Intermittent Hypoxic Exposure with High Dose of Arginine Impact on Circulating Mediators of Tissue Regeneration

 , ,
, ,
Abstract
1. Introduction
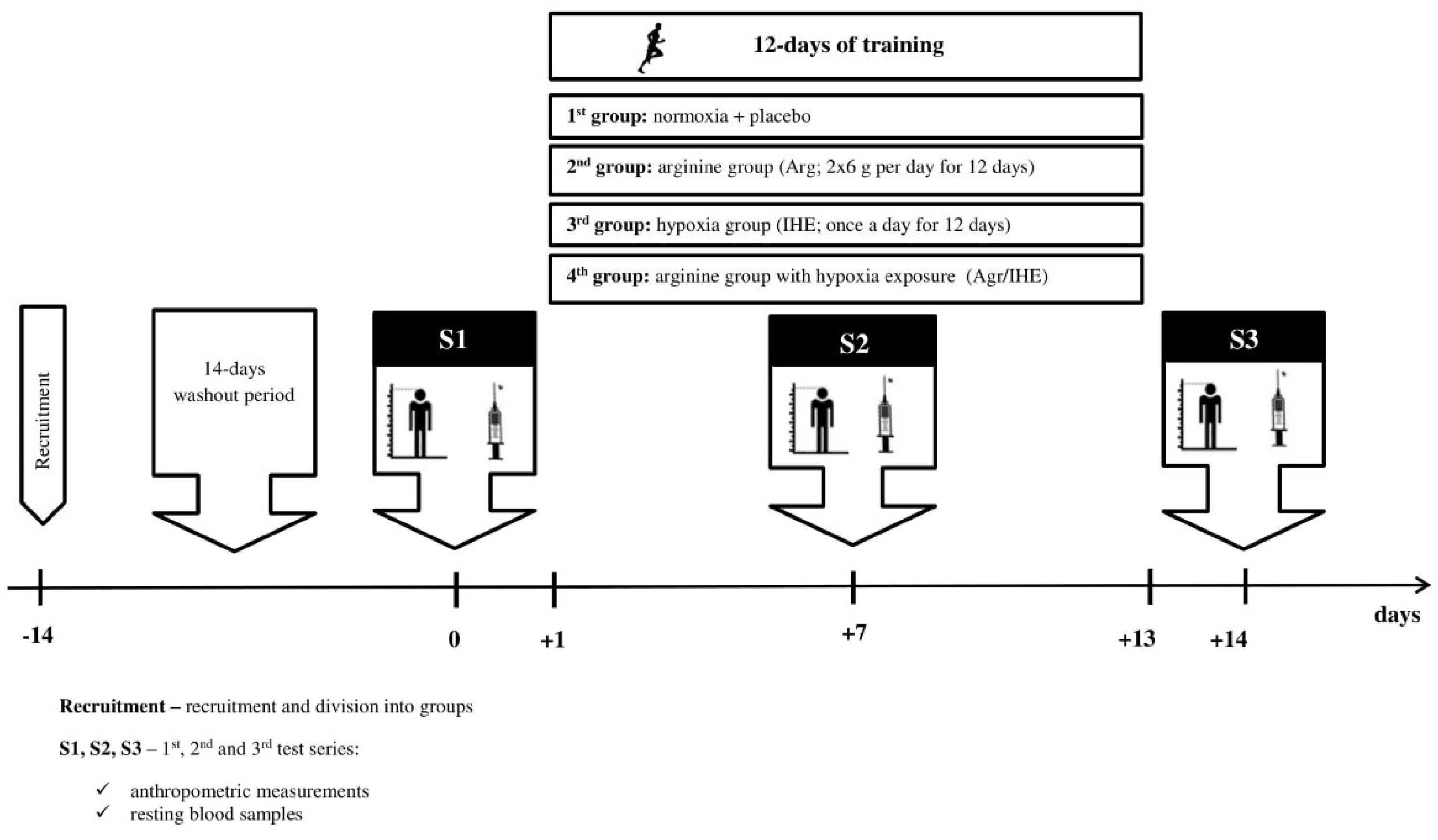
2. Materials and Methods
2.1. Subjects
2.2. Arginine Supplementation
2.3. Intermittent Hypoxic Exposure
2.4. Body Composition
2.5. Blood Sampling
2.6. Skeletal Muscle Damage
2.7. Oxi-Inflammatory Mediators
2.8. Growth Factors
2.9. Lipoprotein-Lipid Profile
2.10. Haematological Variables
2.11. Statistical Analysis
3. Results
3.1. Skeletal Muscle Damage
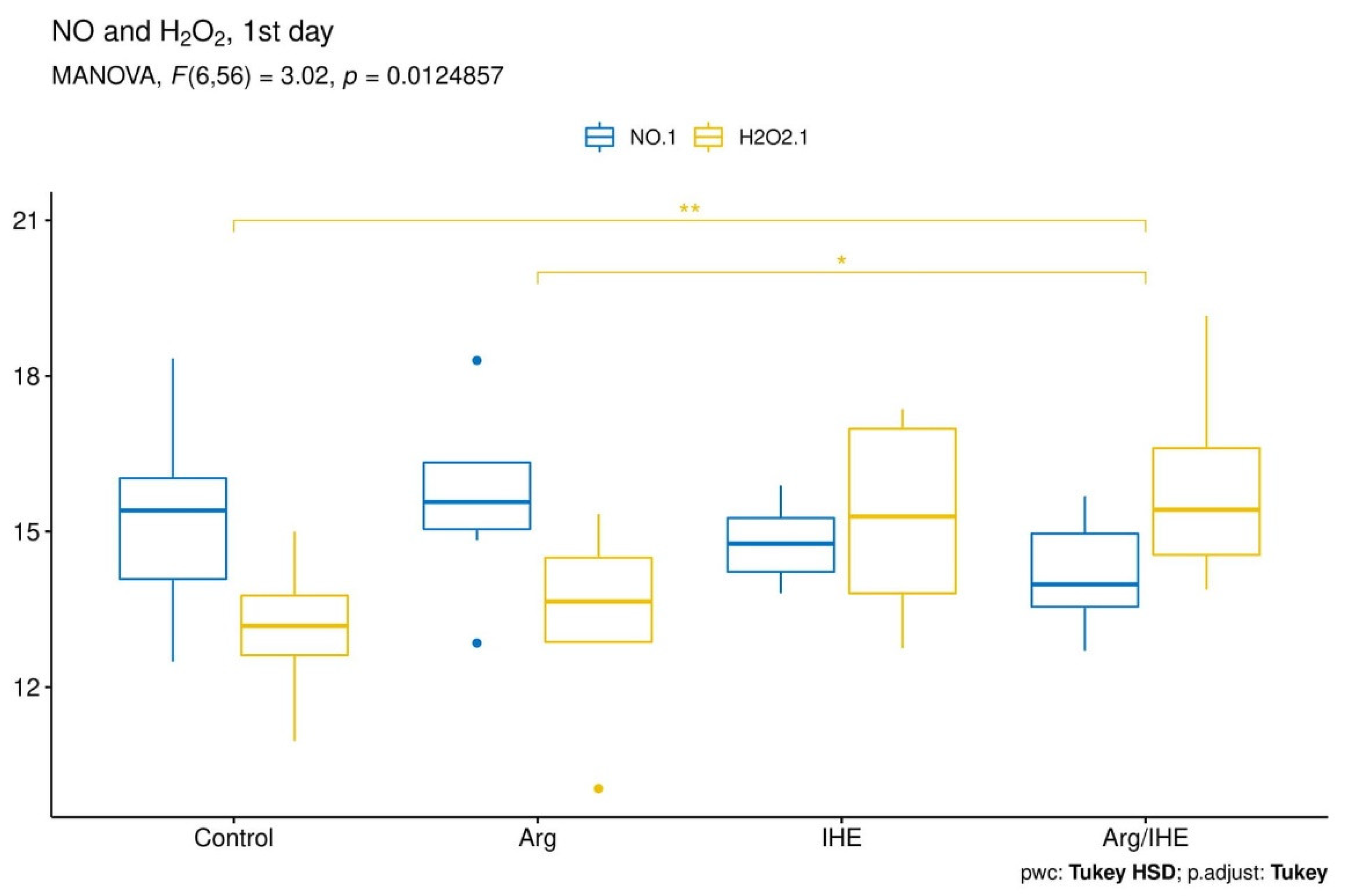
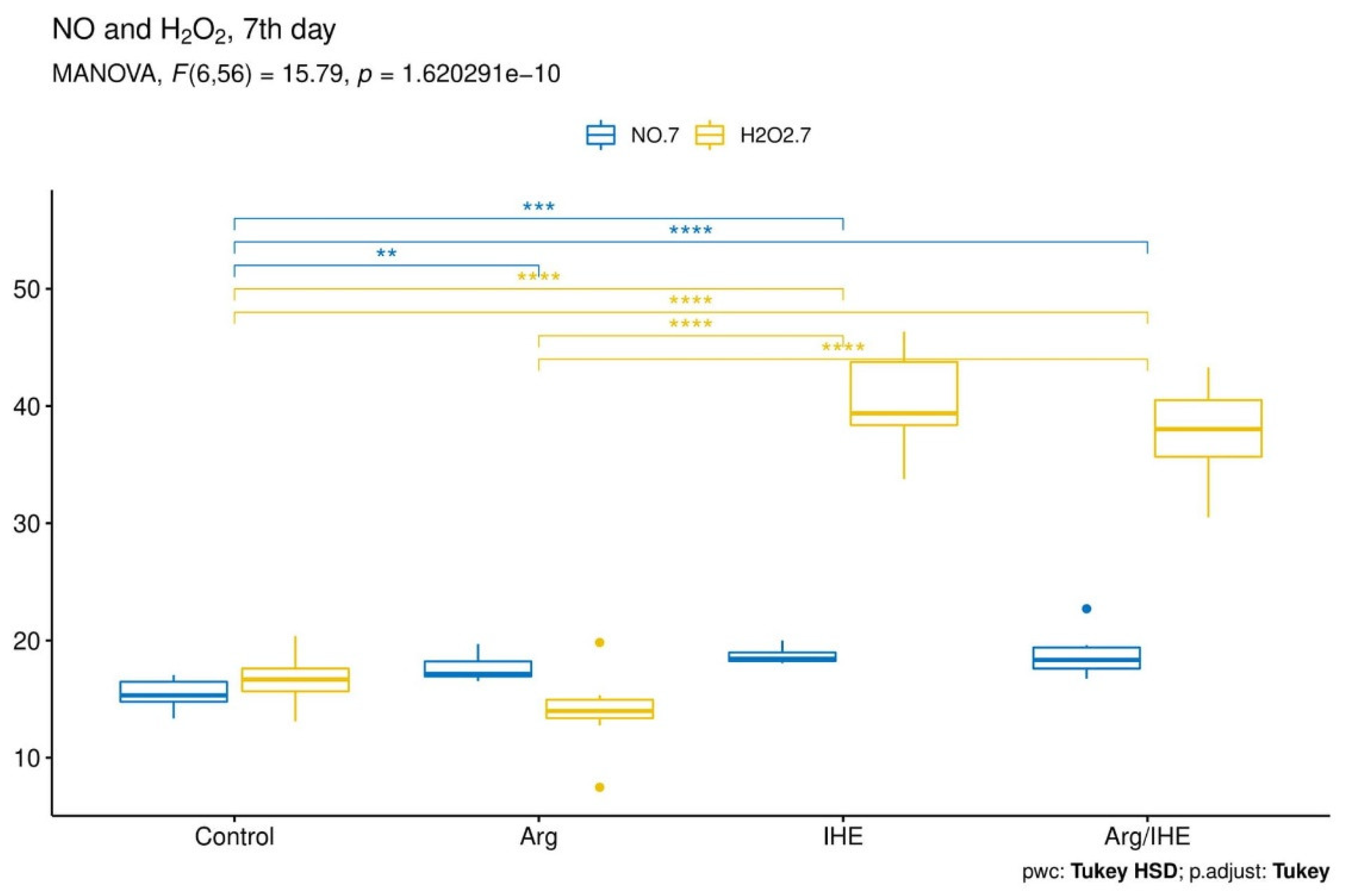
3.2. Oxi-Inflammatory Mediators
3.3. Growth Factors
3.4. Haematological Variables
3.5. Lipoprotein–Lipid Profile
4. Discussion
5. Conclusions
6. Limitations
Author Contributions
Funding
Conflicts of Interest
References
- Filippin, L.I.; Cuevas, M.J.; Lima, E.; Marroni, N.P.; Gonzalezgallego, J.; Xavier, R.M. Nitric oxide regulates the repair of injured skeletal muscle. Nitric Oxide 2011, 24, 43–49. [Google Scholar] [CrossRef]
- Kuang, S.; Gillespie, M.A.; Rudnicki, M.A. Niche regulation of muscle satellite cell self-renewal and differentiation. Cell Stem. Cell 2008, 2, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Tengan, C.H.; Rodrigues, G.S.; Godinho, R.O. Nitric oxide in skeletal muscle: Role on mitochondrial biogenesis and function. Int. J. Mol. Sci. 2012, 13, 17160–17184. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.M.; Davies, B.; Baker, J. Training in hypoxia: Modulation of metabolic and cardiovascular risk factors in men. Med. Sci. Sports Exerc. 2000, 32, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Czuba, M.; Bril, G.; Ploszczyca, K.; Piotrowicz, Z.; Chalimoniuk, M.; Roczniok, R.; Zembron-Lacny, A.; Gerasimuk, D.; Langfort, J. Intermittent hypoxic training at lactate threshold intensity improves aiming performance in well-trained biathletes with little change of cardiovascular variables. Biomed Res. Int. 2019. [Google Scholar] [CrossRef] [PubMed]
- Haufe, S.; Wiesner, S.; Engeli, S.; Luft, F.C.; Jordan, J. Influence of normobaric hypoxia training on metabolic risk markers in human subjects. Med. Sci. Sports Exerc. 2008, 40, 1939–1944. [Google Scholar] [CrossRef]
- Chaillou, T.; Lanner, J.T. Regulation of myogenesis and skeletal muscle regeneration: Effects of oxygen levels on satellite cell activity. FASEB J. 2016, 30, 3929–3941. [Google Scholar] [CrossRef]
- Savla, J.J.; Levine, B.D.; Sadek, H.A. The effect of hypoxia on cardiovascular disease: Friend or foe? High. Alt. Med. Biol. 2018, 19, 124–130. [Google Scholar] [CrossRef]
- Barbieri, E.; Sestili, P. Reactive oxygen species in skeletal muscle signaling. J. Signal. Transduct. 2012, 2012, 982794. [Google Scholar] [CrossRef] [PubMed]
- Nakada, Y.; Canseco, D.C.; Thet, S.; Abdisalaam, S.; Asaithamby, A.; Santos, C.X.; Shah, A.M.; Zhang, H.; Faber, J.E.; Kinter, M.T.; et al. Hypoxia induces heart regeneration in adult mice. Nature 2017, 541, 222–227. [Google Scholar] [CrossRef]
- Böger, R.H. The pharmacodynamics of L-arginine. J. Nutr. 2007, 137, 1650S–1655S. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.S.; Oh, S.K.; Lee, J.S.; Wu, C.; Lee, S.J. Effects of L-arginine on growth hormone and insulin-like growth factor 1. Food Sci. Biotechnol. 2017, 26, 1749–1754. [Google Scholar] [CrossRef]
