Oral Supplementation of the Vitamin D Metabolite 25(OH)D3 Against Influenza Virus Infection in Mice

 and
and 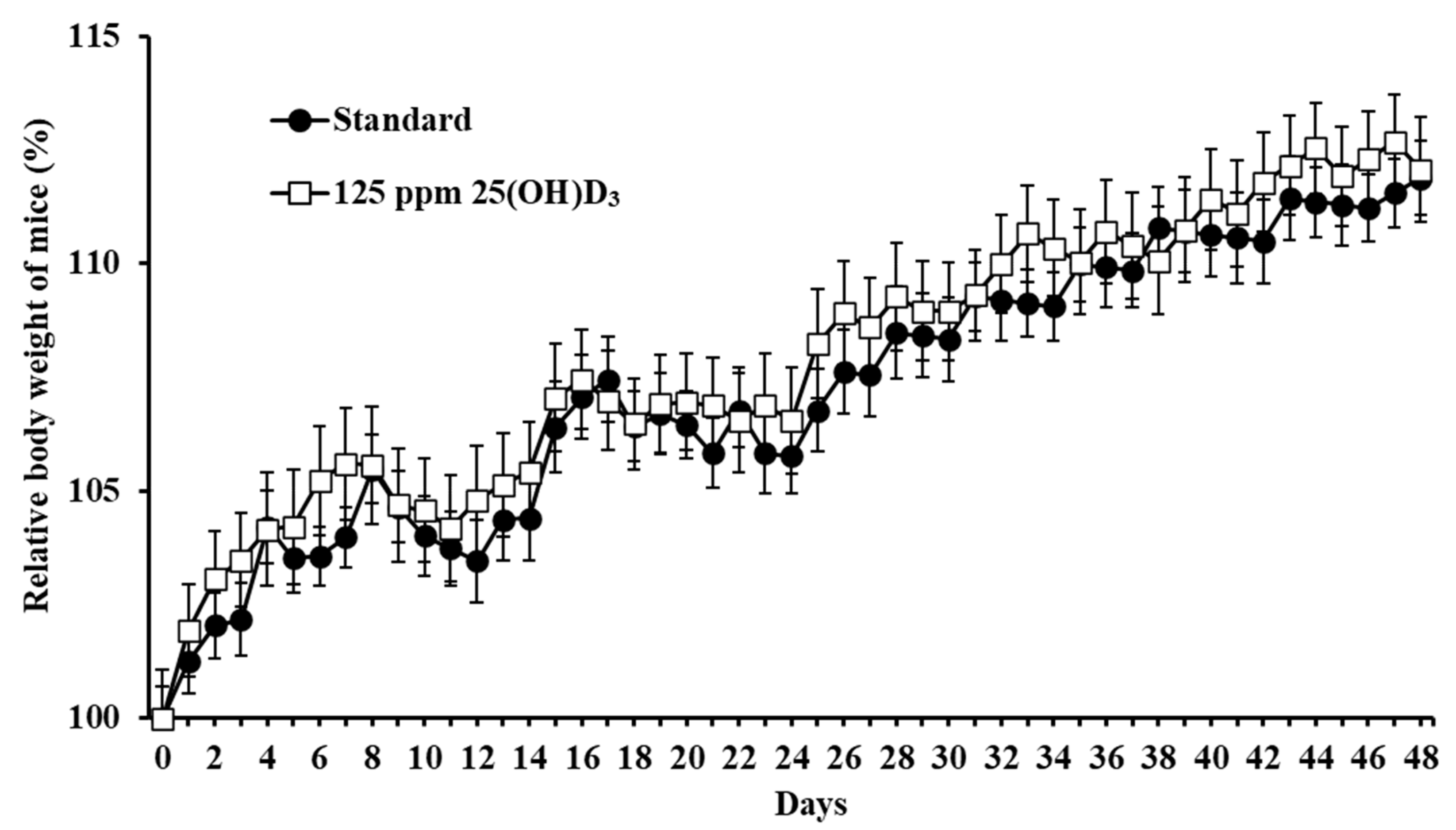{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus and Cells
2.2. Animals and Diet
2.3. Measurement of 25(OH)D3 and 24,25(OH)2D3
2.4. Virus Challenge in Mice
2.5. Preparation of Lung Homogenates
2.6. Virus Titration in Mouse Lungs
2.7. Bio-Plex Assay for the Measurement of Anti-Inflammatory and Proinflammatory Cytokines
2.8. Statistical Analysis
2.9. Ethics Statement
3. Results
3.1. Monitoring of Adverse Effects in Mice
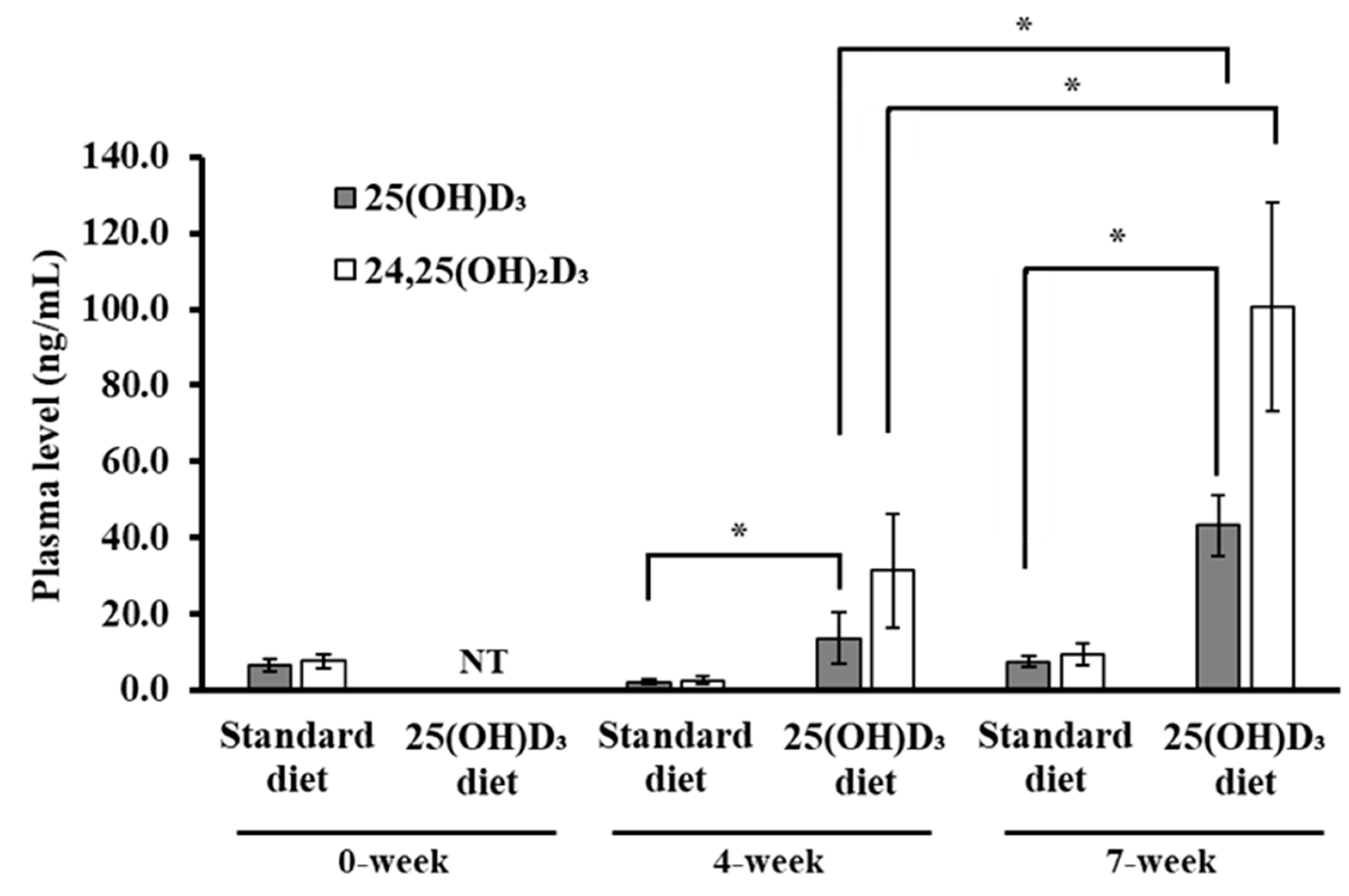
3.2. Blood Concentrations of 25(OH)D3 and 24,25(OH)2D3 in Mice
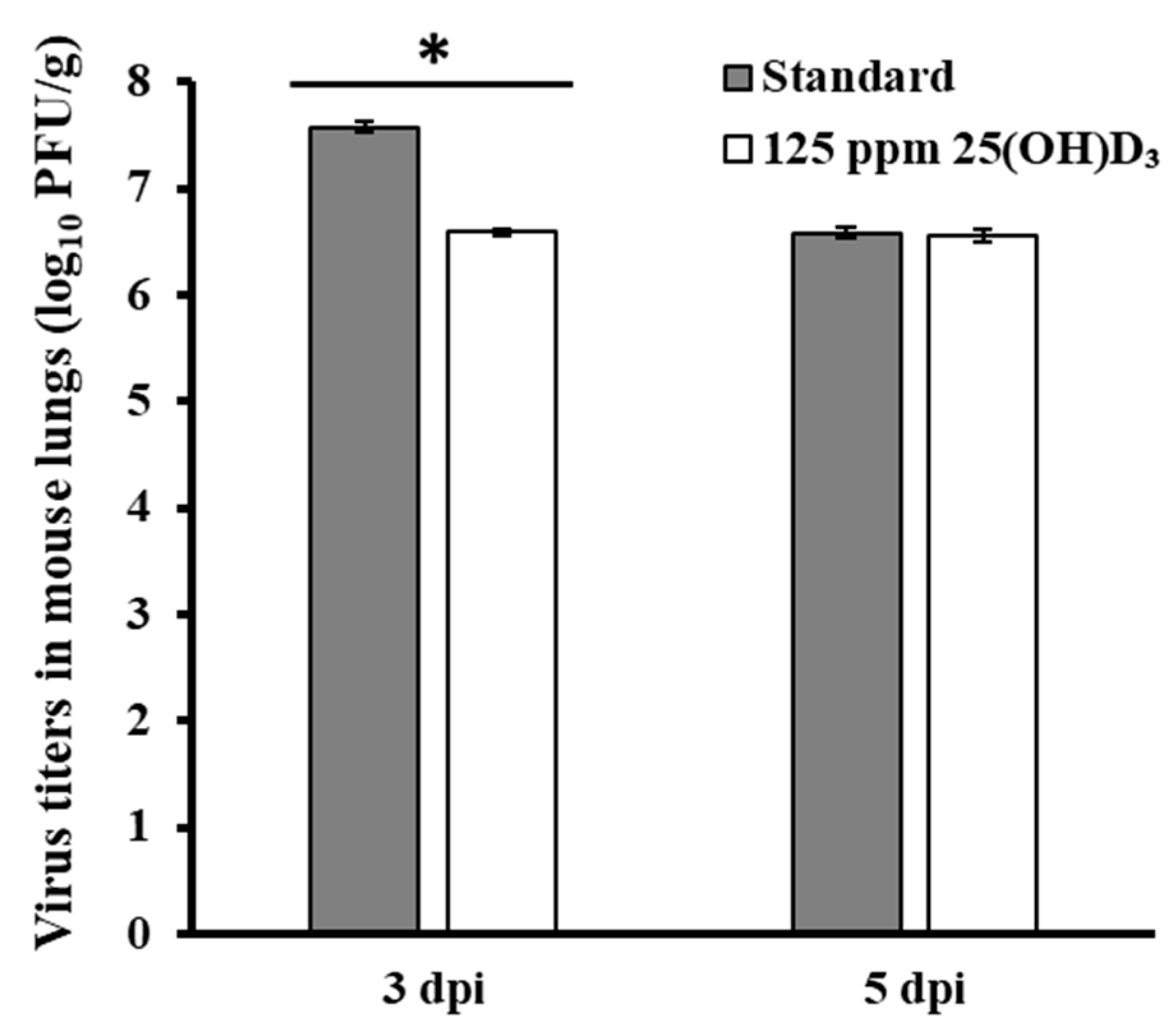
3.3. Immunostimulant Test for 25(OH)D3 Against Influenza Virus Infection
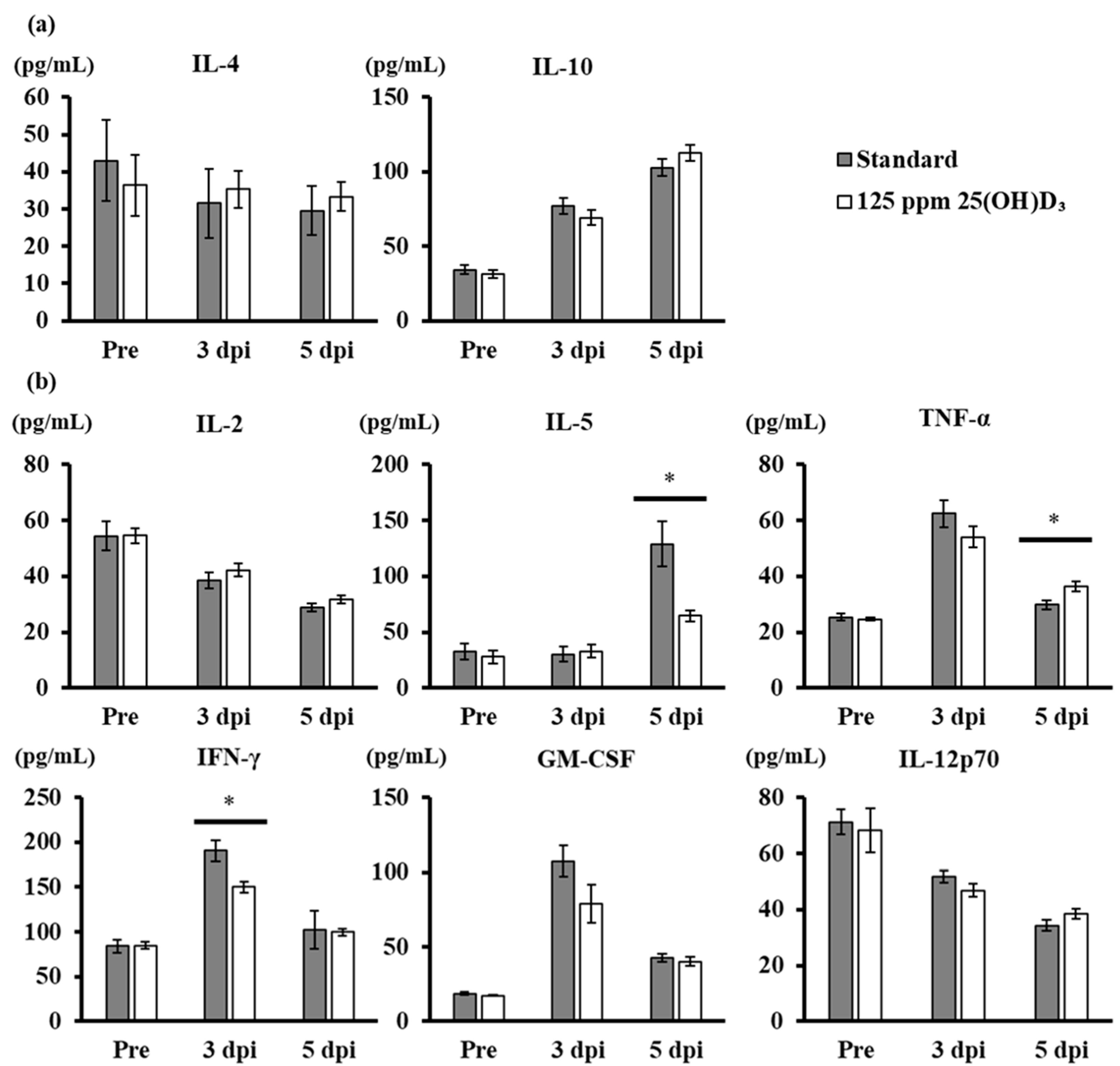
3.4. Production of Anti-Inflammatory and Proinflammatory Cytokines after Viral Challenge in Mice with 25(OH)D3 Supplementation
4. Discussions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lopez Payares, G.M.; Ali, F.A. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar]
- Lin, R. Crosstalk between Vitamin D metabolism, VDR signalling, and innate immunity. Biomed Res. Int. 2016, 2016, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Dong, H.; Zhao, H.; Song, J.; Tang, H.; Yao, L.; Liu, L.; Tong, W.; Zou, M.; Zou, F.; et al. 1,25-Dihydroxyvitamin D3 prevents toluene diisocyanate-induced airway epithelial barrier disruption. Int. J. Mol. Med. 2015, 36, 263–270. [Google Scholar] [CrossRef]
- Cynthia Aranow Vitamin D and the immune system. J. Investig. Med. 2011, 59, 881–886. [CrossRef] [PubMed]
