Effect of the Lactococcus Lactis 11/19-B1 Strain on Atopic Dermatitis in a Clinical Test and Mouse Model


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
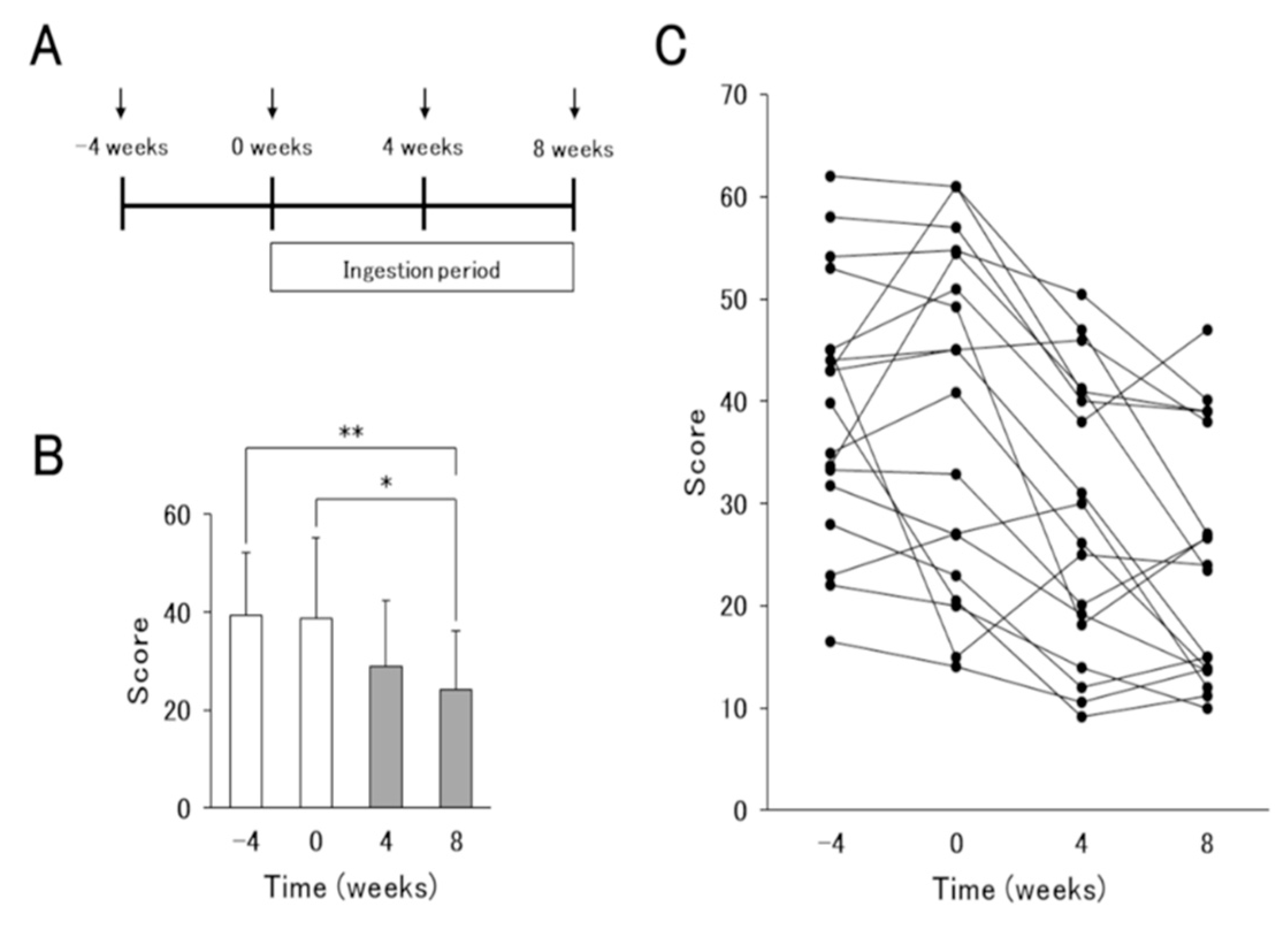
2.1. Subjects and Study Design
2.2. Yogurt
2.3. Animals and Diets
2.4. AD Mouse Model
2.5. Measurement of Total IgE in Plasma
2.6. Flow Cytometric Analysis (FACS)
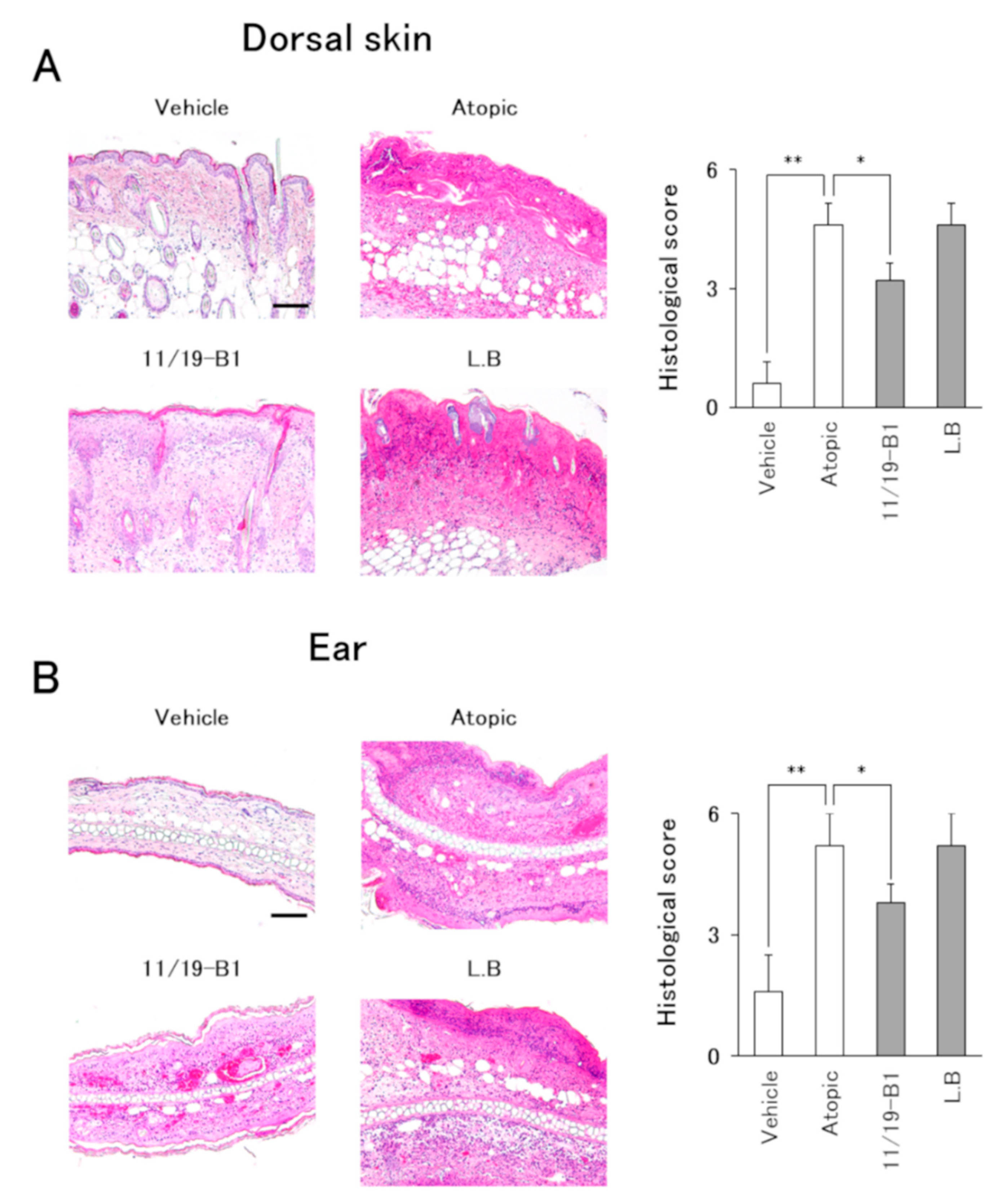
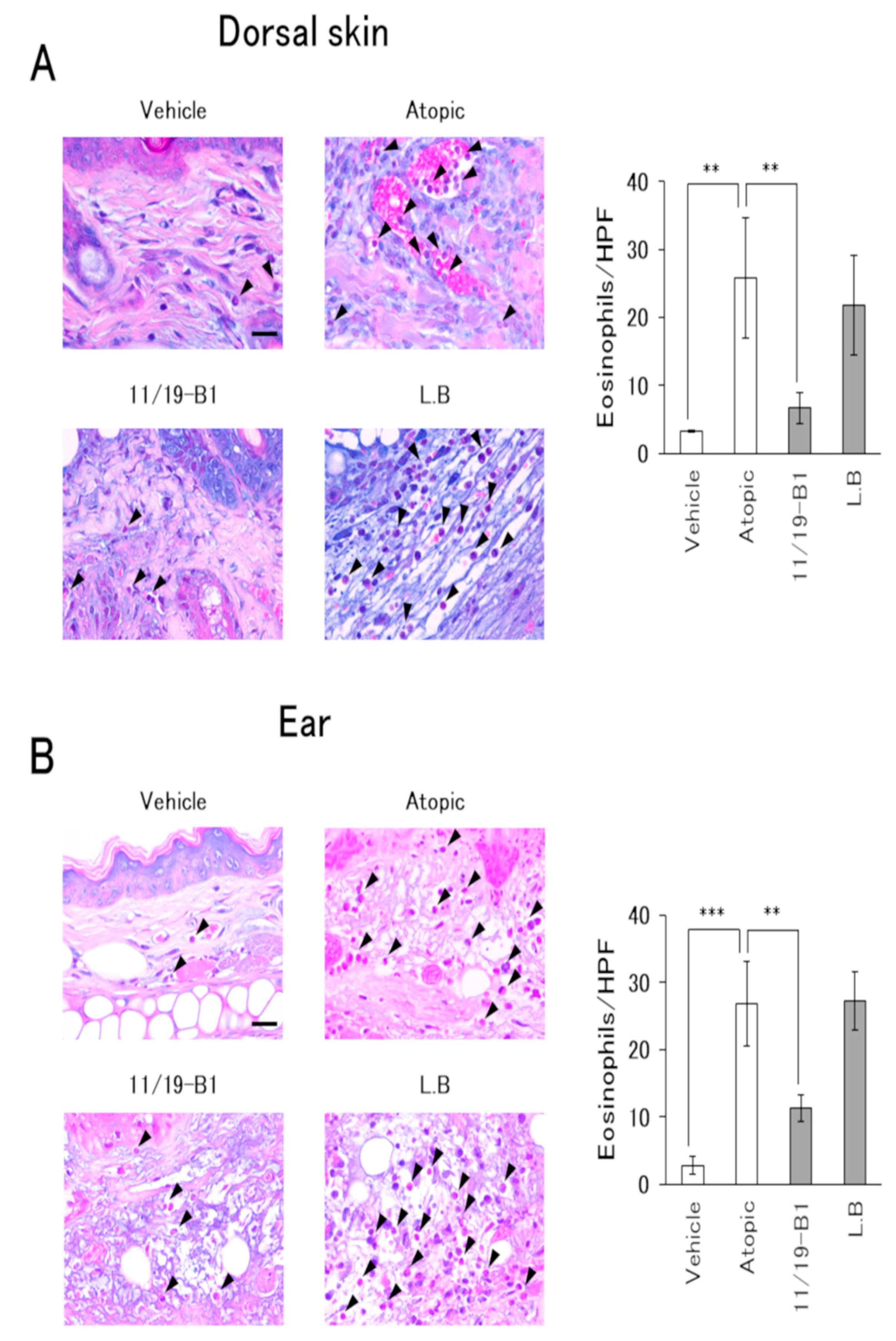
2.7. Histological Evaluation
2.8. Statistics
3. Results
3.1. Effect of L. Lactis 11/19-B1-Containing Yogurt Intake in AD Patients
3.2. Identification of LAB Strains Inducing Improvement in AD Pathology Using a Mouse Model
3.3. Altered Distribution of CD4+ T Cell Subsets with Ingestion of L. Lactis 11/19-B1
3.4. Histological Changes in AD Pathology Associated with the Ingestion of L. Lactis 11/19-B1
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saeki, H.; Furue, M.; Furukawa, F.; Hide, M.; Ohtsuki, M.; Katayama, I.; Sasaki, R.; Suto, H.; Takehara, K. Guidelines for management of atopic dermatitis. J. Dermatol. 2009, 36, 563–577. [Google Scholar] [CrossRef]
- Bieber, T. Atopic Dermatitis. N. Engl. J. Med. 2008, 358, 1483–1494. [Google Scholar] [CrossRef]
- Oyoshi, M.K.; He, R.; Kumar, L.; Yoon, J.; Geha, R.S. Cellular and Molecular Mechanisms in Atopic Dermatitis. Adv. Immunol. 2009, 102, 135–226. [Google Scholar] [PubMed]
- Koga, C.; Kabashima, K.; Shiraishi, N.; Kobayashi, M.; Tokura, Y. Possible pathogenic role of Th17 cells for atopic dermatitis. J. Investig. Dermatol. 2008, 128, 2625–2630. [Google Scholar] [CrossRef] [PubMed]
- Rizello, V.; Bonaccorsi, I.; Dongarra, M.L.; Fink, L.N.; Ferlazzo, G. Role of natural killer and dendritic cell crosstalk in immunomodulation by commensal bacteria probiotics. J. Biomed. Biotechnol. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-T.; Cheng, P.-C.; Pan, T.