The Protective and Long-Lasting Effects of Human Milk Oligosaccharides on Cognition in Mammals




Abstract
1. Introduction
2. Assessing the Effects of HMOs on Cognitive Measures in Animal Models
3. Effects of HMOs on Cognition in Mammals
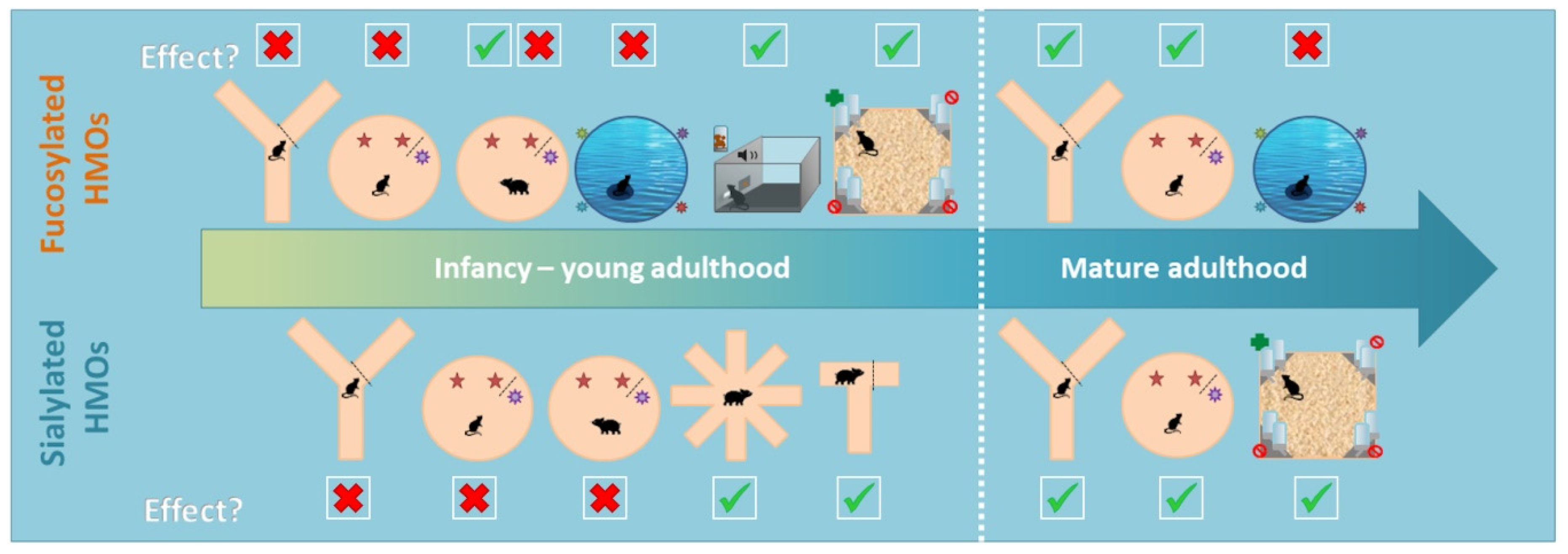
3.1. Main Behavioural Findings
3.1.1. Simple Cognitive Tasks
3.1.2. Complex Cognitive Tasks
3.2. Effects of HMOs on Long Term Potentiation (LTP)
4. Discussion
Potential Underlying Mechanisms
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A
References
- World Health Organization. Available online: https://www.who.int/health-topics/breastfeeding#tab=tab_1 (accessed on 6 May 2020).
