A Ten-Day Grape Seed Procyanidin Treatment Prevents Certain Ageing Processes in Female Rats over the Long Term

 , , , ,
, , , ,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Proanthocyanidin Extract
2.2. Animal Model
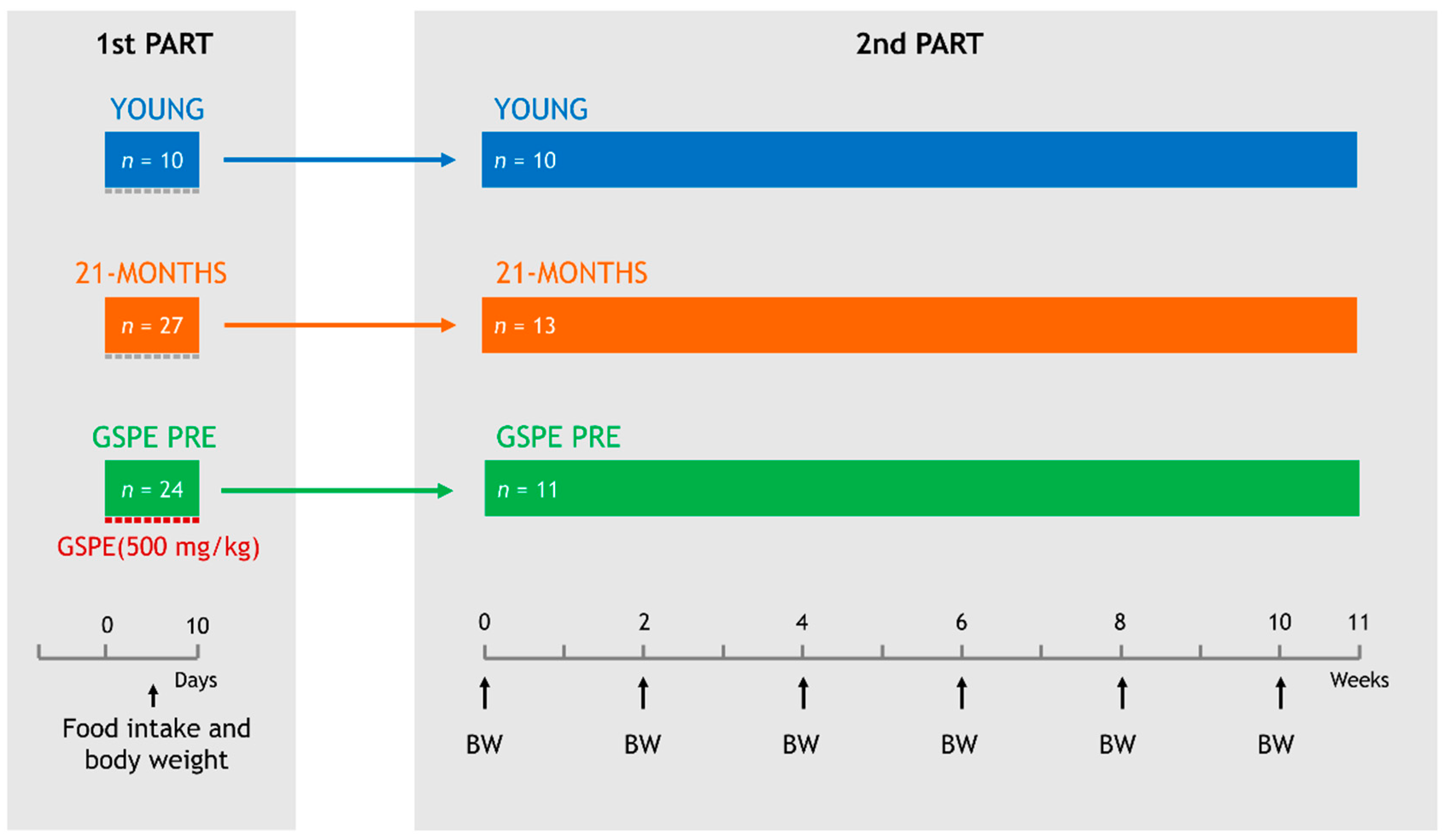
2.3. Experimental Design
- (1)
- YOUNG, which consisted of 10 two-month-old rats.
- (2)
- 21-MONTHS, which consisted of 27 twenty-one-month-old rats.
- (3)
- GSPE PRE, which consisted of 24 twenty-one-month-old rats.
