The Protective Effects of Acer okamotoanum and Isoquercitrin on Obesity and Amyloidosis in a Mouse Model



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Sample Preparation
2.3. Animals
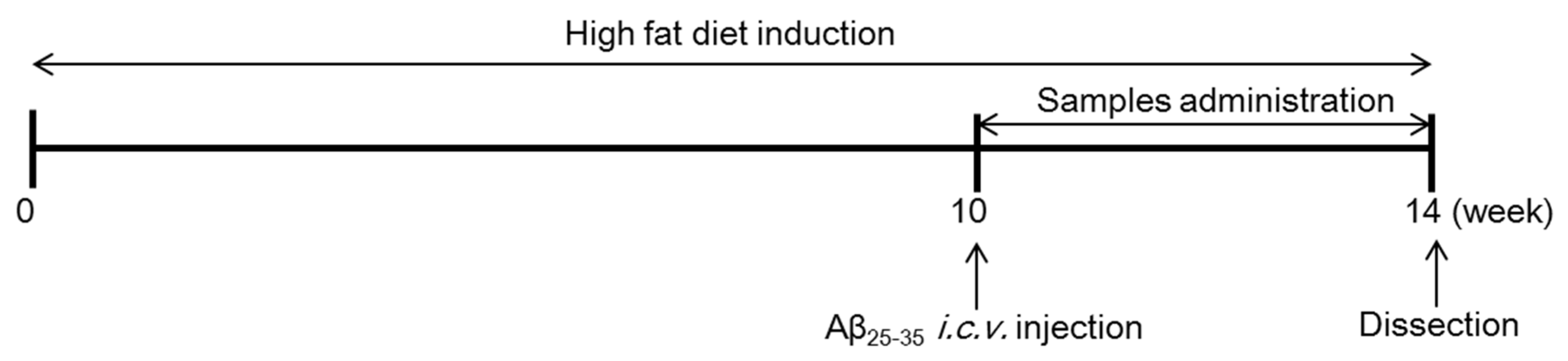
2.4. Groups and Treatment
2.5. Aβ25–35 i.c.v. Injection
2.6. Biochemical Analysis
2.7. Histopathological Analysis
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
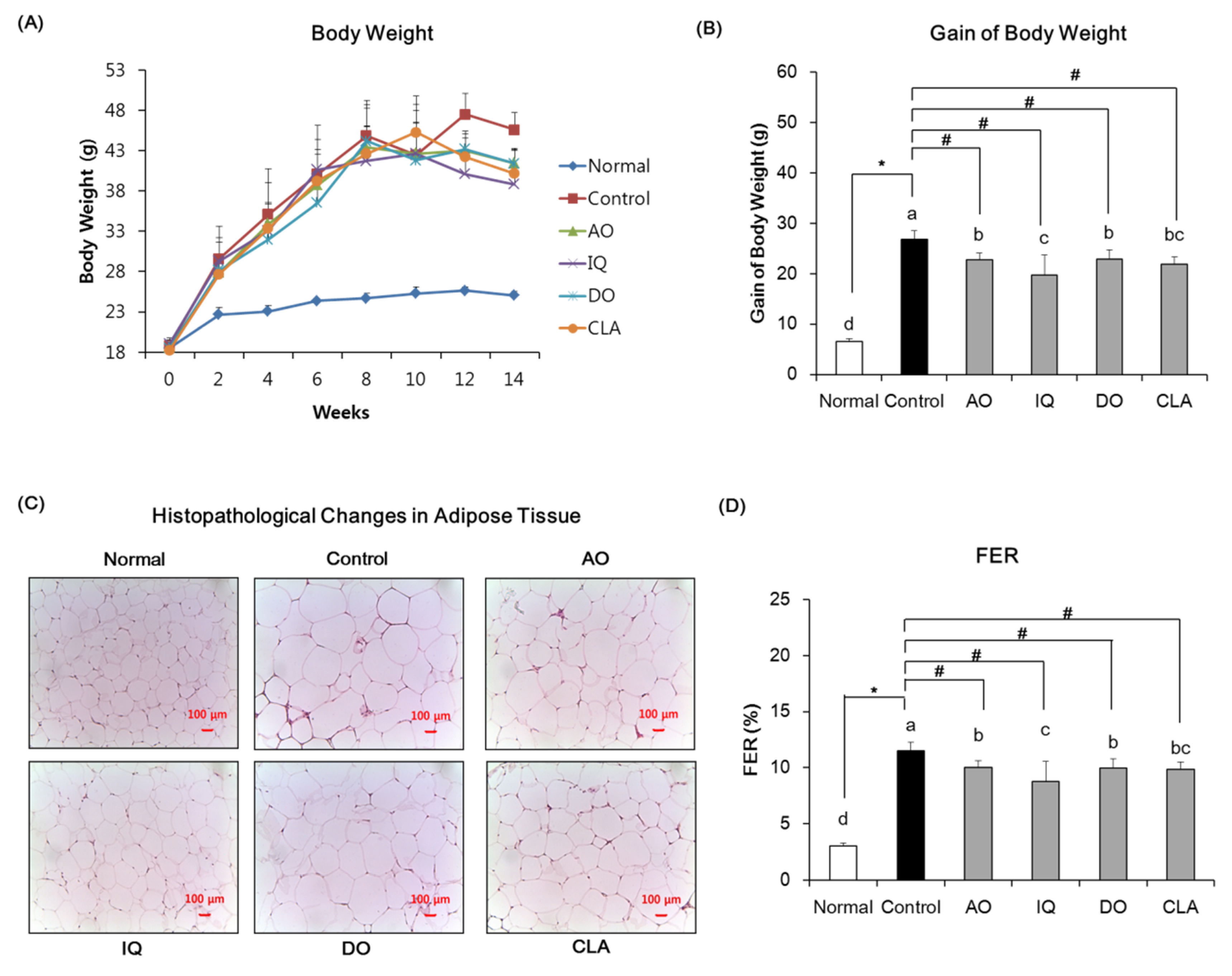
3.1. Effect of EAO and Isoquercitrin on Body Weight
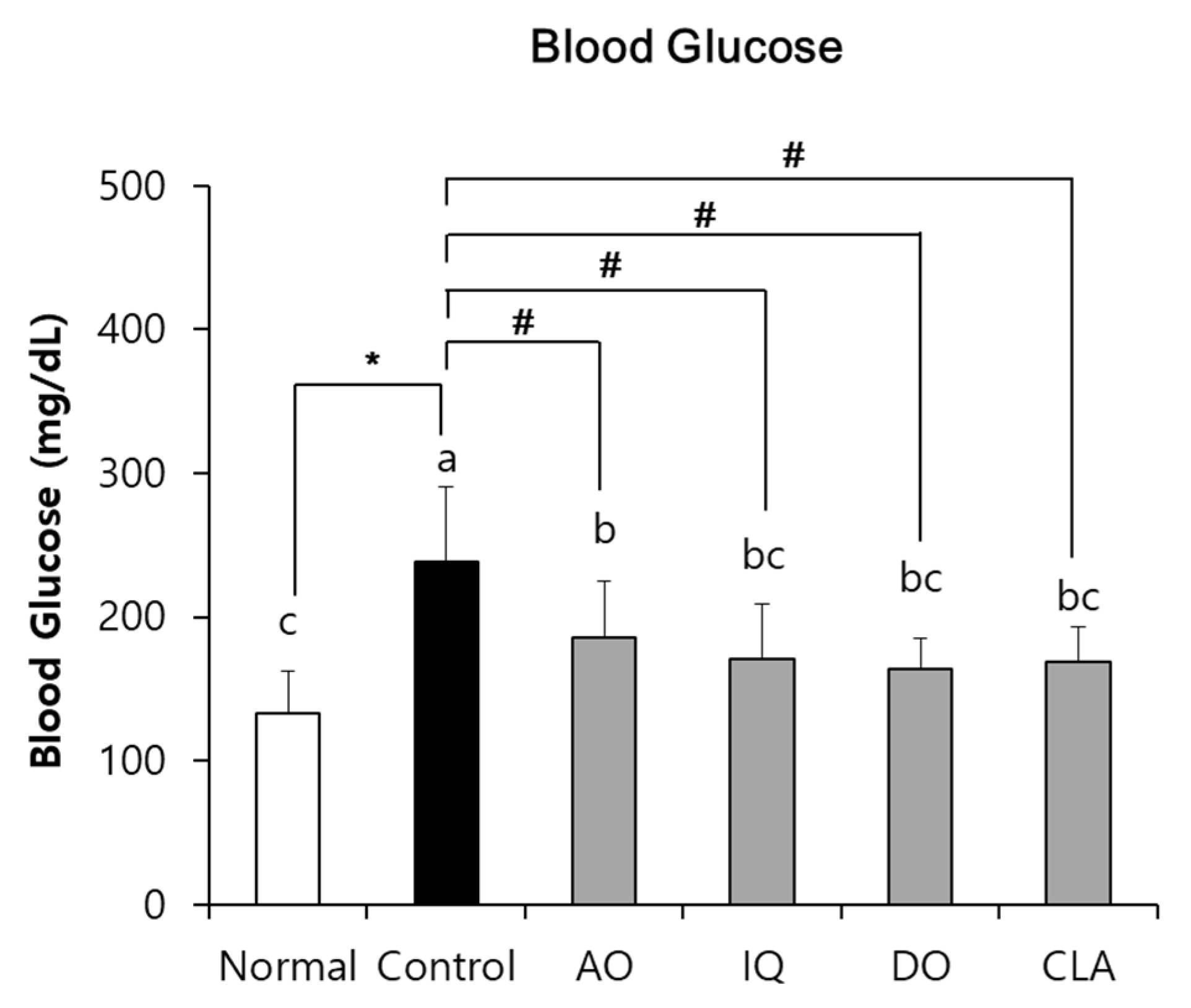
3.2. Effect of EAO and Isoquercitrin on Blood Glucose Level
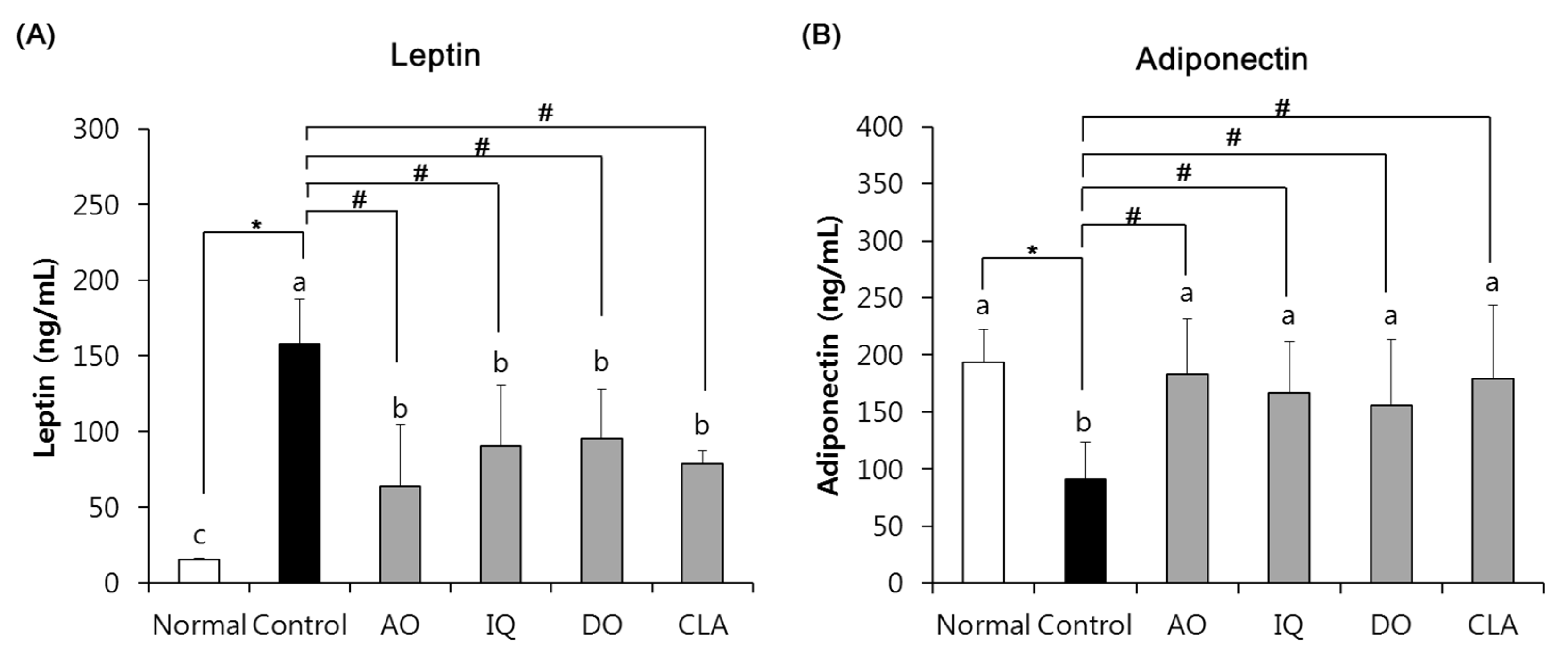
3.3. Effect of EAO and Isoquercitrin on Adipokines Changes
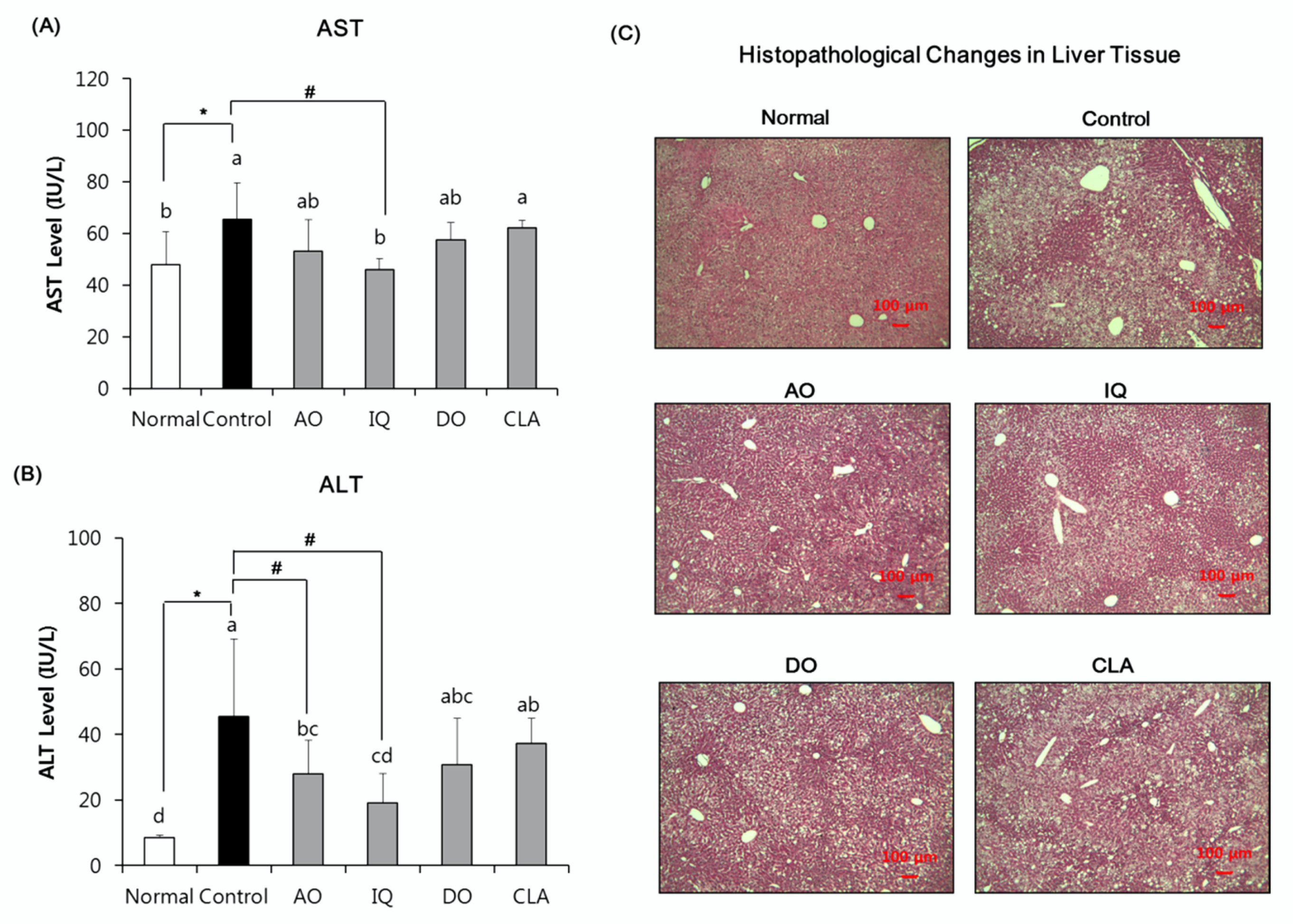
3.4. Effect of EAO and Isoquercitrin on Hepatic Damage
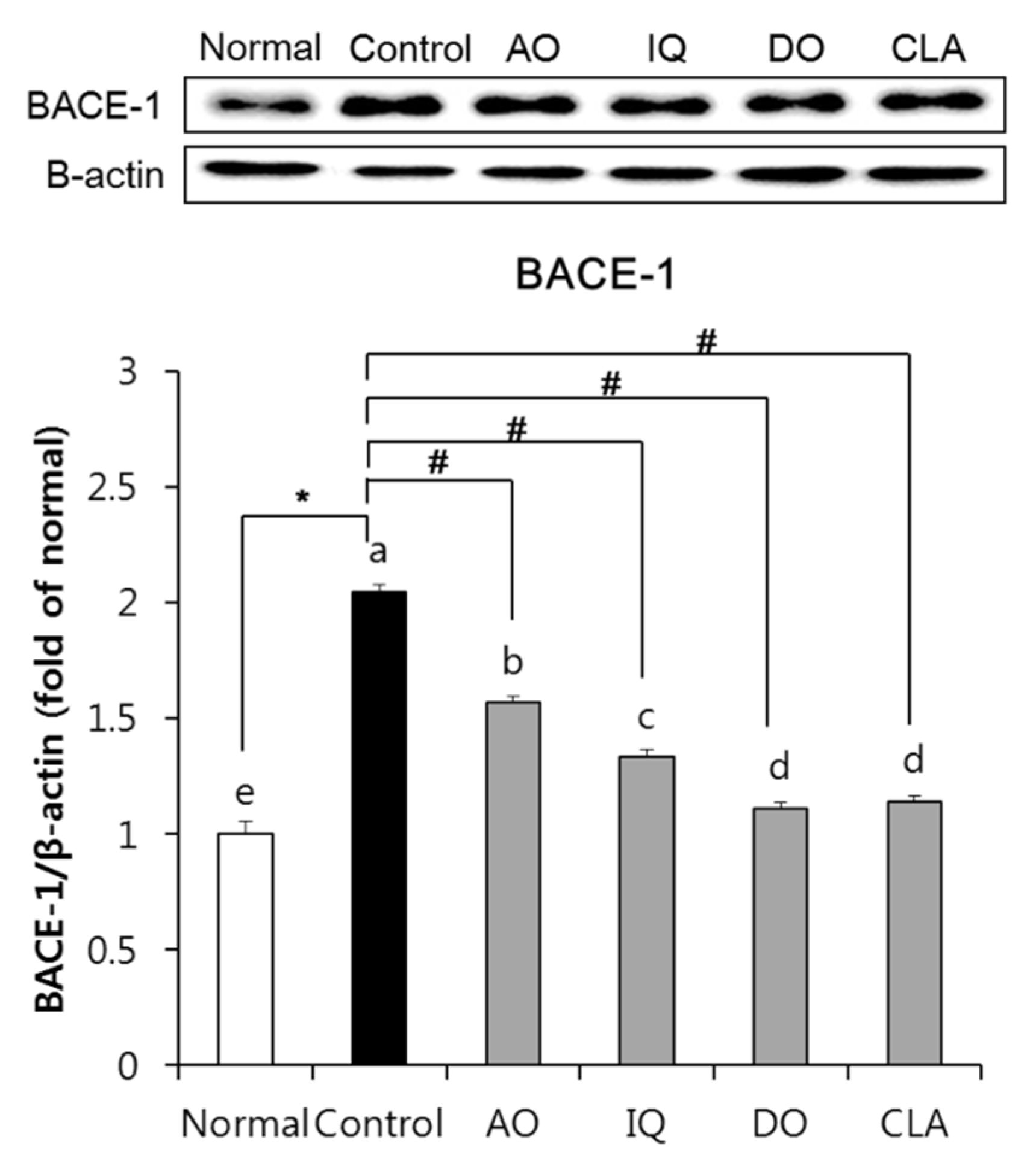
3.5. Effect of EAO and Isoquercitrin on β-Secretase Activity in the Brain Tissue
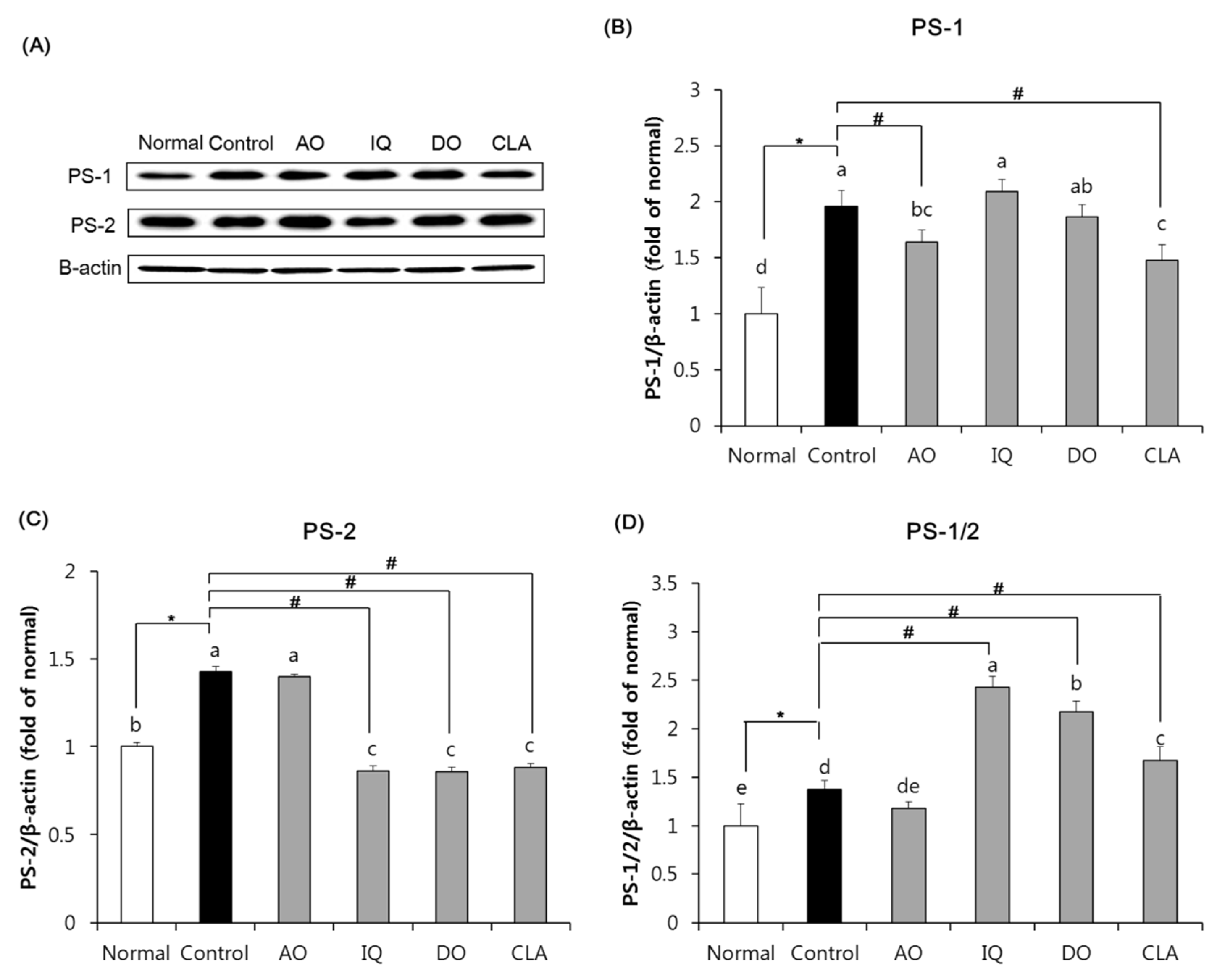
3.6. Effect of EAO and Isoquercitrin on γ-Secretase Activity in the Brain Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Armitage, J.A.; Poston, L.; Taylor, P.D. Developmental origins of obesity and the metabolic syndrome: The role of maternal obesity. Front. Horm. Res. 2008, 36, 73–84. [Google Scholar]
- Sandouk, Z.; Lansang, M.C. Diabetes with obesity-Is there an ideal diet? Clevel. Clin. J. Med. 2017, 84, 4–14. [Google Scholar] [CrossRef]
- Malik, V.S.; Willett, W.C.; Hu, F.B. Global obesity: Trends, risk factors and policy implications. Nat. Rev. Endocrinol. 2012, 9, 13–27. [Google Scholar] [CrossRef]
- Martin-Jiménez, C.A.; Gaitán-Vaca, D.M.; Echeverria, V.; González, J.; Barreto, G.E. Relationship between Obesity, Alzheimer’s Disease, and Parkinson’s Disease: An Astrocentric View. Mol. Neurobiol. 2016, 54, 7096–7115. [Google Scholar] [CrossRef]
- Grant, W.B. Using multicountry ecological and observational studies to determine dietary risk factors for Alzheimer’s disease. J. Am. Coll. Nutr. 2016, 35, 476–489. [Google Scholar] [CrossRef]