- Curran, J.N.; Winter, D.C.; Bouchier-Hayes, D. Biological fate and clinical implications of arginine metabolism in tissue healing. Wound Rep. Reg. 2006, 14, 376–386. [Google Scholar] [CrossRef]
- Filippin, L.I.; Moreira, A.J.; Marroni, N.P.; Xavier, R.M. Nitric oxide and repair of skeletal muscle injury. Nitric Oxide 2009, 21, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Hinckson, E.A.; Hamlin, M.J.; Wood, M.R.; Hopkins, W.G. Game performance and intermittent hypoxic training. Br. J. Sports Med. 2007, 41, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, C.J. An Effect Size Primer: A Guide for Clinicians and Researchers. Prof. Psychol. Res. Pract. 2009, 40, 532–538. [Google Scholar] [CrossRef]
- Zembron-Lacny, A.; Ziemann, E.; Zurek, P.; Hübner-Wozniak, E. Heat shock protein 27 response to wrestling training in relation to the muscle damage and inflammation. J. Strength Cond. Res. 2017, 3, 1221–1228. [Google Scholar] [CrossRef]
- Tidball, J.G. Regulation of muscle growth and regeneration by the immune system. Nat. Rev. Immunol. 2017, 17, 165–178. [Google Scholar] [CrossRef]
- Ambrose, K.R.; Golightly, Y.M. Physical exercise as non-pharmacological treatment of chronic pain: Why and when. Best Pract. Res. Clin. Rheumatol. 2015, 29, 120–130. [Google Scholar] [CrossRef]
- Chang, J.C.; Lien, C.F.; Lee, W.S.; Chang, H.R.; Hsu, Y.C.; Luo, Y.P.; Jeng, J.R.; Hsieh, J.C.; Yang, K.T. Intermittent Hypoxia Prevents Myocardial Mitochondrial Ca2+ Overload and Cell Death during Ischemia/Reperfusion: The Role of Reactive Oxygen Species. Cells 2019. [Google Scholar] [CrossRef]
- Dobson, J.L.; McMillan, J.; Li, L. Benefits of exercise intervention in reducing neuropathic pain. Front. Cell Neurosci. 2014, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, A.; Marsh, D. Ischaemia-reperfusion modulates inflammation and fibrosis of skeletal muscle after contusion injury. Int. J. Exp. Pathol. 2010, 91, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Kashimura, O.; Kano, Y.; Ohno, H.; Ji, L.L.; Izawa, T.; Best, T.M. Role of nitric oxide in muscle regeneration following eccentric muscle contractions in rat skeletal muscle. J. Physiol. Sci. 2013, 63, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Filippin, L.I.; Cuevas, M.J.; Lima, E.; Marroni, N.P.; Gonzalez-Gallego, J.; Xavier, R.M. The role of nitric oxide during healing of trauma to the skeletal muscle. Inflamm. Res. 2011, 60, 347–356. [Google Scholar] [CrossRef]
- Soneja, A.; Drews, M.; Malinski, T. Role of nitric oxide, nitroxidative and oxidative stress in wound healing. Pharmacol. Rep. 2005, 57, 108–119. [Google Scholar]
- Zembron-Lacny, A.; Tylutka, A.; Zeromska, A.; Kasperska, A.; Wolny-Rokicka, E. Does high volume of exercise training increase aseptic vascular inflammation in male athletes? Am. J. Men’s Health 2019, 13. [Google Scholar] [CrossRef]
- Ding, H.L.; Zhu, H.F.; Dong, J.W.; Zhu, W.Z.; Yang, W.W.; Yang, H.T.; Zhou, Z.N. Inducible nitric oxide synthase contributes to intermittent hypoxia against ischemia/reperfusion injury. Acta Pharmacol. Sin. 2005, 26, 315–322. [Google Scholar] [CrossRef]
