Anti-Obesity Effects of a Prunus persica and Nelumbo nucifera Mixture in Mice Fed a High-Fat Diet


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Animals and Experimental Design
2.3. Serum Biochemical Analysis
2.4. Determination of Hepatic Lipid Content
2.5. Real-Time Polymerase Chain Reaction (PCR) Analysis
2.6. Statistical Analysis
3. Results
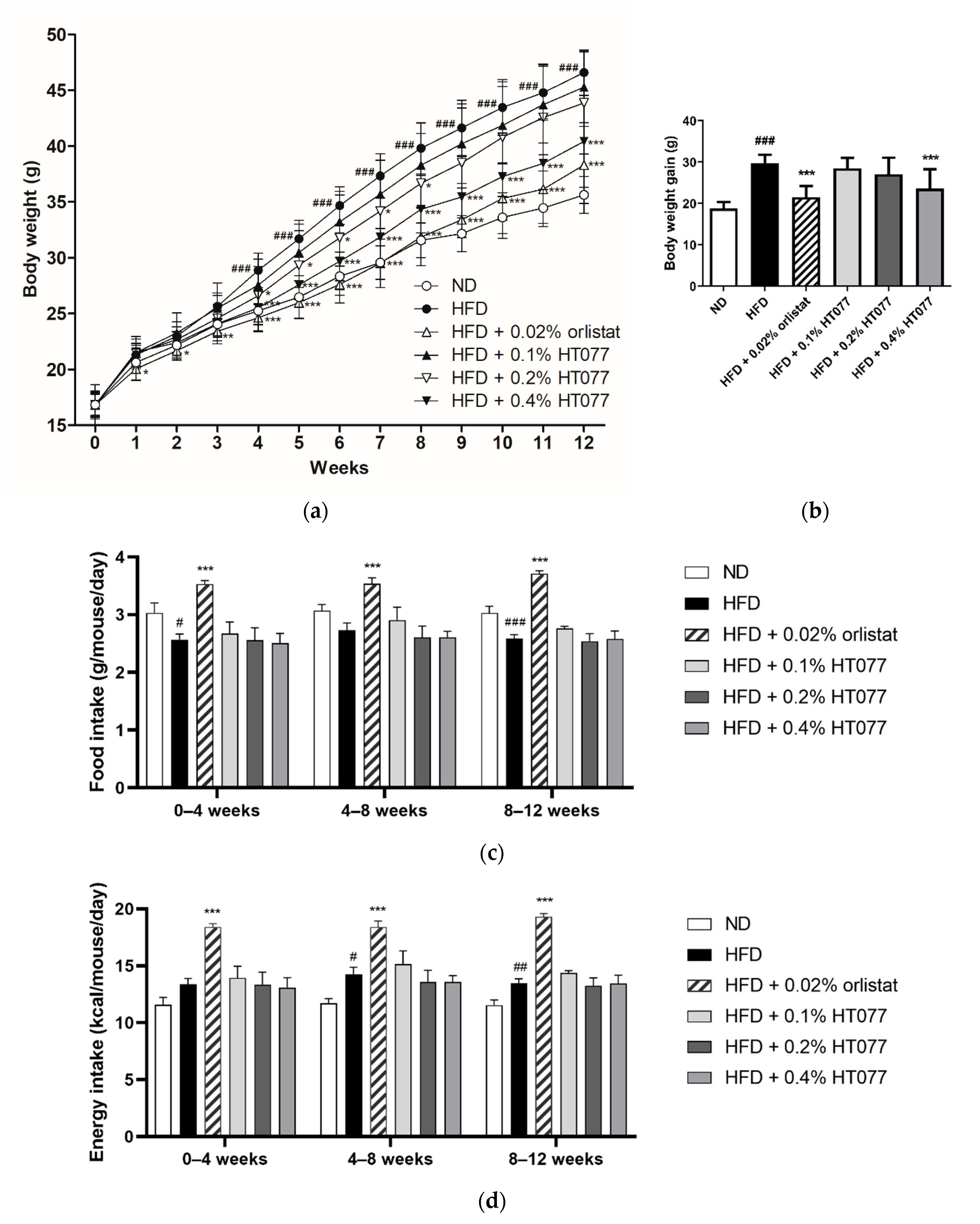
3.1. Effects of HT077 on Body Weight and Food Consumption
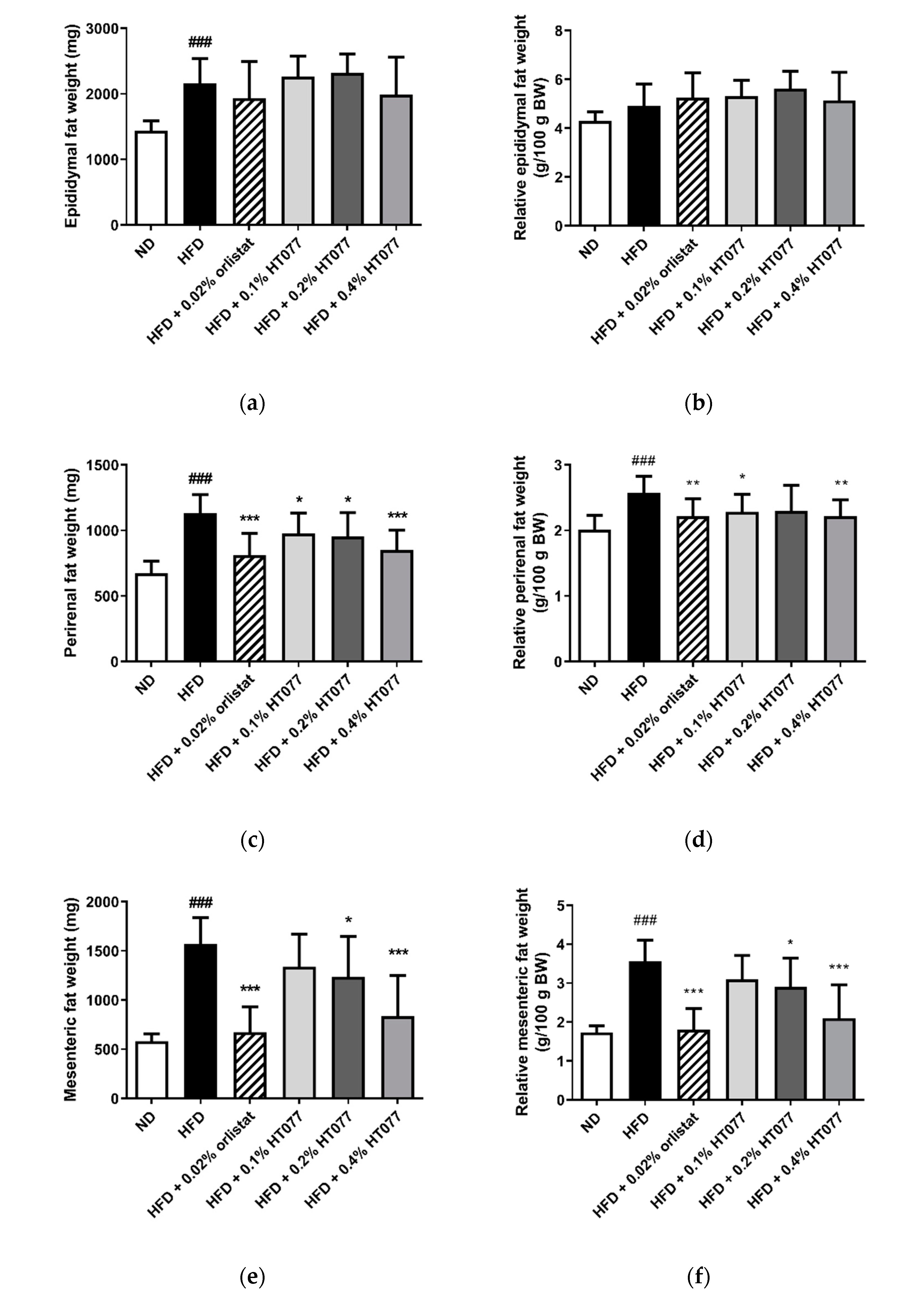
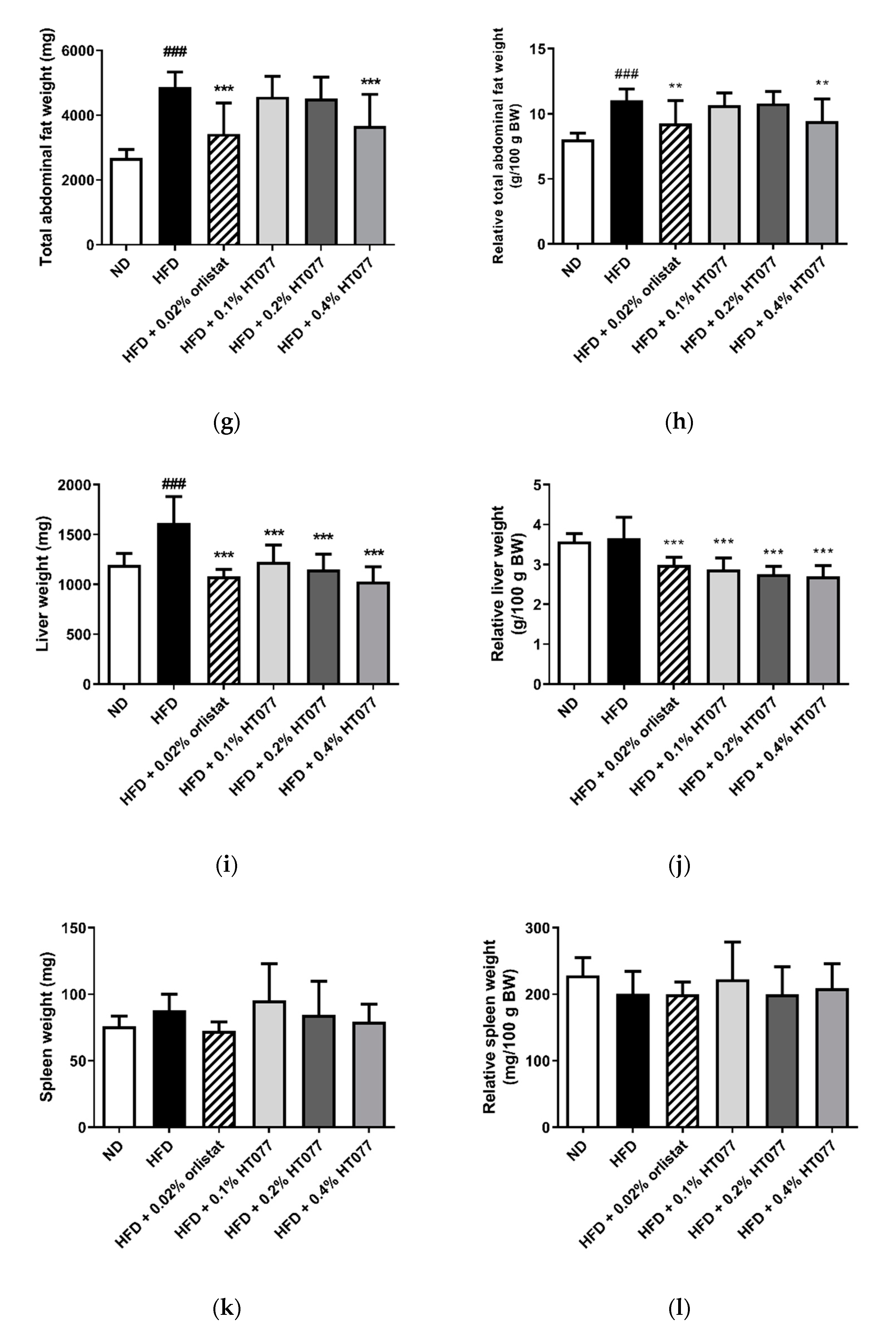
3.2. Effects of HT077 on Abdominal Fat, Liver, and Spleen Wet Weights
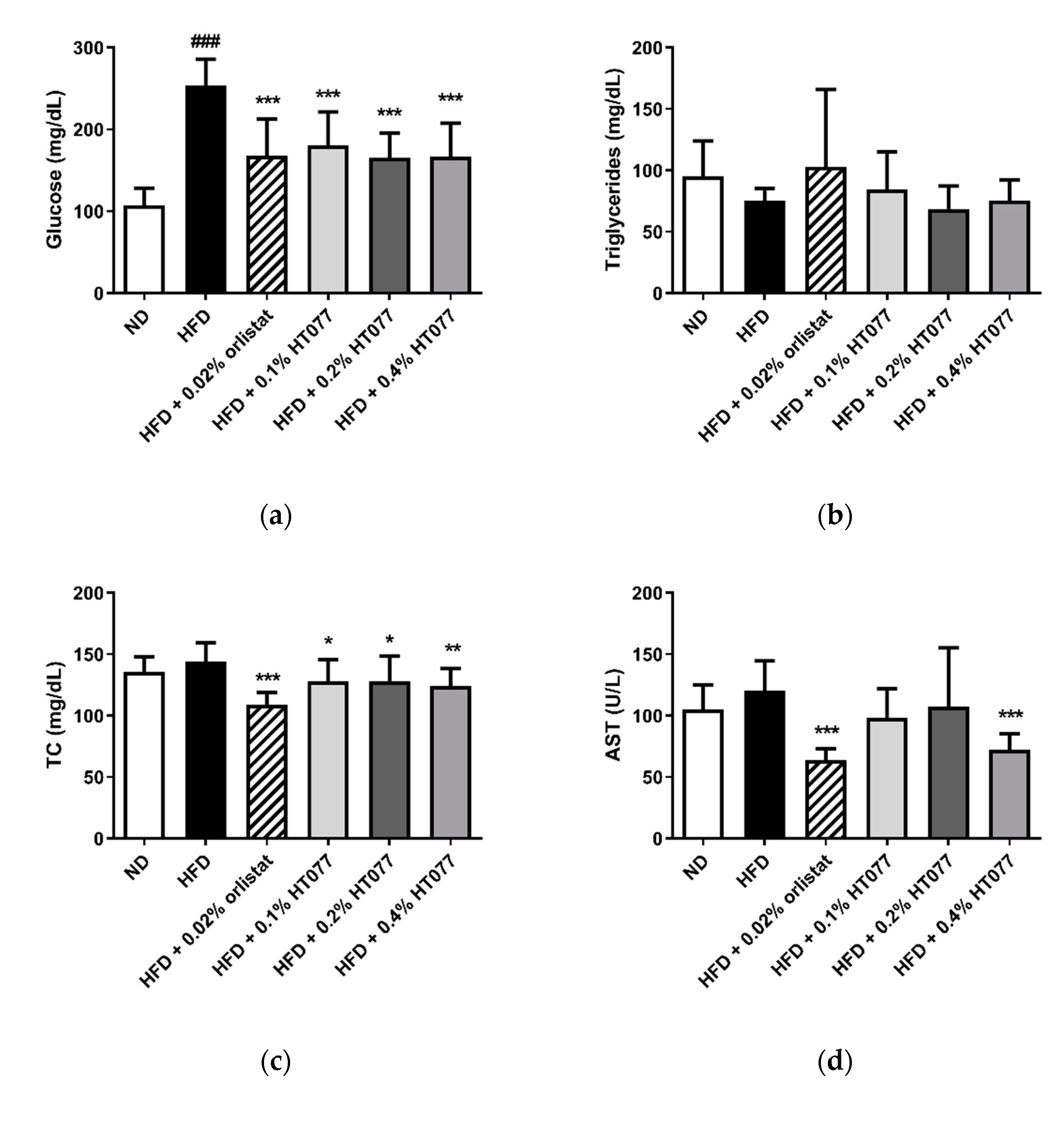
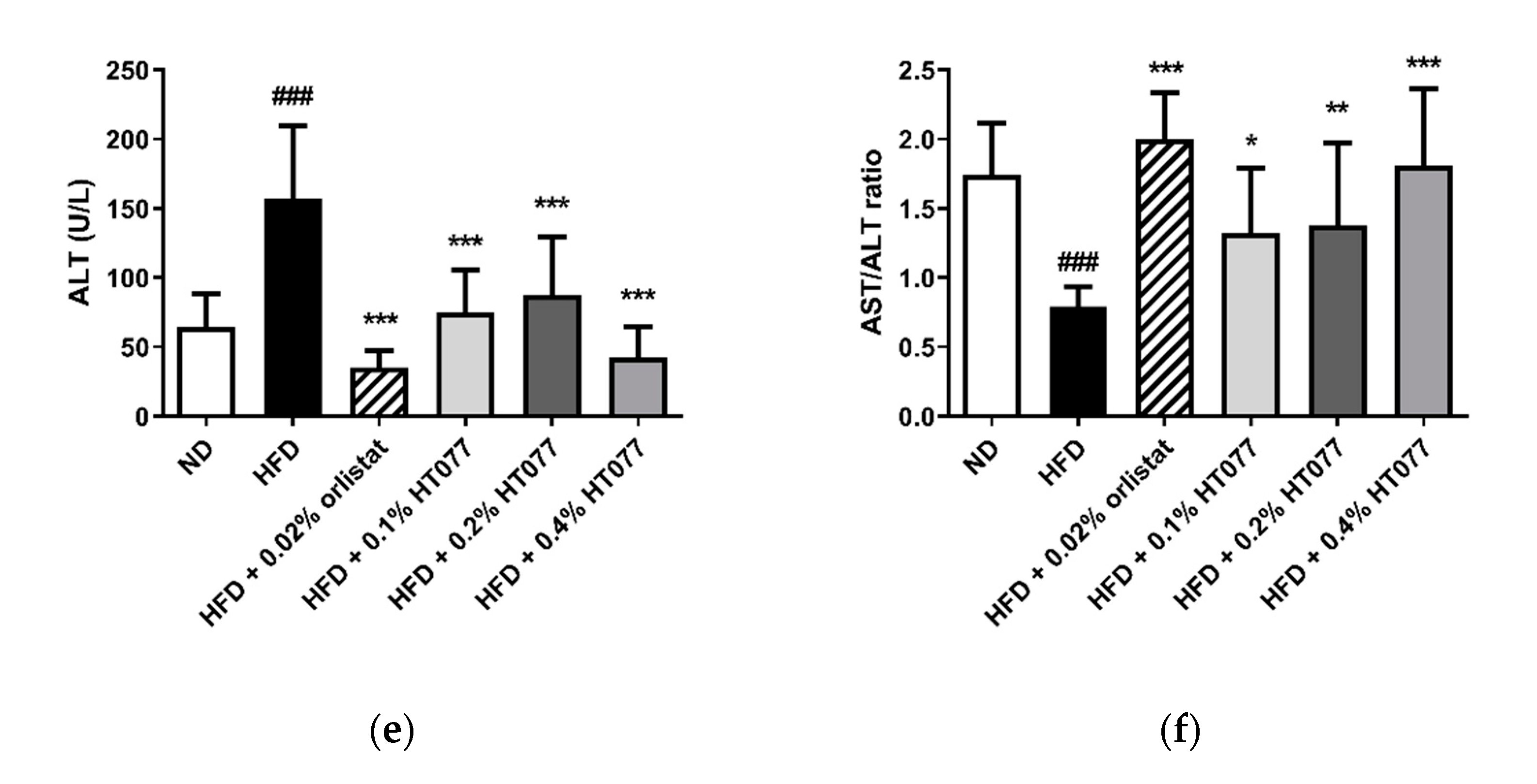
3.3. Effects of HT077 on Serum Levels of Glucose, Lipids, and Liver Enzymes
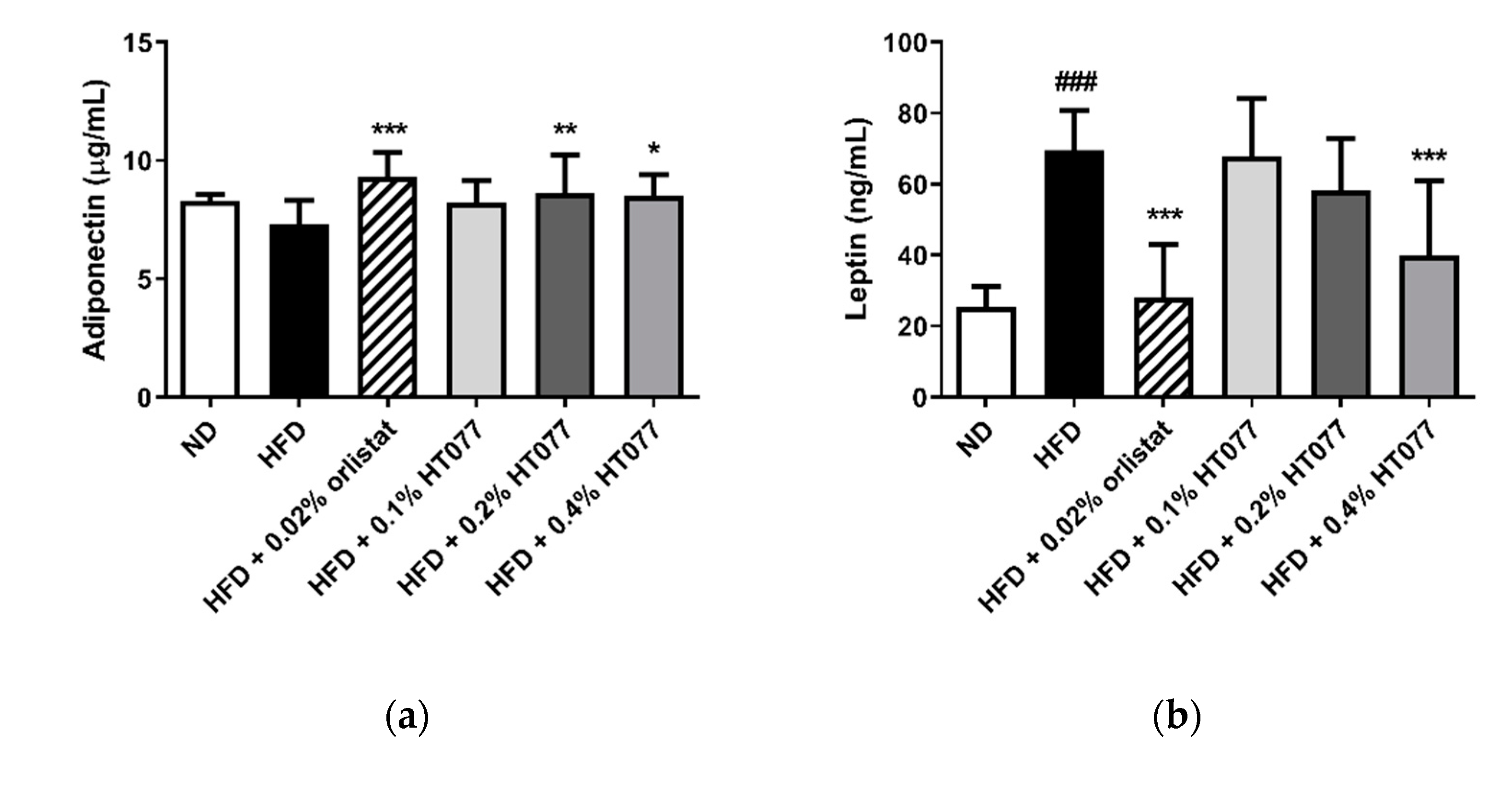
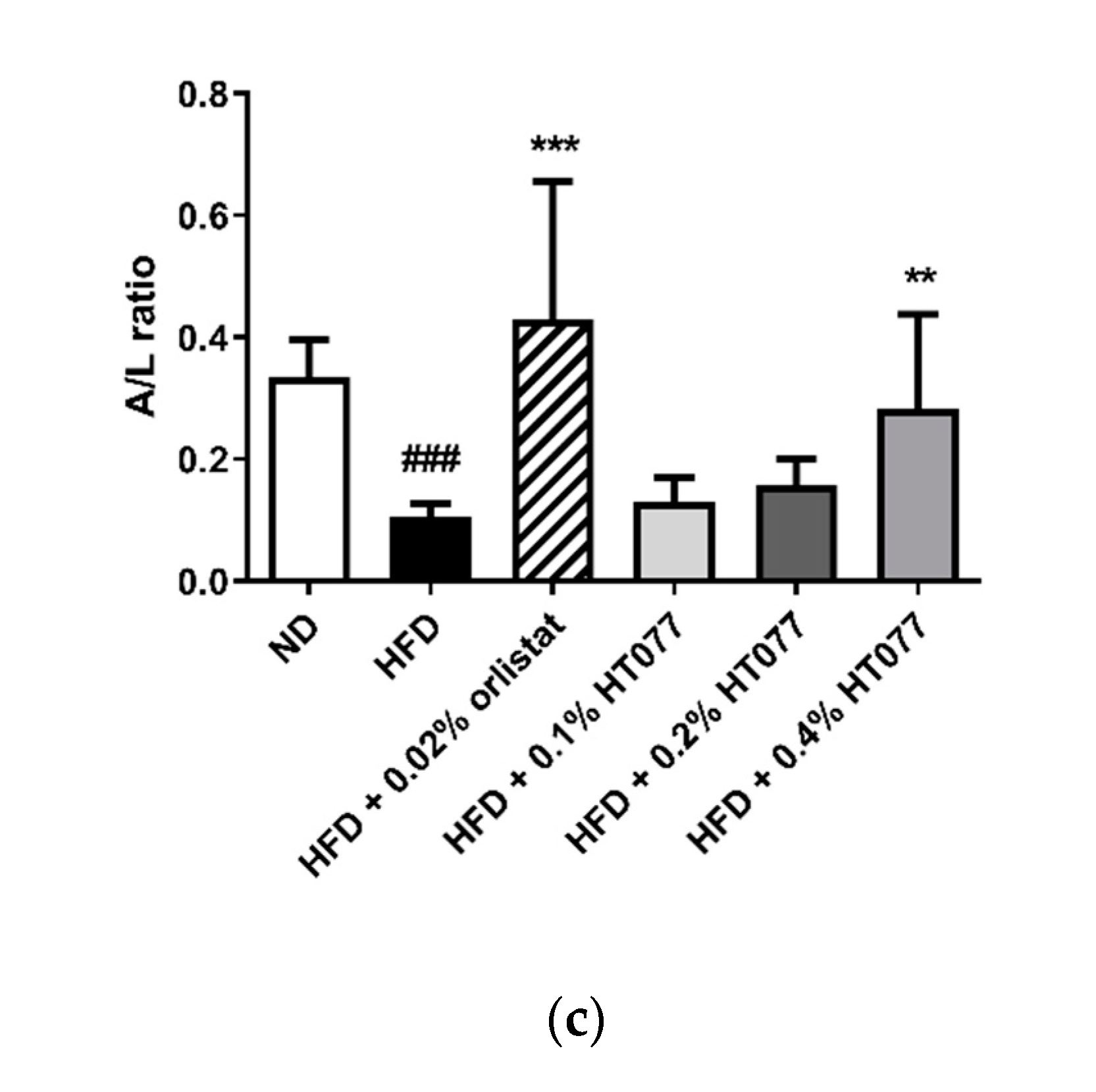
3.4. Effects of HT077on Serum Adipokine Concentrations
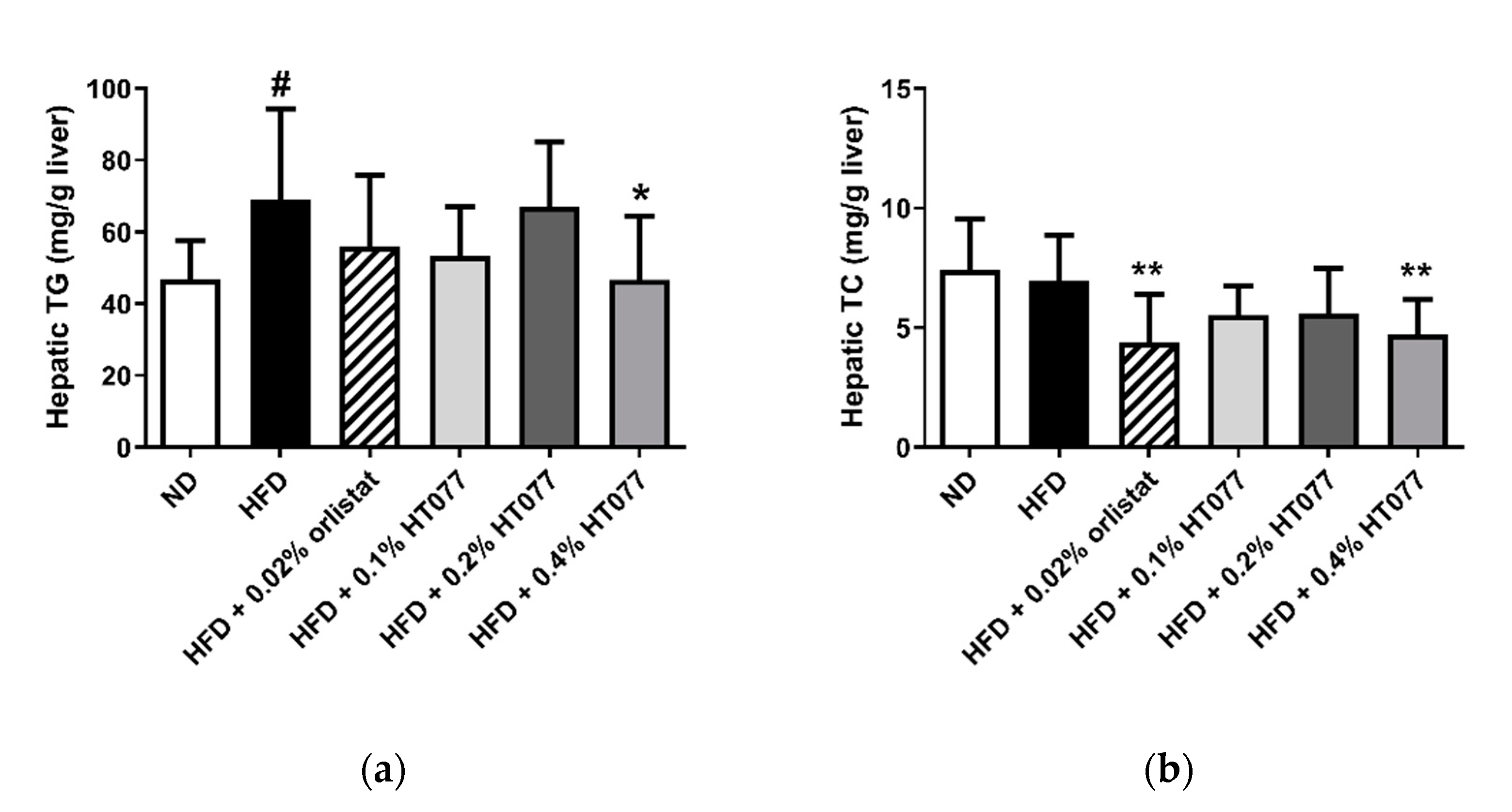
3.5. Effects of HT077on Hepatic Triglyceride and Total Cholesterol Contents
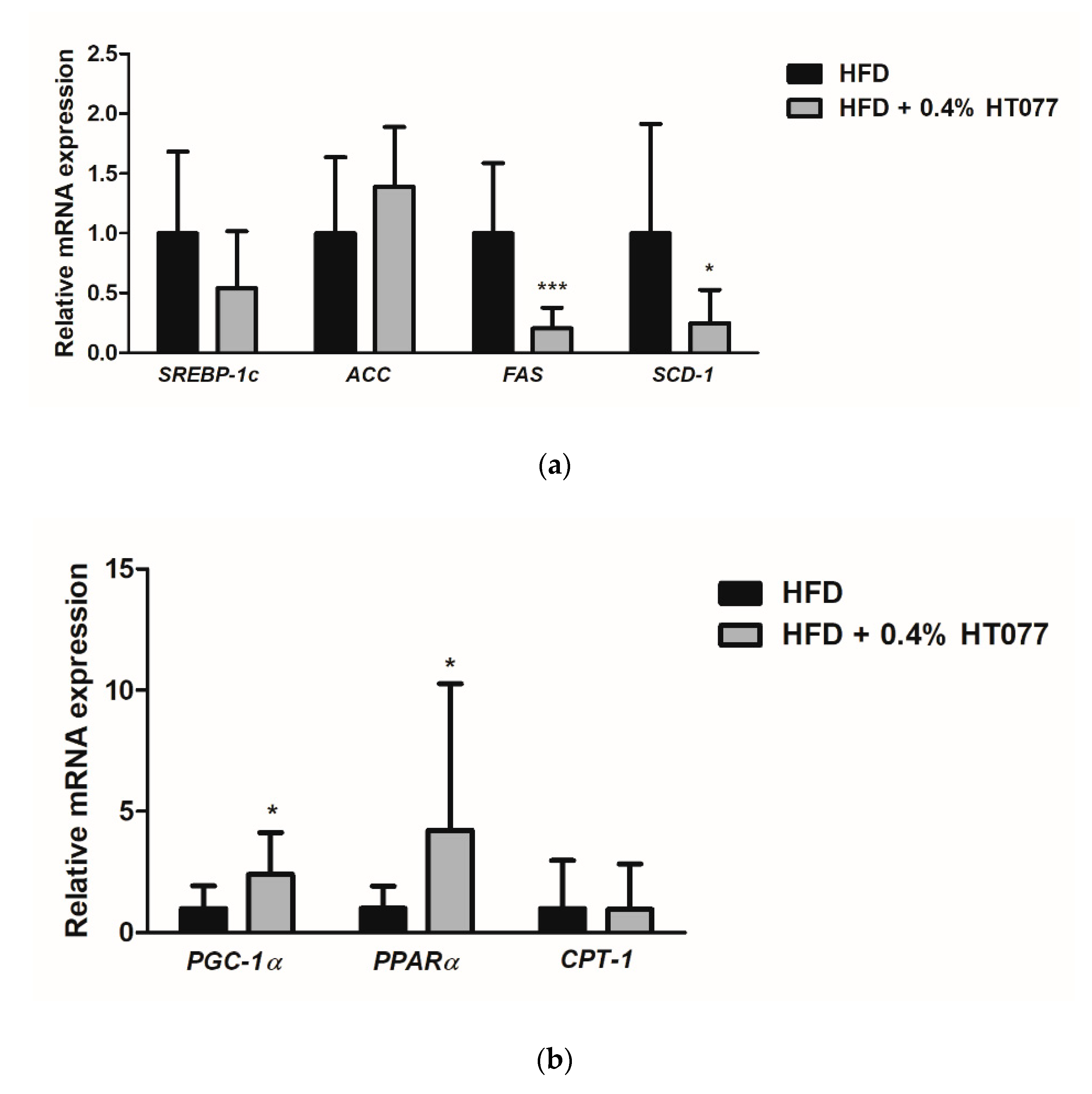
3.6. Effects of HT077on Lipogenesis and Fatty Acid Oxidation-Related Gene Expression in the Adipose Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Collaborators, G.B.D.O.; Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; et al. Health effects of overweight and obesity in 195 countries over 25 years. New Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef]
- Pi-Sunyer, X. The medical risks of obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.N.; Nam, S.S.; Park, Y.K. Effect on 12-week intensive dietary and exercise program on weight reduction and maintenance in obese women with weight cycling history. Clin. Nutr. Res. 2017, 6, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Apovian, C.M.; Aronne, L.J.; Bessesen, D.H.; McDonnell, M.E.; Murad, M.H.; Pagotto, U.; Ryan, D.H.; Still, C.D.; Endocrine, S. Pharmacological management of obesity: An endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2015, 100, 342–362. [Google Scholar] [CrossRef] [PubMed]
- Karri, S.; Sharma, S.; Hatware, K.; Patil, K. Natural anti-obesity agents and their therapeutic role in management of obesity: A future trend perspective. Biomed. Pharmacother. 2019, 110, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Chen, X.X.; Tang, S.C.W.; Lao, L.X.; Sze, S.C.W.; Lee, K.F.; Zhang, K.Y.B. Edible plants from traditional Chinese medicine is a promising alternative for the management of diabetic nephropathy. J. Funct. Foods 2015, 14, 12–22. [Google Scholar] [CrossRef]
