Walnut Oil Alleviates Intestinal Inflammation and Restores Intestinal Barrier Function in Mice

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
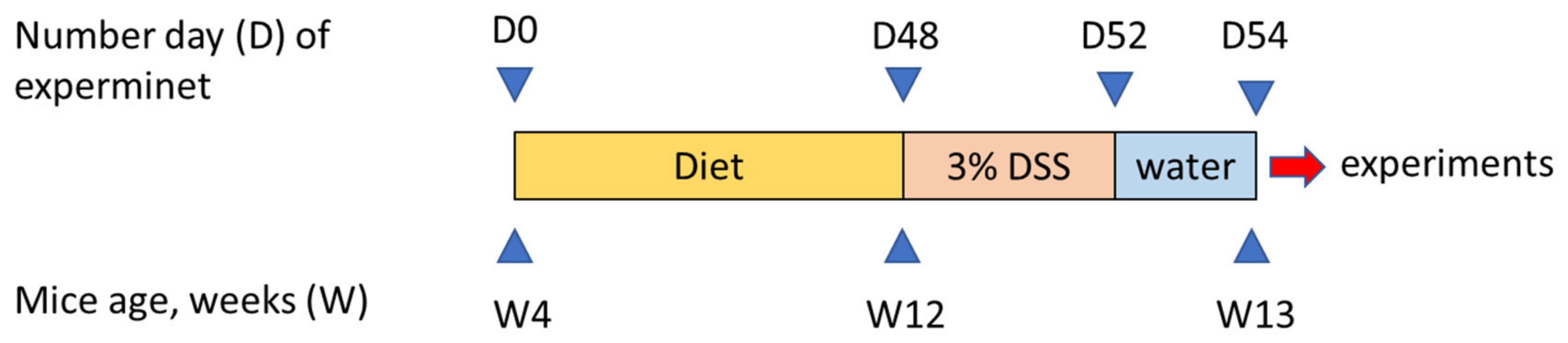
2.1. Animals and Study Design
2.2. Induction of Colitis
2.3. Determination of MPO Activity
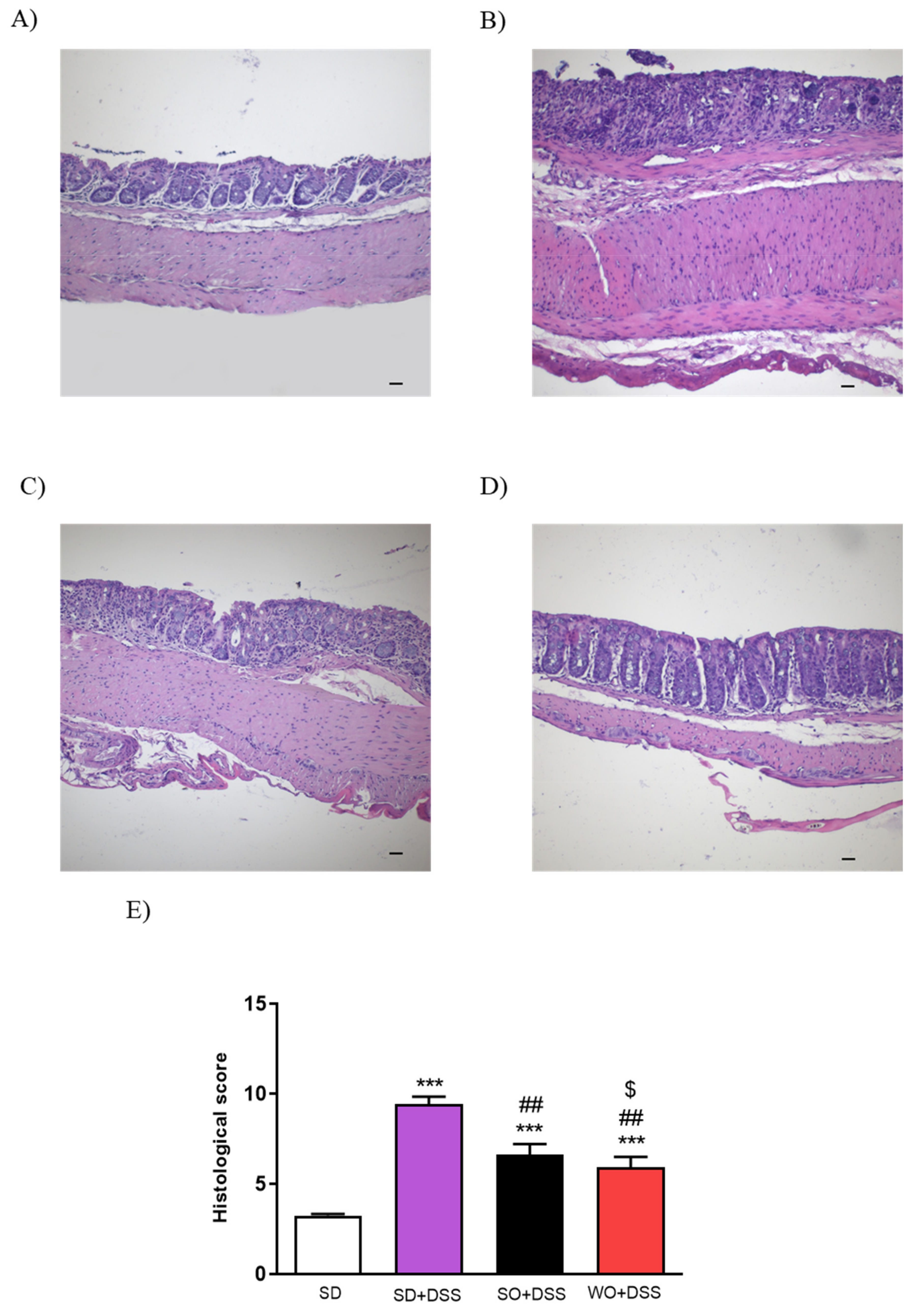
2.4. Histopathological Evaluation
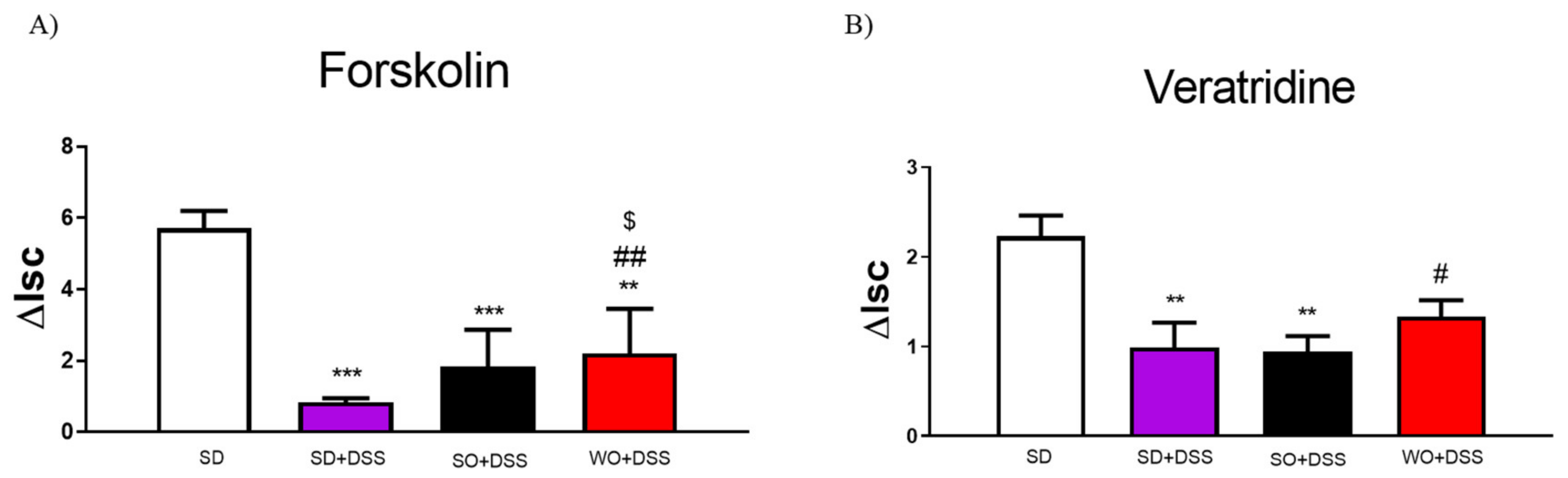
2.5. Ex Vivo Measurement of Epithelial Ion Transport
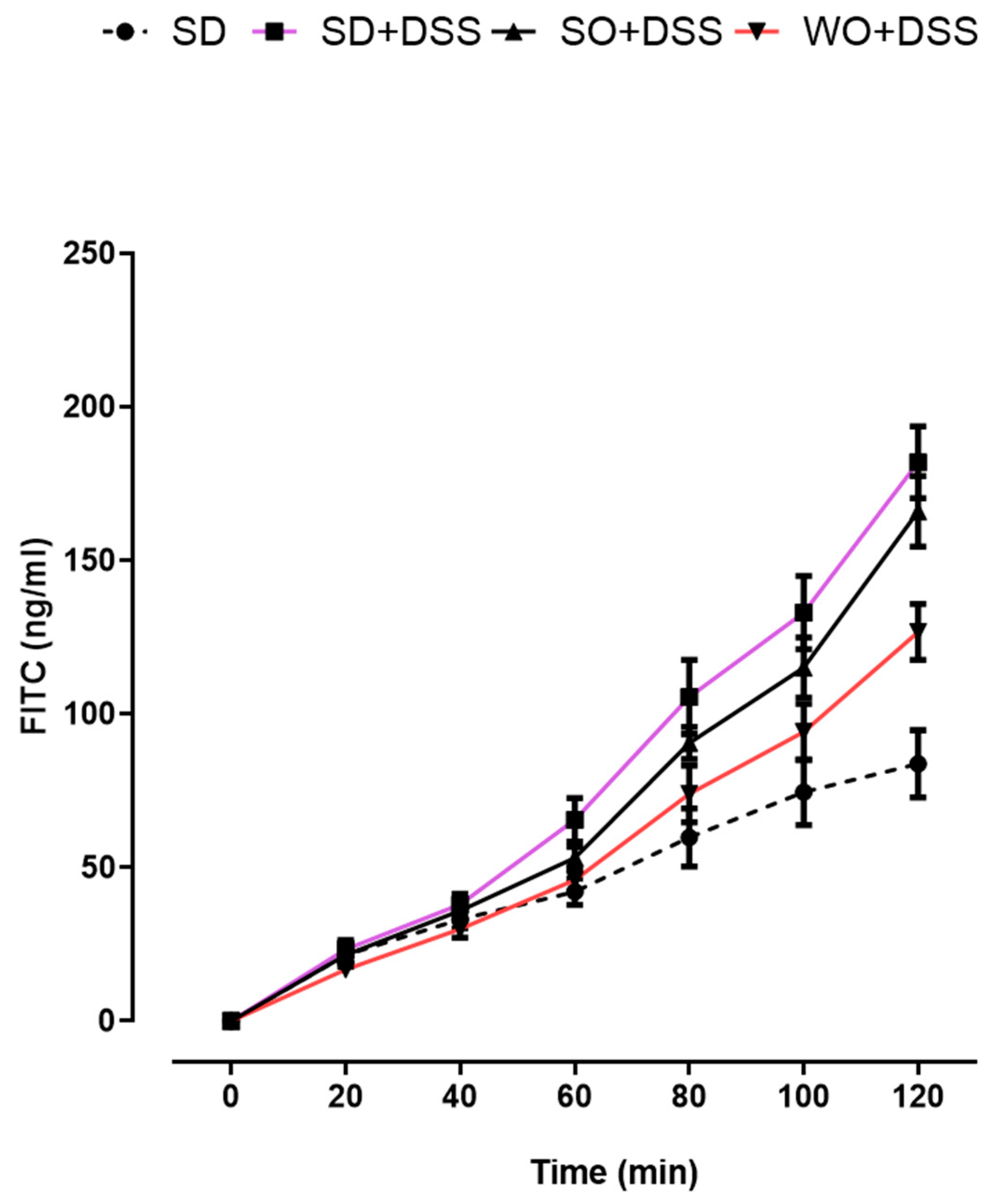
2.6. Ex Vivo Assessment of Intestinal Permeability
2.7. RNA Isolation, Reverse Transcription and qPCR
2.8. Reagents and Drugs
2.9. Statistics
3. Results
3.1. WO Diet Does Not Cause Excessive Body Weight Gain in Mice and Alleviates Symptoms of DSS-Induced Colitis
3.2. WO Inhibits the Effect of DSS on Epithelial Ion Transport in the Mouse Colon
3.3. WO Inhibits the Effect of DSS on Colon Permeability Ex Vivo
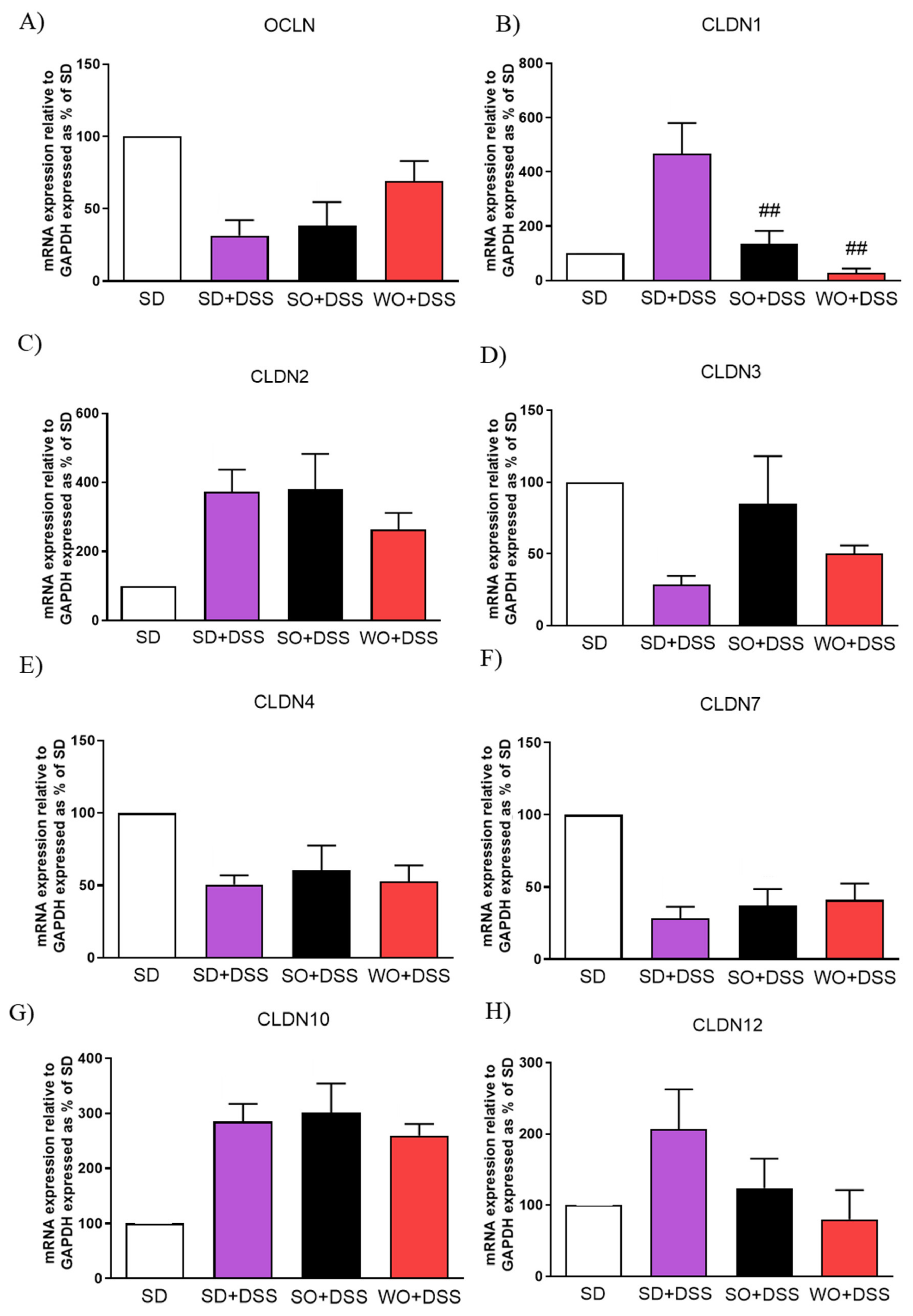
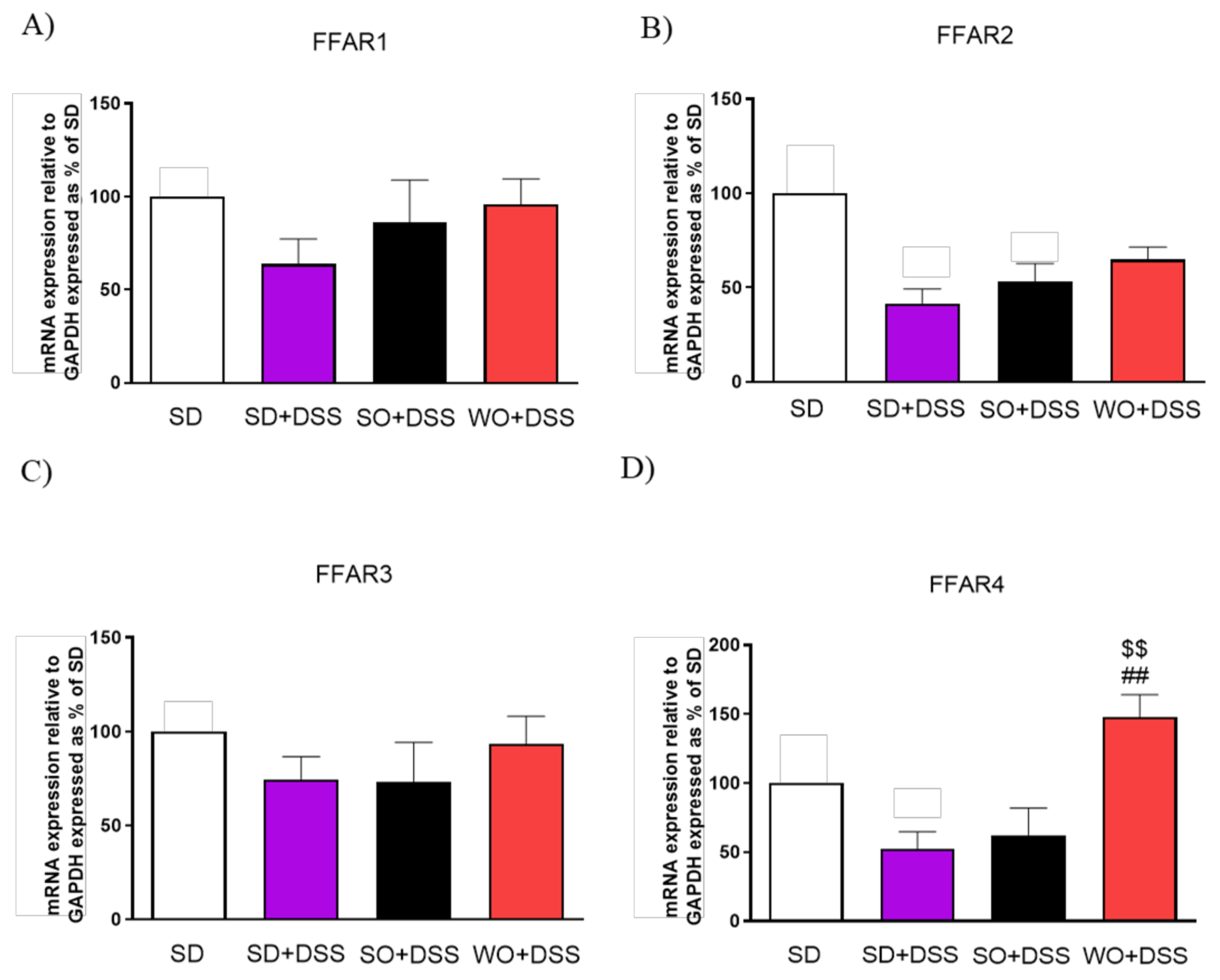
3.4. Regulation of TJs, FFARs, and Pro-Inflammatory Cytokines Gene Expression by the WO in the Mouse Colon
4. Discussion
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/kg Diet) | SD a | SD Supplemented with 7% Sunflower oil (SO) | SD Supplemented with 7% Walnut Oil (WO) |
|---|---|---|---|
| Corn starch | 397.486 | 397.486 | 397.486 |
| Casein | 200.00 | 200.00 | 200.00 |
| Maltodextrin | 132.00 | 132.00 | 132.00 |
| Sucrose | 100.00 | 100.00 | 100.00 |
| Soybean oil | 70.00 | 0.00 | 0.00 |
| Sunflower oil | 0.00 | 70.00 | 0.00 |
| Walnut oil | 0.00 | 0.00 | 70.00 |
| Fiber | 50.00 | 50.00 | 50.00 |
| aAIN93G Mineral mix | 35.00 | 35.00 | 35.00 |
| aAIN93G Vitamin mix | 10.00 | 10.00 | 10.00 |
| L-Cystine | 3.00 | 3.00 | 3.00 |
| Choline bitartrate | 2.50 | 2.50 | 2.50 |
| Tert-butylohydrochinon | 0.014 | 0.014 | 0.014 |
| 1000.0 | 1000.0 | 1000.0 |
References
- Sobczak, M.; Fabisiak, A.; Murawska, N.; Wesołowska, E.; Wierzbicka, P.; Wlazłowski, M.; Wójcikowska, M.; Zatorski, H.; Zwolińska, M.; Fichna, J. Current overview of extrinsic and intrinsic factors in etiology and progression of inflammatory bowel diseases. Pharmacol. Rep. 2014, 66, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Hvas, C.L.; Bendix, M.; Dige, A.; Dahlerup, J.F.; Agnholt, J. Current, experimental, and future treatments in inflammatory bowel disease: A clinical review. Immunopharmacol. Immunotoxicol. 2018, 40, 446–460. [Google Scholar] [CrossRef] [PubMed]
