Two Faces of Vitamin C—Antioxidative and Pro-Oxidative Agent

 , , and
, , and 
Abstract
1. Introduction
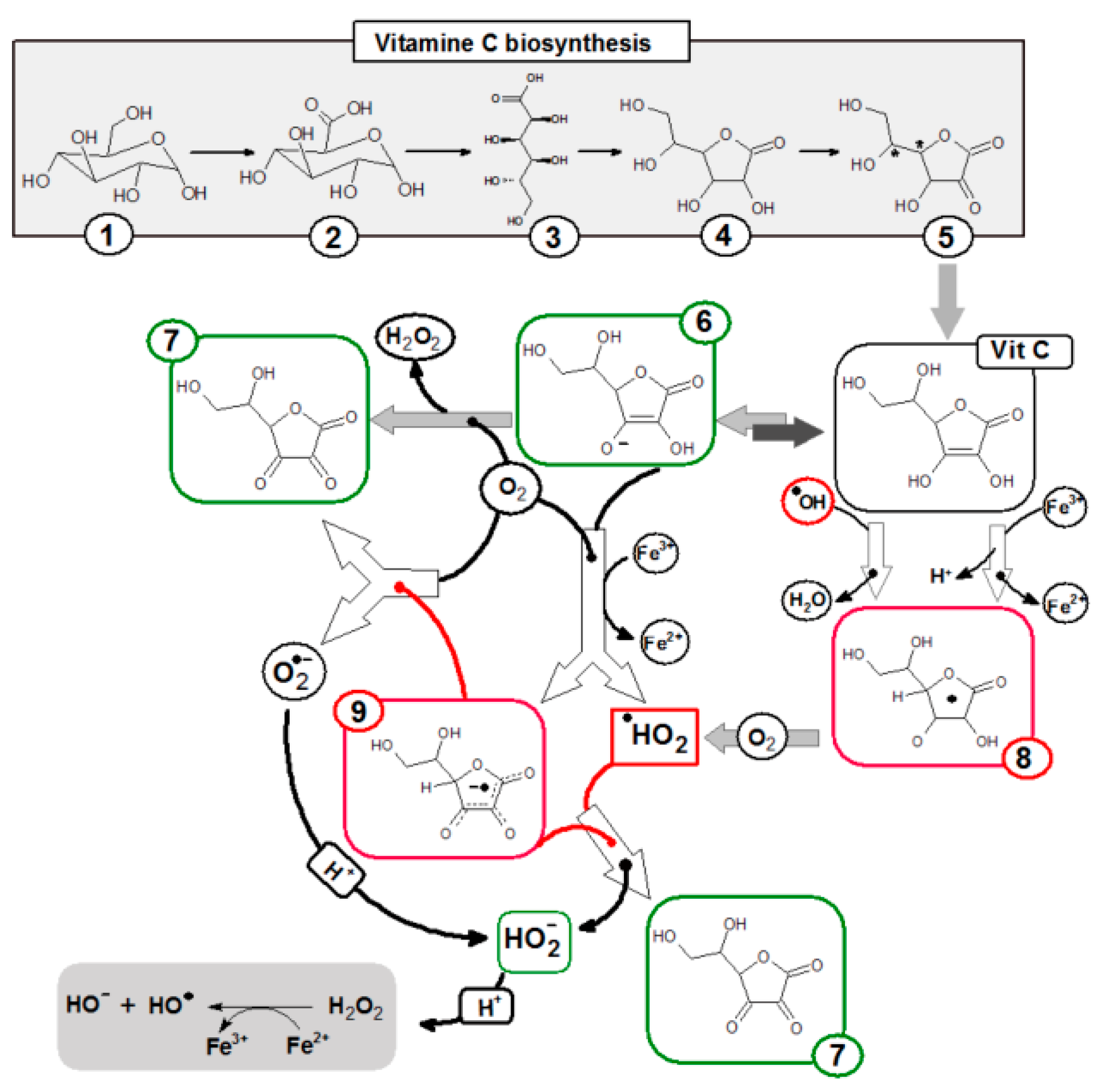
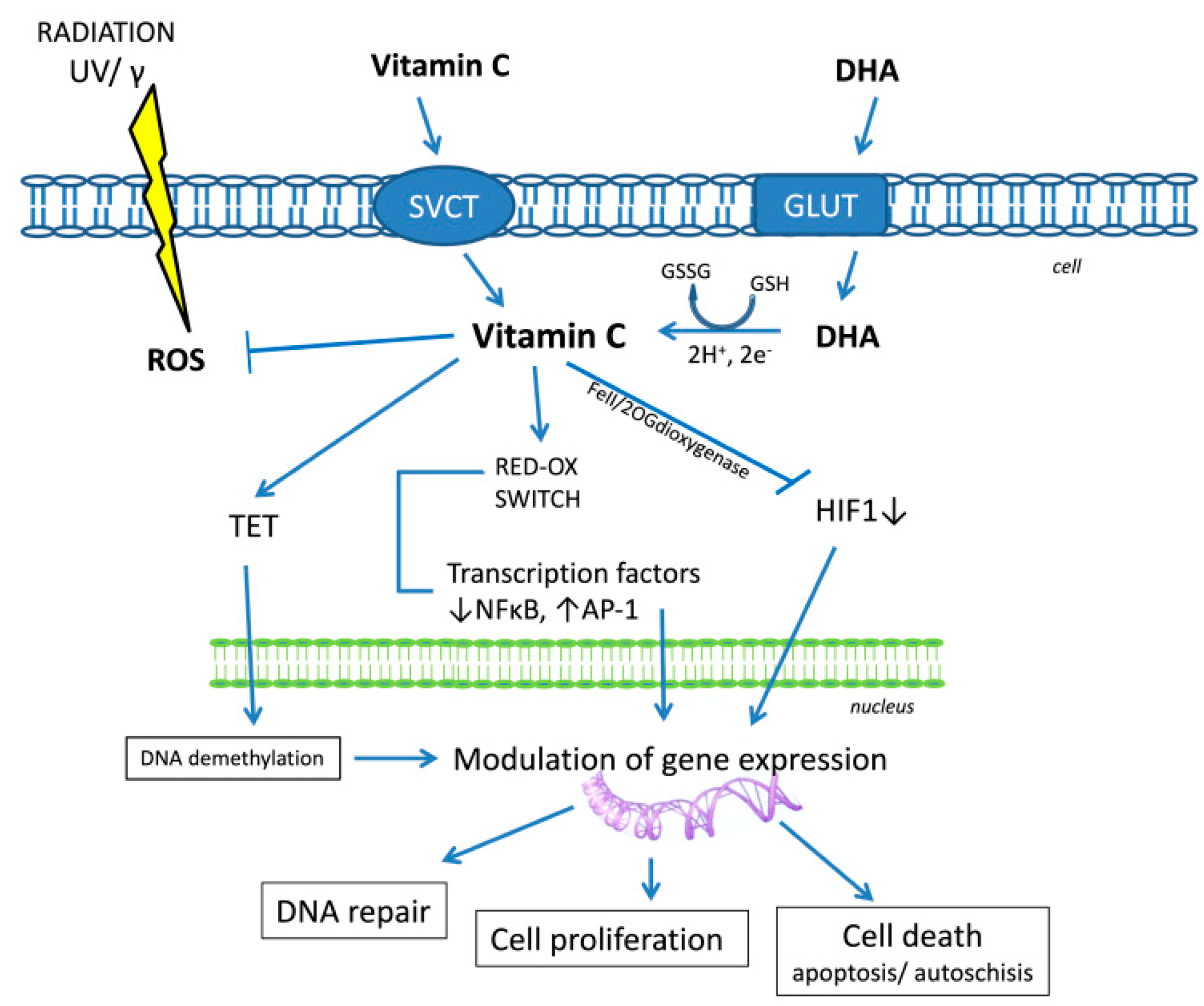
2. Vitamin C: Significant Element of Cellular Metabolism
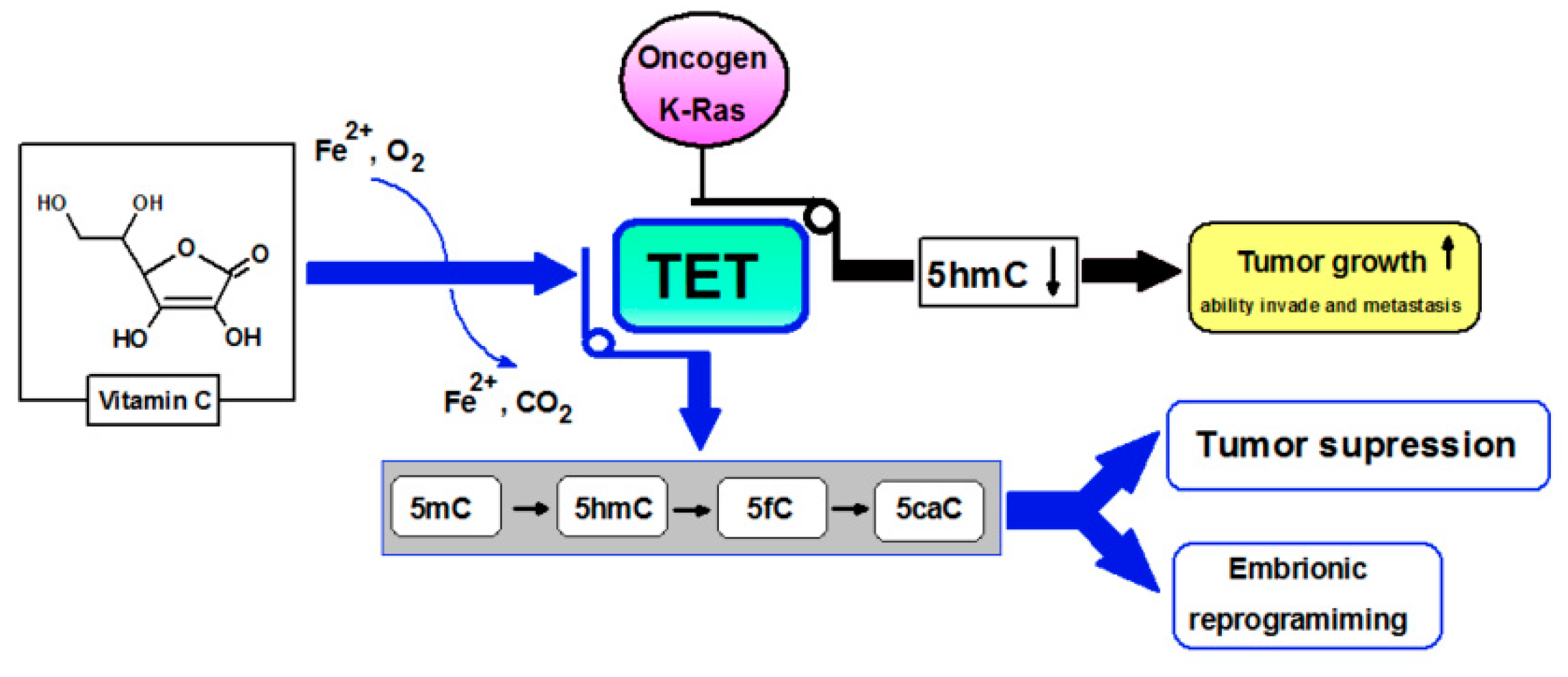
3. Vitamin C: Anti-Cancer Potential
4. Vitamin C: Main Antioxidant of the CNS
5. Vitamin C and Oxidative DNA Damage
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baron, J.H. Sailors scurvy before and after James Lind—A reassessment. Nutr. Rev. 2009, 67, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Varvara, M.; Bozzo, G.; Disanto, C.; Pagliarone, C.N.; Celano, G.V. The use of the ascorbic acid as food additive and technical-legal issues. Ital. J. Food Saf. 2016, 5, 4313. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M.; Pejchal, J.; Snopkova, S.; Havlickova, K.; Karasova, J.Z.; Bostik, P.; Pikula, J. Ascorbic Acid: An Old Player with a Broad Impact on Body Physiology Including Oxidative Stress Suppression and Immunomodulation: A Review. Mini Rev. Med. Chem. 2012, 12, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Jarosz, M. Normy Żywienia dla Populacji Polski; Instytut Żywności i Żywienia: Warszawa, Poland, 2017; Volume 147. [Google Scholar]
- Korcok, J.; Dixon, S.J.; Lo, T.C.Y.; Wilson, J.X. Differential effects of glucose on dehydroascorbic acid transport and intracellular ascorbate accumulation in astrocytes and skeletal myocytes. Brain Res. 2003, 993, 201–207. [Google Scholar] [CrossRef]
- Levine, M.; Wang, Y.; Padayatty, S.J.; Morrow, J. A new recommended dietary allowance of vitamin C for healthy young women. Proc. Natl. Acad. Sci. USA 2001, 98, 9842–9846. [Google Scholar] [CrossRef]
- Harrison, F.E.; May, J.M. Vitamin C function in the brain, vital role of the ascorbate transporter SVCT2. Free Radic. Biol. Med. 2009, 46, 719–730. [Google Scholar] [CrossRef]
- Kalemba-Drożdż, M. Vitamin C for the treatment of cancer and the prevention of non-infectious diseases. Państwo i Społeczeństwo 2018, 3, 91–104. [Google Scholar] [CrossRef]
- Cahill, L.E.; El-Sohemy, A. Vitamin C Transporter Gene Polymorphisms, Dietary Vitamin C and Serum Ascorbic Acid. Lifestyle Genom.J. Nutrigenet. Nutrigenomics 2009, 2, 292–301. [Google Scholar] [CrossRef]
- Castro, M.; Caprile, T.; Astuya, A.; Millán, C.; Reinicke, K.; Vera, J.C. High-affinity sodium-vitamin C co-transporters (SVCT) expression in embryonic mouse neurons. J. Neurochem 2001, 78, 815–823. [Google Scholar] [CrossRef]
- Duell, E.J.; Lujan-Barroso, L.; Llivina, C. Vitamin C transporter gene (SLC23A1 and SLC23A2) polymorphisms, plasma vitamin C levels, and gastric cancer risk in the EPIC cohort. Genes Nutr. 2013, 8, 549–560. [Google Scholar] [CrossRef]
- Granger, M.; Eck, P. Dietary Vitamin C in Human Health. Adv. Food Nutr. Res. 2018, 83, 281–310. [Google Scholar]
- Corti, A.; Raggi, C.; Franzini, M.; Paolicchi, A.; Pompella, A.; Casini, A.F. Plasma membrane γ glutamyltransferase activity facilitates the uptake of vitamin C in melanoma cells. Free Radic. Biol. Med. 2004, 37, 1906–1915. [Google Scholar] [CrossRef] [PubMed]
- Padayatty, S.J.; Levine, M. Vitamin C: The known and the unknown and Goldilocks. Oral Dis. 2016, 22, 463–493. [Google Scholar] [CrossRef] [PubMed]
- Konopacka, M. Rola witaminy C w uszkodzeniach oksydacyjnych DNA. Postepy Hig. Med. Dosw. 2004, 58, 343–348. [Google Scholar]
- Bartosz, G. Druga Twarz Tlenu: Wolne Rodniki w Przyrodzie; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2009. [Google Scholar]