- Scotti, L.; Monteiro, A.F.M.; de Oliveira Viana, J.; Mendonca Junior, F.J.B.; Ishiki, H.M.; Tchouboun, E.N.; Santos, R.; Scotti, M.T. Multi-target drugs against metabolic disorders. Endocr. Metab. Immune Disord. Drug Targets 2019, 19, 402–418. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Kim, Y.S.; Kim, L.; Park, H.J.; Lee, D.; Kim, H. Anti-obesity effects of the flower of Prunus persica in high-fat diet-induced obese mice. Nutrients 2019, 11, 2176. [Google Scholar] [CrossRef]
- Kim, H.; Song, J.; Lee, Y. Compositions for Preventing, Improving or Treating Obesity and Metabolic Diseases Comprising Combined Extracts of Peach Blossom and Lotus Leaf. KR patent registration number 10-1886299, 1 August 2018. [Google Scholar]
- Han, W.; Xu, J.D.; Wei, F.X.; Zheng, Y.D.; Ma, J.Z.; Xu, X.D.; Wei, Z.G.; Wang, W.; Zhang, Y.C. Prokinetic activity of Prunus persica (L.) Batsch flowers extract and its possible mechanism of action in rats. Biomed. Res. Int. 2015, 2015, 569853. [Google Scholar] [CrossRef]
- Kwak, C.S.; Yang, J. Prevention effect of Prunus persica Flos extract from reactive oxygen species generation and matrix metalloproteinases production induced by UVB irradiation in human skin cells. Asian J. Beauty Cosmetol. 2016, 14, 179–190. [Google Scholar] [CrossRef]
- Kim, Y.H.; Yang, H.E.; Park, B.K.; Heo, M.Y.; Jo, B.K.; Kim, H.P. The extract of the flowers of Prunus persica, a new cosmetic ingredient, protects against solar ultraviolet-induced skin damage in vivo. J. Cosmet. Sci. 2002, 53, 27–34. [Google Scholar] [PubMed]
- Kwak, C.S.; Yang, J.; Shin, C.Y.; Chung, J.H. Topical or oral treatment of peach flower extract attenuates UV-induced epidermal thickening, matrix metalloproteinase-13 expression and pro-inflammatory cytokine production in hairless mice skin. Nutr. Res. Pract. 2018, 12, 29–40. [Google Scholar] [CrossRef]
- Takagi, S.; Yamaki, M.; Masuda, K.; Kubota, M.; Minami, J. Studies on the purgative drugs. III. On the constituents of the flowers of Prunus persica Batsch. Yakugaku Zasshi 1977, 97, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.C.; Jiao, Z.G.; Yang, W.B.; Zhang, C.L.; Liu, H.; Lv, Z.Z. Variation in phenolics, flavanoids, antioxidant and tyrosinase inhibitory activity of peach blossoms at different developmental stages. Molecules 2015, 20, 20460–20472. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Takahashi, K.; Nakamura, H.; Itoh, K.; Matsuda, H. Search for skin-whitening agent from Prunus plants and the molecular targets in melanogenesis pathway of active compounds. Nat. Prod. Commun. 2014, 9, 185–188. [Google Scholar] [CrossRef]