- Feldman, R.; Eidelman, A.I. Direct and indirect effects of breast milk on the neurobehavioral and cognitive development of premature infants. Dev. Psychobiol. 2003, 43, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Boquien, C.-Y. Human Milk: An Ideal Food for Nutrition of Preterm Newborn. Front. Pediatr. 2018, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vandenplas, Y.; Berger, B.; Carnielli, V.P.; Ksiazyk, J.; Lagström, H.; Luna, M.S.; Migacheva, N.; Mosselmans, J.M.; Picaud, J.C.; Possner, M.; et al. Human milk oligosaccharides: 2′-fucosyllactose (2′-FL) and lacto-n-neotetraose (LNnT) in infant formula. Nutrients 2018, 10, 1161. [Google Scholar] [CrossRef]
- Bar, S.; Milanaik, R.; Adesman, A. Long-term neurodevelopmental benefits of breastfeeding. Curr. Opin. Pediatr. 2016, 28, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Nolan, L.S.; Parks, O.B.; Good, M. A review of the immunomodulating components of maternal breast milk and protection against necrotizing enterocolitis. Nutrients 2019, 12, 14. [Google Scholar] [CrossRef]
- Carucci, L.; Nocerino, R.; Paparo, L.; Di Scala, C.; Canani, R.B. Dietary Prevention of Atopic March in Pediatric Subjects With Cow’s Milk Allergy. Front. Pediatr. 2020, 8, 1–9. [Google Scholar] [CrossRef]
- Munblit, D.; Peroni, D.G.; Boix-Amoros, A.; Hsu, P.S.; Van‘t Land, B.; Gay, M.C.L.; Kolotilina, A.; Skevaki, C.; Boyle, R.J.; Collado, M.C.; et al. Human Milk and Allergic Diseases: An Unsolved Puzzle. Nutrients 2017, 9, 894. [Google Scholar] [CrossRef]
- Rajani, P.S.; Seppo, A.E.; Järvinen, K.M. Immunologically Active Components in Human Milk and Development of Atopic Disease, With Emphasis on Food Allergy, in the Pediatric Population. Front. Pediatr. 2018, 6, 1–13. [Google Scholar] [CrossRef]
- Den Dekker, H.T.; Sonnenschein-van der Voort, A.M.M.; Jaddoe, V.W.V.; Reiss, I.K.; de Jongste, J.C.; Duijts, L. Breastfeeding and asthma outcomes at the age of 6 years: The Generation R Study. Pediatr. Allergy Immunol. 2016, 27, 486–492. [Google Scholar] [CrossRef]
- Le Doare, K.; Holder, B.; Bassett, A.; Pannaraj, P.S. Mother’s Milk: A purposeful contribution to the development of the infant microbiota and immunity. Front. Immunol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Çatli, G.; Dündar, N.O.; Dündar, B.N. Adipokines in Breast Milk: An Update. J. Clin. Res. Pediatr. Endocrinol. 2014, 6, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Gila-Diaz, A.; Arribas, S.M.; Algara, A.; Martín-Cabrejas, M.A.; López de Pablo, Á.L.; Sáenz de Pipaón, M.; Ramiro-Cortijo, D. A Review of Bioactive Factors in Human Breastmilk: A Focus on Prematurity. Nutrients 2019, 11, 1307. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.S.; Aboud, F.; Mironova, E.; Vanilovich, I.; Platt, R.W.; Matush, L.; Igumnov, S.; Fombonne, E.; Bogdanovich, N.; Ducruet, T.; et al. Breastfeeding and child cognitive development: New evidence from a large randomized trial. Arch. Gen. Psychiatry 2008, 65, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Poton, W.L.; Soares, A.L.G.; de Oliveira, E.R.A.; Gonçalves, H. Breastfeeding and behavior disorders among children and adolescents: A systematic review. Rev. Saude Publica 2018, 52, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Horta, B.L.; Loret De Mola, C.; Victora, C.G. Breastfeeding and intelligence: A systematic review and meta-analysis. Acta Paediatr. Int. J. Paediatr. 