- Cannell, J.J.; Zasloff, M.; Garland, C.F.; Scragg, R.; Giovannucci, E. On the epidemiology of influenza. Virol. J. 2008, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Maria, E. Sundaram and Laura A. Coleman Vitamin D and influenza. Adv. Nutr. 2012, 3, 517–525. [Google Scholar]
- Chang, J.H.; Cha, H.R.; Lee, D.S.; Seo, K.Y.; Kweon, M.N. 1,25-dihydroxyvitamin D3 inhibits the differentiation and migration of TH17 cells to protect against experimental autoimmune encephalomyelitis. PLoS ONE 2010, 5, 1–12. [Google Scholar] [CrossRef]
- Coussens, A.K.; Wilkinson, R.J.; Hanifa, Y.; Nikolayevskyy, V.; Elkington, P.T.; Islam, K.; Timms, P.M.; Venton, T.R.; Bothamley, G.H.; Packe, G.E.; et al. Vitamin D accelerates resolution of inflammatory responses during tuberculosis treatment. Proc. Natl. Acad. Sci. USA 2012, 109, 15449–15454. [Google Scholar] [CrossRef]
- Ivanov, A.P.; Dragunsky, E.M.; Chumakov, K.M. 1,25-Dihydroxyvitamin D3 Enhances Systemic and Mucosal Immune Responses to Inactivated Poliovirus Vaccine in Mice. J. Infect. Dis. 2006, 193, 598–600. [Google Scholar] [CrossRef]
- Medina, R.A.; García-Sastre, A. Influenza A viruses: New research developments. Nat. Rev. Microbiol. 2011, 9, 590–603. [Google Scholar] [CrossRef]
- Castetbon, K.; Vernay, M.; Malon, A.; Salanave, B.; Deschamps, V.; Roudier, C.; Oleko, A.; Szego, E.; Hercberg, S. Dietary intake, physical activity and nutritional status in adults: The French nutrition and health survey (ENNS, 2006-2007). Br. J. Nutr. 2009, 102, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Goncalves-Mendes, N.; Talvas, J.; Dualé, C.; Guttmann, A.; Corbin, V.; Marceau, G.; Sapin, V.; Brachet, P.; Evrard, B.; Laurichesse, H.; et al. Impact of Vitamin D supplementation on influenza vaccine response and immune functions in deficient elderly persons: A randomized placebo-controlled trial. Front. Immunol. 2019, 10, 65. [Google Scholar] [CrossRef]
- Biondi, P.; Pepe, J.; Biamonte, F.; Occhiuto, M.; Parisi, M.; Demofonti, C.; Baffa, V.; Minisola, S.; Cipriani, C. Oral calcidiol is a good form of vitamin D supplementation. Clin. Cases Miner. Bone Metab. 2017, 14, 207–208. [Google Scholar] [CrossRef] [PubMed]
- Shiva Borji, M.R.-K. Vitamin D and its importance on public health. J. Parathyr. Dis. 2015, 4, 20–24. [Google Scholar]
- Tsugawa, N.; Suhara, Y.; Kamao, M.; Okano, T. Determination of 25-Hydroxyvitamin D in Human Plasma Using High-Performance Liquid Chromatography - Tandem Mass Spectrometry. Anal. Chem. 2005, 77, 3001–3007. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.; Yasuda, K.; Takamatsu, M.; Abe, K.; Nakagawa, K.; Tsugawa, N.; Hirota, Y.; Tanaka, K.; Yamashita, S.; Ikushiro, S.; et al. Generation of 1,25-dihydroxyvitamin D 3 in Cyp27b1 knockout mice by treatment with 25-hydroxyvitamin D 3 rescued their rachitic phenotypes. J. Steroid Biochem. Mol. Biol. 2019, 185, 71–79. [Google Scholar] [CrossRef]
- Nakatsukasa, A.; Kuruma, K.; Okamatsu, M.; Hiono, T.; Suzuki, M.; Matsuno, K.; Kida, H.; Oyamada, T.; Sakoda, Y. Potency of whole virus particle and split virion vaccines using dissolving microneedle against challenges of H1N1 and H5N1 influenza viruses in mice. Vaccine 2017, 35, 2855–2861. [Google Scholar] [CrossRef]