- Ono, Y.; Hattori, E.; Fukaya, Y.; Imai, S.; Ohizumi, Y. Anti-obesity effect of Nelumbo nucifera leaves extract in mice and rats. J. Ethnopharmacol. 2006, 106, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, D.; Huang, B.; Chen, Y.; Lu, X.; Wang, Y. Inhibition of pancreatic lipase, alpha-glucosidase, alpha-amylase, and hypolipidemic effects of the total flavonoids from Nelumbo nucifera leaves. J. Ethnopharmacol. 2013, 149, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.H.; He, X.X.; You, C.; Tao, X.; Wang, L.S.; Zhang, M.D.; Zhou, Y.F.; Chang, Q. Pharmacokinetics of nuciferine and n-nornuciferine, two major alkaloids from Nelumbo nucifera leaves, in rat plasma and the brain. Front. Pharmacol. 2018, 9, 902. [Google Scholar] [CrossRef]
- Wang, M.X.; Liu, Y.L.; Yang, Y.; Zhang, D.M.; Kong, L.D. Nuciferine restores potassium oxonate-induced hyperuricemia and kidney inflammation in mice. Eur. J. Pharmacol. 2015, 747, 59–70. [Google Scholar] [CrossRef]
- Yan, M.Z.; Chang, Q.; Zhong, Y.; Xiao, B.X.; Feng, L.; Cao, F.R.; Pan, R.L.; Zhang, Z.S.; Liao, Y.H.; Liu, X.M. Lotus leaf alkaloid extract displays sedative-hypnotic and anxiolytic effects through GABAA receptor. J. Agric. Food Chem. 2015, 63, 9277–9285. [Google Scholar] [CrossRef]
- Do, T.C.; Nguyen, T.D.; Tran, H.; Stuppner, H.; Ganzera, M. Analysis of alkaloids in Lotus (Nelumbo nucifera Gaertn.) leaves by non-aqueous capillary electrophoresis using ultraviolet and mass spectrometric detection. J. Chromatogr. A 2013, 1302, 174–180. [Google Scholar] [CrossRef]
- Bagchi, D.P.; MacDougald, O.A. Identification and dissection of diverse mouse adipose depots. J. Vis. Exp. 2019, 149, e59499. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Melhorn, S.J.; Krause, E.G.; Scott, K.A.; Mooney, M.R.; Johnson, J.D.; Woods, S.C.; Sakai, R.R. Acute exposure to a high-fat diet alters meal patterns and body composition. Physiol. Behav. 2010, 99, 33–39. [Google Scholar] [CrossRef]