- Walker, J.M.; Dixit, S.; Saulsberry, A.C.; May, J.M.; Harrison, F.E. Reversal of high fat diet-induced obesity improves glucose tolerance, inflammatory response, β-amyloid accumulation and cognitive decline in the APP/PSEN1 mouse model of Alzheimer’s disease. Neurobiol. Dis. 2017, 100, 87–98. [Google Scholar] [CrossRef]
- Knight, E.M.; Martins, I.V.; Gümüsgöz, S.; Allan, S.M.; Lawrence, C.B. High-fat diet-induced memory impairment in triple-transgenic Alzheimer’s disease (3xTgAD) mice is independent of changes in amyloid and tau pathology. Neurobiol. Aging 2014, 35, 1821–1832. [Google Scholar] [CrossRef]
- Mohamed, H.E.; Abo-Elmatty, D.M.; Mesbah, N.; Saleh, S.M.; Ali, A.-M.A.; Sakr, A.T. Raspberry ketone preserved cholinergic activity and antioxidant defense in obesity induced Alzheimer disease in rats. Biomed. Pharmacother. 2018, 107, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, D.; Picone, P.; Baldassano, S.; Caruana, L.; Messina, E.; Gammazza, A.; Cappello, F.; Mulè, F.; Di Carlo, M. Insulin Resistance as Common Molecular Denominator Linking Obesity to Alzheimer’s Disease. Curr. Alzheimer Res. 2015, 12, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Kothari, V.; Luo, Y.; Tornabene, T.; O’Neill, A.M.; Greene, M.; Geetha, T.; Babu, J.R. High fat diet induces brain insulin resistance and cognitive impairment in mice. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Si, X.; Li, Y.; Jiang, Y.; Shang, W.; Shui, G.; Lam, S.M.; Blanchard, C.; Strappe, P.; Zhou, Z.; Blanchard, C.L. γ-Aminobutyric Acid Attenuates High-Fat Diet-Induced Cerebral Oxidative Impairment via Enhanced Synthesis of Hippocampal Sulfatides. J. Agric. Food Chem. 2018, 67, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- Maesako, M.; Uemura, K.; Iwata, A.; Kubota, M.; Watanabe, K.; Uemura, M.T.; Noda, Y.; Asada-Utsugi, M.; Kihara, T.; Takahashi, R.; et al. Continuation of Exercise Is Necessary to Inhibit High Fat Diet-Induced β-Amyloid Deposition and Memory Deficit in Amyloid Precursor Protein Transgenic Mice. PLoS ONE 2013, 8, e72796. [Google Scholar] [CrossRef]