2.4. Blood and Tissue Collection
2.5. Biochemical Variables
2.6. Statistical Analysis
3. Results
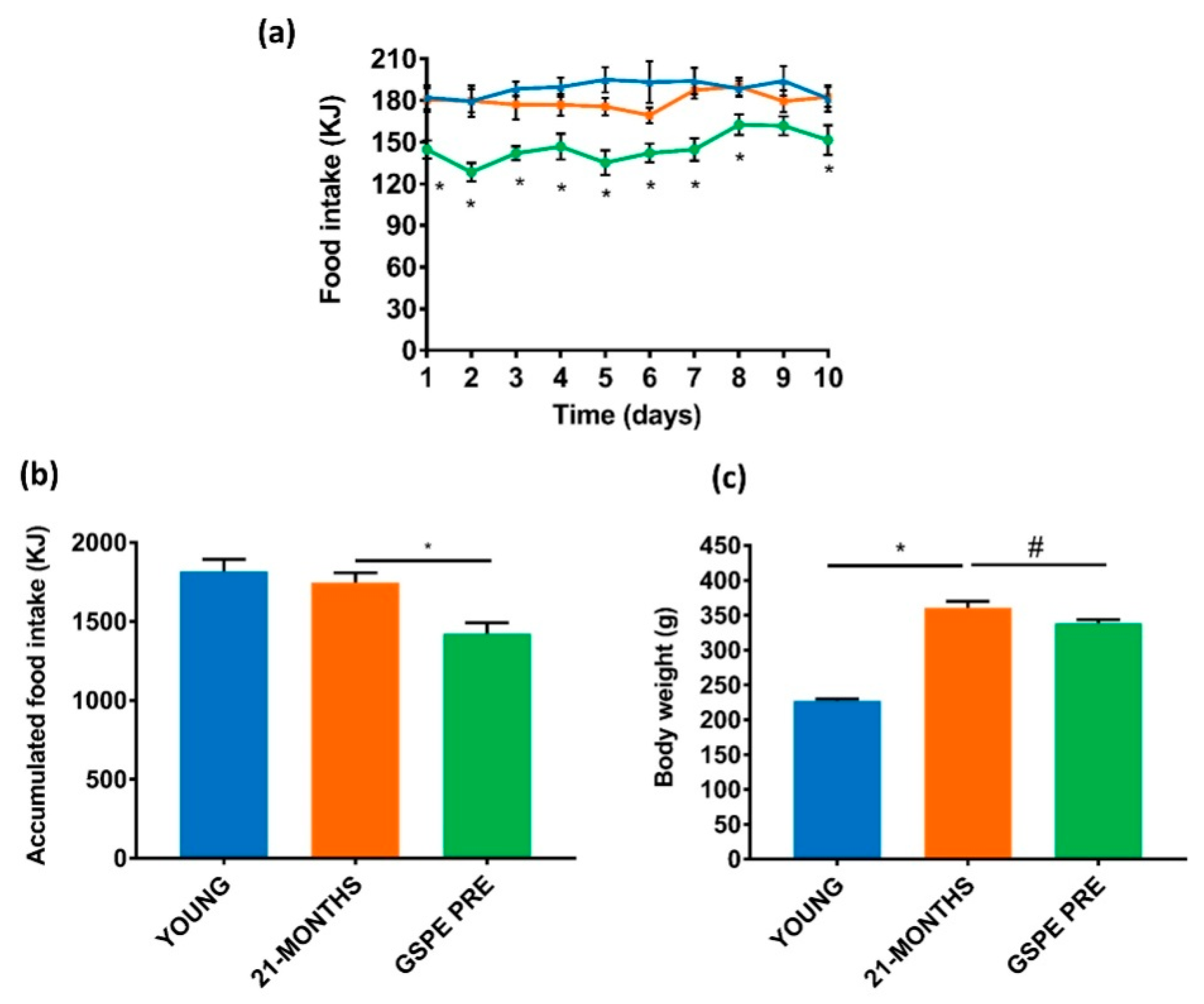
3.1. GSPE Reduces Food Intake and Body Weight in the Short-Term in Aged Rats
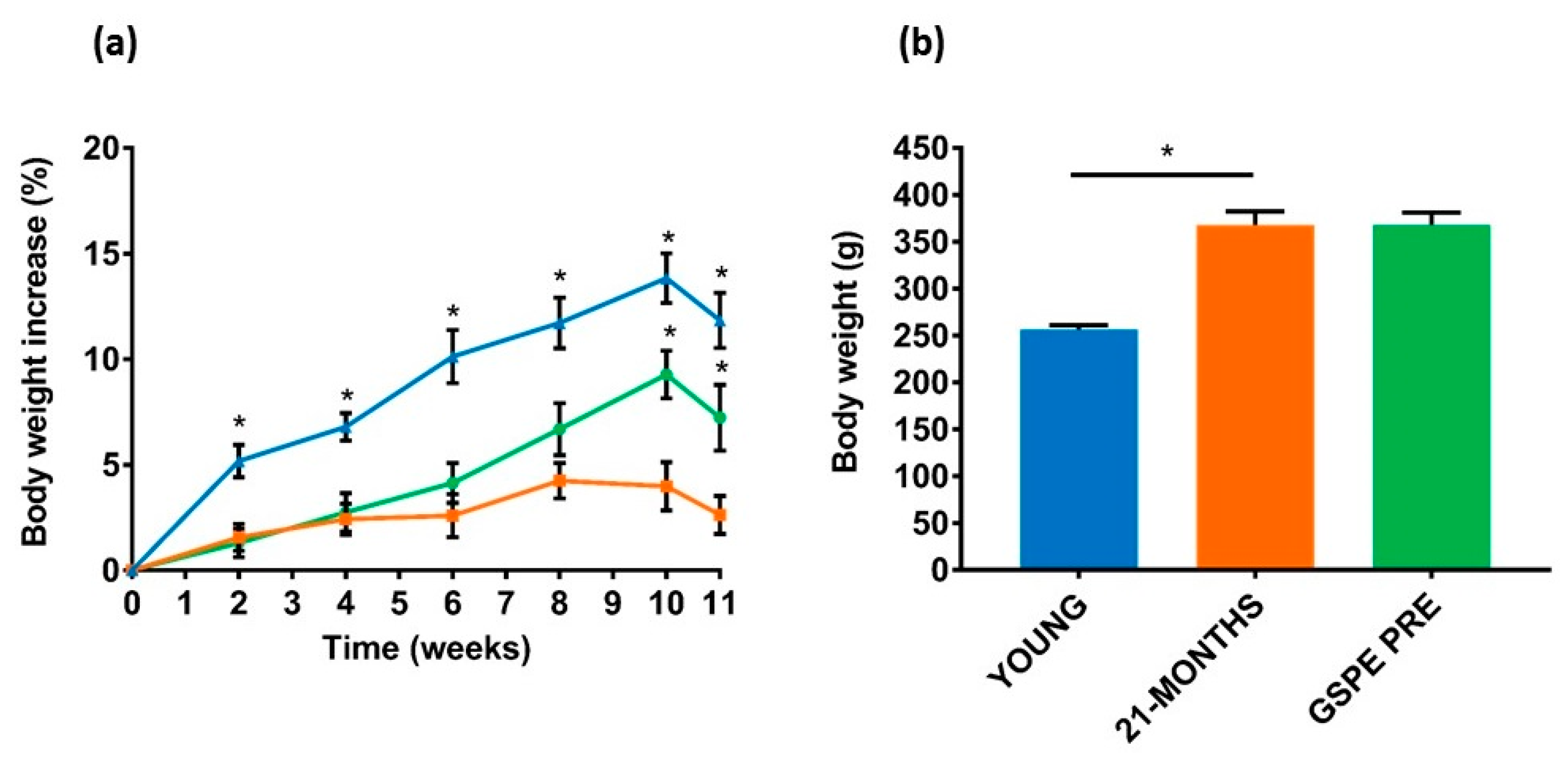
3.2. The GSPE Effect on Body Weight and Adiposity Continued for Several Weeks after Administration
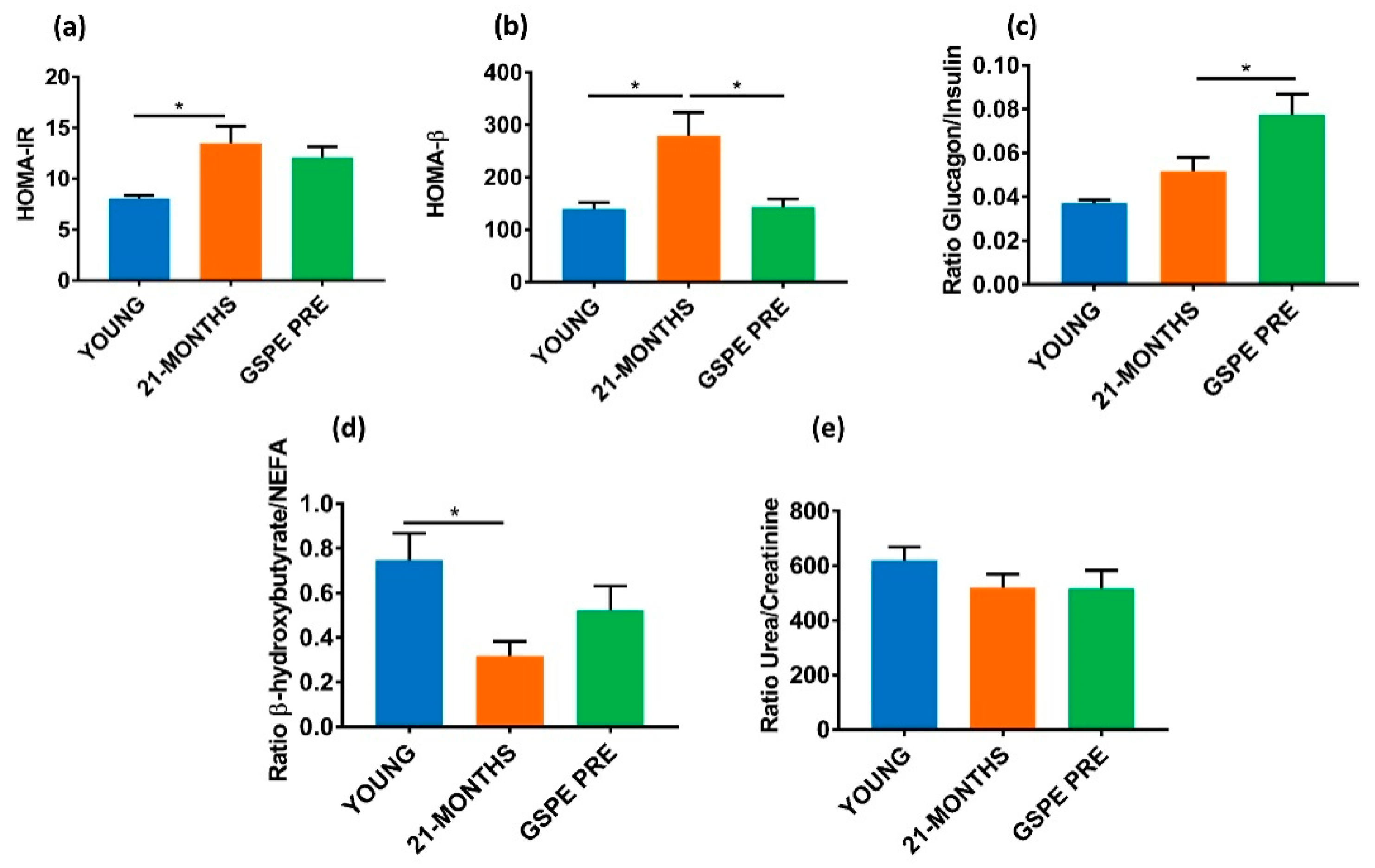
3.3. Aged GSPE Pre-Treated Rats Showed a Higher Fasting Glucagon/Insulin Ratio Eleven Weeks after the Treatment
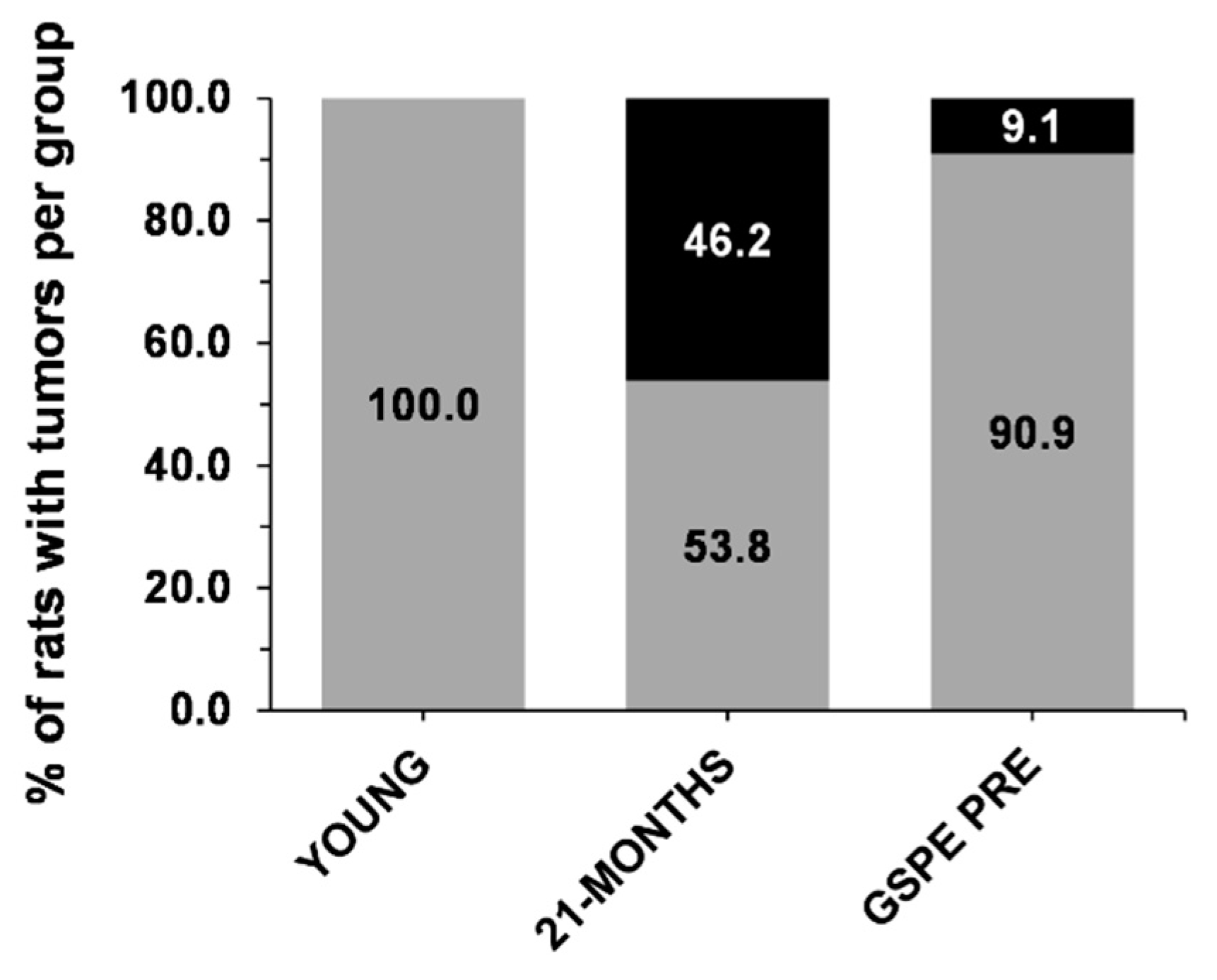
3.4. GSPE Limits the Development of Tumors in 21-MONTHS Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pomatto, L.C.D.; Davies, K.J.A. The role of declining adaptive homeostasis in ageing. J. Physiol. 2017, 595, 7275–7309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campisi, J. Aging, Cellular Senescence, and Cancer. Annu. Rev. Physiol. 2013, 75, 685–705. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Garagnani, P.; Morsiani, C.; Conte, M.; Santoro, A.; Grignolio, A.; Monti, D.; Capri, M.; Salvioli, S. The continuum of aging and age-related diseases: Common mechanisms but different rates. Front. Med. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elahi, D.; Muller, D.C. Carbohydrate metabolism in the elderly. Eur. J. Clin. Nutr. 2000, 54 (Suppl. 3), S112–S120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chia, C.W.; Egan, J.M.; Ferrucci, L. Age-Related Changes in Glucose Metabolism, Hyperglycemia, and Cardiovascular Risk. Circ. Res. 2018, 123, 886–904. [Google Scholar] [CrossRef]
- Shuster, A.; Patlas, M.; Pinthus, J.H.; Mourtzakis, M. The clinical importance of visceral adiposity: A critical review of methods for visceral adipose tissue analysis. Br. J. Radiol. 2012, 85, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Simonson, D.C.; DeFronzo, R.A. Glucagon physiology and aging: Evidence for enhanced hepatic sensitivity. Diabetologia 1983, 25, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bladé, C.; Aragones, G.; Arola-Arnal, A.; Muguerza, B.; Bravo, F.I.; Salvadó, M.J.; Arola, L.; Suárez, M. Proanthocyanidins in Health and Disease. Biofactors 2016, 42, 5–12. [Google Scholar]
- Mirza-Aghazadeh-Attari, M.; Ekrami, E.M.; Aghdas, S.A.M.; Mihanfar, A.; Hallaj, S.; Yousefi, B.; Safa, A.; Majidinia, M. Targeting PI3K/Akt/mTOR signaling pathway by polyphenols: Implication for cancer therapy. Life Sci. 2020, 255, 117481. [Google Scholar] [CrossRef]
- Tasatargil, A.; Tanriover, G.; Barutcigil, A.; Turkmen, E. Protective effect of resveratrol on methylglyoxal-induced endothelial dysfunction in aged rats. Aging Clin. Exp. Res. 2019, 31, 331–338. [Google Scholar] [CrossRef]