- Englard, S.; Seifter, S. The biochemical functions of Ascorbic acid. Ann. Rev. Nutr. 1986, 6, 365–406. [Google Scholar] [CrossRef]
- Montecinos, V.; Guzmán, P.; Barra, V.; Villagrán, M.; Muñoz-Montesino, C.; Sotomayor, K.; Vera, J.C. Vitamin C Is an Essential Antioxidant That Enhances Survival of Oxidatively Stressed Human Vascular Endothelial Cells in the Presence of a Vast Molar Excess of Glutathione. J. Biol. Chem. 2007, 282, 15506–15515. [Google Scholar] [CrossRef] [PubMed]
- Janda, K.; Kasprzak, M.; Wolska, J. Vitamin C—Structure, properties, occurrence and functions. Pomeranian J. Life Sci. 2015, 61, 419–425. [Google Scholar] [PubMed]
- Kohgo, Y.; Ikuta, K.; Ohtake, T.; Torimoto, Y.; Kato, J. Body iron metabolism and pathophysiology of iron overload. Int. J. Hematol. 2008, 88, 7–15. [Google Scholar] [CrossRef]
- Wohlrab, C.; Vissers, M.C.M.; Phillips, E.; Morrin, H.; Robinson, B.A.; Dachs, G.U. The Association Between Ascorbate and the Hypoxia-Inducible Factors in Human Renal Cell Carcinoma Requires a Functional Von Hippel-Lindau Protein. Front. Oncol. 2018, 8, 574. [Google Scholar] [CrossRef]
- Eales, K.L.; Hollinshead, K.E.R.; Tennant, D.A. Hypoxia and metabolic adaptation of cancer cells. Oncogenesis 2016, 5, e190. [Google Scholar] [CrossRef]
- Guz, J.; Oliński, R. The role of vitamin C in epigenetic regulation. Postepy Hig. Med. Dosw. 2017, 71, 747–760. [Google Scholar] [CrossRef] [PubMed]
- Lian, H.; Li, W.B.; Jin, W.L. The emerging insights into catalytic or non-catalytic roles of TET proteins in tumors and neural development. Oncotarget 2016, 7, 64512–64525. [Google Scholar] [CrossRef] [PubMed]
- DiTroia, S.P.; Percharde, M.; Guerquin, M.-J.; Wall, E.; Collignon, E.; Ebata, K.T.; Mesh, K.; Mahesula, S.; Agathocleous, M.; Laird, D.J.; et al. Maternal vitamin C regulates reprogramming of DNA methylation and germline development. Nature 2019, 573, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Mao, S.Q.; Zhao, B.; Chong, Z.; Yang, Y.; Zhao, C.; Wang, H. Ascorbic Acid Enhances Tet-Mediated 5-Methylcytosine Oxidation and Promotes DNA Demethylation in Mammals. J. Am. Chem. Soc. 2013, 135, 10396–10403. [Google Scholar] [CrossRef]
- Fu, H.L.; Ma, Y.; Lu, L.G.; Hou, P.; Li, B.J.; Jin, W.L.; Cui, D.X. TET1 exertsits tumor suppressor function by interacting with p53-EZH2 pathwayin gastric cancer. J. Biomed. Nanotechnol. 2014, 10, 1217–1230. [Google Scholar] [CrossRef]
- Ciesielski, P.; Jóźwiak, P.; Krześlak, A. Białka TET a modyfikacje epigenetyczne w nowotworach. Postepy Hig. Med. Dosw. 2015, 69, 1371–1383. [Google Scholar] [CrossRef]
- Wu, B.K.; Brenner, C. Suppression of TET1-dependent DNAdemethylation is essential for KRAS-mediated transformation. Cell Rep. 2014, 9, 1827–1840. [Google Scholar] [CrossRef]
- Jin, S.-G.; Jiang, Y.; Qiu, R.; Rauch, T.A.; Wang, Y.; Schackert, G. 5-Hydroxymethylcytosine Is Strongly Depleted in Human Cancers but Its Levels Do Not Correlate with IDH1 Mutations. Cancer Res. 2011, 71, 7360–7365. [Google Scholar] [CrossRef]
- Rampal, R.; Alkalin, A.; Madzo, J.; Vasanthakumar, A.; Pronier, E.; Patel, J. DNA Hydroxymethylation Profiling Reveals that WT1 Mutations Result in Loss of TET2 Function in Acute Myeloid Leukemia. Cell Rep. 2014, 9, 1841–1855. [Google Scholar] [CrossRef]
- Pfeifer, G.P.; Kadam, S.; Jin, S.-G. 5-hydroxymethylcytosine and its potential roles in development and cancer. Epigenetics Chromatin 2013, 6, 10. [Google Scholar] [CrossRef]