-M. The immunomodulatory effects of lactic acid bacteria for improving immune functions and benefits. Artic. Appl. Microbiol. Biotechnol. 2012, 96, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Pessi, T.; Su, Y.; Tas, È.; Hurme, M.; Isolauri, E. Interleukin-10 generation in atopic children following oral Lactobacillus rhamnosus GG. Clin. Exp. Allergy 2000, 30, 1804–1808. [Google Scholar] [CrossRef] [PubMed]
- Sawada, J.; Morita, H.; Tanaka, A.; Salminen, S.; He, F.; Matsuda, H. Ingestion of heat-treated Lactobacillus rhamnosus GG prevents development of atopic dermatitis in NC/Nga mice. Clin. Exp. Allergy 2007, 37, 296–303. [Google Scholar] [CrossRef]
- Fujiwara, D.; Inoue, S.; Wakabayashi, H.; Fujii, T. The Anti-Allergic Effects of Lactic Acid Bacteria Are Strain Dependent and Mediated by Effects on both Th1/Th2 Cytokine Expression and Balance. Int. Arch. Allergy Immunol. 2004, 135, 205–215. [Google Scholar] [CrossRef]
- Fujiwara, D.; Wakabayashi, H.; Watanabe, H.; Nishida, S.; Iino, H. A Double-blind Trial of Lactobacillus paracasei Strain KW3110 Administration for Immunomodulation in Patients with Pollen Allergy. Allergol. Int. 2005, 54, 143–149. [Google Scholar] [CrossRef]
- Nishida, S.; Ono, Y.; Sekimizu, K. Lactic acid bacteria activating innate immunity improve survival in bacterial infection model of silkworm. Drug Discov. Ther. 2016, 10, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Hamamoto, H.; Kamimura, M.; Sekimizu, K. Activation of the silkworm cytokine by bacterial and fungal cell wall components via a reactive oxygen species-triggered mechanism. J. Biol. Chem. 2008, 283, 2185–2191. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Kobayashi, T.; Sato, Y.; Watanabe, Y.; Kikuchi, R.; Kanno, R.; Koshizuka, T.; Miyazaki, N.; Ishioka, K.; Suzutani, T.; et al. A Double-Blind Controlled Study to Evaluate the Effects of Yogurt Enriched with Lactococcus lactis 11/19-B1 and Bifidobacterium lactis on Serum Low-Density Lipoprotein Level and Antigen-Specific Interferon-γ Releasing Ability. Nutrients 2018, 10, 1778. [Google Scholar] [CrossRef] [PubMed]