- Chusyd, D.E.; Wang, D.; Huffman, D.M.; Nagy, T.R. Relationships between rodent white adipose fat pads and human white adipose fat depots. Front. Nutr. 2016, 3, 10. [Google Scholar] [CrossRef]
- Hall, J.E.; do Carmo, J.M.; da Silva, A.A.; Wang, Z.; Hall, M.E. Obesity, kidney dysfunction and hypertension: Mechanistic links. Nat. Rev. Nephrol. 2019, 15, 367–385. [Google Scholar] [CrossRef]
- Ricci, M.A.; Scavizzi, M.; Ministrini, S.; De Vuono, S.; Pucci, G.; Lupattelli, G. Morbid obesity and hypertension: The role of perirenal fat. J. Clin. Hypertens. (Greenwich) 2018, 20, 1430–1437. [Google Scholar] [CrossRef]
- Koo, B.K.; Denenberg, J.O.; Wright, C.M.; Criqui, M.H.; Allison, M.A. Associations of perirenal fat thickness with renal and systemic calcified atherosclerosis. Endocrinol. Metab. (Seoul) 2020, 35, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Chan, Y.L.; Chan, J.C.; Chan, W.B.; Kong, W.L. Mesenteric fat thickness as an independent determinant of fatty liver. Int. J. Obes. (London) 2006, 30, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Wueest, S.; Item, F.; Lucchini, F.C.; Challa, T.D.; Muller, W.; Bluher, M.; Konrad, D. Mesenteric fat lipolysis mediates obesity-associated hepatic steatosis and insulin resistance. Diabetes 2016, 65, 140–148. [Google Scholar] [CrossRef]
- Ellrichmann, M.; Kapelle, M.; Ritter, P.R.; Holst, J.J.; Herzig, K.H.; Schmidt, W.E.; Schmitz, F.; Meier, J.J. Orlistat inhibition of intestinal lipase acutely increases appetite and attenuates postprandial glucagon-like peptide-1-(7-36)-amide-1, cholecystokinin, and peptide YY concentrations. J. Clin. Endocrinol. Metab. 2008, 93, 3995–3998. [Google Scholar] [CrossRef]
- Guo, J.; Jou, W.; Gavrilova, O.; Hall, K.D. Persistent diet-induced obesity in male C57BL/6 mice resulting from temporary obesigenic diets. PLoS ONE 2009, 4, e5370. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell. Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef]
- Fabbrini, E.; Magkos, F.; Mohammed, B.S.; Pietka, T.; Abumrad, N.A.; Patterson, B.W.; Okunade, A.; Klein, S. Intrahepatic fat, not visceral fat, is linked with metabolic complications of obesity. Proc. Natl. Acad. Sci. USA 2009, 106, 15430–15435. [Google Scholar] [CrossRef]
- Schindhelm, R.K.; Diamant, M.; Dekker, J.M.; Tushuizen, M.E.; Teerlink, T.; Heine, R.J. Alanine aminotransferase as a marker of non-alcoholic fatty liver disease in relation to type 2 diabetes mellitus and cardiovascular disease. Diabetes Metab. Res. Rev. 2006, 22, 437–443. [Google Scholar] [CrossRef]