- Simon, J.A.; Hudes, E.S.; Tice, J.A. Relation of serum ascorbic acid to mortality among US adults. J. Am. Coll. Nutr. 2001, 20, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Langlois, M.; Duprez, D.; Delanghe, J.; De Buyzere, M.; Clement, D.L. Serum vitamin C concentration is low in peripheral arterial disease and is associated with inflammation and severity of atherosclerosis. Circulation 2001, 103, 1863–1868. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, T.; Date, C.; Kokubo, Y.; Yoshiike, N.; Matsumura, Y.; Tanaka, H. Serum Vitamin C Concentration Was Inversely Associated with Subsequent 20-Year Incidence of Stroke in a Japanese Rural Community: The Shibata Study. Stroke 2000, 31, 2287–2294. [Google Scholar] [CrossRef]
- Lin, J.; Cook, N.R.; Albert, C.; Zaharris, E.; Gaziano, J.M.; Van Denburgh, M. Vitamins C and E and Beta Carotene Supplementation and Cancer Risk: A Randomized Controlled Trial. J Natl. Cancer. Inst. 2008, 101, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Espey, M.G.; Krishna, M.C.; Mitchell, J.B.; Corpe, C.P.; Buettner, G.R. Pharmacologic ascorbic acid concentrations selectively kill cancer cells: Action as a pro-drug to deliver hydrogen peroxide to tissues. Proc. Natl. Acad. Sci. USA 2005, 102, 13604–13609. [Google Scholar] [CrossRef]
- Schoenfeld, J.D.; Sibenaller, Z.A.; Mapuskar, K.A.; Wagner, B.A.; Cramer-Morales, K.L.; Furqan, M. O 2- and H 2 O 2 -Mediated Disruption of Fe Metabolism Causes the Differential Susceptibility of NSCLC and GBM Cancer Cells to Pharmacological Ascorbate. Cancer Cell 2017, 31, 487–500.e8. [Google Scholar] [CrossRef]
- Olney, K.E.; Du, J.; van’t Erve, T.J.; Witmer, J.R.; Sibenaller, Z.A.; Wagner, B.A. Inhibitors of hydroperoxide metabolism enhance ascorbate-induced cytotoxicity. Free Radic. Res. 2013, 47, 154–163. [Google Scholar] [CrossRef]
- Ibrahim, W.H.; Habib, H.M.; Kamal, H.; St Clair, D.K.; and Chow, C.K. Mitochondrial superoxide mediates labile iron level: Evidence from Mn-SODtransgenic mice and heterozygous knockout mice and isolated rat liver mitochondria. Free Radic. Biol. Med. 2013, 65, 143–149. [Google Scholar] [CrossRef]
- Torti, S.V.; Torti, F.M. Iron and cancer: More ore to be mined. Nat. Rev. Cancer 2013, 13, 342–355. [Google Scholar] [CrossRef]
- Uetaki, M.; Tabata, S.; Nakasuka, F.; Soga, T.; Tomita, M. Metabolomic alterations in human cancer cells by vitamin C-induced oxidative stress. Sci. Rep. 2015, 5, 13896. [Google Scholar] [CrossRef]
- Chen, P.; Yu, J.; Chalmers, B.; Drisko, J.; Yang, J.; Li, B.; Chen, Q. Pharmacological ascorbate induces cytotoxicity in prostate cancer cells through ATP depletion and induction of autophagy. Anti-Cancer Drugs 2012, 23, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-W.; Jin, N.-H.; Hahm, E.-S.; Yim, S.-H.; Lim, J.S.; Kim, K.-I.; Yang, Y.; Lee, S.-S.; Kang, J.-S.; Lee, W.-J.; et al. Ascorbate (vitamin C) induces cell death through the apoptosis-inducing factor in human breast cancer cells. Oncol. Rep. 2007, 18, 811–815. [Google Scholar] [CrossRef] [PubMed]
- Mustafi, S.; Sant, D.W.; Liu, Z.-J.; Wang, G. Ascorbate induces apoptosis in melanoma cells by suppressing Clusterin expression. Sci. Rep. 2017, 7, 3671. [Google Scholar] [CrossRef] [PubMed]
- Jamison, J.M.; Gilloteaux, J.; Taper, H.S.; Calderon, P.B. Summers J.L. Autoschizis: A novel cell death. Biochem. Pharmacol. 2002, 63, 1773–1783. [Google Scholar] [CrossRef]
- Gilloteaux, J.; Jamison, J.M.; Arnold, D.; Taper, H.S.; Summers, J.L. Ultrastructural Aspects of Autoschizis: A New Cancer Cell Death Induced by the Synergistic Action of Ascorbate/Menadione on Human Bladder Carcinoma Cells. Ultrastruct. Pathol. 2001, 25, 183–192. [Google Scholar] [CrossRef]