- Dehghani, A.; Hafizibarjin, Z.; Najjari, R.; Kaseb, F.; Safari, F. Resveratrol and 1,25-dihydroxyvitamin D co-administration protects the heart against d-galactose-induced aging in rats: Evaluation of serum and cardiac levels of klotho. Aging Clin. Exp. Res. 2019, 31, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Masodsai, K.; Lin, Y.-Y.; Chaunchaiyakul, R.; Su, C.-T.; Lee, S.-D.; Yang, A.-L. Twelve-Week Protocatechuic Acid Administration Improves Insulin-Induced and Insulin-Like Growth Factor-1-Induced Vasorelaxation and Antioxidant Activities in Aging Spontaneously Hypertensive Rats. Nutrients 2019, 11, 699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annunziata, G.; Jimenez-García, M.; Tejada, S.; Moranta, D.; Arnone, A.; Ciampaglia, R.; Tenore, G.C.; Sureda, A.; Novellino, E.; Capó, X. Grape Polyphenols Ameliorate Muscle Decline Reducing Oxidative Stress and Oxidative Damage in Aged Rats. Nutrients 2020, 12, 1280. [Google Scholar] [CrossRef]
- Meador, B.M.; Mirza, K.A.; Tian, M.; Skelding, M.B.; Reaves, L.A.; Edens, N.K.; Tisdale, M.J.; Pereira, S.L. The Green Tea Polyphenol Epigallocatechin-3-Gallate (EGCg) Attenuates Skeletal Muscle Atrophy in a Rat Model of Sarcopenia. J. Frailty Aging 2015, 4, 209. [Google Scholar] [PubMed]
- Sarubbo, F.; Ramis, M.R.; Aparicio, S.; Ruiz, L.; Esteban, S.; Miralles, A.; Moranta, D. Improving effect of chronic resveratrol treatment on central monoamine synthesis and cognition in aged rats. Age 2015, 37, 9777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhijit, S.; Subramanyam, M.V.V.; Devi, S.A. Grape Seed Proanthocyanidin and Swimming Exercise Protects Against Cognitive Decline: A Study on M1 Acetylcholine Receptors in Aging Male Rat Brain. Neurochem. Res. 2017, 42, 3573–3586. [Google Scholar] [CrossRef]
- Abhijit, S.; Tripathi, S.J.; Bhagya, V.; Shankaranarayana Rao, B.S.; Subramanyam, M.V.; Asha Devi, S. Antioxidant action of grape seed polyphenols and aerobic exercise in improving neuronal number in the hippocampus is associated with decrease in lipid peroxidation and hydrogen peroxide in adult and middle-aged rats. Exp. Gerontol. 2018, 101, 101–112. [Google Scholar] [CrossRef]
- Ksiezak-Reding, H.; Ho, L.; Santa-Maria, I.; Diaz-Ruiz, C.; Wang, J.; Pasinetti, G.M. Ultrastructural alterations of Alzheimer’s disease paired helical filaments by grape seed-derived polyphenols. Neurobiol. Aging 2012, 33, 1427–1439. [Google Scholar] [CrossRef]