- Grebe, A.; Hoss, F.; Latz, E. NLRP3 inflammasome and the IL-1 pathway in atherosclerosis. Circ. Res. 2018, 122, 1722–1740. [Google Scholar] [CrossRef]
- Strijdom, H.; Muller, C.; Lochner, A. Direct intracellular nitric oxide detection in isolated adult cardiomyocytes: Flow cytometric analysis using the fluorescent probe, diaminofluorescein. J. Mol. Cell. Cardiol. 2004, 37, 897–902. [Google Scholar] [CrossRef]
- Strijdom, H.; Jacobs, S.; Hattingh, S.; Page, C.; Lochner, A. Nitric oxide production is higher in rat cardiac microvessel endothelial cells than ventricular cardiomyocytes in baseline and hypoxic conditions: A comparative study. FASEB J. 2006, 20, 314–316. [Google Scholar] [CrossRef]
- Vogt, M.; Hoppeler, H. Is hypoxia training good for muscles and exercise performance? Prog. Cardiovasc. Dis. 2010, 52, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Kolár, F.; Szárszoi, O.; Neckárˇ, J.; Pecháňová, O.; Miková, D.; Hampl, V.; Oštádal, B. Role of nitric oxide and reactive oxygen species in reperfusion-induced arrhythmias and cardioprotection in chronically hypoxic rat. hearts. Physiol. Res. 2003, 52, 52. [Google Scholar]
- Brancaccio, P.; Lippi, G.; Maffulli, N. Biochemical markers of muscular damage. Clin. Chem. Lab. Med. 2010, 48, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.L.; Widstrom, L.; Goodrich, J.; Poddar, S.; Rueda, M.; Holliday, M.; San Millian, I.; Byrnes, W.C. A Retrospective Analysis of Collegiate Athlete Blood Biomarkers at Moderate Altitude. J. Strength Cond. Res. 2019, 33, 2913–2919. [Google Scholar] [CrossRef] [PubMed]
- Hoppeler, H.; Vogt, M. Muscle tissue adaptations to hypoxia. J. Exp. Biol. 2001, 204, 3133–3139. [Google Scholar] [PubMed]
- Alvares, T.S.; Conte-Junior, C.A.; Silva, J.T.; Paschoalin, V.M. L-arginine does not improve biochemical and hormonal response in trained runners after 4 weeks of supplementation. Nutr. Res. 2014, 34, 31–39. [Google Scholar] [CrossRef]
- Forbes, S.C.; Bell, G.J. The acute effects of a low and high dose of oral L-arginine supplementation in young active males at rest. Appl. Physiol. Nutr. Metab. 2011, 36, 405–411. [Google Scholar] [CrossRef]
- Forbes, S.C.; Harber, V.; Bell, G.J. The acute effects of L-arginine on hormonal and metabolic responses during submaximal exercise in trained cyclists. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 369–377. [Google Scholar] [CrossRef]
- Meirelles, C.M.; Matsuura, C. Acute supplementation of L-arginine affects neither strength performance nor nitric oxide production. J. Sports Med. Phys. Fit. 2018, 58, 216–220. [Google Scholar]
- Meirelles, C.M.; Matsuura, C.; Silva, R.S., Jr.; Guimarães, F.F.; Gomes, P.S.C. Acute effects of l-arginine supplementation on oxygen consumption kinetics and muscle oxyhemoglobin and deoxyhemoglobin during treadmill running in male adults. Int. J. Exerc. Sci. 2019, 12, 444–455. [Google Scholar]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zembron-Lacny, A.; Krzywański, J.; Ostapiuk-Karolczuk, J.; Kasperska, A. Cell and molecular mechanisms of regeneration and reorganization of skeletal muscles. Ortop. Traumatol. Rehabil. 2012, 14, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Breen, E.; Tang, K.; Olfert, M.; Knapp, A.; Wagner, P. Skeletal muscle capillarity during hypoxia: VEGF and its activation. High Alt. Med. Biol. 2008, 9, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Shashar, M.; Chernichovski, T.; Pasvolsky, O.; Levi, S.; Grupper, A.; Hershkovitz, R.; Weinstein, T.; Schwartz, I.F. Vascular Endothelial Growth Factor Augments Arginine Transport and Nitric Oxide Generation via a KDR Receptor Signaling Pathway. Kidney Blood Press. Res. 2017, 42, 201–208. [Google Scholar] [CrossRef]