- Panaccione, R.; Underwood, F.E.; Ghosh, S.; Kaplan, G.G.; Hamidi, N.; Benchimol, E.I.; Chan, F.K.L.; Sung, J.J.Y.; Shi, H.Y.; Wu, J.C.Y.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar]
- Moran, C.J. Very early onset inflammatory bowel disease. Semin. Pediatr. Surg. 2017, 26, 356–359. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef]
- Scaioli, E.; Liverani, E.; Belluzzi, A. The Imbalance between n-6/n-3 Polyunsaturated Fatty Acids and Inflammatory Bowel Disease: A Comprehensive Review and Future Therapeutic Perspectives. Int. J. Mol. Sci. 2017, 18, 2619. [Google Scholar] [CrossRef]
- Vinolo, M.A.R.; Hirabara, S.M.; Curi, R. G-protein-coupled receptors as fat sensors. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 112–116. [Google Scholar] [CrossRef]
- Milligan, G.; Shimpukade, B.; Ulven, T.; Hudson, B.D. Complex pharmacology of free fatty acid receptors. Chem. Rev. 2017, 117, 67–110. [Google Scholar] [CrossRef]
- Alvarez-Curto, E.; Milligan, G. Metabolism meets immunity: The role of free fatty acid receptors in the immune system. Biochem. Pharmacol. 2016, 114, 3–13. [Google Scholar] [CrossRef]
- Bartoszek, A.; Moo, E.V.; Binienda, A.; Fabisiak, A.; Krajewska, J.B.; Mosińska, P.; Niewinna, K.; Tarasiuk, A.; Martemyanov, K.; Salaga, M.; et al. Free fatty acid receptors as new potential therapeutic target in inflammatory bowel diseases. Pharmacol. Res. 2020, 152, 104604. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Utsumi, D.; Matsumoto, K. G protein-coupled receptor 40 activation ameliorates dextran sulfate sodium-induced colitis in mice via the upregulation of glucagon-likepeptide-2. J. Pharmacol. Sci. 2019, 140, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Di, Y.; Schilter, H.C.; Rolph, M.S.; MacKay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Masui, R.; Sasaki, M.; Funaki, Y.; Ogasawara, N.; Mizuno, M.; Iida, A.; Izawa, S.; Kondo, Y.; Ito, Y.; Tamura, Y.; et al. G protein-coupled receptor 43 moderates gut inflammation through cytokine regulation from mononuclear cells. Inflamm. Bowel Dis. 2013, 19, 2848–2856. [Google Scholar] [CrossRef]
