Zinc Deficiency—An Independent Risk Factor in the Pathogenesis of Haemorrhagic Stroke?


{kind=link}
Abstract
1. Introduction
2. Functions of Zinc
3. Resorption of Zinc
4. Zinc and Nutrition (Diet)
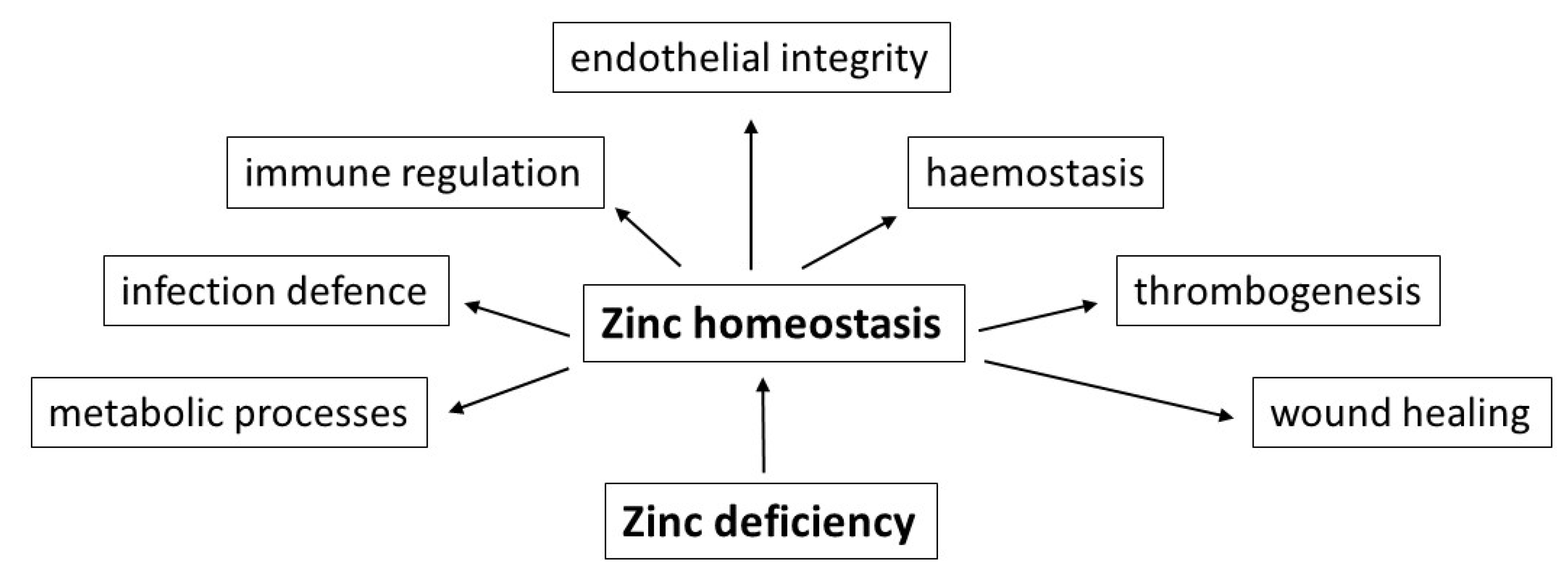
5. Zinc Deficiency
6. Zinc Deficiency: Influence on Vessels, Coagulation and Stroke
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tong, Y.N.; Appleby, P.N.; Bradbury, K.; Perez-Cornago, A.; Travis, R.C.; Clarke, R.; Key, T.J. Risks of ischaemic heart disease and stroke, in meat eaters, fish eaters, and vegetarians over 18 years follow-up: Results from the prospetive EPIC-Oxford study. BMJ 2019, 366, I14897. [Google Scholar] [CrossRef]
- Maret, W. Regulation of cellular zinc ions and their signaling functions. In Zinc Signaling, 2nd ed.; Fukada, T., Kambe, T., Eds.; Springer Nature: Singapore, 2019; pp. 5–22. [Google Scholar]
- Maret, W. The redox biology of redox-inert zinc ions. Free Radical Biol. Med. 2019, 134, 311–326. [Google Scholar] [CrossRef]
- Grüngreiff, K.; Reinhold, D. Zinc and Liver. In Zinc in Human Health, 1st ed.; Rink, L., Ed.; IOS Press: Amsterdam, The Netherlands, 2011; pp. 473–495. [Google Scholar]
- King, J.C. Zinc: An essential but elusive nutrient. Am. J. Clin. Nutr. 2011, 94, 679S–684S. [Google Scholar] [CrossRef]
- Vu, T.T.; Fredenburg, J.C.; Weitz, J.I. Zinc: An important co-factor in hemostasis and thrombosis. Thromb. Hemost. 2013, 109, 421–430. [Google Scholar] [CrossRef]
- Lin, P.-H.; Sermersheim, M.; Li, H.; Lee, P.H.U.; Steinberg, S.M.; Ma, J. Zinc in wound healing modulation. Nutrients 2017, 10, 16. [Google Scholar] [CrossRef]
- Mammadova-Bach, E.; Braun, A. Zinc homeostasis in platelet-related diseases. Int. J. Mol. Sci. 2019, 20, 5228. [Google Scholar] [CrossRef]
- King, J.C. Yet again, serum zinc concentrations are unrelated to zinc intakes. J. Nutr. 2018, 148, 1399–1401. [Google Scholar] [CrossRef]
- Coverdal, P.C.; Barnett, J.P.; Adamu, A.H.; Griffiths, E.J.; Stewart, A.J.; Blindauer, C.A. A metalloproteomic analysis of interactions between plasma proteins and zinc: Elevated fatty levels affect zinc distribution. Metallomics 2019, 11, 1805–1819. [Google Scholar] [CrossRef]
- Lonergan, Z.R.; Skaar, E.P. Nutrient zinc at the host-pathogen interface. Trends Biochem. Sci. 2019, 44, 1041–1056. [Google Scholar] [CrossRef]
- Markowitz, M.E.; Rosen, J.F.; Mizruchi, M. Circadian variations in serum zinc (Zn) concentrations: Correlation with blood ionized calcium, serum total calcium and phosphate in humans. Am. J. Clin. Nutr. 1985, 41, 689–696. [Google Scholar] [CrossRef]