- Sorbi, D.; Boynton, J.; Lindor, K.D. The ratio of aspartate aminotransferase to alanine aminotransferase: Potential value in differentiating nonalcoholic steatohepatitis from alcoholic liver disease. Am. J. Gastroenterol. 1999, 94, 1018–1022. [Google Scholar] [CrossRef]
- Sharma, B.R.; Oh, J.; Kim, H.A.; Kim, Y.J.; Jeong, K.S.; Rhyu, D.Y. Anti-obesity effects of the mixture of Eriobotrya japonica and Nelumbo nucifera in adipocytes and high-fat diet-induced obese mice. Am. J. Chin. Med. 2015, 43, 681–694. [Google Scholar] [CrossRef]
- Lin, M.C.; Kao, S.H.; Chung, P.J.; Chan, K.C.; Yang, M.Y.; Wang, C.J. Improvement for high fat diet-induced hepatic injuries and oxidative stress by flavonoid-enriched extract from Nelumbo nucifera leaf. J. Agric. Food Chem. 2009, 57, 5925–5932. [Google Scholar] [CrossRef]
- Wu, C.H.; Yang, M.Y.; Chan, K.C.; Chung, P.J.; Ou, T.T.; Wang, C.J. Improvement in high-fat diet-induced obesity and body fat accumulation by a Nelumbo nucifera leaf flavonoid-rich extract in mice. J. Agric. Food Chem. 2010, 58, 7075–7081. [Google Scholar] [CrossRef]
- Czech, M.P. Insulin action and resistance in obesity and type 2 diabetes. Nat. Med. 2017, 23, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Shirosaki, M.; Goto, Y.; Hirooka, S.; Masuda, H.; Koyama, T.; Yazawa, K. Peach leaf contains multiflorin a as a potent inhibitor of glucose absorption in the small intestine in mice. Biol. Pharm. Bull. 2012, 35, 1264–1268. [Google Scholar] [CrossRef][Green Version]
- Huang, C.F.; Chen, Y.W.; Yang, C.Y.; Lin, H.Y.; Way, T.D.; Chiang, W.; Liu, S.H. Extract of lotus leaf (Nelumbo nucifera) and its active constituent catechin with insulin secretagogue activity. J. Agric. Food Chem. 2011, 59, 1087–1094. [Google Scholar] [CrossRef]
- Yan, K.; Zhu, H.; Xu, J.; Pan, H.; Li, N.; Wang, L.; Yang, H.; Liu, M.; Gong, F. Lotus leaf aqueous extract reduces visceral fat mass and ameliorates insulin resistance in HFD-induced obese rats by regulating PPARgamma2 expression. Front. Pharmacol. 2017, 8, 409. [Google Scholar] [CrossRef]
- Chen, H.W.; Yang, M.Y.; Hung, T.W.; Chang, Y.C.; Wang, C.J. Nelumbo nucifera leaves extract attenuate the pathological progression of diabetic nephropathy in high-fat diet-fed and streptozotocin-induced diabetic rats. J. Food Drug Anal. 2019, 27, 736–748. [Google Scholar] [CrossRef]