- Santa-Maria, I.; Diaz-Ruiz, C.; Ksiezak-Reding, H.; Chen, A.; Ho, L.; Wang, J.; Pasinetti, G.M. GSPE interferes with tau aggregation in vivo: Implication for treating tauopathy. Neurobiol. Aging 2012, 33, 2072–2081. [Google Scholar] [CrossRef] [Green Version]
- Asha Devi, S.; Sagar Chandrasekar, B.K.; Manjula, K.R.; Ishii, N. Grape seed proanthocyanidin lowers brain oxidative stress in adult and middle-aged rats. Exp. Gerontol. 2011, 46, 958–964. [Google Scholar] [CrossRef] [Green Version]
- Graef, J.L.; Ouyang, P.; Wang, Y.; Rendina-Ruedy, E.; Lerner, M.R.; Marlow, D.; Lucas, E.A.; Smith, B.J. Dried plum polyphenolic extract combined with vitamin K and potassium restores trabecular and cortical bone in osteopenic model of postmenopausal bone loss. J. Funct. Foods 2018, 42, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.L.; Smith, B.J.; Li, J.; Cao, J.J.; Song, X.; Newhardt, M.F.; Corry, K.A.; Tomison, M.D.; Tang, L.; Wang, J.S.; et al. Effect of Long-Term Green Tea Polyphenol Supplementation on Bone Architecture, Turnover, and Mechanical Properties in Middle-Aged Ovariectomized Rats. Calcif. Tissue Int. 2019, 104, 285–300. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Bodakhe, S.H. Resveratrol delay the cataract formation against naphthalene-induced experimental cataract in the albino rats. J. Biochem. Mol. Toxicol. 2020, 34, e22420. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Pérez, C.; García-Villanova, B.; Guerra-Hernández, E.; Verardo, V. Grape seeds proanthocyanidins: An overview of in vivo bioactivity in animal models. Nutrients 2019, 11, 2435. [Google Scholar]
- Serrano, J.; Casanova-Martí, À.; Gual, A.; Pérez-Vendrell, A.M.; Blay, M.T.; Terra, X.; Ardévol, A.; Pinent, M. A specific dose of grape seed-derived proanthocyanidins to inhibit body weight gain limits food intake and increases energy expenditure in rats. Eur. J. Nutr. 2017, 56, 1629–1636. [Google Scholar] [CrossRef]
- Gonzalez-Abuin, N.; Pinent, M.; Casanova-Marti, A.; Arola, L.L.; Blay, M.; Ardevol, A.; González-Abuín, N.; Pinent, M.; Casanova-Martí, A.; Arola, L.L.; et al. Procyanidins and their healthy protective effects against type 2 diabetes. Curr. Med. Chem. 2015, 22, 39–50. [Google Scholar] [CrossRef]
- Quesada, H.; Pajuelo, D.; Fernández-Iglesias, A.; Díaz, S.; Ardevol, A.; Blay, M.; Salvadó, M.J.; Arola, L.; Blade, C. Proanthocyanidins modulate triglyceride secretion by repressing the expression of long chain acyl-CoA synthetases in Caco2 intestinal cells. Food Chem. 2011, 129, 1490–1494. [Google Scholar] [CrossRef]
- Li, Y.; Bao, L.; Zhang, Z.; Dai, X.; Ding, Y.; Jiang, Y.; Li, Y. Effects of grape seed proanthocyanidin extract on renal injury in type 2 diabetic rats. Mol. Med. Rep. 2015, 11, 645–652. [Google Scholar]