- Gianni Barrera, R.; Di Maggio, N.; Melly, L.; Burger, M.G.; Mujagic, E.; Gürke, L.; Schaefer, D.J.; Banfi, A. Therapeutic vascularization in regenerative medicine. Stem Cells Transl. Med. 2020, 9, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.E. Hepatocyte growth factor and satellite cell activation. Adv. Exp. Med. Biol. 2016, 900, 1–25. [Google Scholar]
- Pedersen, B.K.; Pedersen, M.; Krabbe, K.S.; Bruunsgaard, H.; Matthews, V.B.; Febbraio, M.A. Role of exercise-induced brain-derived neurotrophic factor production in the regulation of energy homeostasis in mammals. Exp. Physiol. 2009, 94, 1153–1160. [Google Scholar] [CrossRef]
- Ogborn, D.I.; Gardiner, P.F. Effects of exercise and muscle type on BDNF, NT-4/5, and TrKB expression in skeletal muscle. Muscle Nerve 2010, 41, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, P.; Brassard, P.; Adser, H.; Pedersen, M.V.; Leick, L.; Hart, E.; Secher, N.H.; Pedersen, B.K.; Pilegaard, H. Evidence for a release of brain-derived neurotrophic factor from the brain during exercise. Exp. Physiol. 2009, 94, 1062–1069. [Google Scholar] [CrossRef]
- Lieberman, P.; Protopapas, A.; Reed, E.; Youngs, J.W.; Kanki, B.G. Cognitive defects at altitude. Nature 1994, 372, 325. [Google Scholar] [CrossRef]
- Amann, M.; Romer, L.M.; Subudhi, A.W.; Pegelow, D.F.; Dempsey, J.A. Severity, of arterial hypoxaemia affects the relative contributions of peripheral muscle fatigue to exercise performance in healthy humans. J. Physiol. 2007, 581, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Piotrowicz, Z.; Chalimoniuk, M.; Płoszczyca, K.K.; Czuba, M.; Langfort, J. Acute normobaric hypoxia does not affect the simultaneous exercise-induced increase in circulating BDNF and GDNF in young healthy men: A feasibility study. PLoS ONE 2019, 14, e0224207. [Google Scholar] [CrossRef] [PubMed]
- Suresh, S.; Rajvanshi, P.K.; Noguchi, C.T. The many facets of erythropoietin physiologic and metabolic response. Front. Physiol. 2019, 10, 1534. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Suzuki, N.; Yamamoto, M.; Gassmann, M.; Noguchi, C.T. Endogenous erythropoietin signaling facilitates skeletal muscle repair and recovery following pharmacologically induced damage. FASEB J. 2012, 26, 2847–2858. [Google Scholar] [CrossRef]
- Heeschen, C.; Aicher, A.; Lehmann, R.; Fichtlscherer, S.; Vasa, M.; Urbich, C.; Mildner-Rihm, C.; Martin, H.; Zeiher, A.M.; Dimmeler, S. Erythropoietin is a potent physiologic stimulus for endothelial progenitor cell mobilization. Blood 2003, 102, 1340–1346. [Google Scholar] [CrossRef]
- Beleslin-Cokic, B.B.; Cokic, V.P.; Yu, X.; Weksler, B.B.; Schechter, A.N.; Noguchi, C.T. Erythropoietin and hypoxia stimulate erythropoietin receptor and nitric oxide production by endothelial cells. Blood 2004, 104, 2073–2080. [Google Scholar] [CrossRef]