- Du, H.; You, J.S.; Zhao, X.; Park, J.Y.; Kim, S.H.; Chang, K.J. Antiobesity and hypolipidemic effects of lotus leaf hot water extract with taurine supplementation in rats fed a high fat diet. J. Biomed. Sci. 2010, 17 (Suppl. 1), S42. [Google Scholar] [CrossRef]
- Fruhwurth, S.; Vogel, H.; Schurmann, A.; Williams, K.J. Novel insights into how overnutrition disrupts the hypothalamic actions of leptin. Front. Endocrinol. (Lausanne) 2018, 9, 89. [Google Scholar] [CrossRef]
- Yang, R.; Barouch, L.A. Leptin signaling and obesity: Cardiovascular consequences. Circ. Res. 2007, 101, 545–559. [Google Scholar] [CrossRef]
- Fishman, S.; Muzumdar, R.H.; Atzmon, G.; Ma, X.; Yang, X.; Einstein, F.H.; Barzilai, N. Resistance to leptin action is the major determinant of hepatic triglyceride accumulation in vivo. FASEB J. 2007, 21, 53–60. [Google Scholar] [CrossRef]
- Ziemke, F.; Mantzoros, C.S. Adiponectin in insulin resistance: Lessons from translational research. Am. J. Clin. Nutr. 2010, 91, 258S–261S. [Google Scholar] [CrossRef] [PubMed]
- Fruhbeck, G.; Catalan, V.; Rodriguez, A.; Gomez-Ambrosi, J. Adiponectin-leptin ratio: A promising index to estimate adipose tissue dysfunction. Relation with obesity-associated cardiometabolic risk. Adipocyte 2018, 7, 57–62. [Google Scholar] [CrossRef]
- Inoue, M.; Maehata, E.; Yano, M.; Taniyama, M.; Suzuki, S. Correlation between the adiponectin-leptin ratio and parameters of insulin resistance in patients with type 2 diabetes. Metabolism 2005, 54, 281–286. [Google Scholar] [CrossRef]
- Guo, F.; Yang, X.; Li, X.; Feng, R.; Guan, C.; Wang, Y.; Li, Y.; Sun, C. Nuciferine prevents hepatic steatosis and injury induced by a high-fat diet in hamsters. PLoS ONE 2013, 8, e63770. [Google Scholar] [CrossRef]
- Bodis, K.; Roden, M. Energy metabolism of white adipose tissue and insulin resistance in humans. Eur. J. Clin. Invest. 2018, 48, e13017. [Google Scholar] [CrossRef]
- Catalano, K.J.; Stefanovski, D.; Bergman, R.N. Critical role of the mesenteric depot versus other intra-abdominal adipose depots in the development of insulin resistance in young rats. Diabetes 2010, 59, 1416–1423. [Google Scholar] [CrossRef]
- Item, F.; Konrad, D. Visceral fat and metabolic inflammation: The portal theory revisited. Obes. Rev. 2012, 13 (Suppl. 2), 30–39. [Google Scholar] [CrossRef]
