Impact of Meal Timing and Chronotype on Food Reward and Appetite Control in Young Adults

 ,
,  , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Development of Diurnal Leeds Food Preference Questionnaire and Test Meal
2.2. Laboratory Study Design
2.3. Participants
2.4. Measurements
2.4.1. Initial Session
2.4.2. Habitual Energy Intake
2.4.3. Test Meal Sessions
2.5. Statistical Analyses
3. Results
3.1. Participant Characteristics
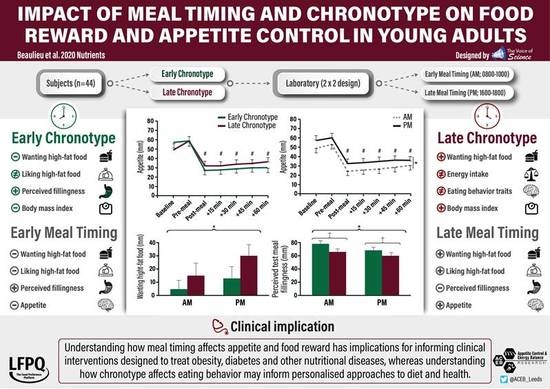
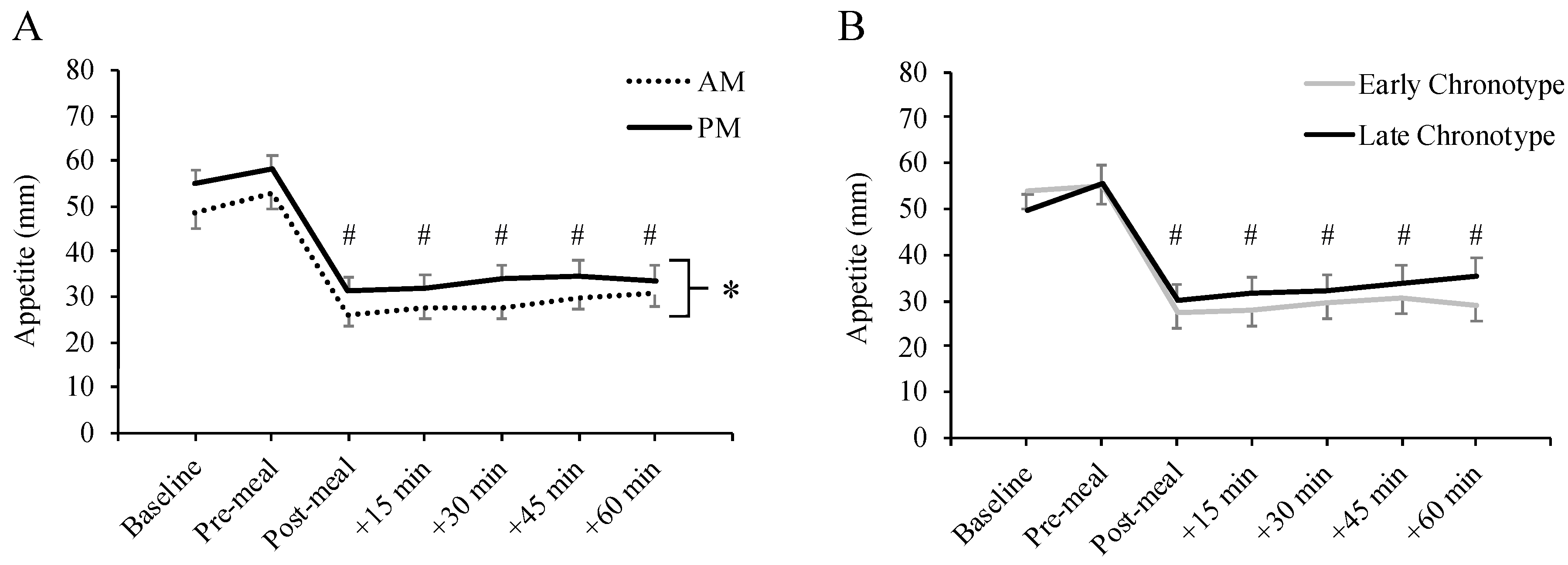
3.2. Subjective Appetite Sensations by Meal Timing and Chronotype
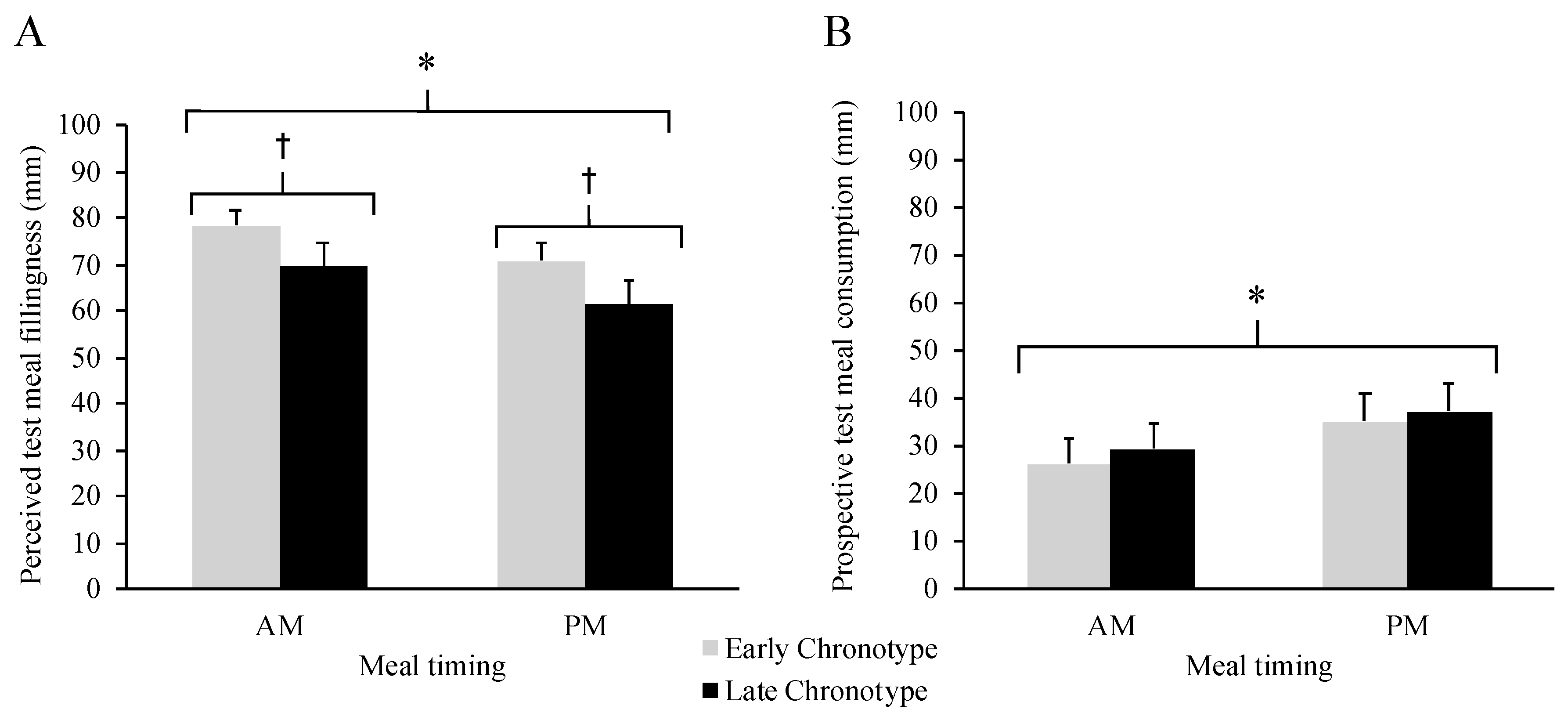
3.3. Test Meal Ratings by Meal Timing and Chronotype
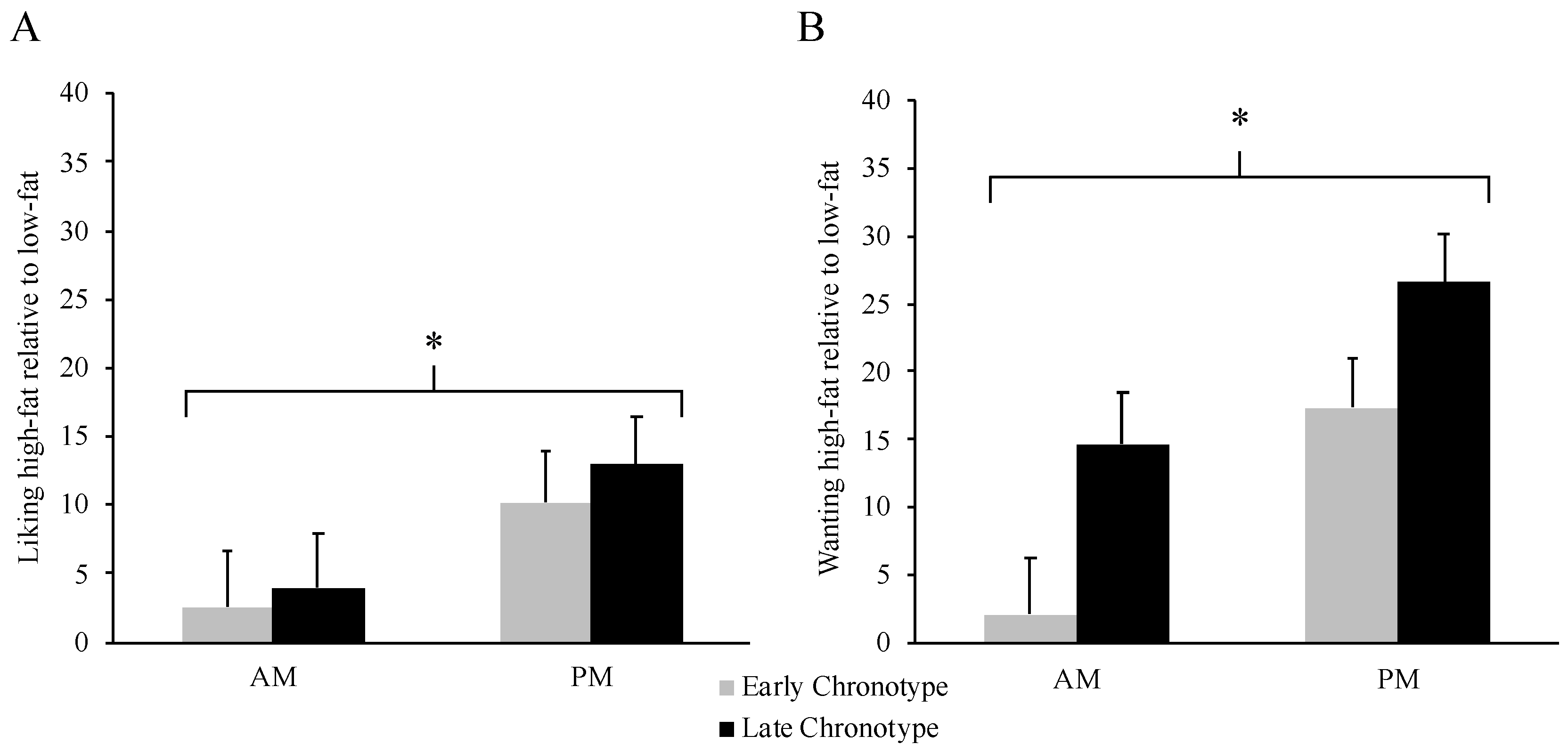
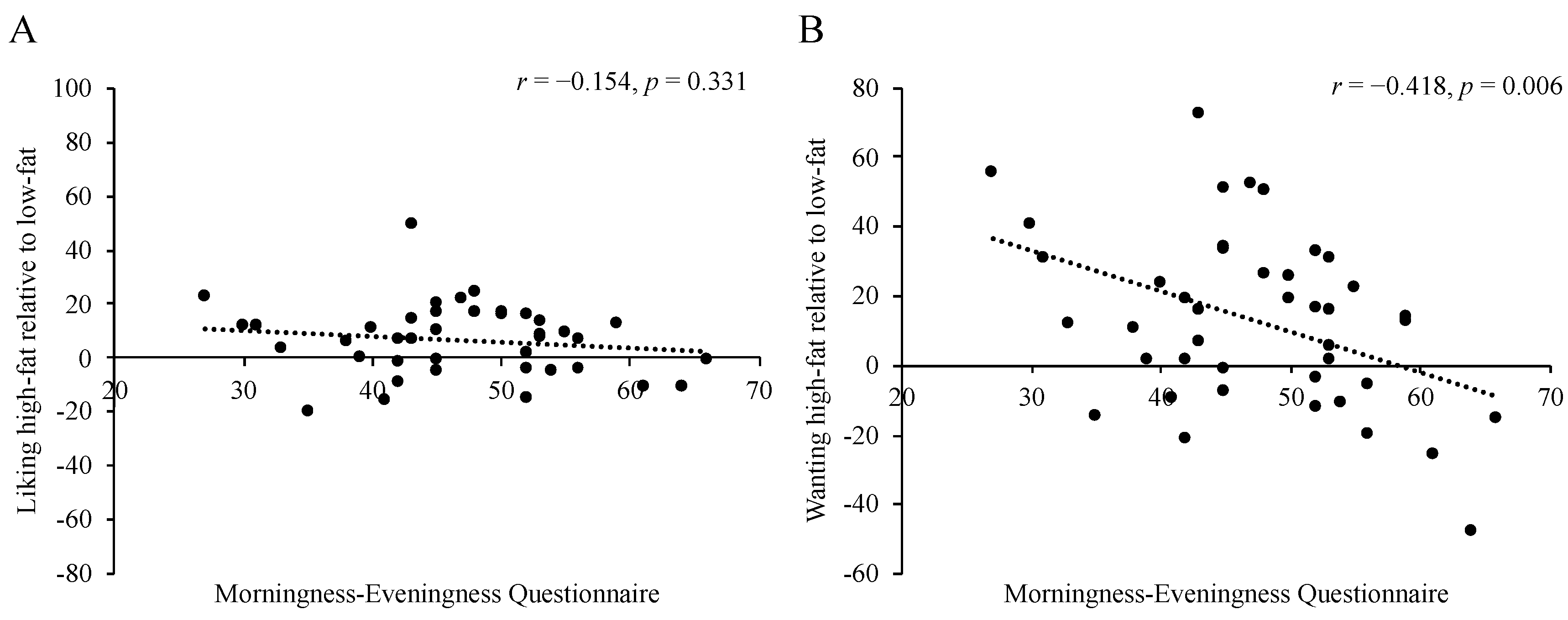
3.4. Food Reward by Meal Timing and Chronotype
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gill, S.; Panda, S. A Smartphone App Reveals Erratic Diurnal Eating Patterns in Humans that Can Be Modulated for Health Benefits. Cell Metab. 2015, 22, 789–798. [Google Scholar] [CrossRef] [PubMed]
- de Castro, J.M. Socio-cultural determinants of meal size and frequency. Br. J. Nutr. 1997, 77, S39–S54. [Google Scholar] [CrossRef] [PubMed]
- de Castro, J.M. Heritability of diurnal changes in food intake in free-living humans. Nutrition 2001, 17, 713–720. [Google Scholar] [CrossRef]
- Baron, K.G.; Reid, K.J.; Kern, A.S.; Zee, P.C. Role of sleep timing in caloric intake and BMI. Obesity 2011, 19, 1374–1381. [Google Scholar] [CrossRef]
- Wang, J.B.; Patterson, R.E.; Ang, A.; Emond, J.A.; Shetty, N.; Arab, L. Timing of energy intake during the day is associated with the risk of obesity in adults. J. Hum. Nutr. Diet. 2014, 27, 255–262. [Google Scholar] [CrossRef]