- Sina, C.; Gavrilova, O.; Forster, M.; Till, A.; Derer, S.; Hildebrand, F.; Raabe, B.; Chalaris, A.; Scheller, J.; Rehmann, A.; et al. G Protein-Coupled Receptor 43 Is Essential for Neutrophil Recruitment during Intestinal Inflammation. J. Immunol. 2009, 183, 7514–7522. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology 2013, 145, 396–406. [Google Scholar] [CrossRef]
- Calder, P.C. Polyunsaturated fatty acids, inflammatory processes and inflammatory bowel diseases. Mol. Nutr. Food Res. 2008, 52, 885–897. [Google Scholar] [CrossRef]
- Poggetti, L.; Ferfuia, C.; Chiabà, C.; Testolin, R.; Baldini, M. Kernel oil content and oil composition in walnut ( Juglans regia L.) accessions from north-eastern Italy. J. Sci. Food Agric. 2018, 98, 955–962. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Sobczak, M.; Mokrowiecka, A.; Cygankiewicz, A.I.; Zakrzewski, P.K.; Sałaga, M.; Storr, M.; Kordek, R.; Małecka-Panas, E.; Krajewska, W.M.; Fichna, J. Anti-inflammatory and antinociceptive action of an orally available nociceptin receptor agonist SCH 221510 in a mouse model of inflammatory bowel diseases. J. Pharmacol. Exp. Ther. 2014, 348, 401–409. [Google Scholar] [CrossRef]
- Fichna, J.; Dicay, M.; Lewellyn, K.; Janecka, A.; Zjawiony, J.K.; MacNaughton, W.K.; Storr, M.A. Salvinorin A has antiinflammatory and antinociceptive effects in experimental models of colitis in mice mediated by KOR and CB1 receptors. Inflamm. Bowel Dis. 2012, 18, 1137–1145. [Google Scholar] [CrossRef]
- Fichna, J.; Schicho, R.; Andrews, C.N.; Bashashati, M.; Klompus, M.; Mckay, D.M.; Sharkey, K.A.; Zjawiony, J.K.; Janecka, A.; Storr, M.A. Salvinorin A inhibits colonic transit and neurogenic ion transport in mice by activating κ-opioid and cannabinoid receptors. Neurogastroenterol. Motil. 2009, 21, 1326-e128. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Ishibashi, A.; Kani, K.; Yakabi, K. Optimized management of ulcerative proctitis: When and how to use mesalazine suppository. Digestion 2018, 97, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Cross, R.K. Safety considerations with the use of corticosteroids and biologic therapies in mild-to-moderate ulcerative colitis. Inflamm. Bowel. Dis. 2017, 23, 1689–1701. [Google Scholar] [CrossRef] [PubMed]