- Schallreuter, K.U. Severity scoring of atopic dermatitis: The SCORAD index. Consensus Report of the European Task Force on Atopic Dermatitis. Dermatology 1993, 186, 23–31. [Google Scholar]
- Hussain, Z.; Katas, H.; Mohd Amin, M.C.I.; Kumolosasi, E.; Sahudin, S. Downregulation of immunological mediators in 2,4-dinitrofluorobenzene-induced atopic dermatitis-like skin lesions by hydrocortisone-loaded chitosan nanoparticles. Int. J. Nanomed. 2014, 9, 5143–5156. [Google Scholar]
- Lee, J.H.; Lee, Y.S.; Lee, E.J.; Kim, T.Y. Capsiate Inhibits DNFB-Induced Atopic Dermatitis in NC/Nga Mice through Mast Cell and CD4+ T-Cell Inactivation. J. Investig. Dermatol. 2015, 135, 1977–1985. [Google Scholar] [CrossRef]
- Tamura, A.; Kumai, H.; Nakamichi, N.; Sugiyama, T.; Deguchi, R.; Takagi, A.; Koga, Y. Suppression of Helicobacter pylori-induced interleukin-8 production in vitro and within the gastric mucosa by a live Lactobacillus strain. J. Gastroenterol. Hepatol. 2006, 21, 1399–1406. [Google Scholar] [CrossRef]
- Kabir, A.M.; Aiba, Y.; Takagi, A.; Kamiya, S.; Miwa, T.; Koga, Y. Prevention of Helicobacter pylori infection by lactobacilli in a gnotobiotic murine model. Gut 1997, 41, 49–55. [Google Scholar] [CrossRef]
- Jungersen, M.; Wind, A.; Johansen, E.; Christensen, J.; Stuer-Lauridsen, B.; Eskesen, D.; Jungersen, M.; Wind, A.; Johansen, E.; Christensen, J.E.; et al. The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®. Microorganisms 2014, 2, 92–110. [Google Scholar] [CrossRef]
- De Vrese, M.; Winkler, P.; Rautenberg, P.; Harder, T.; Noah, C.; Laue, C.; Ott, S.; Hampe, J.; Schreiber, S.; Heller, K.; et al. Probiotic bacteria reduced duration and severity but not the incidence of common cold episodes in a double blind, randomized, controlled trial. Vaccine 2006, 24, 6670–6674. [Google Scholar] [CrossRef]