- Mallet, R.T.; Manukhina, E.B.; Ruelas, S.S.; Caffrey, J.L.; Downey, H.F. Cardioprotection by intermittent hypoxia conditioning: Evidence, mechanisms, and therapeutic potential. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H216–H232. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control n = 10 | Arg n = 7 | IHE n = 6 | Arg/IHE n = 9 | Control vs. Arg IHE Arg/IHE | |
|---|---|---|---|---|---|
| Age [yr.] | 24.6 ± 3.0 | 20.0 ± 1.6 | 22.8 ± 2.6 | 24.7 ± 4.4 | <0.05 0.622 0.999 |
| Height [cm] | 173.6 ± 8.8 | 179.0 ± 9.5 | 181.2 ± 7.3 | 175.6 ± 8.3 | 0.559 0.320 0.947 |
| Weight [kg] | 81.4 ± 21.8 | 79.9 ± 13.0 | 97.1 ± 22.7 | 87.9 ± 20.7 | 0.989 0.560 0.957 |
| BMI [kg/m2] | 26.6 ± 4.5 | 24.4 ± 1.4 | 29.3 ± 5.2 | 27.8 ± 4.5 | 0.552 0.773 0.990 |
| %FM | 18.1 ± 4.8 | 9.3 ± 3.0 | 14.5 ± 6.0 | 21.3 ± 6.5 | <0.05 0.844 0.252 |
| FM [kg] | 15.4 ± 7.4 | 7.6 ± 3.2 | 15.1 ± 9.5 | 19.3 ± 10.8 | 0.355 0.998 0.569 |
| FFM [kg] | 66.0 ± 15.0 | 72.3 ± 10.6 | 81.9 ± 14.0 | 67.2 ± 11.5 | 0.952 0.270 0.996 |
| 1st Day of Camp | 7th Day of Camp | 14th Day of Camp | 1st Day vs. 7th Day | 1st Day vs. 14th Day | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | |||
| NO [µmol/L] | |||||||||||
| Control | 15.37 ± 1.83 | - | 0.166 | 15.36 ± 1.26 | - | 0.569 | 17.33 ± 0.99 | - | 0.701 | 0.995 | <0.05 |
| Arg | 15.64 ± 1.66 | 0.98 | 17.67 ± 1.19 | <0.01 | 18.97 ± 1.67 | 0.326 | <0.01 | <0.01 | |||
| IHE | 14.78 ± 0.79 | 0.856 | 18.71 ± 0.73 | <0.001 | 22.26 ± 0.95 | <0.001 | <0.05 | <0.001 | |||
| Arg/IHE | 14.12 ± 0.95 | 0.251 | 18.67 ± 1.78 | <0.001 | 23.93 ± 3.02 | <0.001 | <0.001 | <0.01 | |||
| H2O2 [µmol/L] | |||||||||||
| Control | 13.16 ± 1.13 | - | 0.382 | 16.67 ± 2.00 | - | 0.92 | 17.87 ± 3.15 | - | 0.678 | <0.001 | <0.01 |
| Arg | 13.39 ± 1.76 | 0.992 | 13.99 ± 3.66 | 0.46 | 17.34 ± 4.45 | 0.994 | 0.748 | <0.05 | |||
| IHE | 15.26 ± 1.99 | 0.098 | 40.36 ± 4.64 | <0.001 | 30.93 ± 4.81 | <0.001 | <0.001 | <0.001 | |||
| Arg/IHE | 15.97 ± 1.91 | <0.01 | 39.47 ± 4.36 | <0.001 | 27.41 ± 4.63 | <0.001 | <0.001 | <0.001 | |||
| NO/H2O2 ratio [µmol/L] | |||||||||||
| Control | 1.17 ± 0.10 | - | 0.456 | 0.93 ± 0.13 | - | 0.99 ± 0.15 | - | 0.386 | <0.01 | <0.05 | |
| Arg | 1.19 ± 0.23 | 0.986 | 1.35 ± 0.42 | <0.01 | 1.13 ± 0.19 | 0.383 | 0.505 | 0.675 | |||
| IHE | 0.98 ± 0.13 | 0.095 | 0.47 ± 0.05 | <0.01 | 0.74 ± 0.15 | <0.05 | <0.001 | <0.05 | |||
| Arg/IHE | 0.90 ± 0.12 | <0.01 | 0.51 ± 0.10 | <0.01 | 0.90 ± 0.20 | 0.615 | <0.001 | 0.974 | |||
| CRP [mg/L] | |||||||||||
| Control | 1.57 ± 0.53 | - | 0.024 | 2.02 ± 0.35 | - | 0.552 | 2.29 ± 0.48 | - | 0.115 | <0.05 | <0.01 |
| Arg | 1.62 ± 0.49 | 0.997 | 1.99 ± 0.55 | 0.999 | 2.14 ± 0.41 | 0.917 | 0.182 | 0.097 | |||
| IHE | 1.45 ± 0.15 | 0.968 | 2.69 ± 0.41 | <0.05 | 2.46 ± 0.28 | 0.913 | <0.001 | <0.001 | |||
| Arg/IHE | 1.66 ± 0.57 | 0.977 | 3.09 ± 0.52 | <0.001 | 2.58 ± 0.63 | 0.589 | <0.001 | <0.01 | |||
| 1st Day of Camp | 7th Day of Camp | 14th Day of Camp | 1st Day vs. 7th Day | 1st Day vs. 14th Day | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | |||
| HGF [pg/mL] | |||||||||||
| Control | 587 ± 73 | - | 0.237 | 534 ± 77 | - | 0.467 | 602 ± 68 | - | 0.653 | <0.01 | 0.625 |