- Kanabrocki, E.L.; Scheving, L.E.; Olwin, J.H.; Marks, G.E.; McCormick, J.B.; Halberg, F.; Pauly, J.E.; Greco, J.; De Bartolo, M.; Nemchausky, B.A.; et al. Circadian variation in the urinary excretion of electrolytes and trace elements in men. Am. J. Anat. 1983, 166, 121–148. [Google Scholar] [CrossRef]
- Hambidge, K.M.; Goodall, M.J.; Stall, C.; Pritts, J. Post-prandial and daily changes in plasma zinc. J. Trace Elem. Elektrolytes Health Dis. 1989, 3, 55–57. [Google Scholar]
- Henningar, S.R.; Lieberman, H.R.; Fulgoni, V.L.; McClung, J.P. Serum zinc concentrations in the US population are related to sex, age, and time of blood draw but not dietary or supplemental zinc. J. Nutr. 2018, 148, 1341–1351. [Google Scholar] [CrossRef]
- Gröber, U.; Kisters, K.; Classen, H.G. Zinkmangel im Fokus: Ursachen, Symptome, Diagnose und Therapie. EHK 2019, 68, 278–292. [Google Scholar] [CrossRef]
- Krezel, A.; Maret, W. Zinc-buffering capacity of eukaryotic cells at physiological Zn. J. Biol. Inorg. Chem. 2006, 11, 1049–1062. [Google Scholar] [CrossRef]
- Andrews, G.K. Cellular zinc sensors: MTF-1 regulation of gene expression. BioMetals 2001, 14, 223–237. [Google Scholar] [CrossRef]
- Wessels, I.; Rink, L. Micronutrients in autoimmune diseases: Possible therapeutic benefits of zinc and vitamin D. J. Nutr. Biochem. 2020, 77, 108240. [Google Scholar] [CrossRef]
- Cousins, R.J.; Lichten, L.A. Zinc transporters. In Zinc in Human Health, 1st ed.; Rink, L., Ed.; IOS Press: Amsterdam, The Netherlands, 2011; pp. 163–194. [Google Scholar]
- Ishida, T.; Takechi, S. ß-Naphthoflavone, an exogenous ligand of aryl hydrocarbon receptor, disrupts zinc homeostasis in human hepatoma HepG2 cells. J. Toxicol. Sci. 2019, 44, 711–720. [Google Scholar] [CrossRef]
- King, J.C.; Brown, K.H.; Gibson, R.S.; Krebs, N.F.; Lowe, N.M.; Siekmann, J.H.; Raiten, D.J. Biomarkers of nutrition for development (BOND)-Zinc review. J. Nutr. 2015, 146, 858S–885S. [Google Scholar] [CrossRef]
- Hamibdge, K.M.; Miller, L.V.; Westcott, J.E.; Sheng, X.; Krebs, N.F. Zinc bioavailability and homeostasis. Am. J. Clin. Nutr. 2010, 91, 1478S–1483S. [Google Scholar] [CrossRef]
- Grüngreiff, K.; Reinhold, D. Zink: Bedeutung in der Ärztlichen Praxis, 1st ed.; Jürgen Hartmann Verlag: Heßdorf-Klebheim, Germany, 2007; pp. 1–96. [Google Scholar]
- Lönnerdal, B. Dietary factors influencing zinc absorption. J. Nutr. 2000, 130, 1378S–1383S. [Google Scholar] [CrossRef]
- Maret, W.; Sandstaedt, H.H. Zinc requirements and risks and benefits of zinc supplementation. J. Trace Elem. Med. Biol. 2006, 20, 3–18. [Google Scholar] [CrossRef]
- Maret, W. (King’s College London, London, UK). Personal communication, 2020.