- Mendiola-Precoma, J.; Berumen, L.C.; Padilla, K.; García-Alcocer, G. Therapies for Prevention and Treatment of Alzheimer’s Disease. BioMed Res. Int. 2016, 2016, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, F.; Khazaei, M.; Komaki, A.; Amiri, I.; Jalili, C. Virgin coconut oil (VCO) by normalizing NLRP3 inflammasome showed potential neuroprotective effects in Amyloid-β induced toxicity and high-fat diet fed rat. Food Chem. Toxicol. 2018, 118, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Asadbegi, M.; Komaki, A.; Salehi, I.; Yaghmaei, P.; Ebrahim-Habibi, A.; Shahidi, S.; Sarihi, A.; Asl, S.S.; Golipoor, Z. Effects of thymol on amyloid-β-induced impairments in hippocampal synaptic plasticity in rats fed a high-fat diet. Brain Res. Bull. 2018, 137, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Asadbegi, M.; Yaghmaei, P.; Salehi, I.; Komaki, A.; Ebrahim-Habibi, A. Investigation of thymol effect on learning and memory impairment induced by intrahippocampal injection of amyloid beta peptide in high fat diet- fed rats. Metab. Brain Dis. 2017, 32, 827–839. [Google Scholar] [CrossRef]
- Jin, W.; Thuong, P.T.; Su, N.D.; Min, B.S.; Son, K.H.; Chang, H.W.; Kim, H.P.; Kang, S.S.; Sok, D.E.; Bae, K. Antioxidant activity of cleomiscosins A and C isolated from Acer okamotoanum. Arch. Pharmacal Res. 2007, 30, 275–281. [Google Scholar] [CrossRef]
- An, B.-S.; Kang, J.-H.; Yang, H.; Yang, M.-P.; Jeung, E.-B. Effects of Acer okamotoanum sap on the function of polymorphonuclear neutrophilic leukocytes in vitro and in vivo. Mol. Med. Rep. 2012, 7, 654–658. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, S.; Cho, E.J. Acer okamotoanum protects SH-SY5Y neuronal cells against hydrogen peroxide-induced oxidative stress. Food Sci. Biotechnol. 2018, 28, 191–200. [Google Scholar] [CrossRef]
