 (27 healthy, 25 PE, 22 non-pregnant age-matched) | Delivery | Serum, plasma, umbilical venous Se. Plasma and umbilical venous TBARS. Plasma and placental GPx. | Decreasing trend of plasma Se from non-pregnant to normal pregnant and PE; Lower serum Se and plasma GPx in PE compared to healthy pregnancies; higher level of maternal and umbilical venous TBARS in PE group; Lower placental GPx in PE |
| [20] | Hungary 2008 | 61 (20 healthy, 17 GDM, 24 healthy non-pregnant) | 24–28 weeks | Serum Se, high sensitivity C-reactive protein | Negative correlation between serum Se and high-sensitive C-reactive protein |
| [21] | Turkey 2008 | 180 (101 healthy, 30 GDM, 49 glucose intolerance) | 24–28 weeks | Serum Se | Lower Se in GDM and glucose intolerance compared to healthy pregnancies |
| [137] | Kuwait 2007 | 20 obese (10 GDM, 10 control) | Delivery | Blood Se, GPx, SOD and total antioxidant enzyme activity | Higher SOD activity in maternal vein, umbilical artery and umbilical vein blood of control obese women |
| [96] | Turkey 2005 | 85 (32 PE, 28 healthy pregnant, 25 non-pregnant) | 28–39 weeks | Serum and placenta MDA, SOD and catalase in erythrocytes. Placental GSH and GPx. Serum Se | Lower serum Se, erythrocyte SOD activity, and placental GPx and higher erythrocyte catalase activity in PE; negative correlation between placental MDA level and serum Se in PE |
| [67] | Italy 2005 | 504 (210 gestational hyperglycemic, 294 normoglycemic) | 24–28 weeks | Serum Se Dietary intake of Se | Lower dietary intake of Se in gestational hyperglycemic women; lower serum Se in women with impaired glucose tolerance; negative association between Se (OR 0.92, 95% CI = 0.87 to 0.95, p < 0.0001) and gestational hyperglycemia |
| [97] | Kuwait 2004 | 30 (15 healthy, 15 GDM) | Delivery | Blood Se from maternal vein, umbilical artery and umbilical vein | Lower maternal vein Se in GDM compared to healthy pregnancies |
| [108] | Turkey2003 | 36 (16 healthy, 9 PIH and PE, 3 IDDP; 3 GDM, 3 OP, 2 PAP) | 3rd trimester and immediately after delivery | GST, GPx and catalase activity and TBARS in maternal erythrocyte, plasma, and umbilical cord blood | Higher erythrocyte GPx activity and increased plasma TBARS in PIH and IDDP; higher cord blood GST activity (2–3 fold) in PE and PIH and IDDP compared to maternal activity before delivery; lower cord blood GPx activity compared to before delivery in PE and PIH; lower cord blood GPx (in PE and PIH, IDDP) and catalase activity (in PE and PIH) compared to maternal values; higher plasma TBARS in PE and PIH and IDDP in the antenatal period; lower cord blood erythrocytic TBARS in PE and PIH compared to maternal value |
| [102] | UK 2003 | 106 (53 healthy, 53 PE) | Not reported | Toenail Se | Lower Se in PE; more severe PE (delivery <32 weeks’) with lower Se within the PE group; higher PE risk in the bottom tertile of Se (OR 4.4, 95% CI 1.6–14.9) |
| [103] | China 2001 | 251 (98 IGT, 46 GDM, 90 healthy, 17 healthy non-pregnant) | 20 and 42 weeks | Serum Se | Lower Se at 33–42 weeks’ than at 20–33 weeks’ in all pregnant women; lower Se in IGT and GDM; lower Se in healthy pregnancies compared to healthy non-pregnant |
| Ref. | Country Year | Sample Size (Pregnant Women) | Time of Sampling | Sample Type and Assays | Outcome |
|---|---|---|---|---|---|
| [153] | UK 2018 | 3182 (3140 with child alive at 1st year, 42 pregnancy/infant loss) | Not reported | Urinary iodine-to-creatinine ratio(spot urine) | No association between iodine status and pregnancy complications or infant loss |