- Ma, J.; Fang, B.; Zeng, F.; Pang, H.; Ma, C.; Xia, J. Grape seed proanthocyanidins extract inhibits pancreatic cancer cell growth through down-regulation of miR-27a expression. J. Cent. South Univ. Med. Sci. 2015, 40, 46–52. [Google Scholar]
- Serrano, J.; Casanova-Martí, À.; Gil-Cardoso, K.; Blay, M.T.; Terra, X.; Pinent, M.; Ardévol, A. Acutely administered grape-seed proanthocyanidin extract acts as a satiating agent. Food Funct. 2016, 7, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Gil-Cardoso, K.; Ginés, I.; Pinent, M.; Ardévol, A.; Blay, M.; Terra, X. The co-administration of proanthocyanidins and an obesogenic diet prevents the increase in intestinal permeability and metabolic endotoxemia derived to the diet. J. Nutr. Biochem. 2018, 62, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Ginés, I.; Gil-Cardoso, K.; Terra, X.; Blay, M.; Pérez-Vendrell, A.M.; Pinent, M.; Ardévol, A. Grape Seed Proanthocyanidins Target the Enteroendocrine System in Cafeteria-Diet-Fed Rats. Mol. Nutr. Food Res. 2019, 63, 1800912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginés, I.; Gil-Cardoso, K.; Serrano, J.; Casanova-Martí, À.; Blay, M.; Pinent, M.; Ardévol, A.; Terra, X. Effects of an Intermittent Grape-Seed Proanthocyanidin (GSPE) Treatment on a Cafeteria Diet Obesogenic Challenge in Rats. Nutrients 2018, 10, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginés, I.; Gil-cardoso, K.; Serrano, J.; Casanova-marti, À.; Lobato, M.; Terra, X.; Blay, M.T.; Ard, A. Proanthocyanidins Limit Adipose Accrual Induced by a Cafeteria Diet, Several Weeks after the End of the Treatment. Genes 2019, 10, 598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, S.J.; Madrigal-Matute, J.; Scheibye-Knudsen, M.; Fang, E.; Aon, M.; González-reyes, J.A.; Cortassa, S.; Kaushik, S.; Gonzalez-Freire, M.; Patel, B.; et al. Effects of sex, strain, and energy intake on hallmarks of aging in mice. Cell Metab. 2016, 23, 1093–1112. [Google Scholar] [CrossRef] [Green Version]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194. [Google Scholar] [CrossRef] [Green Version]
- Ginés, I.; Gil-Cardoso, K.; D’addario, C.; Falconi, A.; Bellia, F.; Blay, M.T.; Terra, X.; Ardévol, A.; Pinent, M.; Beltrán-Debón, R. Long-lasting effects of gspe on ileal GLP-1R gene expression are associated with a hypomethylation of the GLP-1R promoter in female wistar rats. Biomolecules 2019, 9, 865. [Google Scholar] [CrossRef] [Green Version]