- Hiono, T.; Okamatsu, M.; Yamamoto, N.; Ogasawara, K.; Endo, M.; Kuribayashi, S.; Shichinohe, S.; Motohashi, Y.; Chu, D.H.; Suzuki, M.; et al. Experimental infection of highly and low pathogenic avian influenza viruses to chickens, ducks, tree sparrows, jungle crows, and black rats for the evaluation of their roles in virus transmission. Vet. Microbiol. 2016, 182, 108–115. [Google Scholar] [CrossRef]
- Ashton, J.C. ANOVA and the analysis of drug combination experiments. Nat. Methods 2015, 12, 1108. [Google Scholar] [CrossRef]
- Nagai, T.; Makino, S.; Ikegami, S.; Itoh, H.; Yamada, H. Effects of oral administration of yogurt fermented with Lactobacillus delbrueckii ssp. bulgaricus OLL1073R-1 and its exopolysaccharides against influenza virus infection in mice. Int. Immunopharmacol. 2011, 11, 2246–2250. [Google Scholar] [CrossRef]
- Cannell, J.J.; Vieth, R.; Umhau, J.C.; Holick, M.F.; Grant, W.B.; Madronich, S.; Garland, C.F.; Giovannucci, E. Epidemic influenza and vitamin D. Epidemiol. Infect. 2006, 134, 1129–1140. [Google Scholar] [CrossRef]
- Zhou, J.; Du, J.; Huang, L.; Wang, Y.; Shi, Y.; Lin, H. Preventive Effects of Vitamin D on Seasonal Influenza A in Infants: A Multicenter, Randomized, Open, Controlled Clinical Trial. Pediatr. Infect. Dis. J. 2018, 37, 749–754. [Google Scholar] [CrossRef] [PubMed]
- Daniela, C. Romero. and JoAnn Vitamin D and Your Heart. Circulation 2015, 132, e391–e392. [Google Scholar] [CrossRef][Green Version]