- Connors, T.A.; Jones, M.; Ross, W.C.J.; Braddock, P.D.; Khokhar, A.R.; Tobe, M.L. New platinum complexes with anti tumour activity. Chem.-Biol. Interact. 1972, 5, 415–424. [Google Scholar] [CrossRef]
- Shahid, F.; Farooqui, Z.; Khan, F. Cisplatin-induced gastrointestinal toxicity: An update on possible mechanisms and on available gastroprotective strategies. Eur. J. Pharmacol. 2018, 827, 49–57. [Google Scholar] [CrossRef]
- Leekha, A.; Gurjar, B.S.; Tyagi, A.; Rizvi, M.A.; Verma, A.K. Vitamin C in synergism with cisplatin induces cell death in cervical cancer cells through altered redox cycling and p53 upregulation. J. Cancer Res. Clin. Oncol. 2016, 142, 2503–2514. [Google Scholar] [CrossRef]
- Asaduzzaman Khan, M.; Tania, M.; Zhang, D.; Chen, H. Antioxidant Enzymes and Cancer. Chin. J. Cancer Res. 2010, 22, 87–92. [Google Scholar] [CrossRef]
- Kim, J.; Lee, S.-D.; Chang, B.; Jin, N.-H.; Jung, S.-I.; Park, M.-Y.; Han, Y.; Yang, Y.; Kim, K.I.; Lim, J.S.; et al. Enhanced antitumor activity of vitamin C via p53 in Cancer cells. Free Radic. Biol. Med. 2012, 53, 1607–1615. [Google Scholar] [CrossRef]
- Yun, J.; Mullarky, E.; Lu, C.; Bosch, K.N.; Kavalier, A.; Rivera, K.; Roper, J.; Chio, I.I.C.; Giannopoulou, E.G.; Rago, C.; et al. Vitamin C selectively kills KRAS and BRAF mutant colorectal cancer cells by targeting GAPDH. Science 2015, 350, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Blaszczak, W.; Barczak, W.; Masternak, J.; Kopczyński, P.; Zhitkovich, A.; Rubiś, B. Vitamin C as a Modulator of the Response to Cancer Therapy. Molecules 2019, 24, 453. [Google Scholar] [CrossRef] [PubMed]
- Verrax, J.; Calderon, P. Pharmacologic Concentrations of Ascorbate Are Achieved by Parenteral Administration and Exhibit Antitumoral Effects. Free Radic. Biol. Med. 2009, 47, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Doskey, C.M.; Buranasudja, V.; Wagner, B.A.; Wilkes, J.G.; Du, J.; Cullen, J.J.; Buettner, G.R. Tumor cells have decreased ability to metabolize H2O2: Implications for pharmacological ascorbate in cancer therapy. Redox Biol. 2016, 10, 274–284. [Google Scholar] [CrossRef]
- Chen, Q.; Espey, M.G.; Sun, A.; Pooput, C.; Kirk, K.L.; Krishna, M.C.; Khosh, D.B.; Drisko, J.; Levine, M. Pharmacologic doses of ascorbate act as a prooxidant and decrease growth of aggressive tumor xenografts in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 11105–11109. [Google Scholar] [CrossRef]
- Du, J.; Martin, S.M.; Levine, M.; Wagner, B.A.; Buettner, G.R.; Wang, S.-H.; Taghiyev, A.F.; Du, C.; Knudson, C.M.; Cullen, J.J. Mechanisms of Ascorbate-Induced Cytotoxicity in Pancreatic Cancer. Clin. Cancer Res. 2010, 16, 509–520. [Google Scholar] [CrossRef]
- Roomi, M.W.; Kalinovsky, T.; Cha, J.; Roomi, N.W.; Niedzwiecki, A.; Rath, M. Effects of a nutrient mixture on immunohistochemical localization of cancer markers in human cervical cancer HeLa cell tumor xenografts in female nude mice. Exp. Ther. Med. 2014, 9, 294–302. [Google Scholar] [CrossRef]
- Vollbracht, C.; Schneider, B.; Leendert, V.; Weiss, G.; Auerbach, L.; Beuth, J. Intravenous Vitamin C Administration Improves Quality of Life in Breast Cancer Patients during Chemo/Radiotherapy and Aftercare: Results of a Retrospective, Multicentre, Epidemiological Cohort Study in Germany. In Vivo 2011, 25, 983–990. [Google Scholar]
- Ma, Y.; Chapman, J.; Levine, M.; Polireddy, K.; Drisko, J.; Chen, Q. High-Dose Parenteral Ascorbate Enhanced Chemosensitivity of Ovarian Cancer and Reduced Toxicity of Chemotherapy. Sci. Transl. Med. 2014, 6, 222ra18. [Google Scholar] [CrossRef]