- Kim, J.H.; Quilantang, N.G.; Kim, H.Y.; Lee, S.; Cho, E.J. Attenuation of hydrogen peroxide-induced oxidative stress in SH-SY5Y cells by three flavonoids from Acer okamotoanum. Chem. Pap. 2018, 73, 1135–1144. [Google Scholar] [CrossRef]
- Kim, E.J.; Kang, M.J.; Seo, Y.B.; Nam, S.W.; Kim, G.D. Acer okamotoanum Nakai leaf extract inhibits adipogenesis via suppressing expression of PPARγ and C/EBPα in 3T3-L1 cells. J. Microbiol. Biotechnol. 2018, 28, 1645–1653. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, D.-G.; Rodriguez, J.P.; Park, J.Y.; Cho, E.J.; Jacinto, S.D.; Lee, S. Determination of flavonoids in Acer okamotoanum and their aldose reductase inhibitory activities. Hortic. Environ. Biotechnol. 2018, 59, 131–137. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, S.; Cho, E.J. Acer okamotoanum and isoquercitrin improve cognitive function via attenuation of oxidative stress in high fat diet- and amyloid beta-induced mice. Food Funct. 2019, 10, 6803–6814. [Google Scholar] [CrossRef] [PubMed]
- Knowles, J. Donepezil in Alzheimer’s disease: An evidence-based review of its impact on clinical and economic outcomes. Core Évid. 2006, 1, 195–219. [Google Scholar]
- Koba, K.; Yanagita, T. Health benefits of conjugated linoleic acid (CLA). Obes. Res. Clin. Pract. 2014, 8, e525–e532. [Google Scholar] [CrossRef]
- Laursen, S.E.; Belknap, J. Intracerebroventricular injections in mice. J. Pharmacol. Methods 1986, 16, 355–357. [Google Scholar] [CrossRef]
- Choi, J.G.; Moon, M.; Kim, H.G.; Mook-Jung, I.; Chung, S.Y.; Kang, T.H.; Kim, S.Y.; Lee, E.H.; Oh, M.S. Gami-Chunghyuldan ameliorates memory impairment and neurodegeneration induced by intrahippocampal Aβ1–42 oligomer injection. Neurobiol. Learn. Mem. 2011, 96, 306–314. [Google Scholar] [CrossRef]
- Prickett, C.; Brennan, L.; Stolwyk, R. Examining the relationship between obesity and cognitive function: A systematic literature review. Obes. Res. Clin. Pract. 2015, 9, 93–113. [Google Scholar] [CrossRef]