- Iniguez, M.; Perez-Matute, P.; Villanueva-Millan, M.J.; Recio-Fernandez, E.; Roncero-Ramos, I.; Perez-Clavijo, M.; Oteo, J.A. Agaricus bisporus supplementation reduces high-fat diet-induced body weight gain and fatty liver development. J. Physiol. Biochem. 2018, 74, 635–646. [Google Scholar] [CrossRef]
- Kim, B.; Kwon, J.; Kim, M.S.; Park, H.; Ji, Y.; Holzapfel, W.; Hyun, C.K. Protective effects of Bacillus probiotics against high-fat diet-induced metabolic disorders in mice. PLoS ONE 2018, 13, e0210120. [Google Scholar] [CrossRef]
- Lim, S.M.; Kim, E.; Shin, J.H.; Seok, P.R.; Jung, S.; Yoo, S.H.; Kim, Y. Xylobiose prevents high-fat diet induced mice obesity by suppressing mesenteric fat deposition and metabolic dysregulation. Molecules 2018, 23, 705. [Google Scholar] [CrossRef]
- Li, Z.S.; Zheng, J.W.; Manabe, Y.; Hirata, T.; Sugawara, T. Anti-obesity properties of the dietary green alga, codium cylindricum, in high-fat diet-induced obese mice. J. Nutr. Sci. Vitaminol. (Tokyo) 2018, 64, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Kim, B.; Park, H.; Ji, Y.; Holzapfel, W.; Kim, D.Y.; Hyun, C.K. Long-term fermented soybean paste improves metabolic parameters associated with non-alcoholic fatty liver disease and insulin resistance in high-fat diet-induced obese mice. Biochem. Biophys. Res. Commun. 2018, 495, 1744–1751. [Google Scholar] [CrossRef]
- Song, Z.; Xiaoli, A.M.; Yang, F. Regulation and metabolic significance of de novo lipogenesis in adipose tissues. Nutrients 2018, 10, 1383. [Google Scholar] [CrossRef]
- Berndt, J.; Kovacs, P.; Ruschke, K.; Kloting, N.; Fasshauer, M.; Schon, M.R.; Korner, A.; Stumvoll, M.; Bluher, M. Fatty acid synthase gene expression in human adipose tissue: Association with obesity and type 2 diabetes. Diabetologia 2007, 50, 1472–1480. [Google Scholar] [CrossRef]
- Jiang, G.; Li, Z.; Liu, F.; Ellsworth, K.; Dallas-Yang, Q.; Wu, M.; Ronan, J.; Esau, C.; Murphy, C.; Szalkowski, D.; et al. Prevention of obesity in mice by antisense oligonucleotide inhibitors of stearoyl-CoA desaturase-1. J. Clin. Invest. 2005, 115, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.F.; Ku, H.C.; Lin, H. PGC-1alpha as a pivotal factor in lipid and metabolic regulation. Int. J. Mol. Sci. 2018, 19, 3447. [Google Scholar] [CrossRef]
- Goto, T.; Lee, J.Y.; Teraminami, A.; Kim, Y.I.; Hirai, S.; Uemura, T.; Inoue, H.; Takahashi, N.; Kawada, T. Activation of peroxisome proliferator-activated receptor-alpha stimulates both differentiation and fatty acid oxidation in adipocytes. J. Lipid Res. 2011, 52, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef]