- Forslund, H.B.; Lindroos, A.K.; Sjöström, L.; Lissner, L. Meal patterns and obesity in Swedish women—A simple instrument describing usual meal types, frequency and temporal distribution. Eur. J. Clin. Nutr. 2002, 56, 740–747. [Google Scholar] [CrossRef]
- St-Onge, M.-P.; Ard, J.; Baskin, M.L.; Chiuve, S.E.; Johnson, H.M.; Kris-Etherton, P.; Varady, K. Meal Timing and Frequency: Implications for Cardiovascular Disease Prevention: A Scientific Statement from the American Heart Association. Circulation 2017, 135, 96–121. [Google Scholar] [CrossRef]
- Sofer, S.; Stark, A.H.; Madar, Z. Nutrition targeting by food timing: Time-related dietary approaches to combat obesity and metabolic syndrome. Adv. Nutr. 2015, 6, 214–223. [Google Scholar] [CrossRef]
- Garaulet, M.; Gómez-Abellán, P.; Béjar, J.J.A.; Lee, Y.-C.; Ordovas, J.M.; Scheer, F.A.J.L. Timing of food intake predicts weight loss effectiveness. Int. J. Obes. 2013, 37, 604–611. [Google Scholar] [CrossRef]
- Jakubowicz, D.; Barnea, M.; Wainstein, J.; Froy, O. High Caloric intake at breakfast vs. dinner differentially influences weight loss of overweight and obese women. Obesity 2013, 21, 2504–2512. [Google Scholar] [CrossRef]
- Johnston, J.D. Physiological responses to food intake throughout the day. Nutr. Res. Rev. 2014, 27, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Ruddick-Collins, L.C.; Johnston, J.D.; Morgan, P.J.; Johnstone, A.M. The Big Breakfast Study: Chrono-nutrition influence on energy expenditure and bodyweight. Nutr. Bull. 2018, 43, 174–183. [Google Scholar] [CrossRef] [PubMed]
- De Castro, J.M. The Time of Day of Food Intake Influences Overall Intake in Humans. J. Nutr. 2004, 134, 104–111. [Google Scholar] [CrossRef] [PubMed]
- De Castro, J.M. Circadian rhythms of the spontaneous meal pattern, macronutrient intake, and mood of humans. Physiol. Behav. 1987, 40, 437–446. [Google Scholar] [CrossRef]