2015, 104, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Deoni, S.C.L.; Dean, D.C.; Piryatinsky, I.; O’Muircheartaigh, J.; Waskiewicz, N.; Lehman, K.; Han, M.; Dirks, H. Breastfeeding and early white matter development: A cross-sectional study. Neuroimage 2013, 82, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Herba, C.M.; Roza, S.; Govaert, P.; Hofman, A.; Jaddoe, V.; Verhulst, F.C.; Tiemeier, H. Breastfeeding and early brain development: The Generation R study. Matern. Child Nutr. 2013, 9, 332–349. [Google Scholar] [CrossRef]
- Allen-Blevins, C.R.; You, X.; Hinde, K.; Sela, D.A. Handling stress may confound murine gut microbiota studies. PeerJ 2017, 5, 1–21. [Google Scholar] [CrossRef]
- Andreas, N.J.; Kampmann, B.; Mehring Le-Doare, K. Human breast milk: A review on its composition and bioactivity. Early Hum. Dev. 2015, 91, 629–635. [Google Scholar] [CrossRef]
- Mudd, A.T.; Dilger, R.N. Early-Life Nutrition and Neurodevelopment: Use of the Piglet as a Translational Model. Adv. Nutr. Int. Rev. J. 2017, 8, 92–104. [Google Scholar] [CrossRef]
- Wang, H.J.; Hua, C.Z.; Ruan, L.L.; Hong, L.Q.; Sheng, S.Q.; Shang, S.Q. Sialic Acid and Iron Content in Breastmilk of Chinese Lactating Women. Indian Pediatr. 2017, 54, 1029–1031. [Google Scholar] [CrossRef] [PubMed]
- Ayechu-Muruzabal, V.; van Stigt, A.H.; Mank, M.; Willemsen, L.E.M.; Stahl, B.; Garssen, J.; van’t Land, B. Diversity of human milk oligosaccharides and effects on early life immune development. Front. Pediatr. 2018, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hegar, B.; Wibowo, Y.; Basrowi, R.W.; Ranuh, R.G.; Sudarmo, S.M.; Munasir, Z.; Atthiyah, A.F.; Widodo, A.D.; Supriatmo Kadim, M.; Suryawan, A.; et al. The Role of Two Human Milk Oligosaccharides, 2′-Fucosyllactose and Lacto-N-Neotetraose, in Infant Nutrition. Pediatr. Gastroenterol. Hepatol. Nutr. 2019, 22, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Urashima, T.; Saito, T.; Nakamura, T.; Messer, M. Oligosaccharides of milk and colostrum in non-human mammals. Glycoconj. J. 2001, 18, 357–371. [Google Scholar] [CrossRef]
- Ten Bruggencate, S.J.; Bovee-Oudenhoven, I.M.; Feitsma, A.L.; van Hoffen, E.; Schoterman, M.H. Functional role and mechanisms of sialyllactose and other sialylated milk oligosaccharides. Nutr. Rev. 2014, 72, 377–389. [Google Scholar] [CrossRef]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; Garcia-Mantrana, I.; Bertua-Rios, B.; Martinez-Costa, C.; Borsch, C.; Rudloff, S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef]
- Austin, S.; de Castro, C.A.; Bénet, T.; Hou, Y.; Sun, H.; Thakkar, S.K.; Vinyes-Pares, G.; Zhang, Y.; Wang, P. Temporal change of the content of 10 oligosaccharides in the milk of Chinese urban mothers. Nutrients 2016, 8, 346. [Google Scholar] [CrossRef]
- Thurl, S.; Munzert, M.; Henker, J.; Boehm, G.; Müller-Werner, B.; Jelinek, J.; Stahl, B. Variation of human milk oligosaccharides in relation to milk groups and lactational periods. Br. J. Nutr. 2010, 104, 1261–1271. [Google Scholar] [CrossRef]
- Austin, S.; De Castro, C.A.; Sprenger, N.; Binia, A.; Affolter, M.; Garcia-Rodenas, C.L.; Beauport, L.; Tolsa, J.F.; Fischer Fumeaux, C.J. Human milk oligosaccharides in the milk of mothers delivering term versus preterm infants. Nutrients 2019, 11, 1282. [Google Scholar] [CrossRef]