- Harris, H.R.; Orsini, N.; Wolk, A. Vitamin C and survival among women with breast cancer: A Meta-analysis. Eur. J. Cancer 2014, 50, 1223–1231. [Google Scholar] [CrossRef]
- Bjelakovic, G.; Nikolova, D.; Simonetti, R.G.; Gluud, C. Antioxidant Supplements for Preventing Gastrointestinal Cancers. Cochrane Database Syst. Rev. 2008, 3, CD004183. [Google Scholar] [CrossRef]
- Almendra Freitas, B.; de Lloret, G.R.; Visacri, M.B.; Tuan, B.T.; Amaral, L.S.; Baldini, D.; Moriel, P. High 15 F2t-Isoprostane Levels in Patients with a Previous History of Nonmelanoma Skin Cancer: The Effects of Supplementary Antioxidant Therapy. BioMed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Satheesh, N.; Samuel, S.; Büsselberg, D. Combination Therapy with Vitamin C Could Eradicate Cancer Stem Cells. Biomolecules 2020, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, E.M.; Bonuccelli, G.; Maggiolini, M.; Sotgia, F.; Lisanti, M.P. Vitamin C and Doxycycline: A synthetic lethal combination therapy targeting metabolic flexibility in cancer stem cells (CSCs). Oncotarget 2017, 8, 67269–67286. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Fu, L. Targeting Cancer Stem Cells: A New Therapy to Cure Cancer Patients. Am. J. Cancer Res. 2012, 2, 340–356. [Google Scholar] [PubMed]
- Popa-Wagner, A.; Mitran, S.; Sivanesan, S.; Chang, E.; Buga, A.-M. ROS and Brain Diseases: The Good, the Bad, and the Ugly. Oxidative Med. Cell. Longev. 2013, 2013, 1–14. [Google Scholar] [CrossRef]
- Rice, M.E. Ascorbate regulation and its neuroprotective role in the brain. Trends Neurosci. 2000, 23, 209–216. [Google Scholar] [CrossRef]
- Covarrubias-Pinto, A.; Acuña, A.; Beltrán, F.; Torres-Díaz, L.; Castro, M. Old Things New View: Ascorbic Acid Protects the Brain in Neurodegenerative Disorders. Int. J. Mol. Sci. 2015, 16, 28194–28217. [Google Scholar] [CrossRef]
- Majewska, M.D.; Bell, J.A. Ascorbic Acid Protects Neurons From Injury Induced by Glutamate and NMDA. Neuroreport 1990, 1, 194–196. [Google Scholar] [CrossRef]
- Du, J.; Cullen, J.J.; Buettner, G.R. Ascorbic acid: Chemistry, biology and the treatment of cancer. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2012, 1826, 443–457. [Google Scholar] [CrossRef]
- Perez Ortiz, J.M.; Swerdlow, R.H. Mitochondrial dysfunction in Alzheimer’s disease: Role in pathogenesis and novel therapeutic opportunities. Br. J. Pharmacol. 2019, 176, 3489–3507. [Google Scholar] [CrossRef] [PubMed]
- Kook, S.-Y.; Lee, K.-M.; Kim, Y.; Cha, M.-Y.; Kang, S.; Baik, S.H.; Lee, H.; Park, R.; Mook-Jung, I. High-dose of vitamin C supplementation reduces amyloid plaque burden and ameliorates pathological changes in the brain of 5XFAD mice. Cell Death Dis. 2014, 5, 1083. [Google Scholar] [CrossRef] [PubMed]
- Sagun, K.C.; Ca´rcamo, J.M.; Golde, D.W. Vitamin C enters mitochondria via facilitative glucose transporter 1 (Glut1) and confers mitochondrial protection against oxidative injury. FASEB J. 2005, 19, 1657–1667. [Google Scholar]
- May, J.M.; Qu, Z.; Nazarewicz, R.; Dikalov, S. Ascorbic acid efficiently enhances neuronal synthesis of norepinephrine from dopamine. Brain Res. Bull. 2013, 90, 35–42. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Trueba, G.P.; Poulsen, H.E.; Christen, S. Vitamin C deficiency in weanling guinea pigs: Differential expression of oxidative stress and DNA repair in liver and brain. Br. J. Nutr. 2007, 98, 1116–1119. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Galiniak, S.; Bartosz, G. Reakcja Fentona. Kosmos Problemy Nauk Biologicznych 2014, 63, 309–314. [Google Scholar]