- Saunders, A.V.; Craig, W.J.; Baines, S.K. Zinc in vegetarian diets. Med. J. Aust. 2013, 199, 17–22. [Google Scholar] [CrossRef]
- Gibson, R.S. Zinc nutrition in developing countries. Nutr. Res. Rev. 1994, 7, 151–173. [Google Scholar] [CrossRef]
- Hahn, A.; Ströhle, A.; Wolters, M. Ernährung: Physiologische Grundlagen, Prävention, Therapie, 3rd ed.; Wissenschaftliche Verlagsgesellschaft Stuttgart: Stuttgart, Germany, 2016; pp. 1–1182. [Google Scholar]
- Udechukuwu, M.C.; Collins, S.A.; Udenigwe, C. Prospects of enhancing dietary zinc bioavailability with food-derived zinc-chelating peptides. Food Funct. 2016, 7, 4137–4144. [Google Scholar] [CrossRef]
- Trame, S.; Wessels, I.; Haase, H.; Rink, L. A short 18 items food frequeny questionaire biochemically validated to estimate zinc statur in human. J. Trace Elem. Med. Biol. 2018, 49, 285–295. [Google Scholar] [CrossRef]
- Hunt, J.R. Biovailabilty of iron, zinc, and other trace minerals from vegetarian diets. Am. J. Clin. Nutr. 2003, 78, 633S–639S. [Google Scholar] [CrossRef]
- Turnlund, J.R.; King, J.C.; Keyes, W.R.; Gong, B.; Michel, M.C. A stable isotope study of zinc absorption in young men: Effects of phytate and α-cellulose. Am. J. Clin. Nutr. 1982, 40, 1071–1077. [Google Scholar] [CrossRef]
- Hunt, J.R.; Johnson, L.K.; Lykken, G.I. High- versus low-meat diets: Effects on zinc absorption, iron status, and calcium, copper, iron, magnesium, manganese, nitrogen, phosphorus, and zinc balance in postmenopausal women. Am. J. Clin. Nutr. 1995, 62, 621–632. [Google Scholar] [CrossRef]
- Tran, C.D.; Miller, L.V.; Krebs, N.F.; Lei, S.; Hambidge, K.M. Zinc absorption as a function of the dose of zinc sulfate in aequeous solution. Am. J. Clin. Nutr. 2004, 80, 1570–1573. [Google Scholar] [CrossRef]
- Rose, S.D.; Strombom, A.J. Diverticular disease risk reduced with a plant-based diet. Adv. Res. Gastroenterol. Hepatol. 2019, 14, 32–34. [Google Scholar]
- Skrikumar, T.S.; Johansson, G.K.; Öckerman, P.; Gustafsson, P.; Äkesson, B. Trace element status in healthy subjects switching from a mixed to a lactovegetarian diet for 12 months. Am. J. Clin. Nutr. 1992, 55, 885–890. [Google Scholar] [CrossRef]
- World Health Organization. Trace Elements in Human Nutrition and Health, 1st ed.; WHO: Geneva, Switzerland, 1996; pp. 1–361. [Google Scholar]
- Gibson, R.S.; Hess, S.Y.; Hotz, C.; Brown, K.H. Indicators of zinc status at the population level: A review of the evidence. Br. J. Nutr. 2008, 99, S14–S23. [Google Scholar] [CrossRef]
- Grüngreiff, K. Zink in der Pathogenese chronischer Lebererkrankungen. Z. Gastroenterol. 2018, 56, 1301–1302. [Google Scholar]