- Gui, B.; Chen, Q.; Hu, C.; Zhu, C.; He, G. Effects of calcitriol (1,25-dihydroxy-vitamin D3) on the inflammatory response induced by H9N2 influenza virus infection in human lung A549 epithelial cells and in mice. Virol. J. 2017, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- European Food Safe Authority, (EFSA). Scientific Opinion onthe Tolerable Upper Intake Level ofvitamin D. Available online: https://efsa.onlinelibrary.wiley.com/doi/pdf/10.2903/j.efsa.2012.2813 (accessed on 12 April 2020).
- Marcinowska-Suchowierska, E.; Kupisz-Urbanska, M.; Lukaszkiewicz, J.; Pludowski, P.; Jones, G. Vitamin D Toxicity-A clinical perspective. Front. Endocrinol. (Lausanne) 2018, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Deluca, H.F.; Prahl, J.M.; Plum, L.A. 1,25-Dihydroxyvitamin D is not responsible for toxicity caused by vitamin D or 25-hydroxyvitamin D. Arch. Biochem. Biophys. 2011, 505, 226–230. [Google Scholar] [CrossRef]
- Urashima, M.; Segawa, T.; Okazaki, M.; Kurihara, M.; Wada, Y.; Ida, H. Randomized trial of vitamin D supplementation to prevent seasonal influenza A in schoolchildren. Am. J. Clin. Nutr. 2010, 91, 1255–1260. [Google Scholar] [CrossRef]
- Mallya, S.M.; Corrado, K.R.; Saria, E.A.; Yuan, F.; Tran, H.Q.; Saucier, K.; Atti, E.; Tetradis, S.; Arnold, A. Modeling vitamin D insufficiency and moderate deficiency in adult mice via dietary cholecalciferol restriction. Endocr. Res. 2016, 41, 290–299. [Google Scholar] [CrossRef]
- Chan, R.W.Y.; Leung, C.Y.H.; Nicholls, J.M.; Peiris, J.S.M.; Chan, M.C.W. Proinflammatory Cytokine Response and Viral Replication in Mouse Bone Marrow Derived Macrophages Infected with Influenza H1N1 and H5N1 Viruses. PLoS ONE 2012, 7, 1–8. [Google Scholar] [CrossRef]