- Winocur, G.; Greenwood, C.E. Studies of the effects of high fat diets on cognitive function in a rat model. Neurobiol. Aging 2005, 26, 46–49. [Google Scholar] [CrossRef]
- Ow, S.-Y.; Dunstan, D.E. A brief overview of amyloids and Alzheimer’s disease. Protein Sci. 2014, 23, 1315–1331. [Google Scholar] [CrossRef]
- Lin, B.; Hasegawa, Y.; Takane, K.; Koibuchi, N.; Cao, C.; Kim-Mitsuyama, S. High-fat-diet intake enhances cerebral amyloid angiopathy and cognitive impairment in a mouse model of Alzheimer’s disease, independently of metabolic disorders. J. Am. Heart Assoc. 2016, 5, e003154. [Google Scholar] [CrossRef] [PubMed]
- Barron, A.; Tokunaga, M.; Zhang, M.-R.; Ji, B.; Suhara, T.; Higuchi, M. Assessment of neuroinflammation in a mouse model of obesity and β-amyloidosis using PET. J. Neuroinflammation 2016, 13, 221. [Google Scholar] [CrossRef]
- Kim, D.-G.; Krenz, A.; Toussaint, L.E.; Maurer, K.J.; Robinson, S.-A.; Yan, A.; Torres, L.; Bynoe, M.S. Non-alcoholic fatty liver disease induces signs of Alzheimer’s disease (AD) in wild-type mice and accelerates pathological signs of AD in an AD model. J. Neuroinflammation 2016, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Kim, J.H.; Lee, J.; Lee, S.; Cho, E.J. Protective effect of Acer okamotoanum from oxidative stress in C6 glial cells. J. Appl. Biol. Chem. 2017, 60, 141–147. [Google Scholar] [CrossRef]
- Choi, S.Y.; Lee, J.; Lee, D.G.; Lee, S.; Cho, E.J. Acer okamotoanum improves cognition and memory function in Aβ25–35-induced Alzheimer’s mice model. Appl. Biol. Chem. 2016, 60, 1–9. [Google Scholar] [CrossRef]
- Lee, S.-H.; Kim, B.; Oh, M.J.; Yoon, J.; Kim, H.Y.; Lee, K.J.; Lee, J.D.; Choi, K.-Y. Persicaria hydropiper(L.) Spach and its Flavonoid Components, Isoquercitrin and Isorhamnetin, Activate the Wnt/β-catenin Pathway and Inhibit Adipocyte Differentiation of 3T3-L1 Cells. Phytother. Res. 2011, 25, 1629–1635. [Google Scholar] [CrossRef]