- Van Gaal, L.F.; Broom, J.I.; Enzi, G.; Toplak, H. Efficacy and tolerability of orlistat in the treatment of obesity: A 6-month dose-ranging study. Orlistat dose-ranging study group. Eur. J. Clin. Pharmacol. 1998, 54, 125–132. [Google Scholar] [CrossRef]
- Lodhi, I.J.; Yin, L.; Jensen-Urstad, A.P.; Funai, K.; Coleman, T.; Baird, J.H.; El Ramahi, M.K.; Razani, B.; Song, H.; Fu-Hsu, F.; et al. Inhibiting adipose tissue lipogenesis reprograms thermogenesis and PPARgamma activation to decrease diet-induced obesity. Cell Metab. 2012, 16, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Lin, L.; Li, Q.; Xue, Y.; Zheng, F.; Wang, G.; Zheng, C.; Du, L.; Hu, M.; Huang, Y.; et al. Scd1 controls de novo beige fat biogenesis through succinate-dependent regulation of mitochondrial complex II. Proc. Natl Acad Sci USA 2020, 117, 2462–2472. [Google Scholar] [CrossRef]
- Hondares, E.; Rosell, M.; Diaz-Delfin, J.; Olmos, Y.; Monsalve, M.; Iglesias, R.; Villarroya, F.; Giralt, M. Peroxisome proliferator-activated receptor alpha (PPARalpha) induces PPARgamma coactivator 1alpha (PGC-1alpha) gene expression and contributes to thermogenic activation of brown fat: Involvement of PRDM16. J. Biol. Chem. 2011, 286, 43112–43122. [Google Scholar] [CrossRef]
- Wang, W.; Seale, P. Control of brown and beige fat development. Nat. Rev. Mol. Cell Biol. 2016, 17, 691–702. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Kim, J.; Park, H.J.; Kim, H. Anti-Obesity Effects of a Prunus persica and Nelumbo nucifera Mixture in Mice Fed a High-Fat Diet. Nutrients 2020, 12, 3392. https://doi.org/10.3390/nu12113392
Song J, Kim J, Park HJ, Kim H. Anti-Obesity Effects of a Prunus persica and Nelumbo nucifera Mixture in Mice Fed a High-Fat Diet. Nutrients. 2020; 12(11):3392. https://doi.org/10.3390/nu12113392
Chicago/Turabian StyleSong, Jungbin, Jiye Kim, Hyo Jin Park, and Hocheol Kim. 2020. "Anti-Obesity Effects of a Prunus persica and Nelumbo nucifera Mixture in Mice Fed a High-Fat Diet" Nutrients 12, no. 11: 3392. https://doi.org/10.3390/nu12113392
APA StyleSong, J., Kim, J., Park, H. J., & Kim, H. (2020). Anti-Obesity Effects of a Prunus persica and Nelumbo nucifera Mixture in Mice Fed a High-Fat Diet. Nutrients, 12(11), 3392. https://doi.org/10.3390/nu12113392