- Vogel, A.J.; Harris, S.; Marsteller, N.; Condon, S.A.; Brown, D.M. Early cytokine dysregulation and viral replication are associated with mortality during lethal influenza infection. Viral Immunol. 2014, 27, 214–224. [Google Scholar] [CrossRef]
- Chen, S.; Liu, G.; Chen, J.; Hu, A.; Zhang, L.; Sun, W.; Tang, W.; Liu, C.; Zhang, H.; Ke, C.; et al. Ponatinib protects mice from lethal influenza infection by suppressing cytokine storm. Front. Immunol. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Bzura, B.M. Vitamin D and influenza—Prevention or therapy? Int. J. Mol. Sci. 2018, 19, 2419. [Google Scholar] [CrossRef] [PubMed]
- Keating, P.; Munim, A.; Hartmann, J.X. Effect of vitamin D on T-helper type 9 polarized human memory cells in chronic persistent asthma. Ann. Allergy Asthma Immunol. 2014, 112, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhang, C.; Liu, Q.; Zhao, Y.; Zhang, Y.; Qin, Y.; Li, X.; Li, C.; Zhou, C.; Jin, N.; et al. Identification of amitriptyline HCl, flavin adenine dinucleotide, azacitidine and calcitriol as repurposing drugs for influenza A H5N1 virus-induced lung injury. PLoS Pathog. 2020, 16, 1–16. [Google Scholar] [CrossRef]
- Grant, W.B.; Giovannucci, E. The possible roles of solar ultraviolet-B radiation and vitamin D in reducing case-fatality rates from the 1918–1919 influenza pandemic in the United States. Dermatoendocrinology 2009, 1, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Grant, W.B.; Lahore, H.; McDonnell, S.L.; Baggerly, C.A.; French, C.B.; Aliano, J.L.; Bhattoa, H.P. Evidence that Vitamin D Supplementation Could Reduce Risk of Influenza and COVID-19 Infections and Deaths. Nutrients 2020, 12, 988. [Google Scholar] [CrossRef]