| [69] | China 2018 | 1569 euthyroid and primapara | 1st trimester | Urinary iodine concentration (spot urine) | Mild iodine deficiency (urinary iodine 100–150 μg/L) was an independent risk factor for GDM (OR 1.669, 95% CI 1.114–2.501, p < 0.05). More than adequate and excessive iodine (urinary iodine ≥250 μg/L) was an independent risk factor for macrosomia (OR = 2.116, 95% CI 1.218–3.676, p < 0.05) |
| [154] | China 2018 | 2347 | 1st, 2nd, 3rd trimester | Urinary iodine concentration (spot urine) | Lower incidences of PE in pregnant women with UIC 150–249 μg/L compared to the reference group of UIC < 50 μg/L (OR = 0.12, 95% CI 0.01–0.87, p < 0.05) |
| [66] | Mexico 2017 | 57 (37 normotensive, 20 HPD) | 3rd trimester | Urinary iodine concentration (spot urine), SOD, CAT, TBARS | Significant association between iodine deficiency and hypertensive disease of pregnancy (HPD); lower level of urinary iodine, SOD and CAT and higher level of TBARS in HPD compared to normotensive |
| [64] | Thailand 2016 | 390 | Each trimester | Urinary iodine concentration (spot urine) | Higher PTB risk (OR 2.69, 95% CI 1.38–5.24, p = 0.004) and low birthweight (OR 2.66, 95% CI 1.40–5.05, p = 0.003) among urinary iodine < 150 μg/L |
| [155] | Argentina 2012 | 77 | Not reported | Urinary iodine concentration (morning and evening urine samples, placental weight, placental index | Higher risk of lower placental weight in iodine deficiency (urinary iodine < 150 μg/L) (OR 3, 95% CI 1.06–8.5) |
| [156] | Turkey 2010 | 58 (40 severe PE, 18 healthy) | Not reported | Urinary iodine concentration (spot urine), thyroid hormone levels (T3, T4, TSH, fT3, fT4), blood magnesium | Positive correlation between urinary iodine and blood magnesium level in PE; higher T3 and fT3 levels in PE |
| [144] | Turkey 2009 | 40 (24 severe PE, 16 healthy) | Not reported for blood | Serum protein-bound iodine | Lower level of serum protein-bound iodine in maternal blood in PE; higher serum protein-bound iodine level in umbilical cord blood of infants in severe PE |
| [65] | Turkey 2007 | 35 (20 severe PE, 15 healthy) | Not reported for blood | Placental tissue iodine content, blood magnesium | Lower placental iodine in PE; positive correlation between placental iodine and blood magnesium level in PE |
| [145] | Senegal 2000 | 882 (462 pregnant, 420 non-pregnant) | Not reported | Urinary iodine concentration (spot urine), rate of miscarriage and stillbirth | Higher risk of miscarriage and stillbirth in iodine deficiency; highest rate in severe iodine deficiency (urinary iodine ≤ 20 μg/L) (OR 3.64, 95% CI 2.92–4.55) |
| [157] | China 1997 | >60,000 Iodine supplementation to water | Not reported | Neonatal and infant mortality rate after iodine supplementation | Large reduction in both neonatal and infant mortality with iodine supplementation of water among all population in three severely iodine-deficient townships |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habibi, N.; Grieger, J.A.; Bianco-Miotto, T. A Review of the Potential Interaction of Selenium and Iodine on Placental and Child Health. Nutrients 2020, 12, 2678. https://doi.org/10.3390/nu12092678
Habibi N, Grieger JA, Bianco-Miotto T. A Review of the Potential Interaction of Selenium and Iodine on Placental and Child Health. Nutrients. 2020; 12(9):2678. https://doi.org/10.3390/nu12092678
Chicago/Turabian StyleHabibi, Nahal, Jessica A. Grieger, and Tina Bianco-Miotto. 2020. "A Review of the Potential Interaction of Selenium and Iodine on Placental and Child Health" Nutrients 12, no. 9: 2678. https://doi.org/10.3390/nu12092678
APA StyleHabibi, N., Grieger, J. A., & Bianco-Miotto, T. (2020). A Review of the Potential Interaction of Selenium and Iodine on Placental and Child Health. Nutrients, 12(9), 2678. https://doi.org/10.3390/nu12092678