- Viljanen, M.; Pohjavuori, E.; Haahtela, T.; Korpela, R.; Kuitunen, M.; Sarnesto, A.; Vaarala, O.; Savilahti, E. Induction of inflammation as a possible mechanism of probiotic effect in atopic eczema–dermatitis syndrome. J. Allergy Clin. Immunol. 2005, 115, 1254–1259. [Google Scholar] [CrossRef] [PubMed]
- Vanderpool, C.; Yan, F.; Polk, B.D. Mechanisms of probiotic action: Implications for therapeutic applications in inflammatory bowel diseases. Inflamm. Bowel Dis. 2008, 14, 1585–1596. [Google Scholar] [CrossRef] [PubMed]
- Liong, M.-T. Roles of Probiotics and Prebiotics in Colon Cancer Prevention: Postulated Mechanisms and In-vivo Evidence. Int. J. Mol. Sci. 2008, 9, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, K.; Yamasaki, T.; Akira, S.; Gantner, F.; Bacon, K.B. Essential role of MHC II-independent CD4 + T cells, IL-4 and STAT6 in contact hypersensitivity induced by fluorescein isothiocyanate in the mouse. Int. Immunol. 2004, 16, 685–695. [Google Scholar] [CrossRef]
- Grewe, M.; Bruijnzeel-Koomen, C.A.F.M.; Schöpf, E.; Thepen, T.; Langeveld-Wildschut, A.G.; Ruzicka, T.; Krutmann, J. A role for Th1 and Th2 cells in the immunopathogenesis of atopic dermatitis. Immunol. Today 1998, 19, 359–361. [Google Scholar] [CrossRef]
- Kitagaki, H.; Ono, N.; Hayakawa, K.; Kitazawa, T.; Watanabe, K.; Shiohara, T. Repeated Elicitation of Contact Hypersensitivity Induces a Shift in Cutaneous Cytokine Milieu from a T Helper Cell Type 1 to a T Helper Cell Type 2 Profile’. J. Immunol. 1997, 159, 2484–2491. [Google Scholar]
- Inagaki, N.; Shiraishi, N.; Igeta, K.; Nagao, M.; Kim, J.F.; Chikumoto, T.; Itoh, T.; Katoh, H.; Tanaka, H.; Nagai, H. Depletion of substance P, a mechanism for inhibition of mouse scratching behavior by tacrolimus. Eur. J. Pharmacol. 2010, 626, 283–289. [Google Scholar] [CrossRef]