- Horne, J.A.; Ostberg, O. A self-assessment questionnaire to determine morningness-eveningness in human circadian rhythms. Int. J. Chrono 1976, 4, 97–110. [Google Scholar]
- Xiao, Q.; Garaulet, M.; Scheer, F.A. Meal timing and obesity: Interactions with macronutrient intake and chronotype. Int. J. Obes. 2019, 43, 1701–1711. [Google Scholar] [CrossRef]
- Harb, A.; Levandovski, R.; Oliveira, C.; Caumo, W.; Allison, K.C.; Stunkard, A.; Hidalgo, M.P. Night eating patterns and chronotypes: A correlation with binge eating behaviors. Psychiatry Res. 2012, 200, 489–493. [Google Scholar] [CrossRef]
- Maukonen, M.; Kanerva, N.; Partonen, T.; Kronholm, E.; Tapanainen, H.; Kontto, J.; Männistö, S. Chronotype differences in timing of energy and macronutrient intakes: A population-based study in adults. Obesity 2017, 25, 608–615. [Google Scholar] [CrossRef]
- Fleig, D.; Randler, C. Association between chronotype and diet in adolescents based on food logs. Eat. Behav. 2009, 10, 115–118. [Google Scholar] [CrossRef]
- Schubert, E.; Randler, C. Association between chronotype and the constructs of the Three-Factor-Eating-Questionnaire. Appetite 2008, 51, 501–505. [Google Scholar] [CrossRef]
- Finlayson, G.; King, N.; Blundell, J.E. Liking vs. wanting food: Importance for human appetite control and weight regulation. Neurosci. Biobehav. Rev. 2007, 31, 987–1002. [Google Scholar] [CrossRef] [PubMed]
- Dalton, M.; Finlayson, G. Hedonics, satiation and satiety. In Satiation, Satiety and the Control of Food Intake; Blundell, J.E., Bellisle, F., Eds.; Woodhead Publishing: Sawston, UK, 2013; pp. 221–237. [Google Scholar]