| Arg | 620 ± 60 | 0.687 | 568 ± 89 | 0.827 | 770 ± 69 | <0.001 | <0.05 | <0.01 | |||
| IHE | 623 ± 61 | 0.666 | 546 ± 95 | 0.992 | 792 ± 49 | <0.001 | 0.094 | <0.05 | |||
| Arg/IHE | 544 ± 42 | 0.425 | 708 ± 72 | <0.001 | 789 ± 71 | <0.001 | <0.001 | <0.001 | |||
| IGF-1 [ng/mL] | |||||||||||
| Control | 120 ± 41 | - | 0.19 | 116 ± 22 | - | 0.867 | 100 ± 15 | - | 0.911 | 0.16 | 0.084 |
| Arg | 126 ± 23 | 0.982 | 103 ± 19 | 0.834 | 98 ± 12 | 0.998 | 0.073 | <0.01 | |||
| IHE | 149 ± 11 | 0.231 | 248 ± 26 | <0.001 | 206 ± 29 | <0.001 | <0.001 | <0.01 | |||
| Arg/IHE | 148 ± 26 | 0.186 | 280 ± 51 | <0.001 | 293 ± 43 | <0.001 | <0.001 | <0.001 | |||
| PDGFBB [pg/mL] | |||||||||||
| Control | 2281 ± 513 | - | 0.084 | 2646 ± 289 | - | 0.381 | 2068 ± 368 | - | 0.706 | 0.074 | 0.063 |
| Arg | 2327 ± 418 | 0.995 | 2836 ± 293 | 0.806 | 1980 ± 161 | 0.927 | 0.062 | 0.088 | |||
| IHE | 2582 ± 231 | 0.456 | 2762 ± 160 | 0.953 | 3133 ± 263 | <0.001 | 0.15 | <0.01 | |||
| Arg/IHE | 2307 ± 266 | 0.999 | 3418 ± 686 | <0.01 | 2478 ± 274 | <0.05 | <0.01 | 0.226 | |||
| BDNF [pg/mL] | |||||||||||
| Control | 23,447 ± 3237 | - | 0.067 | 27,486 ± 1974 | - | 0.781 | 27,426 ± 2452 | - | 0.789 | <0.05 | <0.01 |
| Arg | 23,922 ± 3040 | 0.987 | 29,567 ± 2651 | 0.301 | 26,626 ± 1250 | 0.88 | <0.05 | 0.073 | |||
| IHE | 22,402 ± 3184 | 0.899 | 18,817 ± 1118 | <0.001 | 18,154 ± 1377 | <0.001 | 0.059 | 0.053 | |||
| Arg/IHE | 22,120 ± 2177 | 0.756 | 21,218 ± 3025 | <0.001 | 19,952 ± 2791 | <0.001 | 0.198 | <0.05 | |||
| VEGF [pg/mL] | |||||||||||
| Control | 341 ± 68 | - | 0.03 | 405 ± 54 | - | 0.112 | 234 ± 65 | - | 0.782 | 0.085 | <0.001 |
| Arg | 361 ± 64 | 0.861 | 408 ± 63 | 0.958 | 238 ± 77 | 0.999 | 0.154 | <0.05 | |||
| IHE | 330 ± 44 | 0.999 | 406 ± 46 | 0.974 | 389 ± 45 | <0.001 | <0.05 | 0.135 | |||
| Arg/IHE | 344 ± 78 | 0.991 | 452 ± 97 | 0.284 | 495 ± 64 | <0.001 | 0.054 | <0.01 | |||
| HGF [pg/mL] | IGF-1 [ng/mL] | PDGFBB [pg/mL] | BDNF [pg/mL] | VEGF [pg/mL] | |
|---|---|---|---|---|---|
| NO [µmol/L] | 0.662 | 0.554 | 0.160 | −0.286 | 0.274 |
| <0.001 | <0.001 | >0.05 | <0.01 | <0.01 | |
| H2O2 [µmol/L] | 0.321 | 0.780 | 0.479 | −0.525 | 0.368 |
| <0.01 | <0.001 | <0.001 | <0.001 | <0.001 |
| 1st Day of Camp | 7th Day of Camp | 14th Day of Camp | 1st Day vs. 7th Day | 1st day vs. 14th day | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | |||
| HB [g/dL] | |||||||||||
| Control | 15.3 ± 0.8 | - | 0.043 | 15.2 ± 0.7 | - | 0.165 | 15.5 ± 0.2 | - | 0.462 | 0.918 | 0.335 |
| Arg | 14.9 ± 0.8 | 0.757 | 14.8 ± 0.5 | 0.617 | 14.6 ± 0.3 | <0.01 | 0.271 | 0.306 | |||
| IHE | 15.1 ± 0.8 | 0.961 | 14.4 ± 1.0 | 0.137 | 14.3 ± 0.6 | <0.01 | 0.071 | 0.218 | |||
| Arg/IHE | 15.3 ± 0.7 | 1 | 15.1 ± 0.7 | 0.947 | 15.2 ± 0.8 | 0.517 | 0.16 | 0.851 | |||
| RBC [mln/mm3] | |||||||||||
| Control | 5.4 ± 0.3 | - | 0.189 | 5.4 ± 0.2 | - | 0.392 | 5.1 ± 0.2 | - | 0.736 | 0.411 | <0.05 |
| Arg | 5.3 ± 0.5 | 0.781 | 5.2 ± 0.3 | 0.286 | 4.6 ± 0.1 | <0.001 | 0.636 | <0.05 | |||
| IHE | 5.2 ± 0.4 | 0.852 | 5.0 ± 0.5 | <0.05 | 4.7 ± 0.3 | <0.01 | 0.055 | <0.05 | |||
| Arg/IHE | 5.0 ± 0.2 | 0.076 | 4.9 ± 0.2 | <0.01 | 5.5 ± 0.3 | <0.05 | 0.065 | <0.01 | |||
| RET [‰] | |||||||||||