- Grüngreiff, K. Zinc and the Liver, 1st ed.; Dr. Falk Pharma GmbH: Freiburg, Germany, 2013; pp. 1–84. [Google Scholar]
- Prasad, A.S.; Miale, A.; Farid, Z.; Sandstaed, H.H.; Schulert, A.R.; Darby, W.J. Biochemical studies of dwarfism, hypogonadism and anemia. Arch. Int. Med. 1963, 111, 407–428. [Google Scholar] [CrossRef]
- Garnica, A.D. Trace elements and hemoglobin metabolism. Ann. Clin. Lab. Sci. 1981, 11, 2220–2228. [Google Scholar]
- Siyame, E.W.; Hurst, R.; Waver, A.A.; Young, S.D.; Broadley, M.R.; Chilimba, A.D.; Ander, L.E.; Watts, M.J.; Chilima, B.; Gondwe, J.; et al. A high prevalence of zinc- but not iron-deficiency among women in rural Malawi: A cross-sectional study. Int. J. Vitam. Nutr. Res. 2013, 83, 176–186. [Google Scholar] [CrossRef]
- Spivak, J.L.; Fischer, J.; Isaacs, M.A.; Hankins, W.D. Protein kinases and phosphatases are involved in erythropoetin-mediated signal transduction. Expt. Hematol. 1992, 20, 500–504. [Google Scholar]
- Clair, J.; Talwakar, M.; McClain, R.J. Selective removal of zinc from cell media. J. Trace Elem. Exp. Med. 1995, 7, 143–150. [Google Scholar]
- Hennig, B.; Toborek, M.; McClain, C.J. Antiatherogenic properties of zinc: Implications in endothelial cell metabolism. J. Clin. Nutr. 1996, 12, 711–717. [Google Scholar] [CrossRef]
- Hennig, B.; Wang, Y.; Ramasamy, S.; McClain, C.J. Zinc deficiency alters barrier function of cultured porcine endothelial cells. J. Nutr. 1992, 122, 1242–1247. [Google Scholar] [CrossRef]
- Hennig, B.; McClain, C.J.; Wang, Y.; Ramasamy, S. Zinc protects against linoleic acid-induced disruption of endothelial barrier function in culture. J. Am. Coll. Nutr. 1990, 9, 535–538. [Google Scholar]
- Connell, P.; Young, V.M.; Toborek, M.; Cohen, D.A.; Barve, S.; McClain, C.J.; Hennig, B. Zinc attenuates tumor necrosis factor-mediated activation of transcription factors in endothelial cells. J. Am. Coll. Nutr. 1997, 16, 411–417. [Google Scholar] [CrossRef]
- Geng, Y.I. Molecular signal transduction in vascular cell apoptosis. Cell Res. 2001, 11, 253–264. [Google Scholar] [CrossRef]
- Beattie, J.H.; Kwun, I.S. Is zinc deficiency a risk factor for atherosclerosis? Br. J. Nutr. 2004, 91, 177–181. [Google Scholar] [CrossRef]
- Nakamura, H.; Sekiguchi, A.; Ogawa, Y.; Kawamura, T.; Akai, R.; Iwawaki, T.; Makiguchi, T.; Yokoo, S.; Ishikawa, O.; Motegi, S.I. Zinc deficiency exacerbates pressure ulcers by increasing oxidative stress and ATP in the skin. J. Dermatol. Sci. 2019, 95, 62–69. [Google Scholar] [CrossRef]