- Qin, G.; Ma, J.; Huang, Q.; Yin, H.; Han, J.; Li, M.; Deng, Y.; Wang, B.; Hassan, W.; Shang, J. Isoquercetin Improves Hepatic Lipid Accumulation by Activating AMPK Pathway and Suppressing TGF-β Signaling on an HFD-Induced Nonalcoholic Fatty Liver Disease Rat Model. Int. J. Mol. Sci. 2018, 19, 4126. [Google Scholar] [CrossRef]
- Masuyama, H.; Mitsui, T.; Eguchi, T.; Tamada, S.; Hiramatsu, Y. The effects of paternal high-fat diet exposure on offspring metabolism with epigenetic changes in the mouse adiponectin and leptin gene promoters. Am. J. Physiol. Metab. 2016, 311, E236–E245. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Flier, J.S. Adipose Tissue as an Endocrine Organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Myers, M.G. Leptin receptor signaling and the regulation of mammalian physiology. Recent Prog. Horm. Res. 2004, 59, 287–304. [Google Scholar] [CrossRef]
- Stefan, N.; Stumvoll, M. Adiponectin - Its Role in Metabolism and Beyond. Horm. Metab. Res. 2002, 34, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-W.; Liu, H.-K.; Lin, L.-C.; Liou, K.-T.; Huang, Y.-C.; Lin, C.-H.; Tzeng, T.-T.; Shie, F.-S.; Tsay, H.-J.; Shiao, Y.-J. Xuefu Zhuyu decoction ameliorates obesity, hepatic steatosis, neuroinflammation, amyloid deposition and cognition impairment in metabolically stressed APPswe/PS1dE9 mice. J. Ethnopharmacol. 2017, 209, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Shie, F.-S.; Shiao, Y.-J.; Yeh, C.-W.; Lin, C.-H.; Tzeng, T.-T.; Hsu, H.-C.; Huang, F.-L.; Tsay, H.-J.; Liu, H.-K. Obesity and Hepatic Steatosis Are Associated with Elevated Serum Amyloid Beta in Metabolically Stressed APPswe/PS1dE9 Mice. PLoS ONE 2015, 10, e0134531. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yoshitomi, H.; Liu, T.H.; Zhou, B.; Sun, W.; Qin, L.; Guo, X.; Huang, L.; Wu, L.; Gao, M. Isoquercitrin activates the AMP–activated protein kinase (AMPK) signal pathway in rat H4IIE cells. BMC Complement. Altern. Med. 2014, 14, 42. [Google Scholar] [CrossRef] [PubMed]