- Azad, M.B.; Robertson, B.; Atakora, F.; Becker, A.B.; Subbarao, P.; Moraes, T.J.; Mandhane, P.J.; Turvey, S.E.; Lefebvre, D.L.; Sears, M.R.; et al. Human Milk Oligosaccharide Concentrations Are Associated with Multiple Fixed and Modifiable Maternal Characteristics, Environmental Factors, and Feeding Practices. J. Nutr. 2018, 148, 1733–1742. [Google Scholar] [CrossRef]
- Paganini, D.; Uyoga, M.; Kortman, G.A.M.; Boekhorst, J.; Schneeberger, S.; Karanja, S.; Hennet, T.; Zimmerman, M.B. Maternal Human Milk Oligosaccharide Profile Modulates the Impact of an Intervention with Iron and galacto-Oligosaccharides in Kenyan Infants. Nutrients 2019, 11, 2596. [Google Scholar] [CrossRef] [PubMed]
- Lis-Kuberka, J.; Orczyk-Pawilowicz, M. Sialylated Oligosaccharides and Glycoconjugates of Human Milk. The Impact on Infant and Newborn Protection, Development and Well-Being. Nutrients 2019, 11, 306. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.L.; Cubero, J.; Sánchez, J.; Franco, L.; Rodriguez, A.B.; Rivero, M. Evolution of the circadian profile of human milk amino acids during breastfeeding. J. Appl. Biomed. 2013, 11, 59–70. [Google Scholar] [CrossRef]
- Goehring, K.C.; Marriage, B.J.; Oliver, J.S.; Wilder, J.A.; Barrett, E.G.; Buck, R.H. Similar to Those Who Are Breastfed, Infants Fed a Formula Containing 2′-Fucosyllactose Have Lower Inflammatory Cytokines in a Randomized Controlled Trial. J. Nutr. 2016, 146, 2559–2566. [Google Scholar] [CrossRef] [PubMed]
- Steenhout, P.; Sperisen, P.; Martin, F.P.; Sprenger, N.; Wernimont, S.; Pecquet, S.; Berger, B. Term infant formula supplemented with human milk oligosaccharides (2′fucosyllactose and lacto-N-neotetraose) shifts stool microbiota and metabolic signatures closer to that of breastfed infants. FASEB J. 2016, 30, 275–277. [Google Scholar] [CrossRef]
- Berger, P.K.; Plows, J.F.; Jones, R.B.; Alderete, T.L.; Yonemitsu, C.; Poulsen, M.; Ryoo, J.H.; Peterson, B.S.; Bode, L.; Goran, M.I. Human milk oligosaccharide 2′-fucosyllactose links feedings at 1 month to cognitive development at 24 months in infants of normal and overweight mothers. PLoS ONE 2020, 15, e228323. [Google Scholar] [CrossRef]
- Bilbo, S.D.; Schwarz, J.M. The Immune System and Developmental Programming of Brain Front Neuroendocrinol Author Manuscript and Behavior. NIH Public Access 2012, 33, 267–286. [Google Scholar] [CrossRef]
- Wallace, T.L.; Ballard, T.M.; Glavis-Bloom, C. Animal Paradigms to Assess Cognition with Translation to Humans. Handb. Exp. Pharmacol. 2015, 228, 27–57. [Google Scholar] [CrossRef]
- Wiera, G.; Nowak, D.; van Hove, I.; Dziegiel, P.; Moons, L.; Mozrzymas, J.W. Mechanisms of NMDA receptor- and voltage-gated L-type calcium channel-dependent hippocampal LTP critically rely on proteolysis that is mediated by distinct metalloproteinases. J. Neurosci. 2017, 37, 1240–1256. [Google Scholar] [CrossRef]
- Gieling, E.T.; Nordquist, R.E.; van der Staay, F.J. Assessing learning and memory in pigs. Anim. Cogn. 2011, 14, 151–173. [Google Scholar] [CrossRef]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Vorhees, C.V.; Williams, M.T. Assessing Spatial Learning and Memory in Rodents. ILAR J. 2014, 55, 310–332. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Takahashi, S.; Sakurai, Y. Reinforcement schedules differentially affect learning in neuronal operant conditioning in rats. Neurosci. Res. 2020, 153, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Kiryk, A.; Janusz, A.; Zglinicki, B.; Turkes, E.; Knapska, E.; Konopka, W.; Lipp, H.P.; Kaczmarek, L. IntelliCage as a tool for measuring mouse behaviour—20 years perspective. Behav. Brain Res. 2020, 388, 1–17. [Google Scholar] [CrossRef]