- Thorne, K.; Alrubaiy, L.; Akbari, A.; Samuel, D.G.; Morrison-Rees, S.; Roberts, S.E. Colectomy rates in patients with ulcerative colitis following treatment with infliximab or ciclosporin: A systematic literature review. Eur. J. Gastroenterol. Hepatol. 2016, 28, 369–382. [Google Scholar] [CrossRef]
- Khan, K.J.; Dubinsky, M.C.; Ford, A.C.; Ullman, T.A.; Talley, N.J.; Moayyedi, P. Efficacy of immunosuppressive therapy for inflammatory bowel disease: A systematic review and meta-analysis. Am. J. Gastroenterol. 2011, 106, 630–642. [Google Scholar] [CrossRef]
- Mosińska, P.; Tarasiuk, A.; Fabisiak, A.; Krajewska, J.; Niewinna, K.; Bartoszek, A.; Binienda, A.; Sałaga, M.; Fichna, J. Dietary fatty acid content influences the expression of genes involved in the lipid turnover and inflammation in mouse colon and spleen. Pharmacol. Reports 2019, 71, 899–908. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, H.; Shi, P.; Wang, W.; Sun, Y. GPR120, a potential therapeutic target for experimental colitis in IL-10 deficient mice. Oncotarget 2017, 8, 8397–8405. [Google Scholar] [CrossRef]
- Stenson, W.F.; Cort, D.; Rodgers, J.; Burakoff, R.; DeSchryver-Kecskemeti, K.; Gramlich, T.L.; Beeken, W. Dietary supplementation with fish oil in ulcerative colitis. Ann. Intern. Med. 1992, 116, 609. [Google Scholar] [CrossRef]
- Hawthorne, A.B.; Daneshmend, T.K.; Hawkey, C.J.; Belluzzi, A.; Everitt, S.J.; Holmes, G.K.; Malkinson, C.; Shaheen, M.Z.; Willars, J.E. Treatment of ulcerative colitis with fish oil supplementation: A prospective 12 month randomised controlled trial. Gut 1992, 33, 922–928. [Google Scholar] [CrossRef]