- Jeung, E.-B.; Yoo, Y.-M.; Jung, E.-M.; Kang, H.-Y.; Choi, I.-G.; Choi, K.-C. The sap of Acer okamotoanum decreases serum alcohol levels after acute ethanol ingestion in rats. Int. J. Mol. Med. 2011, 28, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Pinçon, A.; De Montgolfier, O.; Akkoyunlu, N.; Daneault, C.; Pouliot, P.; Villeneuve, L.; Lesage, F.; Levy, B.; Thorin-Trescases, N.; Thorin, E.; et al. Non-Alcoholic Fatty Liver Disease, and the Underlying Altered Fatty Acid Metabolism, Reveals Brain Hypoperfusion and Contributes to the Cognitive Decline in APP/PS1 Mice. Metabolites 2019, 9, 104. [Google Scholar] [CrossRef]
- Estrada, L.D.; Ahumada, P.; Cabrera, D.; Arab, J.P. Liver Dysfunction as a Novel Player in Alzheimer’s Progression: Looking Outside the Brain. Front. Aging Neurosci. 2019, 11, 174. [Google Scholar] [CrossRef]
- Vetrivel, K.S.; Thinakaran, G. Amyloidogenic processing of -amyloid precursor protein in intracellular compartments. Neurology 2005, 66, 69–73. [Google Scholar] [CrossRef]
- Thériault, P.; ElAli, A.; Rivest, S. High fat diet exacerbates Alzheimer’s disease-related pathology in APPswe/PS1 mice. Oncotarget 2016, 7, 67808–67827. [Google Scholar] [CrossRef]
- Sah, S.K.; Lee, C.; Jang, J.H.; Park, G.H. Effect of high-fat diet on cognitive impairment in triple-transgenic mice model of Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2017, 493, 731–736. [Google Scholar] [CrossRef]
- Tucsek, Z.; Toth, P.; Sosnowska, D.; Gautam, T.; Mitschelen, M.; Koller, A.; Szalai, G.; Sonntag, W.E.; Ungvari, Z.; Csiszar, A. Obesity in aging exacerbates blood-brain barrier disruption, neuroinflammation, and oxidative stress in the mouse hippocampus: Effects on expression of genes involved in beta-amyloid generation and Alzheimer’s disease. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2013, 69, 1212–1226. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Lee, M.H.; Lee, S.; Cho, E.J. Alpha-linolenic acid from Perilla frutescens var. japonica oil protects Aβ-induced cognitive impairment through regulation of APP processing and Aβ degradation. J. Agric. Food Chem. 2017, 65, 10719–10729. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Vassar, R. Targeting the β secretase BACE1 for Alzheimer’s disease therapy. Lancet Neurol. 2014, 13, 319–329. [Google Scholar] [CrossRef]