| Control | 4.1 ± 1.1 | - | 0.222 | 5.2 ± 1.2 | - | 0.459 | 7.1 ± 2.2 | - | 0.505 | <0.05 | <0.01 |
| Arg | 4.1 ± 1.2 | 1 | 5.0 ± 1.4 | 0.994 | 10.1 ± 1.8 | <0.05 | 0.2 | <0.001 | |||
| IHE | 3.0 ± 0.6 | 0.139 | 8.7 ± 2.5 | <0.001 | 12.0 ± 1.4 | <0.001 | <0.01 | <0.001 | |||
| Arg/IHE | 3.3 ± 0.7 | 0.295 | 6.4 ± 1.1 | 0.319 | 9.3 ± 1.7 | 0.071 | <0.001 | <0.01 | |||
| HCT [%] | |||||||||||
| Control | 48.1 ± 3.0 | - | 0.4 | 48.2 ± 2.8 | - | 0.45 | 45.8 ± 1.5 | - | 0.403 | 0.825 | <0.05 |
| Arg | 47.9 ± 3.0 | 0.996 | 47.2 ± 1.8 | 0.878 | 43.0 ± 1.2 | <0.01 | 0.444 | <0.01 | |||
| IHE | 45.7 ± 1.1 | 0.221 | 45.1 ± 2.4 | 0.164 | 44.2 ± 2.4 | 0.176 | 0.598 | 0.096 | |||
| Arg/IHE | 43.8 ± 1.7 | <0.01 | 42.3 ± 3.7 | <0.001 | 45.8 ± 1.0 | 1 | 0.41 | <0.05 | |||
| MCV [fL] | |||||||||||
| Control | 89.3 ± 3.9 | - | 0.316 | 89.9 ± 3.1 | - | 0.427 | 89.8 ± 2.0 | - | 0.732 | 0.43 | 0.626 |
| Arg | 92.1 ± 3.0 | 0.35 | 91.8 ± 2.8 | 0.559 | 93.5 ± 0.3 | <0.05 | 0.647 | 0.278 | |||
| IHE | 85.5 ± 3.4 | 0.163 | 84.8 ± 2.4 | <0.05 | 83.8 ± 1.9 | <0.001 | 0.328 | 0.067 | |||
| Arg/IHE | 88.0 ± 3.2 | 0.842 | 87.8 ± 3.1 | 0.406 | 85.0 ± 3.5 | <0.001 | 0.681 | <0.05 | |||
| MCH [pg/RBC] | |||||||||||
| Control | 28.4 ± 1.0 | - | 0.393 | 28.5 ± 1.5 | - | 0.311 | 30.5 ± 0.9 | - | 0.643 | 1 | <0.001 |
| Arg | 28.7 ± 1.5 | 0.963 | 28.6 ± 0.9 | 1 | 32.0 ± 0.6 | <0.05 | 1 | <0.001 | |||
| IHE | 28.8 ± 1.4 | 0.934 | 29.7 ± 1.9 | 0.377 | 28.5 ± 1.0 | <0.01 | 0.079 | 0.451 | |||
| Arg/IHE | 30.7 ± 1.2 | <0.01 | 30.6 ± 1.1 | <0.05 | 28.6 ± 1.5 | <0.01 | 0.681 | <0.05 | |||
| MCHC [g/dL] | |||||||||||
| Control | 31.7 ± 0.9 | - | 0.826 | 31.7 ± 1.1 | - | 0.863 | 33.7 ± 0.4 | - | 0.158 | 0.968 | <0.01 |
| Arg | 31.0 ± 0.6 | 0.241 | 31.2 ± 0.3 | 0.522 | 34.2 ± 0.3 | 0.221 | 0.359 | <0.05 | |||
| IHE | 33.5 ± 0.2 | <0.001 | 35.1 ± 0.9 | <0.001 | 33.7 ± 0.6 | 1 | <0.001 | 0.336 | |||
| Arg/IHE | 34.8 ± 0.8 | <0.001 | 34.9 ± 0.3 | <0.001 | 33.8 ± 0.5 | 1 | 0.155 | <0.05 | |||
| RDW [%] | |||||||||||
| Control | 15,2 ± 1,7 | - | 0.525 | 15.0 ± 1.3 | - | 0.594 | 15,0 ± 0.9 | - | 0.85 | 0.797 | 0.743 |
| Arg | 14,7 ± 0.4 | 0.839 | 14.9 ± 0.6 | 0.973 | 14.5 ± 0.0 | 0.282 | 0.352 | 0.26 | |||
| IHE | 12.4 ± 0.4 | <0.001 | 12.4 ± 0.4 | <0.001 | 12.4 ± 0.3 | <0.001 | 0.741 | 0.618 | |||
| Arg/IHE | 15.3 ± 0.9 | 0.991 | 14.9 ± 0.6 | 0.975 | 12.3 ± 0.4 | <0.001 | 0.214 | <0.001 | |||
| EPO [mIU/mL] | |||||||||||
| Control | 3.25 ± 0.88 | - | 0.618 | 4.3 ± 1.37 | - | 0.466 | 4.84 ± 1.17 | - | 0.654 | <0.01 | <0.01 |
| Arg | 3.51 ± 0.42 | 0.922 | 6.08 ± 1.04 | <0.05 | 4.40 ± 1.68 | 0.994 | 0.618 | <0.05 | |||
| IHE | 3.12 ± 0.86 | 0.99 | 6.57 ± 0.71 | <0.01 | 9.70 ± 2.32 | <0.001 | <0.01 | <0.001 | |||
| Arg/IHE | 4.62 ± 0.8 | <0.01 | 6.49 ± 0.73 | <0.01 | 8.13 ± 1.62 | <0.001 | <0.001 | <0.01 | |||
| WBC [103/µL] | |||||||||||
| Control | 5.9 ± 1.0 | - | 0.149 | 6.7 ± 0.7 | - | 0.25 | 6.9 ± 0.4 | - | 0.173 | 0.066 | <0.05 |
| Arg | 5.6 ± 0.1 | 0.774 | 5.9 ± 0.8 | 0.3 | 6.5 ± 0.8 | 0.957 | 0.281 | <0.05 | |||
| IHE | 5.0 ± 0.3 | 0.148 | 5.3 ± 0.7 | <0.05 | 6.1 ± 0.3 | 0.619 | 0.114 | <0.01 | |||
| Arg/IHE | 5.5 ± 1.0 | 0.654 | 6.2 ± 1.2 | 0.699 | 7.6 ± 2.2 | 0.591 | 0.322 | <0.05 | |||