- Dalton, M.; Blundell, J.; Finlayson, G. Effect of BMI and binge eating on food reward and energy intake: Further evidence for a binge eating subtype of obesity. Obes. Facts 2013, 6, 348–359. [Google Scholar] [CrossRef]
- Finlayson, G.; King, N.; Blundell, J. The role of implicit wanting in relation to explicit liking and wanting for food: Implications for appetite control. Appetite 2008, 50, 120–127. [Google Scholar] [CrossRef]
- Stunkard, A.J.; Messick, S. The three-factor eating questionnaire to measure dietary restraint, disinhibition and hunger. J. Psychosom. Res. 1985, 29, 71–83. [Google Scholar] [CrossRef]
- Wark, P.A.; Hardie, L.J.; Frost, G.S.; Alwan, N.A.; Carter, M.; Elliott, P.; Ford, H.E.; Hancock, N.; Morris, M.A.; Mulla, U.Z.; et al. Validity of an online 24-h recall tool (myfood24) for dietary assessment in population studies: Comparison with biomarkers and standard interviews. BMC Med. 2018, 16, 136. [Google Scholar] [CrossRef]
- Food Standards Agency. McCance and Widdowson’s The Composition of Foods; Royal Society of Chemistry: London, UK, 2014. [Google Scholar]
- Carter, M.; Albar, S.; Morris, M.A.; Mulla, U.Z.; Hancock, N.; Evans, C.E.L.; Alwan, N.A.; Greenwood, D.; Hardie, L.J.; Frost, G.; et al. Development of a UK Online 24-h Dietary Assessment Tool: myfood24. Nutrients 2015, 7, 4016–4032. [Google Scholar] [CrossRef] [PubMed]
- Flint, A.; Raben, A.; Blundell, J.; Astrup, A. Reproducibility, power and validity of visual analogue scales in assessment of appetite sensations in single test meal studies. Int. J. Obes. 2000, 24, 38–48. [Google Scholar] [CrossRef]
- Stubbs, R.J.; Hughes, D.A.; Johnstone, A.M.; Rowley, E.; Reid, C.; Elia, M.; Stratton, R.; Delargy, H.; King, N.; Blundell, J.E. The use of visual analogue scales to assess motivation to eat in human subjects: A review of their reliability and validity with an evaluation of new hand-held computerized systems for temporal tracking of appetite ratings. Br. J. Nutr. 2000, 84, 405–415. [Google Scholar] [CrossRef]