- Haber, F.; Weiss, J. On the catalysis of hydroperoxide. Naturwissenschaften 1932, 20, 948–950. [Google Scholar] [CrossRef]
- Koppenol, W.H. The Haber-Weiss cycle—70 years later. Redox Rep. 2001, 6, 229–234. [Google Scholar] [CrossRef]
- Turowski, P.; Uhrynowska, M.; Brojer, E. Thalassemias—Pathophysiology, molecular basics and diagnostics. Hematologia 2013, 4, 239–256. [Google Scholar]
- Sikorska, K.; Bielawski, K.; Romanowski, T.; Stalke, P. Hereditary hemochromatosis: The most frequent inherited human disease. Postepy Hig. Med. Dosw. 2006, 60, 667–676. [Google Scholar]
- Sies, H.; Jones, D.P. Reactive Oxygen Species (ROS) as Pleiotropic Physiological Signalling Agents. Nat. Rev. Mol. Cell Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Chandel, N.S. Fundamentals of cancer metabolism. Sci. Adv. 2016, 2, e1600200. [Google Scholar] [CrossRef] [PubMed]
- Lenton, K.J.; Sané, A.T.; Therriault, H.; Cantin, A.M.; Payette, H.; Wagner, J.R. Vitamin C augments lymphocyte glutathione in subjects with ascorbate deficiency. Am. J. Clin. Nutr. 2003, 77, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Boyacioğlu, M.; Sekkin, S.; Kum, C.; Korkmaz, D.; Kiral, F.; Yalinkilinc, H.S.; Ak, M.O.; Akar, F. The protective effects of vitamin C on the DNA damage, antioxidant defenses and aorta histopathology in chronic hyperhomocysteinemia induced rats. Exp. Toxicol. Pathol. 2014, 66, 407–413. [Google Scholar] [CrossRef]
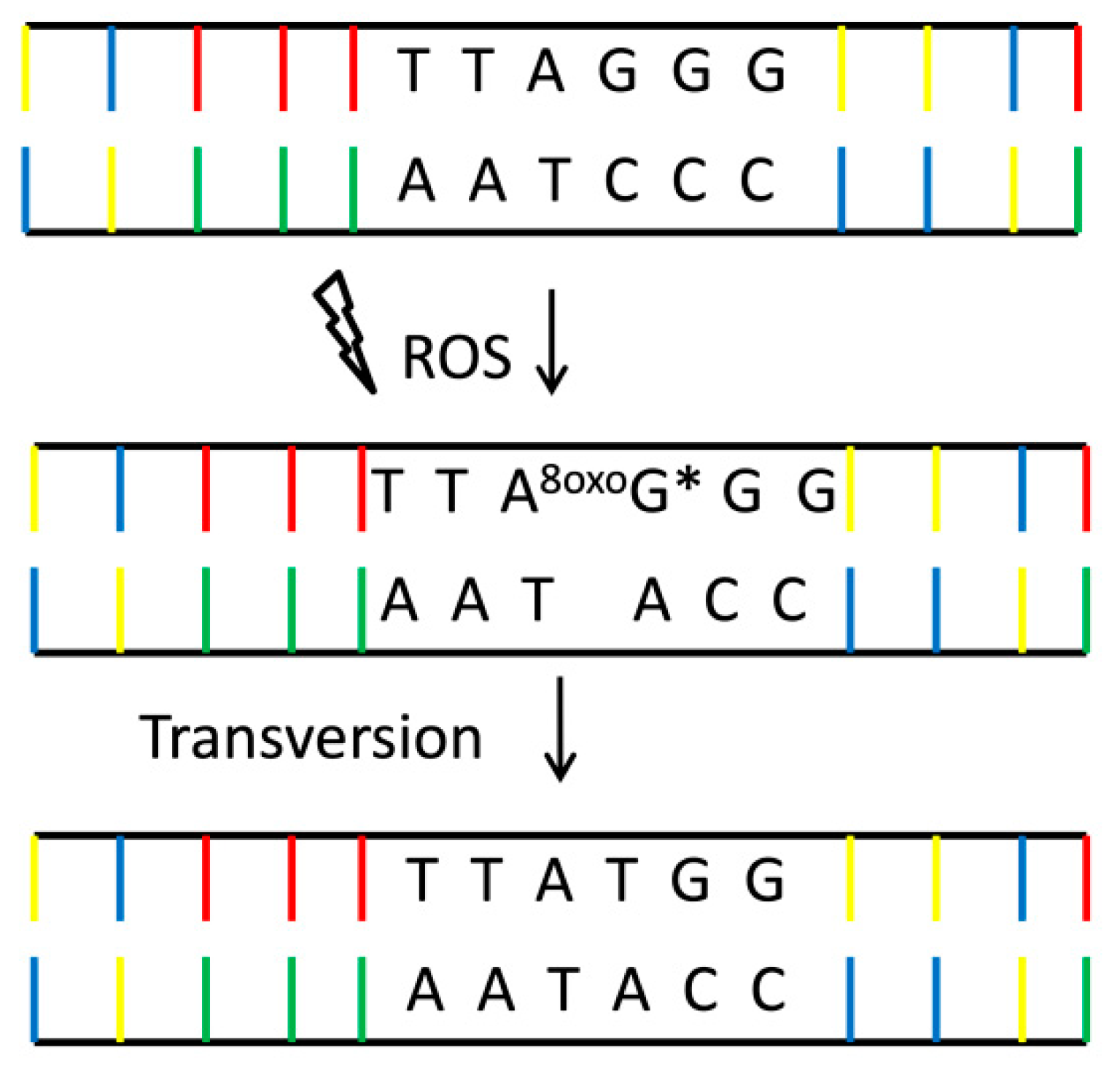
- Ohno, M.; Miura, T.; Furuichi, M.; Tominaga, Y.; Tsuchimoto, D.; Sakumi, K.; Nakabeppu, Y. A genome-wide distribution of 8-oxoguanine correlates with the preferred regions for recombination and single nucleotide polymorphism in the human genome. Genome Res. 2006, 16, 567–575. [Google Scholar] [CrossRef]
- Lutsenko, E.A.; Carcamo, J.M.; Golde, D.W. Vitamin C Prevents DNA Mutation Induced by Oxidative Stress. J. Biol. Chem. 2002, 277, 16895–16899. [Google Scholar] [CrossRef]
- Youn, C.-K.; Song, P.I.; Kim, M.-H.; Kim, J.; Hyun, J.-W.; Choi, S.-J.; Yoon, S.-P.; Chung, M.-H.; Chang, I.-Y.; You, H.J. Human 8-Oxoguanine DNA Glycosylase Suppresses the Oxidative Stress Induced Apoptosis through a p53-Mediated Signaling Pathway in Human Fibroblasts. Mol. Cancer Res. 2007, 5, 1083–1098. [Google Scholar] [CrossRef]
- Kamiya, H. Mutagenicity of oxidized DNA precursors in living cells: Roles of nucleotide pool sanitization and DNA repair enzymes, and translesion synthesis DNA polymerases. Mutat. Res./Genet. Toxicol. Environ. Mutagenesis 2010, 703, 32–36. [Google Scholar] [CrossRef]
- Ghosh, R.; Mitchell, D.L. Effect of oxidative DNA damage in promoter elements on transcription factor binding. Nucleic Acids Res. 1999, 27, 3213–3218. [Google Scholar] [CrossRef]
- Raffoul, J.J.; Heydari, A.R.; Hillman, G.G. DNA repair and cancer therapy: Targeting APE1/Ref-1 using dietary agents. J. Oncol. 2012, 2012, 370481. [Google Scholar] [CrossRef]
- Tudek, B.; Swoboda, M.; Kowalczyk, P.; Oliński, R. Modulation of oxidative DNA damage repair by the diet, inflammation and neoplastic transformation. J. Physiol. Pharmacol. 2006, 57, 33–49. [Google Scholar] [PubMed]
- Li, Y.; Schellhorn, H.E. New Developments and Novel Therapeutic Perspectives for Vitamin C. J. Nutr. 2007, 137, 2171–2184. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.S.; Evans, M.D.; Podmore, I.D.; Herbert, K.E.; Mistry, N.; Mistry, P.; Lunec, J. Novel repair action of vitamin C upon in vivo oxidative DNA damage. FEBS Lett. 1998, 439, 363–367. [Google Scholar] [CrossRef]