| 1st Day of Camp | 7th Day of Camp | 14th Day of Camp | 1st Day vs. 7th Day | 1st Day vs. 14th Day | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | mean ± SD | Control vs. Arg, IHE or Arg/IHE | η2 | |||
| TG [mg/dL] | |||||||||||
| Control | 97 ± 34 | - | 0.057 | 83 ± 24 | - | 0.231 | 118 ± 42 | - | 0.359 | <0.05 | 0.31 |
| Arg | 85 ± 24 | 0.925 | 76 ± 12 | 0.951 | 64 ± 4 | <0.01 | 0.331 | 0.057 | |||
| IHE | 111 ± 41 | 0.898 | 112 ± 18 | 0.138 | 122 ± 27 | 0.995 | 0.975 | 0.357 | |||
| Arg/IHE | 105 ± 50 | 0.969 | 78 ± 35 | 0.978 | 111 ± 32 | 0.964 | 0.06 | 0.764 | |||
| TC [mg/dL] | |||||||||||
| Control | 196 ± 25 | - | 0.143 | 189 ± 38 | - | 0.315 | 175 ± 19 | - | 0.306 | 0.323 | <0.05 |
| Arg | 160 ± 47 | 0.238 | 138 ± 25 | <0.05 | 148 ± 7 | 0.123 | 0.097 | 0.522 | |||
| IHE | 188 ± 38 | 0.976 | 161 ± 31 | 0.399 | 163 ± 27 | 0.789 | <0.05 | <0.05 | |||
| Arg/IHE | 196 ± 41 | 1 | 189 ± 34 | 1 | 188 ± 30 | 0.601 | 0.399 | 0.359 | |||
| LDL [mg/dL] | |||||||||||
| Control | 118 ± 27 | - | 0.149 | 121 ± 33 | - | 0.173 | 111 ± 18 | - | 0.059 | 0.707 | 0.232 |
| Arg | 93 ± 32 | 0.395 | 77 ± 17 | 0.555 | 97 ± 10 | 0.692 | 0.094 | 0.698 | |||
| IHE | 112 ± 38 | 0.985 | 103 ± 38 | 0.956 | 105 ± 37 | 0.963 | 0.406 | 0.386 | |||
| Arg/IHE | 128 ± 35 | 0.913 | 157 ± 117 | 0.664 | 112 ± 30 | 0.999 | 0.432 | 0.129 | |||
| HDL [mg/dL] | |||||||||||
| Control | 64 ± 43 | - | 0.094 | 51 ± 11 | - | 0.114 | 53 ± 10 | - | 0.188 | 0.234 | 0.76 |
| Arg | 50 ± 13 | 0.716 | 45 ± 9 | 0.675 | 53 ± 5 | 0.996 | 0.182 | 0.47 | |||
| IHE | 45 ± 10 | 0.498 | 56 ± 12 | 0.787 | 48 ± 7 | 0.667 | <0.01 | <0.05 | |||
| Arg/IHE | 47 ± 7 | 0.493 | 49 ± 7 | 0.998 | 45 ± 8 | 0.176 | <0.05 | 0.641 | |||
| Non-HDL [mg/dL] | |||||||||||
| Control | 132 ± 44 | - | 0.126 | 138 ± 36 | - | 0.312 | 122 ± 15 | - | 0.385 | 1 | 0.454 |
| Arg | 110 ± 35 | 1 | 92 ± 17 | <0.05 | 95 ± 2 | 0.105 | 0.084 | 0.298 | |||
| IHE | 143 ± 42 | 1 | 106 ± 39 | 0.234 | 116 ± 28 | 0.951 | <0.05 | <0.001 | |||
| Arg/IHE | 149 ± 40 | 1 | 139 ± 31 | 1 | 143 ± 33 | 0.215 | 0.205 | 0.313 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zembron-Lacny, A.; Gramacki, A.; Wawrzyniak-Gramacka, E.; Tylutka, A.; Hertmanowska, N.; Kasperska, A.; Czuba, M. Intermittent Hypoxic Exposure with High Dose of Arginine Impact on Circulating Mediators of Tissue Regeneration. Nutrients 2020, 12, 1933. https://doi.org/10.3390/nu12071933
Zembron-Lacny A, Gramacki A, Wawrzyniak-Gramacka E, Tylutka A, Hertmanowska N, Kasperska A, Czuba M. Intermittent Hypoxic Exposure with High Dose of Arginine Impact on Circulating Mediators of Tissue Regeneration. Nutrients. 2020; 12(7):1933. https://doi.org/10.3390/nu12071933
Chicago/Turabian StyleZembron-Lacny, Agnieszka, Artur Gramacki, Edyta Wawrzyniak-Gramacka, Anna Tylutka, Natalia Hertmanowska, Anna Kasperska, and Miłosz Czuba. 2020. "Intermittent Hypoxic Exposure with High Dose of Arginine Impact on Circulating Mediators of Tissue Regeneration" Nutrients 12, no. 7: 1933. https://doi.org/10.3390/nu12071933
APA StyleZembron-Lacny, A., Gramacki, A., Wawrzyniak-Gramacka, E., Tylutka, A., Hertmanowska, N., Kasperska, A., & Czuba, M. (2020). Intermittent Hypoxic Exposure with High Dose of Arginine Impact on Circulating Mediators of Tissue Regeneration. Nutrients, 12(7), 1933. https://doi.org/10.3390/nu12071933