- Patel, N.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Surman, S.L.; Sun, Y.; Tang, L.; De Beauchamp, J.; Webb, A.; Richardson, J.; et al. Baseline serum Vitamin A and D levels determine benefit of oral Vitamin A&D supplements to humoral immune responses following pediatric influenza vaccination. Viruses 2019, 11, 907. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayashi, H.; Okamatsu, M.; Ogasawara, H.; Tsugawa, N.; Isoda, N.; Matsuno, K.; Sakoda, Y. Oral Supplementation of the Vitamin D Metabolite 25(OH)D3 Against Influenza Virus Infection in Mice. Nutrients 2020, 12, 2000. https://doi.org/10.3390/nu12072000
Hayashi H, Okamatsu M, Ogasawara H, Tsugawa N, Isoda N, Matsuno K, Sakoda Y. Oral Supplementation of the Vitamin D Metabolite 25(OH)D3 Against Influenza Virus Infection in Mice. Nutrients. 2020; 12(7):2000. https://doi.org/10.3390/nu12072000
Chicago/Turabian StyleHayashi, Hirotaka, Masatoshi Okamatsu, Honami Ogasawara, Naoko Tsugawa, Norikazu Isoda, Keita Matsuno, and Yoshihiro Sakoda. 2020. "Oral Supplementation of the Vitamin D Metabolite 25(OH)D3 Against Influenza Virus Infection in Mice" Nutrients 12, no. 7: 2000. https://doi.org/10.3390/nu12072000
APA StyleHayashi, H., Okamatsu, M., Ogasawara, H., Tsugawa, N., Isoda, N., Matsuno, K., & Sakoda, Y. (2020). Oral Supplementation of the Vitamin D Metabolite 25(OH)D3 Against Influenza Virus Infection in Mice. Nutrients, 12(7), 2000. https://doi.org/10.3390/nu12072000