- Vilaseca, J.; Salas, A.; Guarner, F.; Rodríguez, R.; Martínez, M.; Malagelada, J.R. Dietary fish oil reduces progression of chronic inflammatory lesions in a rat model of granulomatous colitis. Gut 1990, 31, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Andoh, A.; Tsujikawa, T.; Ishizuka, I.; Araki, Y.; Sasaki, M.; Koyama, S.; Fujiyama, Y. N-3 fatty acid-rich diet prevents early response of interleukin-6 elevation in trinitrobenzene sulfonic acid-induced enteritis. Int. J. Mol. Med. 2003, 12, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Shoda, R.; Matsueda, K.; Yamato, S.; Umeda, N. Therapeutic efficacy of N-3 polyunsaturated fatty acid in experimental Crohn’s disease. J. Gastroenterol. 1995, 30, 98–101. [Google Scholar] [PubMed]
- Nieto, N.; Torres, M.I.; Ríos, A.; Gil, A. Dietary polyunsaturated fatty acids improve histological and biochemical alterations in rats with experimental ulcerative colitis. J. Nutr. 2002, 132, 11–19. [Google Scholar] [CrossRef]
- Yuceyar, H.; Ozutemiz, O.; Huseyinov, A.; Saruç, M.; Alkanat, M.; Bor, S.; Coker, I.; Batur, Y. Is administration of n-3 fatty acids by mucosal enema protective against trinitrobenzene-induced colitis in rats? Prostaglandins. Leukot. Essent. Fatty Acids 1999, 61, 339–345. [Google Scholar] [CrossRef]
- McMurray, D.N.; Ly, L.; Weeks, B.R.; Lupton, J.R.; Davidson, L.A.; Chapkin, R.S. Immunomodulatory effects of (n-3) fatty acids: Putative link to inflammation and colon cancer. J. Nutr. 2018, 137, 200S–204S. [Google Scholar]
- Lu, Y.; Serhan, C.N.; Hudert, C.A.; Weylandt, K.H.; Wang, J.; Hong, S.; Kang, J.X.; Dignass, A. Transgenic mice rich in endogenous omega-3 fatty acids are protected from colitis. Proc. Natl. Acad. Sci. USA 2006, 103, 11276–11281. [Google Scholar]
- Reifen, R.; Karlinsky, A.; Stark, A.H.; Berkovich, Z.; Nyska, A. α-Linolenic acid (ALA) is an anti-inflammatory agent in inflammatory bowel disease. J. Nutr. Biochem. 2015, 26, 1632–1640. [Google Scholar] [CrossRef]
- Wen, J.; Khan, I.; Li, A.; Chen, X.; Yang, P.; Song, P.; Jing, Y.; Wei, J.; Che, T.; Zhang, C. Alpha-linolenic acid given as an anti-inflammatory agent in a mouse model of colonic inflammation. Food Sci. Nutr. 2019, 7, 3873–3882. [Google Scholar] [CrossRef]
- Batirel, S.; Yilmaz, A.M.; Sahin, A.; Perakakis, N.; Kartal Ozer, N.; Mantzoros, C.S. Antitumor and antimetastatic effects of walnut oil in esophageal adenocarcinoma cells. Clin. Nutr. 2018, 37, 2166–2171. [Google Scholar] [CrossRef]