- Foster, S.R.; Porrello, E.R.; Stefani, M.; Smith, N.J.; Molenaar, P.; Dos Remedios, C.G.; Thomas, W.G.; Ramialison, M. Cardiac gene expression data and in silico analysis provide novel insights into human and mouse taste receptor gene regulation. Naunyn. Schmiedebergs. Arch. Pharmacol. 2015, 388, 1009–1027. [Google Scholar] [CrossRef]
- Barzilai, N.; Banerjee, S.; Hawkins, M.; Chen, W.; Rossetti, L. Caloric restriction reverses hepatic insulin resistance in aging rats by decreasing visceral fat. J. Clin. Investig. 1998, 101, 1353–1361. [Google Scholar] [CrossRef]
- Margolis, L.M.; Rivas, D.A.; Ezzyat, Y.; Gaffney-Stomberg, E.; Young, A.J.; McClung, J.P.; Fielding, R.A.; Pasiakos, S.M. Calorie restricted high protein diets downregulate lipogenesis and lower intrahepatic triglyceride concentrations in male rats. Nutrients 2016, 8, 571. [Google Scholar] [CrossRef]
- Shanik, M.H.; Xu, Y.; Skrha, J.; Dankner, R.; Zick, Y.; Roth, J. Insulin resistance and hyperinsulinemia: Is hyperinsulinemia the cart or the horse? Diabetes Care 2008, 31 (Suppl. 2), S262–S268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, K.F.; Dufour, S.; Savage, D.B.; Bilz, S.; Solomon, G.; Yonemitsu, S.; Cline, G.W.; Befroy, D.; Zemany, L.; Kahn, B.B.; et al. The role of skeletal muscle insulin resistance in the pathogenesis of the metabolic syndrome. Proc. Natl. Acad. Sci. USA 2007, 104, 12587–12594. [Google Scholar] [CrossRef] [Green Version]
- Salamanca, A.; Bárcena, B.; Arribas, C.; Fernández-Agulló, T.; Martínez, C.; Carrascosa, J.M.; Ros, M.; Andrés, A.; Gallardo, N. Aging impairs the hepatic subcellular distribution of ChREBP in response to fasting/feeding in rats: Implications on hepatic steatosis. Exp. Gerontol. 2015, 69, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, A.; Mazuecos, L.; Pintado, C.; Rubio, B.; López, V.; de Solís, A.J.; Rodríguez, M.; Andrés, A.; Gallardo, N. Effects of moderate chronic food restriction on the development of postprandial dyslipidemia with ageing. Nutrients 2019, 11, 1865. [Google Scholar]
- Gebert, N.; Cheng, C.W.; Kirkpatrick, J.M.; Di Fraia, D.; Yun, J.; Schädel, P.; Pace, S.; Garside, G.B.; Werz, O.; Rudolph, K.L.; et al. Region-Specific Proteome Changes of the Intestinal Epithelium during Aging and Dietary Restriction. Cell Rep. 2020, 31, 107565. [Google Scholar] [CrossRef] [PubMed]
- Brégère, C.; Rebrin, I.; Gallaher, T.K.; Sohal, R.S. Effects of age and calorie restriction on tryptophan nitration, protein content, and activity of succinyl-CoA:3-ketoacid CoA transferase in rat kidney mitochondria. Free Radic. Biol. Med. 2010, 48, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soeters, M.R.; Sauerwein, H.P.; Faas, L.; Smeenge, M.; Duran, M.; Wanders, R.J.; Ruiter, A.F.; Ackermans, M.T.; Fliers, E.; Houten, S.M.; et al. Effects of Insulin on Ketogenesis Following Fasting in Lean and Obese Men. Obesity 2009, 17, 1326–1331. [Google Scholar] [CrossRef]