- Verschoor, E.; Finlayson, G.; Blundell, J.; Markus, C.R.; King, N.A. Effects of an acute α-lactalbumin manipulation on mood and food hedonics in high- and low-trait anxiety individuals. Br. J. Nutr. 2010, 104, 595–602. [Google Scholar] [CrossRef]
- Griffioen-Roose, S.; Finlayson, G.; Mars, M.; Blundell, J.E.; De Graaf, K. Measuring food reward and the transfer effect of sensory specific satiety. Appetite 2010, 55, 648–655. [Google Scholar] [CrossRef]
- Finlayson, G.; Arlotti, A.; Dalton, M.; King, N.; Blundell, J.E. Implicit wanting and explicit liking are markers for trait binge eating. A susceptible phenotype for overeating. Appetite 2011, 57, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Oustric, P.; Thivel, D.; Dalton, M.; Beaulieu, K.; Gibbons, C.; Hopkins, M.; Blundell, J.; Finlayson, G. Measuring food preference and reward: Application and cross-cultural adaptation of the Leeds Food Preference Questionnaire in human experimental research. Food Qual. Prefer. 2020, 80, 103824. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Erlbaum: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Levine, T.R.; Hullett, C.R. Eta Squared, Partial Eta Squared, and Misreporting of Effect Size in Communication Research. Hum. Commun. Res. 2002, 28, 612–625. [Google Scholar] [CrossRef]
- Almiron-Roig, E.; Green, H.; Virgili, R.; Aeschlimann, J.-A.; Moser, M.; Erkner, A. Validation of a new hand-held electronic appetite rating system against the pen and paper method. Appetite 2009, 53, 465–468. [Google Scholar] [CrossRef]