- Lundgren, O. 5-Hydroxytryptamine, enterotoxins, and intestinal fluid secretion. Gastroenterology 1998, 115, 1009–1012. [Google Scholar] [CrossRef]
- Wasilewski, A.; Misicka, A.; Sacharczuk, M.; Fichna, J. Modulation of the endocannabinoid system by fatty acid amide hydrolase, mono-and diacylglycerol lipase inhibitors as an attractive target for secretory diarrhea therapy. United Eur. Gastroenterol. J. 2017, 5, A94. [Google Scholar]
- Seamon, K.B.; Daly, J.W. Forskolin: A unique diterpene activator of cyclic AMP-generating systems. J. Cyclic Nucleotide Res. 1981, 7, 201–204. [Google Scholar] [PubMed]
- Yun, C.H.C.; Oh, S.; Zizak, M.; Steplock, D.; Tsao, S.; Tse, C.M.; Weinman, E.J.; Donowitz, M. cAMP-mediated inhibition of the epithelial brush border Na+/H+ exchanger, NHE3, requires an associated regulatory protein. Proc. Natl. Acad. Sci. USA 1997, 94, 3010–3015. [Google Scholar] [CrossRef]
- Barrett, K.E.; McCole, D.F. Hydrogen peroxide scavenger, catalase, alleviates ion transport dysfunction in murine colitis. Clin. Exp. Pharmacol. Physiol. 2016, 43, 1097–1106. [Google Scholar] [CrossRef]
- Catterall, W.A. Neurotoxins that act on voltage-sensitive sodium channels in excitable membranes. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 15–43. [Google Scholar] [CrossRef]
- Chen, C.; Shah, Y.M.; Morimura, K.; Krausz, K.W.; Miyazaki, M.; Richardson, T.A.; Morgan, E.T.; Ntambi, J.M.; Idle, J.R.; Gonzalez, F.J. Metabolomics reveals that Hhepatic stearoyl-CoA desaturase 1 downregulation exacerbates inflammation and acute colitis. Cell Metab. 2008, 7, 135–147. [Google Scholar] [CrossRef]
- Zeisel, M.B.; Dhawan, P.; Baumert, T.F. Tight junction proteins in gastrointestinal and liver disease. Gut 2019, 68, 547–561. [Google Scholar] [CrossRef]
- Pope, J.L.; Ahmad, R.; Bhat, A.A.; Washington, M.K.; Singh, A.B.; Dhawan, P. Claudin-1 overexpression in intestinal epithelial cells enhances susceptibility to adenamatous polyposis coli-mediated colon tumorigenesis. Mol. Cancer 2014, 13, 167. [Google Scholar] [CrossRef]