- Man, M.-Q.; Hatano, Y.; Lee, S.H.; Man, M.; Chang, S.; Feingold, K.R.; Leung, D.Y.M.; Holleran, W.; Uchida, Y.; Elias, P.M. Characterization of a Hapten-Induced, Murine Model with Multiple Features of Atopic Dermatitis: Structural, Immunologic, and Biochemical Changes following Single Versus Multiple Oxazolone Challenges. J. Investig. Dermatol. 2008, 128, 79–86. [Google Scholar] [CrossRef]
- Nagai, H. Immunopharmacological Approach to Elucidating the Mechanism of Allergic Inflammation. Allergol. Int. 2005, 54, 251–261. [Google Scholar] [CrossRef]
- Sareneva, T.; Matikainen, S.; Kurimoto, M.; Julkunen, I. Influenza A Virus-Induced IFN-alpha/beat and IL-18 Synergistically Enhance IFN-Gene Expression in Human T Cells 1. J. Immunol. 1998, 160, 6032–6038. [Google Scholar]
- Pène, J.; Rousset, F.; Brière, F.; Chrétien, I.; Bonnefoy, J.Y.; Spits, H.; Yokota, T.; Arai, N.; Arai, K.; Banchereau, J. IgE production by normal human lymphocytes is induced by interleukin 4 and suppressed by interferons gamma and alpha and prostaglandin E2. Proc. Natl. Acad. Sci. USA 1988, 85, 6880–6884. [Google Scholar] [CrossRef] [PubMed]
- Jounai, K.; Ikado, K.; Sugimura, T.; Ano, Y.; Braun, J.; Fujiwara, D. Spherical Lactic Acid Bacteria Activate Plasmacytoid Dendritic Cells Immunomodulatory Function via TLR9-Dependent Crosstalk with Myeloid Dendritic Cells. PLoS ONE 2012, 7, e32588. [Google Scholar] [CrossRef] [PubMed]
- Guy-Grand, D.; Griscelli, C.; Vassalli, P. The mouse gut T lymphocyte, a novel type of T cell: Nature, origin, and traffic in mice in normal and graft-versus-host conditions. J. Exp. Med. 1978, 148, 1661–1677. [Google Scholar] [CrossRef] [PubMed]
- Ruedl, C.; Albin, B.; Böck, G.; Wick, G.; Wolf, H. Oral administration of a bacterial immunomodulator enhances murine intestinal lamina propria and Peyer’s patch lymphocyte traffic to the lung: Possible implications for infectious disease prophylaxis and therapy. Int. Immunol. 1993, 5, 29–36. [Google Scholar] [CrossRef]