- Scheer, F.A.J.L.; Morris, C.J.; Shea, S.A. The internal circadian clock increases hunger and appetite in the evening independent of food intake and other behaviors. Obesity 2013, 21, 421–423. [Google Scholar] [CrossRef]
- Zhou, X.; Matthews, R.; Darwent, D.; Roach, G.D. Daily Rhythms of Hunger and Satiety in Healthy Men during One Week of Sleep Restriction and Circadian Misalignment. Int. J. Environ. Res. Public Health 2016, 13, 170. [Google Scholar] [CrossRef]
- Qian, J.; Morris, C.J.; Caputo, R.; Garaulet, M.; Scheer, F.A.J.L. Ghrelin is impacted by the endogenous circadian system and by circadian misalignment in humans. Int. J. Obes. 2019, 43, 1644–1649. [Google Scholar] [CrossRef]
- de Castro, J.M. 13-Satiation, satiety and the daily intake pattern. In Satiation, Satiety and the Control of Food Intake; Blundell, J.E., Bellisle, F., Eds.; Woodhead Publishing: Sawston, UK, 2013; pp. 238–252. [Google Scholar]
- Itzhacki, J.; Lindert, B.H.W.T.; Van Der Meijden, W.P.; Kringelbach, M.L.; Mendoza, J.; Van Someren, E.J. Environmental light and time of day modulate subjective liking and wanting. Emotion 2019, 19, 10–20. [Google Scholar] [CrossRef]
- Byrne, J.E.; Murray, G. Diurnal rhythms in psychological reward functioning in healthy young men: ‘Wanting’, liking, and learning. Chrono. Int. 2017, 34, 287–295. [Google Scholar] [CrossRef]
- Hasler, B.P.; Forbes, E.E.; Franzen, P.L. Time-of-day differences and short-term stability of the neural response to monetary reward: A pilot study. Psychiatry Res. 2014, 224, 22–27. [Google Scholar] [CrossRef]
- Murray, G.; Nicholas, C.L.; Kleiman, J.; Dwyer, R.; Carrington, M.J.; Allen, N.B.; Trinder, J. Nature’s clocks and human mood: The circadian system modulates reward motivation. Emotion 2009, 9, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, J.D.; Patrician, T.M.; Breslin, F.J.; Martin, L.E.; Donnelly, J.E.; Savage, C.R. Evening hyperphagia and food motivation: A preliminary study of neural mechanisms. Eat. Behav. 