- Ahmad, R.; Chaturvedi, R.; Olivares-Villagómez, D.; Habib, T.; Asim, M.; Shivesh, P.; Polk, D.B.; Wilson, K.T.; Washington, M.K.; Van Kaer, L.; et al. Targeted colonic claudin-2 expression renders resistance to epithelial injury, induces immune suppression, and protects from colitis. Mucosal Immunol. 2014, 7, 1340–1353. [Google Scholar] [CrossRef]
- Ding, L.; Lu, Z.; Foreman, O.; Tatum, R.; Lu, Q.; Renegar, R.; Cao, J.; Chen, Y. Inflammation and disruption of the mucosal architecture in claudin-7-deficient mice. Gastroenterology 2012, 142, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.S.L.; Enck, A.H.; Lencer, W.I.; Schneeberger, E.E. Claudin-8 expression in Madin-Darby canine kidney cells augments the paracellular barrier to cation permeation. J. Biol. Chem. 2003, 278, 17350–17359. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Pridgen, T.A.; Blikslager, A.T. Pharmaceutical activation or genetic absence of ClC-2 alters tight junctions during experimental colitis. Inflam. Bowel Dis. 2015, 21, 2747–2757. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Q.; Xu, H.; Tao, L.; Lu, J.; Cai, L.; Wang, C. Somatostatin regulates tight junction proteins expression in colitis mice. Int. J. Clin. Exp. Pathol. 2014, 7, 2153–2162. [Google Scholar]
- Hara, T.; Kashihara, D.; Ichimura, A.; Kimura, I.; Tsujimoto, G.; Hirasawa, A. Role of free fatty acid receptors in the regulation of energy metabolism. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2014, 1841, 1292–1300. [Google Scholar] [CrossRef]
- Cheshmehkani, A.; Senatorov, I.S.; Kandi, P.; Singh, M.; Britt, A.; Hayslett, R.; Moniri, N.H. Fish oil and flax seed oil supplemented diets increase FFAR4 expression in the rat colon. Inflamm. Res. 2015, 64, 809–815. [Google Scholar] [CrossRef]
- Tsukahara, T.; Watanabe, K.; Watanabe, T.; Yamagami, H.; Sogawa, M.; Tanigawa, T.; Shiba, M.; Tominaga, K.; Fujiwara, Y.; Maeda, K.; et al. Tumor necrosis factor α decreases glucagon-like peptide-2 expression by up-regulating G-protein-coupled receptor 120 in crohn disease. Am. J. Pathol. 2015, 185, 185–196. [Google Scholar] [CrossRef]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free fatty acids regulate gut incretin glucagon-like peptide-1 secretion through GPR120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef]