- Lim, L.H.; Li, H.Y.; Huang, C.H.; Lee, B.W.; Lee, Y.K.; Chua, K.Y. The Effects of Heat-Killed Wild-Type Lactobacillus casei Shirota on Allergic Immune Responses in an Allergy Mouse Model. Int. Arch. Allergy Immunol. 2009, 148, 297–304. [Google Scholar] [CrossRef]
- Choi, E.-J.; Iwasa, M.; Han, K.-I.; Kim, W.-J.; Tang, Y.; Hwang, Y.; Chae, J.; Han, W.; Shin, Y.-S.; Kim, E.-K. Heat-Killed Enterococcus faecalis EF-2001 Ameliorates Atopic Dermatitis in a Murine Model. Nutrients 2016, 8, 146. [Google Scholar] [CrossRef]
- Laske, N.; Niggemann, B. Does the severity of atopic dermatitis correlate with serum IgE levels? Pediatr. Allergy Immunol. 2004, 15, 86–88. [Google Scholar] [CrossRef]
- Schaerli, P.; Willimann, K.; Lang, A.B.; Lipp, M.; Loetscher, P.; Moser, B. Cxc Chemokine Receptor 5 Expression Defines Follicular Homing T Cells with B Cell Helper Function. J. Exp. Med. 2000, 192, 1553–1562. [Google Scholar] [CrossRef]
- Smith, K.; Nossal, G.; Tarlinton, D.; Ueno, H. FAS is highly expressed in the germinal center but is not required for regulation of the B-cell response to antigen. Proc. Natl. Acad. Sci. USA 2011, 92, 11628–11632. [Google Scholar] [CrossRef]
- Morita, R.; Schmitt, N.; Bentebibel, S.-E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human Blood CXCR5+CD4+ T Cells Are Counterparts of T Follicular Cells and Contain Specific Subsets that Differentially Support Antibody Secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef]
- Vijayanand, P.; Seumois, G.; Simpson, L.J.; Abdul-Wajid, S.; Baumjohann, D.; Panduro, M.; Huang, X.; Interlandi, J.; Djuretic, I.M.; Brown, D.R.; et al. Interleukin-4 Production by Follicular Helper T Cells Requires the Conserved Il4 Enhancer Hypersensitivity Site V. Immunity 2012, 36, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Harada, Y.; Tanaka, S.; Motomura, Y.; Harada, Y.; Ohno, S.I.; Ohno, S.; Yanagi, Y.; Inoue, H.; Kubo, M. The 3′ Enhancer CNS2 Is a Critical Regulator of Interleukin-4-Mediated Humoral Immunity in Follicular Helper T Cells. Immunity 2012, 36, 188–200. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Atopic Group | |
|---|---|
| N | 18 |
| Age | 8.4 ± 3.6 |
| Sex (% of male) | 66.7 |
| SCORAD index | 38.2 ± 16.4 |
| IgE (mg/dL) | 1757.4 ± 1724.3 |
| TARC (pg/mL) | 1246.0 ± 1206.0 |
| Treatment (%) | |
| Tropical steroid | 100 |
| Heparinoid | 83.3 |
| Antihistamines | 55.6 |
| Tacrolims | 16.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, T.; Nishiyama, K.; Kawata, K.; Sugimoto, K.; Isome, M.; Suzuki, S.; Nozawa, R.; Ichikawa, Y.; Watanabe, Y.; Suzutani, T. Effect of the Lactococcus Lactis 11/19-B1 Strain on Atopic Dermatitis in a Clinical Test and Mouse Model. Nutrients 2020, 12, 763. https://doi.org/10.3390/nu12030763
Suzuki T, Nishiyama K, Kawata K, Sugimoto K, Isome M, Suzuki S, Nozawa R, Ichikawa Y, Watanabe Y, Suzutani T. Effect of the Lactococcus Lactis 11/19-B1 Strain on Atopic Dermatitis in a Clinical Test and Mouse Model. Nutrients. 2020; 12(3):763. https://doi.org/10.3390/nu12030763
Chicago/Turabian StyleSuzuki, Takato, Kyoko Nishiyama, Koji Kawata, Kotaro Sugimoto, Masato Isome, Shigeo Suzuki, Ruriko Nozawa, Yoko Ichikawa, Yoshihisa Watanabe, and Tatsuo Suzutani. 2020. "Effect of the Lactococcus Lactis 11/19-B1 Strain on Atopic Dermatitis in a Clinical Test and Mouse Model" Nutrients 12, no. 3: 763. https://doi.org/10.3390/nu12030763
APA StyleSuzuki, T., Nishiyama, K., Kawata, K., Sugimoto, K., Isome, M., Suzuki, S., Nozawa, R., Ichikawa, Y., Watanabe, Y., & Suzutani, T. (2020). Effect of the Lactococcus Lactis 11/19-B1 Strain on Atopic Dermatitis in a Clinical Test and Mouse Model. Nutrients, 12(3), 763. https://doi.org/10.3390/nu12030763