- Epis, R.; Marcello, E.; Gardoni, F.; Di Luca, M. Alpha, beta-and gamma-secretases in Alzheimer’s disease. Front. Biosci. 2012, 4, 1126–1150. [Google Scholar]
- Medoro, A.; Bartollino, S.; Mignogna, D.; Passarella, D.; Porcile, C.; Pagano, A.; Florio, T.; Nizzari, M.; Guerra, G.; Di Marco, R.; et al. Complexity and Selectivity of γ-Secretase Cleavage on Multiple Substrates: Consequences in Alzheimer’s Disease and Cancer. J. Alzheimer’s Dis. 2017, 61, 1–15. [Google Scholar] [CrossRef]
- Ismail, N.; Ismail, M.; Azmi, N.H.; Abu Bakar, M.F.; Yida, Z.; Abdullah, M.A.; Basri, H. Thymoquinone-rich fraction nanoemulsion (TQRFNE) decreases Aβ40 and Aβ42 levels by modulating APP processing, up-regulating IDE and LRP1, and down-regulating BACE1 and RAGE in response to high fat/cholesterol diet-induced rats. Biomed. Pharmacother. 2017, 95, 780–788. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.H.; Lee, S.; Cho, E.J. The Protective Effects of Acer okamotoanum and Isoquercitrin on Obesity and Amyloidosis in a Mouse Model. Nutrients 2020, 12, 1353. https://doi.org/10.3390/nu12051353
Kim JH, Lee S, Cho EJ. The Protective Effects of Acer okamotoanum and Isoquercitrin on Obesity and Amyloidosis in a Mouse Model. Nutrients. 2020; 12(5):1353. https://doi.org/10.3390/nu12051353
Chicago/Turabian StyleKim, Ji Hyun, Sanghyun Lee, and Eun Ju Cho. 2020. "The Protective Effects of Acer okamotoanum and Isoquercitrin on Obesity and Amyloidosis in a Mouse Model" Nutrients 12, no. 5: 1353. https://doi.org/10.3390/nu12051353
APA StyleKim, J. H., Lee, S., & Cho, E. J. (2020). The Protective Effects of Acer okamotoanum and Isoquercitrin on Obesity and Amyloidosis in a Mouse Model. Nutrients, 12(5), 1353. https://doi.org/10.3390/nu12051353