- Kerndt, P.R.; Naughton, J.L.; Driscoll, C.E.; Loxterkamp, D.A. Fasting: The history, pathophysiology and complications. West. J. Med. 1982, 137, 379–399. [Google Scholar]
- Panickar, K.S.; Jewell, D.E. The benefit of anti-inflammatory and renal-protective dietary ingredients on the biological processes of aging in the kidney. Biology 2018, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Zhan, J.; Wang, K.; Zhang, C.C.; Zhang, C.C.; Li, Y.; Zhang, Y.; Chang, X.; Zhou, Q.; Yao, Y.; Liu, Y.; et al. GSPE inhibits HMGB1 release, attenuating renal IR-induced acute renal injury and chronic renal fibrosis. Int. J. Mol. Sci. 2016, 17, 1647. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, A.; Fonarow, G.C. The cardiorenal connection in heart failure. Curr. Cardiol. Rep. 2008, 10, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Maita, K.; Matsunuma, N.; Masuda, H.; Suzuki, Y. The age-related tumor incidence in Wistar-Imamichi rat. Exp. Anim. 1979, 28, 555–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinicola, S.; Cucina, A.; Pasqualato, A.; D’Anselmi, F.; Proietti, S.; Lisi, E.; Pasqua, G.; Antonacci, D.; Bizzarri, M. Antiproliferative and apoptotic effects triggered by grape seed extract (GSE) versus epigallocatechin and procyanidins on colon cancer cell lines. Int. J. Mol. Sci. 2012, 13, 651–664. [Google Scholar] [CrossRef]
- Yousef, M.I.; Khalil, D.K.A.M.; Abdou, H.M. Neuro- and nephroprotective effect of grape seed proanthocyanidin extract against carboplatin and thalidomide through modulation of inflammation, tumor suppressor protein p53, neurotransmitters, oxidative stress and histology. Toxicol. Rep. 2018, 5, 568–578. [Google Scholar] [CrossRef]
- Guo, F.; Hu, Y.; Niu, Q.; Li, Y.; Ding, Y.; Ma, R.; Wang, X.; Li, S.; Xie, J. Grape Seed Proanthocyanidin Extract Inhibits Human Esophageal Squamous Cancerous Cell Line ECA109 via the NF-κB Signaling Pathway. Mediators Inflamm. 2018, 2018. [Google Scholar] [CrossRef]
- Gil-cardoso, K.; Ginés, I.; Ardévol, A.; Blay, M.; Terra, X. Effects of fl avonoids on intestinal in fl ammation, barrier integrity and changes in gut microbiota during diet-induced obesity. Nutr. Res. Rev. 2018, 29, 234–248. [Google Scholar] [CrossRef]
- Spranger, I. Chemical characterization and antioxidant activities of oligomeric and polymeric procyanidin fractions from grape seeds. Food Chem. 2008, 108, 519–532. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | YOUNG | 21-MONTHS | GSPE PRE |