- Lunec, J.; Holloway, K.A.; Cooke, M.S.; Faux, S.; Griffiths, H.R.; Evans, M.D. Urinary 8-oxo-2′-deoxyguanosine: Redox regulation of DNA repair in vivo? Free Radic. Biol. Med. 2002, 33, 875–885. [Google Scholar] [CrossRef]
- Bevan, R.J.; Mistry, N.; Patel, P.R.; Halligan, E.P.; Dove, R.; Lunec, J. Can vitamin C induce nucleotide excision repair? Support from in vitro evidence. Br. J. Nutr. 2009, 103, 686–695. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Duarte, T.L.; Cooke, M.S.; Jones, G.D.D. Gene expression profiling reveals new protective roles for vitamin C in human skin cells. Free Radic. Biol. Med. 2009, 46, 78–87. [Google Scholar] [CrossRef]
- Tarng, D.-C.; Liu, T.-Y.; Huang, T.-P. Protective effect of vitamin C on 8-hydroxy-2′-deoxyguanosine level in peripheral blood lymphocytes of chronic hemodialysis patients. Kidney Int. 2004, 66, 820–831. [Google Scholar] [CrossRef]
- Guarnieri, S.; Loft, S.; Riso, P.; Porrini, M.; Risom, L.; Poulsen, H.E.; Møller, P. DNA repair phenotype and dietary antioxidant supplementation. Br. J. Nutr. 2008, 99, 1018–1024. [Google Scholar] [CrossRef]
- Bonucci, J.; Gragnani, A.; Trincado, M.M.; Vincentin, V.; Correa, S.A.A.; Ferreira, L.M. The role of vitamin C in the gene expression of oxidative stress markers in fibroblasts from burn patients. Acta Cir. Bras. 2018, 33, 703–712. [Google Scholar] [CrossRef]
- Cooper, S.E.; Hodimont, E.; Green, C.M. A fluorescent bimolecular complementation screen reveals MAF1, RNF7 and SETD3 as PCNAassociated proteins in human cells. Cell Cycle 2015, 14, 2509–2519. [Google Scholar] [CrossRef]
- Karademirci, M.; Kutlu, R.; Kilinc, I. Relationship between smoking and total antioxidant status, total oxidant status, oxidative stress index, vit C, vit E. Clin. Respir. J. 2018, 12, 2006–2012. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, Y.; Aydemir, Ö.; Şengül, A.; Güngen, A.C.; Çoban, H.; Taşdemir, C.; Şehitoğulları, A. Comparison of oxidant/antioxidant balance in COPD and non-COPD smokers. Heart Lung 2019, 48, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Fearon, I.M.; Phillips, G.; Carr, T.; Taylor, M.; Breheny, D.; Faux, S.P. The Role of Oxidative Stress in Smoking-Related Diseases. Mini-Rev. Org. Chem. 2011, 8, 360–371. [Google Scholar] [CrossRef]
- Perfetti, T.; Rodgman, A. The Complexity of Tobacco and Tobacco Smoke. Beiträge Zur Tab. Int. /Contrib. Tob. Res. 2011, 24, 215–232. [Google Scholar] [CrossRef]
- Moller, P.; Viscovich, M.; Lykkesfeldt, J.; Loft, S.; Jensen, A.; Poulsen, H.E. Vitamin C supplementation decreases oxidative DNA damage in mononuclear blood cells of smokers. Eur. J. Nutr. 2004, 43, 267–274. [Google Scholar] [CrossRef]
- Lee, B.M.; Lee, S.K.; Kim, H.S. Inhibition of oxidative DNA damage, 8-OHdG, and carbonyl contents in smokers treated with antioxidants (vitamin E, vitamin C, β-carotene and red ginseng). Cancer Lett. 1998, 132, 219–227. [Google Scholar] [CrossRef]
- Rossner, P.; Uhlirova, K.; Beskid, O.; Rossnerova, A.; Svecova, V.; Sram, R.J. Expression of XRCC5 in peripheral blood lymphocytes is upregulated in subjects from a heavily polluted region in the Czech Republic. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2011, 713, 76–82. [Google Scholar] [CrossRef]
- Konopacka, M.; Palyvoda, O.; Rzeszowska-Wolny, J. Inhibitory effect of ascorbic acid post-treatment on radiation-induced chromosomal damage in human lymphocytes in vitro. Teratog. Carcinog. Mutagenesis 2002, 22, 443–450. [Google Scholar] [CrossRef]
- Kobashigawa, S.; Kashino, G.; Suzuki, K.; Yamashita, S.; Mori, H. Ionizing Radiation-Induced Cell Death Is Partly Caused by Increase of Mitochondrial Reactive Oxygen Species in Normal Human Fibroblast Cells. Radiat. Res. 2015, 183, 455–464. [Google Scholar] [CrossRef]