- Karadas, S.; Sayın, R.; Aslan, M.; Gonullu, H.; Katı, C.; Dursun, R.; Duran, L.; Gonullu, E.; Demir, H. Serum levels of trace elements and heavy metals in patients with acute hemorrhagic stroke. J. Membr. Biol. 2014, 247, 175–180. [Google Scholar] [CrossRef]
- Munshi, A.; Babu, S.; Kaul, S.; Shafi, G.; Rajeswar, K.; Alladi, S.; Jyothy, A. Depletion of serum zinc in ischemic stroke patients. Methods Find. Exp. Clin. Pharmacol. 2010, 32, 433–436. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, J.; Zhang, Y.; Li, H.; Zhang, H.; Huo, Y.; Li, J.; Liu, X.; Wang, X.; Qin, X.; et al. Baseline plasma zinc and risk of first stroke in hypertensive patients. A nested case-control study. Stroke 2019, 50, 3255–3258. [Google Scholar] [CrossRef]
- Khorsadi, H.; Nikpayam, O.; Yousefi, R.; Parandoosh, M.; Hosseinzadeh, N.; Saidpour, A.; Ghorbani, A. Zinc supplementation improves body weight management, inflammatory biomarkers and insulin resistance in individuals with obesity: A randomized, placebo-controlled, double-blind trial. Diabetol. Metab. Syndr. 2019, 11, 101–112. [Google Scholar] [CrossRef]
- Grüngreiff, K. Non-alcoholic Fatty Liver Disease, Diabetes mellitus, and Zinc/Zinc Transporters: There is a Connection? In Liver Research and Clinical Management, 1st ed.; Rodrigo, L., Ed.; Tech Open Ltd.: London, UK, 2018; pp. 43–53. [Google Scholar]
- Qi, Z.; Liu, K.J. The interaction of zinc and the blood-brain- barrier under physiological and ischemic conditions. Toxicol. Appl. Pharmacol. 2019, 364, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Morais, J.B.S.; Severo, J.S.; Beserra, J.B.; Soares de Oiveira, A.R.; Climaco Cruz, K.J.; de Sousa Melo, S.R.; Ribeiro do Nascimento, G.V.; Soares de Macedo, G.F.; do Nascimento Marreiro, D. Association between cortisol, insulin resistance and zinc in obesity: A Mini-review. Biol. Trace Elem. Res. 2019, 191, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Bury, N.R.; Chung, N.J.; Sturm, A.; Walker, P.A.; Hogstrand, C. Cortisol stimulates the zinc signaling pathway and expression of metallothioneins and Znt1 in rainbow trout gill epithelial cells. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Olechnowitz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zinc status is associated with inflammation, oxidative stress, lipid, and glucose metabolism. J. Physiol. Sci. 2018, 68, 19–31. [Google Scholar] [CrossRef]
- Ferro, F.E.D.; de Sousa Lima, V.B.; Mello Soares, N.R.; Ma Francisato Cozzolino, S.; do Nascimento Marreiro, D. Biomarkers of metabolic syndrome and its relashionship with zinc nutrional status in obese women. Nutr. Hosp. 2011, 650–654. [Google Scholar]