- Oliveros, E.; Ramirez, M.; Vazquez, E.; Barranco, A.; Gruart, A.; Delgado-Garcia, J.M.; Buck, R.; Rueda, R.; Martin, M.J. Oral supplementation of 2′-fucosyllactose during lactation improves memory and learning in rats. J. Nutr. Biochem. 2015, 31, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, E.; Vázquez, E.; Barranco, A.; Ramírez, M.; Gruart, A.; Delgado-Garcia, J.M.; Buck, R.; Rueda, R.; Martín, M.J. Sialic acid and sialylated oligosaccharide supplementation during lactation improves learning and memory in rats. Nutrients 2018, 10, 1519. [Google Scholar] [CrossRef]
- Vazquez, E.; Barranco, A.; Ramirez, M.; Gruart, A.; Delgado-Garcia, J.M.; Jimenez, M.L.; Buck, R.; Rueda, R. Dietary 2′-fucosyllactose enhances operant conditioning and long-term potentiation via gut-brain communication through the vagus nerve in rodents. PLoS ONE 2016, 11, e166070. [Google Scholar] [CrossRef]
- Vázquez, E.; Barranco, A.; Ramírez, M.; Delgado-Garcia, J.M.; Martínez-Lara, E.; Blanco, S.; Martín, M.J.; Castanys, E.; Buck, R.; Prieto, P.; et al. Effects of a human milk oligosaccharide, 2′-fucosyllactose, on hippocampal long-term potentiation and learning capabilities in rodents. J. Nutr. Biochem. 2015, 26, 455–465. [Google Scholar] [CrossRef]
- Wang, B.; Yu, B.; Karim, M.; Hu, H.; Sun, Y.; McGreevy, P.; Petocz, P.; Held, S.; Brand-Miller, J. Dietary sialic acid supplementation improves learning and memory in piglets. Am. J. Clin. Nutr. 2007, 85, 561–569. [Google Scholar] [CrossRef]
- Obelitz-Ryom, K.; Bering, S.B.; Overgaard, S.H.; Eskildsen, S.F.; Ringgaard, S.; Olesen, J.L.; Skovgaard, K.; Pankratova, S.; Wang, B.; Brunse, A.; et al. Bovine Milk Oligosaccharides with Sialyllactose Improves Cognition in Preterm Pigs. Nutrients 2019, 11, 1335. [Google Scholar] [CrossRef]
- Fleming, S.A.; Mudd, A.T.; Hauser, J.; Yan, J.; Metairon, S.; Steiner, P.; Donovan, S.M.; Dilger, R.N. Human and Bovine Milk Oligosaccharides Elicit Improved Recognition Memory Concurrent With Alterations in Regional Brain Volumes and Hippocampal mRNA Expression. Front. Neurosci. 2020, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Fleming, S.A.; Mudd, A.T.; Hauser, J.; Yan, J.; Metairon, S.; Steiner, P.; Donovan, S.M.; Dilger, R.N. Dietary Oligofructose Alone or in Combination with 2′-Fucosyllactose Differentially Improves Recognition Memory and Hippocampal mRNA Expression. Nutrients 2020, 12, 2131. [Google Scholar] [CrossRef] [PubMed]
- Fleming, S.A.; Chichlowski, M.; Berg, B.M.; Donovan, S.M.; Dilger, R.N. Dietary sialyllactose does not influence measures of recognition memory or diurnal activity in the young pig. Nutrients 2018, 10, 395. [Google Scholar] [CrossRef] [PubMed]
- Pressler, R.; Auvin, S. Comparison of brain maturation among species: An example in translational research suggesting the possible use of bumetanide in newborn. Front. Neurol. 2013, 4, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106–107, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Radlowski, E.C.; Conrad, M.S.; Lezmi, S.; Dilger, R.N.; Sutton, B.; Larsen, R.; Johnson, R.W. A Neonatal Piglet Model for Investigating Brain and Cognitive Development in Small for Gestational Age Human Infants. PLoS ONE 2014, 9, e91951. [Google Scholar] [CrossRef]