- Maeda, J.; Allum, A.; Mussallem, J.T.; Froning, C.E.; Haskins, A.; Buckner, M.; Kato, T. Ascorbic Acid 2-Glucoside Pretreatment Protects Cells from Ionizing Radiation, UVC, and Short Wavelength of UVB. Genes 2020, 11, 238. [Google Scholar] [CrossRef]
- Kobashigawa, S.; Kashino, G.; Mori, H.; Watanabe, M. Relief of delayed oxidative stress by ascorbic acid can suppress radiation-induced cellular senescence in mammalian fibroblast cells. Mech. Ageing Dev. 2015, 146, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, S.; Funakoshi, T.; Sato, Y.; Saito, N.; Ohsawa, H.; Kurita, K.; Ishigami, A. Protective effect of pre- and post-vitamin C treatments on UVB-irradiation-induced skin damage. Sci. Rep. 2018, 8, 16199. [Google Scholar] [CrossRef] [PubMed]
- Velauthapillai, N.; Barfett, J.; Jaffer, H.; Mikulis, D.; Murphy, K. Antioxidants Taken Orally prior to Diagnostic Radiation Exposure Can Prevent DNA Injury. J. Vasc. Interv. Radiol. 2017, 28, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Domina, E.A.; Pylypchuk, O.P.; Mikhailenko, V.M. Destabilization of human cell genome under the combined effect of radiation and ascorbic acid. Exp. Oncol. 2014, 36, 236–240. [Google Scholar]
- Lenton, K.J.; Therriault, H.; Cantin, A.M.; Fülöp, T.; Payette, H.; Wagner, J.R. Direct correlation of glutathione and ascorbate and their dependence on age and season in human lymphocytes. Am. J. Clin. Nutr. 2000, 71, 1194–1200. [Google Scholar] [CrossRef]
- Mezzetti, A.; Lapenna, D.; Romano, F.; Costantini, F.; Pierdomenico, S.D.; De Cesare, D. Systemic Oxidative Stress and Its Relationship with Age and Illness. J. Am. Geriatr. Soc. 1996, 44, 823–827. [Google Scholar] [CrossRef]
- Jacob, R.; Sotoudeh, G. Vitamin C function and status in chronic disease. Nutr. Clin. Care 2002, 5, 66–74. [Google Scholar] [CrossRef]
- Moszczyński, P.; Pyć, R. Biochemia Witamin. Witaminy Lipofilne i Kwas Askorbinowy. Część II; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2001. [Google Scholar]
- Carpenter, K.J. The discovery of Vitamin C. Ann. Nutr. Metab. 2012, 61, 259–264. [Google Scholar] [CrossRef]
- Cárcamo, J.M.; Pedraza, A.; Bórquez-Ojeda, O.; Zhang, B.; Sanchez, R.; Golde, D.W. Vitamin C is a kinase inhibitor: Dehydroascorbic acid inhibits IkappaBalpha kinase beta. Mol. Cell. Biol. 2004, 24, 6645–6652. [Google Scholar] [CrossRef]







{kind=link}
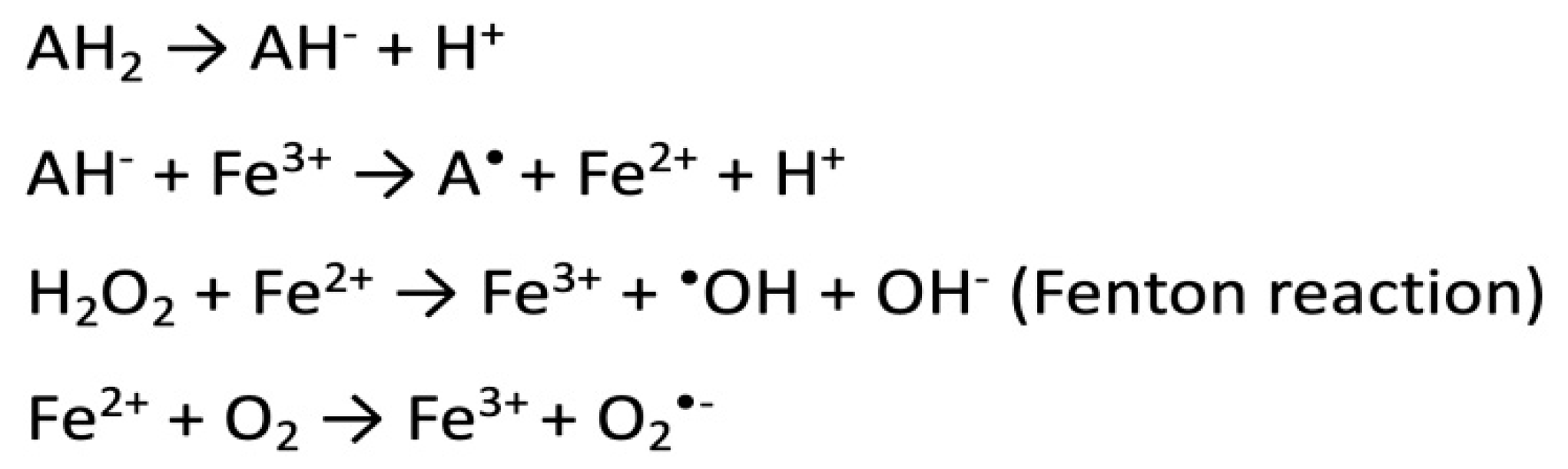{kind=link}
{kind=link}
{kind=link}
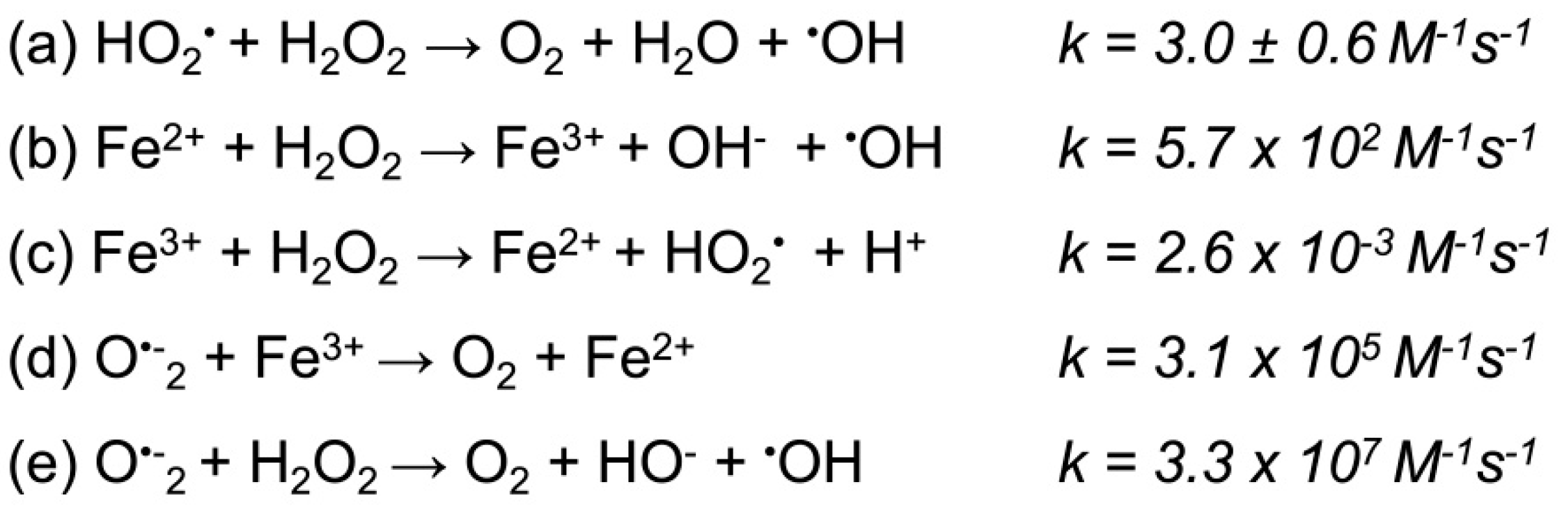{kind=link}
{kind=link}
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaźmierczak-Barańska, J.; Boguszewska, K.; Adamus-Grabicka, A.; Karwowski, B.T. Two Faces of Vitamin C—Antioxidative and Pro-Oxidative Agent. Nutrients 2020, 12, 1501. https://doi.org/10.3390/nu12051501
Kaźmierczak-Barańska J, Boguszewska K, Adamus-Grabicka A, Karwowski BT. Two Faces of Vitamin C—Antioxidative and Pro-Oxidative Agent. Nutrients. 2020; 12(5):1501. https://doi.org/10.3390/nu12051501
Chicago/Turabian StyleKaźmierczak-Barańska, Julia, Karolina Boguszewska, Angelika Adamus-Grabicka, and Bolesław T. Karwowski. 2020. "Two Faces of Vitamin C—Antioxidative and Pro-Oxidative Agent" Nutrients 12, no. 5: 1501. https://doi.org/10.3390/nu12051501
APA StyleKaźmierczak-Barańska, J., Boguszewska, K., Adamus-Grabicka, A., & Karwowski, B. T. (2020). Two Faces of Vitamin C—Antioxidative and Pro-Oxidative Agent. Nutrients, 12(5), 1501. https://doi.org/10.3390/nu12051501