|---|---|---|---|
| n | 10 | 13 | 11 |
| Body weight (g) | 256.6 ± 4.3 * | 367.4 ± 15.0 | 366.8 ± 14.2 |
| mWAT (g) | 3.7 ± 0.2 * | 13.1 ± 1.2 | 10.5 ± 1.3 |
| oWAT (g) | 6.8 ± 0.2 * | 16.6 ± 1.5 | 15.5 ± 1.5 |
| rWAT (g) | 4.0 ± 0.3 * | 11.1 ± 1.1 | 10.6 ± 1.1 |
| Total visceral WAT (g) | 14.6 ± 0.2 * | 39.5 ± 3.4 | 34.8 ± 3.3 |
| BAT (g) | 0.4 ± 0.0 * | 0.7 ± 0.1 | 0.7 ± 0.1 |
| % visceral adiposity | 5.4 ± 0.2 * | 11.3 ± 0.6 | 9.5 ± 0.7 * |
| Liver (g) | 6.2 ± 0.2 * | 8.7 ± 0.4 | 7.7 ± 0.2 * |
| Spleen (g) | 0.5 ± 0.0 * | 0.8 ± 0.0 | 0.8 ± 0.0 |
| Kidney (g) | 0.8 ± 0.0 * | 1.0 ± 0.0 | 0.9 ± 0.0 # |
| Variable | YOUNG | 21-MONTHS | GSPE PRE |
|---|---|---|---|
| Plasma | |||
| Glucose (mM) | 7.3 ± 0.3 | 7.0 ± 0.3 | 8.1 ± 0.6 |
| TAG (mM) | 0.4 ± 0.1 # | 0.6 ± 0.1 | 0.5 ± 0.1 |
| NEFA (mM) | 1.0 ± 0.1 | 1.0 ± 0.1 | 0.9 ± 0.1 |
| Cholesterol (mM) | 2.6 ± 0.1 * | 4.5 ± 0.4 | 4.2 ± 0.4 |
| β-Hydroxybutyrate (mM) | 0.7 ± 0.1 * | 0.3 ± 0.1 | 0.5 ± 0.1 |
| Urea (mM) | 4.2 ± 0.2 | 3.8 ± 0.2 | 4.3 ± 0.1 # |
| Creatinine (μM) | 7.1 ± 0.3 | 7.7 ± 0.6 | 7.3 ± 0.7 |
| Insulin (pM) | 182.7 ± 1.0 * | 322.2 ± 36.4 | 233.7 ± 13.4 * |
| Glucagon (pM) | 7.2 ± 1.3 * | 18.2 ± 2.5 | 18.8 ± 1.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grau-Bové, C.; Sierra-Cruz, M.; Miguéns-Gómez, A.; Rodríguez-Gallego, E.; Beltrán-Debón, R.; Blay, M.; Terra, X.; Pinent, M.; Ardévol, A. A Ten-Day Grape Seed Procyanidin Treatment Prevents Certain Ageing Processes in Female Rats over the Long Term. Nutrients 2020, 12, 3647. https://doi.org/10.3390/nu12123647
Grau-Bové C, Sierra-Cruz M, Miguéns-Gómez A, Rodríguez-Gallego E, Beltrán-Debón R, Blay M, Terra X, Pinent M, Ardévol A. A Ten-Day Grape Seed Procyanidin Treatment Prevents Certain Ageing Processes in Female Rats over the Long Term. Nutrients. 2020; 12(12):3647. https://doi.org/10.3390/nu12123647
Chicago/Turabian StyleGrau-Bové, Carme, Marta Sierra-Cruz, Alba Miguéns-Gómez, Esther Rodríguez-Gallego, Raúl Beltrán-Debón, Mayte Blay, Ximena Terra, Montserrat Pinent, and Anna Ardévol. 2020. "A Ten-Day Grape Seed Procyanidin Treatment Prevents Certain Ageing Processes in Female Rats over the Long Term" Nutrients 12, no. 12: 3647. https://doi.org/10.3390/nu12123647
APA StyleGrau-Bové, C., Sierra-Cruz, M., Miguéns-Gómez, A., Rodríguez-Gallego, E., Beltrán-Debón, R., Blay, M., Terra, X., Pinent, M., & Ardévol, A. (2020). A Ten-Day Grape Seed Procyanidin Treatment Prevents Certain Ageing Processes in Female Rats over the Long Term. Nutrients, 12(12), 3647. https://doi.org/10.3390/nu12123647