- Shumake, J.; Barrett, D.W.; Lane, M.A.; Wittke, A.J. Behavioral effects of bovine lactoferrin administration during postnatal development of rats. BioMetals 2014, 27, 1039–1055. [Google Scholar] [CrossRef]
- Bonfanti, L. PSA-NCAM in mammalian structural plasticity and neurogenesis. Prog. Neurobiol. 2006, 80, 129–164. [Google Scholar] [CrossRef]
- Weledji, E.P.; Assob, J.C. The ubiquitous neural cell adhesion molecule (N-CAM). Ann. Med. Surg. 2014, 3, 77–81. [Google Scholar] [CrossRef]
- Murrey, H.E.; Hsieh-Wilson, L.C. The chemical neurobiology of carbohydrates. Chem Rev. 2008, 108, 1708–1731. [Google Scholar] [CrossRef]
- Sahay, A.; Scobie, K.N.; Hill, A.S.; O’Carroll, C.M.; Kheirbek, M.A.; Burghardt, N.S.; Fenton, A.A.; Dranovsky, A.; Hen, R. Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 2011, 472, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.T.; Nanthakumar, N.N.; Newburg, D.S. The human milk oligosaccharide 2′-fucosyllactose quenches Campylobacter jejuni-induced inflammation in human epithelial cells HEp-2 and HT-29 and in mouse intestinal Mucosa1-3. J. Nutr. 2016, 146, 1980–1990. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Bode, L. The functional biology of human milk oligosaccharides. Early Hum. Dev. 2015, 91, 619–622. [Google Scholar] [CrossRef] [PubMed]
- Tarr, A.J.; Galley, J.D.; Fisher, S.; Chichlowski, M.; Berg, B.M.; Bailey, M.T. The prebiotics 3′Sialyllactose and 6′Sialyllactose diminish stressor-induced anxiety-like behavior and colonic microbiota alterations: Evidence for effects on the gut-brain axis. Brain Behav. Immun. 2016, 50, 166–177. [Google Scholar] [CrossRef]
- Kuntz, S.; Kunz, C.; Borsch, C.; Vazquez, E.; Buck, R.; Reutzel, M.; Eckert, G.P.; Rudloff, S. Metabolic Fate and Distribution of 2′-Fucosyllactose: Direct Influence on Gut Microbial Activity but not on Brain. Mol. Nutr. Food Res. 2019, 63, 1–8. [Google Scholar] [CrossRef]
- Thongaram, T.; Hoeflinger, J.L.; Chow, J.M.; Miller, M.J. Human milk oligosaccharide consumption by probiotic and human-associated bifidobacteria and lactobacilli. J. Dairy Sci. 2017, 100, 7825–7833. [Google Scholar] [CrossRef]
- Lawson, M.; O’Neill, I.J.; Kujawska, M.; Javvadi, S.G.; Wijeyesekera, A.; Flegg, Z.; Chalklen, L.; Hall, L.J. Breast milk-derived human milk oligosaccharides promote Bifidobacterium interactions within a single ecosystem. ISME J. 2020, 14, 635–648. [Google Scholar] [CrossRef]
- Lees, H.; Swann, J.; Poucher, S.M.; Nicholson, J.K.; Holmes, E.; Wilson, I.D.; Marchesi, J.R. Age and microenvironment outweigh genetic influence on the Zucker rat microbiome. PLoS ONE 2014, 9, e100916. [Google Scholar] [CrossRef]
- Moossavi, S.; Sepehri, S.; Robertson, B.; Bode, L.; Goruk, S.; Field, C.J.; Lix, L.M.; de Souza, R.J.; Becker, A.B.; Mandhane, P.J.; et al. Composition and Variation of the Human Milk Microbiota Are Influenced by Maternal Early-Life Factors. Cell Host Microbe 2019, 25, 324–335.e4. [Google Scholar] [CrossRef]
- Ramani, S.; Stewart, C.J.; Laucirica, D.R.; Ajami, N.J.; Robertson, B.; Autran, C.A.; Shinge, D.; Rani, S.; Anandan, S.; Hu, L.; et al. Human milk oligosaccharides, milk microbiome and infant gut microbiome modulate neonatal rotavirus infection. Nature 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Borewicz, K.; Gu, F.; Saccenti, E.; Hechler, C.; Beijers, R.; de Weerth, C.; van Leeuwen, S.S.; Schols, H.A.; Smidt, H. The association between breastmilk oligosaccharides and faecal microbiota in healthy breastfed infants at two, six and twelve weeks of age. Sci Rep 2020, 10, 4270. [Google Scholar] [CrossRef] [PubMed]