- Gu, K.; Xiang, W.; Zhang, Y.; Sun, K.; Jiang, X. The association between zinc level and overweight/obesity: A meta-analysis. Eur. J. Nutr. 2019, 58, 2971–2982. [Google Scholar] [CrossRef] [PubMed]
- Iso, H.; Stampfer, M.J.; Manson, J.E.; Rexrode, K.; Hu, F.; Hennekens, C.H.; Colditz, G.A.; Speizer, F.E.; Willett, W.C. Prospective study of fat and protein intake and risk of intraparenchymal hemorrhage in women. Circulation 2001, 103, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Keys, A.; Parlin, R.W. Serum cholesterol response to changes in dietary lipids. Am. J. Clin. Nutr. 1966, 19, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Mensink, R.P.; Katan, M.B. Effect of dietary trans fatty acids on high-density and low-density lipoprotein cholesterol levels in healthy subjects. N. Engl. J. Med. 1990, 323, 439–445. [Google Scholar] [CrossRef]
- Arleth, T.; Olsen, M.H.; Orre, M.; Rasmussen, R.; Bache, S.; Eskesen, V.; Frikke-Schmidt, R.; Møller, K. Hypozincaemia is associated with severity of subarachnoid hemorrhage: A retrospective cohort study. Acta Neurochir. 2020, 162, 1417–1424. [Google Scholar]
- Hong, K.H.; Keen, C.L.; Mizuno, Y.; Johnston, K.E.; Tamura, T. Effects of dietary zinc deficiency on homocysteine and folate metabolism. J. Nutr. Biochem. 2000, 11, 165–169. [Google Scholar] [CrossRef]
- Barbato, J.C.; Catanescu, O.; Murray, K.; DiBello, P.M.; Jacobsen, D.W. Targeting of metallothionein by L-homocysteine: A novel mechanism for disruption of zinc and redox homeostasis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Jing, M.; Rech, L.; Wu, Y.; Goltz, D.; Taylor, C.G.; House, J.D. Effects of zinc deficiency and zinc supplementation on homocysteine levels and related enzyme expression in rats. J. Trace Elem. Med. Biol. 2015, 30, 77–82. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grüngreiff, K.; Gottstein, T.; Reinhold, D. Zinc Deficiency—An Independent Risk Factor in the Pathogenesis of Haemorrhagic Stroke? Nutrients 2020, 12, 3548. https://doi.org/10.3390/nu12113548
Grüngreiff K, Gottstein T, Reinhold D. Zinc Deficiency—An Independent Risk Factor in the Pathogenesis of Haemorrhagic Stroke? Nutrients. 2020; 12(11):3548. https://doi.org/10.3390/nu12113548
Chicago/Turabian StyleGrüngreiff, Kurt, Thomas Gottstein, and Dirk Reinhold. 2020. "Zinc Deficiency—An Independent Risk Factor in the Pathogenesis of Haemorrhagic Stroke?" Nutrients 12, no. 11: 3548. https://doi.org/10.3390/nu12113548
APA StyleGrüngreiff, K., Gottstein, T., & Reinhold, D. (2020). Zinc Deficiency—An Independent Risk Factor in the Pathogenesis of Haemorrhagic Stroke? Nutrients, 12(11), 3548. https://doi.org/10.3390/nu12113548