- Anbazhagan, A.N.; Priyamvada, S.; Gujral, T.; Bhattacharyya, S.; Alrefai, W.A.; Dudeja, P.K.; Borthakur, A. A novel anti-inflammatory role of GPR120 in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2016, 310, C612–C621. [Google Scholar] [CrossRef]
- Hudson, B.D.; Shimpukade, B.; Mackenzie, A.E.; Butcher, A.J.; Pediani, J.D.; Christiansen, E.; Heathcote, H.; Tobin, A.B.; Ulven, T.; Milligan, G. The pharmacology of TUG-891, a potent and selective agonist of the free fatty acid receptor 4 (FFA4/GPR120), demonstrates both potential opportunity and possible challenges to therapeutic agonism. Mol. Pharmacol. 2013, 84, 710–725. [Google Scholar] [CrossRef]









| Fatty Acid Content (%) | Soybean Oil | Sunflower Oil | Walnut Oil |
|---|---|---|---|
| Palmitic acid C 16:0 | 7–10 | 2–9 | 6–8 |
| Stearic acid C 18:0 | 2–5 | 2–7 | 1–3.5 |
| Oleic acid C 18:1 | 22–30 | 75–91 | 14–21 |
| Linoleic acid C 18:2 | 50–60 | 3–17 | 54–65 |
| α-Linolenic acid C 18:3 | 2–5 | Max 0,3 | 9–16 |
| n-3/n-6 balance | 0.04–0.083 | 0.001–0.1 | 0.167–0.246 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartoszek, A.; Makaro, A.; Bartoszek, A.; Kordek, R.; Fichna, J.; Salaga, M. Walnut Oil Alleviates Intestinal Inflammation and Restores Intestinal Barrier Function in Mice. Nutrients 2020, 12, 1302. https://doi.org/10.3390/nu12051302
Bartoszek A, Makaro A, Bartoszek A, Kordek R, Fichna J, Salaga M. Walnut Oil Alleviates Intestinal Inflammation and Restores Intestinal Barrier Function in Mice. Nutrients. 2020; 12(5):1302. https://doi.org/10.3390/nu12051302
Chicago/Turabian StyleBartoszek, Adrian, Adam Makaro, Agnieszka Bartoszek, Radzisław Kordek, Jakub Fichna, and Maciej Salaga. 2020. "Walnut Oil Alleviates Intestinal Inflammation and Restores Intestinal Barrier Function in Mice" Nutrients 12, no. 5: 1302. https://doi.org/10.3390/nu12051302
APA StyleBartoszek, A., Makaro, A., Bartoszek, A., Kordek, R., Fichna, J., & Salaga, M. (2020). Walnut Oil Alleviates Intestinal Inflammation and Restores Intestinal Barrier Function in Mice. Nutrients, 12(5), 1302. https://doi.org/10.3390/nu12051302