- Oomen, C.A.; Hvoslef-Eide, M.; Heath, C.J.; Mar, A.C.; Horner, A.E.; Bussey, T.J.; Saksida, L.M. The touchscreen operant platform for testing working memory and pattern separation in rats and mice. Nat. Protoc. 2013, 8, 2006–2021. [Google Scholar] [CrossRef] [PubMed]
- Higgins, G.A.; Silenieks, L.B. Rodent test of attention and impulsivity: The 5-choice serial reaction time task. Curr. Protoc. Pharmacol. 2017, 78, 5.49.1–5.49.34. [Google Scholar] [CrossRef] [PubMed]
- Dunnett, S.B.; Heuer, A.; Lelos, M.; Brooks, S.P.; Rosser, A.E. Bilateral striatal lesions disrupt performance in an operant delayed reinforcement task in rats. Brain Res. Bull. 2012, 88, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Study | Species | HMO Component and Dose | Age and Duration Administration | Age Test | Tests | Key Results |
|---|---|---|---|---|---|---|
| Fucosylated HMOs | ||||||
| Oliveros et al., 2016 [46] | Lister Hooded Rats | 2′-FL (1g/KG/BW) | From PND 3–weaning | Long Term study: (1) 4–6 weeks (2) 1 year old | NORT Y maze MWM LTP (only at 1 year) | At 6 weeks of age no differences in behaviour (n = 12) were found. At 1 year of age, 2′-FL rats showed improved performance in the NORT and Y- maze paradigms. No effect was observed in the MWM. LTP was more intense and long lasting in the 2′-FL supplemented groups (n = 10) |
| Sprague Dawley Rats | 2′-FL (1g/ KG/BW) | From PND3 until week 6 | Short Term study: 6 weeks | LTP | LTP was more intense and long lasting in the 2′-FL supplemented groups (n = 10). | |
| Vazquez et al., 2016 [48] | Sprague Dawley Rats | 2′FL (350 mg/kg/BW via AIN-93M diet) L-Fucose (Fuc) (equimolar amounts of fuc and 2′-FL via AIN-93M diet) | 3–4.5 months old for 5 weeks | Started at 2.5–4 months old | Operant conditioning (FR1) LTP | 2′-FL but not fuc displayed enhanced LTP. Vagotomy inhibited the effects of oral 2′-FL on LTP (n = 10) and operant learning paradigms (n = 10). |
| Fleming et al., 2020a [52] | Pigs (1050 Cambro genetics) | Three groups: Oligofructose (OF) 5 g/L OF + 2′-FL 5 g/L OF + 1 g/L 2′-FL Control. Nothing | PND 2–33 | PND 22 | NORT | Pigs (n = 12) who received Oligofructose (OF) displayed enhanced object recognition when tested 1 h after being habituated to the two objects. When pigs consumed both 2′-FL and OF, they showed improved recognition memory after a 48-h delay. |
| Fleming et al., 2020b [53] | Pigs (1050 Cambro genetics) | Four groups: HMOs (2′FL + LNnT) 1 g/L 2′-FL + 0.5 g/L LNnT BMOs 12.4 g/L HMOs + BMOs 1 g/L 2′-FL + 0.5 g/L LNnT + 12.4 g/L BMOs Control. Nothing | PND 2–33 | PND 22 | NORT | Pigs (n = 12) who received only HMOs displayed enhanced object recognition when tested 1 h after being habituated to the two objects. When pigs consumed both HMOs and BMOs, they showed improved recognition memory after a 48-h delay. |
| Vazquez et al., 2015 [46] | Sprague Dawley Rats | 2′FL (1 g/kg/BW) via oral gavage during acute administration and 2′-FL (350 mg/Kg/BW) via AIN-93G diet, during short time feeding | Acute administration: when rats were 3 months old Short-time feeding from 2.5–4 months, for 5 weeks | Operant tests started when administration started. LTP was performed after administration period. | Operant conditioning (FR1) LTP | 2′-FL groups performed better in operant learning paradigms (rats n = 10, mice n = 28) and showed an enhanced LTP response (rats and mice n = 8). The long-time supplementation of 2′FL also increased the expression of molecules involved in storage newly acquired memories (BDNF, PSD-95 phosphorylated CamKII, etc.) |