2013, 14, 447–450. [Google Scholar] [CrossRef] [PubMed][Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food | AM Appropr. Mean ± SD | PM Appropr. Mean ± SD | E% Protein | E% CHO | E% Fat | Energy Density (kcal/100 g) | |
|---|---|---|---|---|---|---|---|
| High-Fat Savory | Cashews | 46 ± 32 | 75 ± 18 | 13.7 | 13.8 | 70.2 | 642 |
| Croissant, ham & cheese | 79 ± 24 | 51 ± 23 | 17.3 | 25.3 | 54.9 | 345 | |
| Sausage roll | 51 ± 33 | 67 ± 20 | 10.4 | 22.3 | 64.9 | 361 | |
| Sausage sandwich | 80 ± 24 | 62 ± 23 | 24.4 | 22.8 | 50.1 | 279 | |
| Mean | 65 | 62 | 16.5 | 21.1 | 60.0 | 407 | |
| Low-Fat Savory | Beans on toast | 81 ± 22 | 74 ± 21 | 19.1 | 66.5 | 4.6 | 122 |
| Bread roll | 76 ± 26 | 80 ± 18 | 13.2 | 69.8 | 9.8 | 258 | |
| Rice cake | 57 ± 32 | 67 ± 23 | 8.5 | 77.0 | 6.5 | 386 | |
| Rye crackers (plain) | 61 ± 34 | 67 ± 23 | 9.7 | 71.7 | 4.4 | 350 | |
| Mean | 68 | 71 | 12.6 | 71.2 | 6.3 | 279 | |
| High-Fat Sweet | Blueberry muffin | 59 ± 31 | 64 ± 22 | 5.5 | 44.1 | 46.4 | 367 |
| Cinnamon swirl pastry | 69 ± 27 | 53 ±25 | 4.9 | 37.7 | 53.8 | 445 | |
| Crepe with cream | 76 ± 28 | 59 ± 24 | 4.6 | 16.1 | 79.3 | 263 | |
| Flapjack | 57 ± 29 | 69 ± 21 | 4.9 | 48.3 | 41.4 | 435 | |
| Mean | 65 | 60 | 5.0 | 36.6 | 55.2 | 377 | |
| Low-Fat Sweet | Dried apricots | 75 ± 27 | 73 ± 21 | 8.8 | 75.8 | 3.0 | 178 |
| Red grapes | 82 ± 24 | 86 ± 17 | 2.4 | 87.5 | 1.4 | 66 | |
| Banana | 91 ± 13 | 82 ± 20 | 4.7 | 83.7 | 4.4 | 103 | |
| Light chocolate granola bar | 83 ± 19 | 60 ± 22 | 5.8 | 61.1 | 16.2 | 344 | |
| Mean | 83 | 75 | 5.4 | 77.0 | 6.3 | 173 |
| All (N = 44) | Early Chronotype (N = 22) | Late Chronotype (N = 22) | |