| C57BL/6 mice | 2′FL (350 mg/Kg/BW via AIN-93G diet) | Long-time feeding from 2–3.5 months old, for 12 weeks | Intellicage (FR1, FR4, FR8) LTP | |||
| Sialylated HMOs | ||||||
| Oliveros et al., 2018 [47] | Sprague Dawley Rats | Neu5Ac 6′-SL (Dose ranged from 400 mg/Kg/BW to 2600 mg/Kg/BW based on theoretical model) | From PND 3 until weaning | After weaning | NORT Y maze | No effects detected after weaning (n = 10). At 1 year old, sia (Neu5Ac and 6′-SL) exposed rats (n = 8♀) showed improved performance on all the behavioural tests (NORT, Y-maze, Intellicage) and showed enhanced LTP (n = 10) when compared to the control group. Of the SL supplemented animals, the 6′-SL group performed better than the Neu5AC group |
| 1 year old | NORT Y maze Intellicage LTP | |||||
| Wang et al., 2007 [50] | Piglets Landrace/Large White cross | Sialic Acid (ingredient of Casein glycomacropeptide cGMP)) (4 groups of animals with their own dose each; 0 mg/L (control), 140 mg/L; 300 mg/L; 635 mg/L and 830 mg/L) | From PND 3 until end of experiment | PND 21–PND 35 | 8-arm Radial maze | Supplemented groups (n = 12–14 per group) required less trials to learn the required response, with a dose–response correlation for the difficult task. |
| Obelitz-Ryom et al., 2019 [51] | Pre-term delivered (experimental groups) Piglets Landrace x Yorkshire x Duroc | Sialyllactose (6′-SL + 3′-SL) (380 mg/L) Lactose (control) (6000 mg/L) | PND1–PND19 | PND13–PND18 | Spatial T-maze | Four experimental groups were included in the study; PRE-SAL (n = 10 ♀, 10 ♂), PRE-CON (n = 9 ♀, 11 ♂), TERM-CON (n = 9 ♀, 5 ♂) and TERM-SAL (n = 6 ♀, 6 ♂). TERM CON piglets reached learning criteria of 80% correct choices on day 3, PRE-SAL on day 4 and PRE-CON on day 5. More PRE-SAL piglets reached the T maze learning criteria compared to PRE-CON piglets. Upregulation of genes for sialic acid metabolism, myelination and ganglioside biosynthesis were present in the hippocampus of SL supplemented preterm piglets. |
| Term delivered piglets (reference groups) Landrace x Yorkshire x Duroc | Lactose (control) (6000 mg/L) Pig’s milk (under natural rearing conditions) | |||||
| Fleming et al., 2018 [54] | Piglets (no breed specified) | Sialyllactose (380 mg/L) | PND2–PND22 | PND15–PND22 | NORT | No effects (n = 17) were observed. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Docq, S.; Spoelder, M.; Wang, W.; Homberg, J.R. The Protective and Long-Lasting Effects of Human Milk Oligosaccharides on Cognition in Mammals. Nutrients 2020, 12, 3572. https://doi.org/10.3390/nu12113572
Docq S, Spoelder M, Wang W, Homberg JR. The Protective and Long-Lasting Effects of Human Milk Oligosaccharides on Cognition in Mammals. Nutrients. 2020; 12(11):3572. https://doi.org/10.3390/nu12113572
Chicago/Turabian StyleDocq, Sylvia, Marcia Spoelder, Wendan Wang, and Judith R. Homberg. 2020. "The Protective and Long-Lasting Effects of Human Milk Oligosaccharides on Cognition in Mammals" Nutrients 12, no. 11: 3572. https://doi.org/10.3390/nu12113572
APA StyleDocq, S., Spoelder, M., Wang, W., & Homberg, J. R. (2020). The Protective and Long-Lasting Effects of Human Milk Oligosaccharides on Cognition in Mammals. Nutrients, 12(11), 3572. https://doi.org/10.3390/nu12113572