|---|---|---|---|
| Weight (kg) | 72.9 ± 11.4 | 73.4 ± 10.3 | 72.4 ± 12.7 |
| BMI (kg/m2) | 24.5 ± 3.2 | 24.1 ± 2.7 | 24.9 ± 3.6 |
| Body fat (%) | 27.7 ± 8.3 | 27.3 ± 8.4 | 28.2 ± 8.4 |
| Hip circumference (cm) | 98.4 ± 6.9 | 99.2 ± 4.8 | 97.6 ± 8.6 |
| Waist circumference (cm) | 84.3 ± 7.9 | 84.2 ± 6.2 | 84.3 ± 9.4 |
| Waist-to-hip ratio | 0.86 ± 0.06 | 0.85 ± 0.07 | 0.86 ± 0.06 |
| MEQ score | 48 ± 9 | 55 ± 5 | 40 ± 6 * |
| TFEQ Restraint | 7 ± 4 | 6 ± 3 | 8 ± 5 |
| TFEQ Disinhibition | 8 ± 3 | 7 ± 3 | 8 ± 3 |
| TFEQ Hunger | 7 ± 3 | 7 ± 3 | 7 ± 3 |
| Energy intake (kcal/day) 1 | 1791 ± 663 | 1843 ± 681 | 1737 ± 659 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beaulieu, K.; Oustric, P.; Alkahtani, S.; Alhussain, M.; Pedersen, H.; Quist, J.S.; Færch, K.; Finlayson, G. Impact of Meal Timing and Chronotype on Food Reward and Appetite Control in Young Adults. Nutrients 2020, 12, 1506. https://doi.org/10.3390/nu12051506
Beaulieu K, Oustric P, Alkahtani S, Alhussain M, Pedersen H, Quist JS, Færch K, Finlayson G. Impact of Meal Timing and Chronotype on Food Reward and Appetite Control in Young Adults. Nutrients. 2020; 12(5):1506. https://doi.org/10.3390/nu12051506
Chicago/Turabian StyleBeaulieu, Kristine, Pauline Oustric, Shaea Alkahtani, Maha Alhussain, Hanne Pedersen, Jonas Salling Quist, Kristine Færch, and Graham Finlayson. 2020. "Impact of Meal Timing and Chronotype on Food Reward and Appetite Control in Young Adults" Nutrients 12, no. 5: 1506. https://doi.org/10.3390/nu12051506
APA StyleBeaulieu, K., Oustric, P., Alkahtani, S., Alhussain, M., Pedersen, H., Quist, J. S., Færch, K., & Finlayson, G. (2020). Impact of Meal Timing and Chronotype on Food Reward and Appetite Control in Young Adults. Nutrients, 12(5), 1506. https://doi.org/10.3390/nu12051506