ω-3 Polyunsaturated Fatty Acids on Colonic Inflammation and Colon Cancer: Roles of Lipid-Metabolizing Enzymes Involved


Abstract
1. Introduction
2. Effects of ω-3 PUFAs on CRC and IBD
2.1. Effects of ω-3 PUFAs on CRC
2.2. Effects of ω-3 PUFAs on IBD
2.3. Potential Reasons for the Mixed Results of ω-3 PUFAs
- Both CRC and IBD are highly heterogeneous diseases, and previous studies have shown that ω-3 PUFAs have varied effects on different types of diseases. The plasma level of ω-3 PUFAs was negatively associated with the risks of proximal colon cancer, but with not distal colon cancer or overall CRC risk [19]. The consumption of ω-3 PUFAs decreased the risks of developing rectal cancer but increased the risks of developing distal colon cancer in men [23]. The administration of fish oil reduced the aberrant crypt foci and adenoma incidence, but not the carcinoma incidence, in a DMH-induced CRC model in rats [28]. It is feasible that ω-3 PUFAs target some specific types of colon carcinogenesis or inflammation, which remains to be better defined.
- Interindividual genetic variations could also influence the effects of ω-3 PUFAs on CRC and IBD. Many human studies have demonstrated significant interindividual variations in response to ω-3 PUFAs [14,15,72,73,74,75,76], which has made it difficult to confirm the efficacy of ω-3 PUFAs. The continuation of the current ω-3 PUFA research paradigms that neglect interindividual variation can be expected to keep generating mixed results and to fail to clarify their effects [15,16]. Notably, recent research supports that ω-3 PUFA-metabolizing enzymes contribute to the biological actions of ω-3 PUFAs. A recent study showed that there is a high degree of interindividual variability in metabolizing ω-3 PUFAs to generate lipid metabolites [14]. In addition, many studies support the critical roles of ω-3 lipid metabolizing enzymes in the activities of ω-3 PUFAs. For example, Dwyer et al. [75] showed that a diet rich in ω-3 PUFAs decreased, while a diet rich in ω-6 PUAFs increased, the risks of atherosclerosis in the subpopulation carrying a specific 5-LOX genotype but not in the general population. Other studies have also supported that polymorphism in genes encoding lipid-metabolizing genes affect the effects of ω-3 PUFAs on CRC. Notably, in a population-based case-control study, lower DHA consumption is linked to increased CRC risk in individuals with polymorphic variants in the PTGS1 gene [74]. The ω-3 PUFAs consumption only increased disease-free survival rate in CRC patients with upregulation of the PTGS2 gene [21,77]. These results emphasize the need to better understand the roles of lipid metabolism in the actions of ω-3 PUFAs.
- Contamination and impurities in medication, supplements, and products can potentially compromise the protective effects of ω-3 PUFAs in clinical applications. ω-3 PUFAs are highly unstable and are easily oxidized. Oxidized ω-3 PUFAs release lipid peroxidation/oxidative products, which are cytotoxic and genotoxic to colonic cells [78,79]. Moreover, persistent organic pollutants (POPs) and foreign contaminations in fish oil supplements could exacerbate the colon carcinogenesis by stimulating aberrant crypt foci formation in rats [80]. The use of high-quality ω-3 PUFAs is critical in future human and animal studies to exclude the potential adverse effects from lipid oxidative products and contaminations. In addition, multiple studies have shown that the beneficial effects of ω-3 PUFAs, including anti-inflammation [81,82], anti-atherosclerosis [83], and anti-metastasis [84] effects, are dose-dependent. More studies are needed to determine the optimal dose and treatment time to maximize the beneficial effect of ω-3 PUFAs and to establish the official recommended daily intake for the general public and for CRC patients.
3. Roles of ω-3 PUFA Metabolism in Mediating Inflammation and Cancer
3.1. Enzymatic Metabolism of ω-3 PUFAs
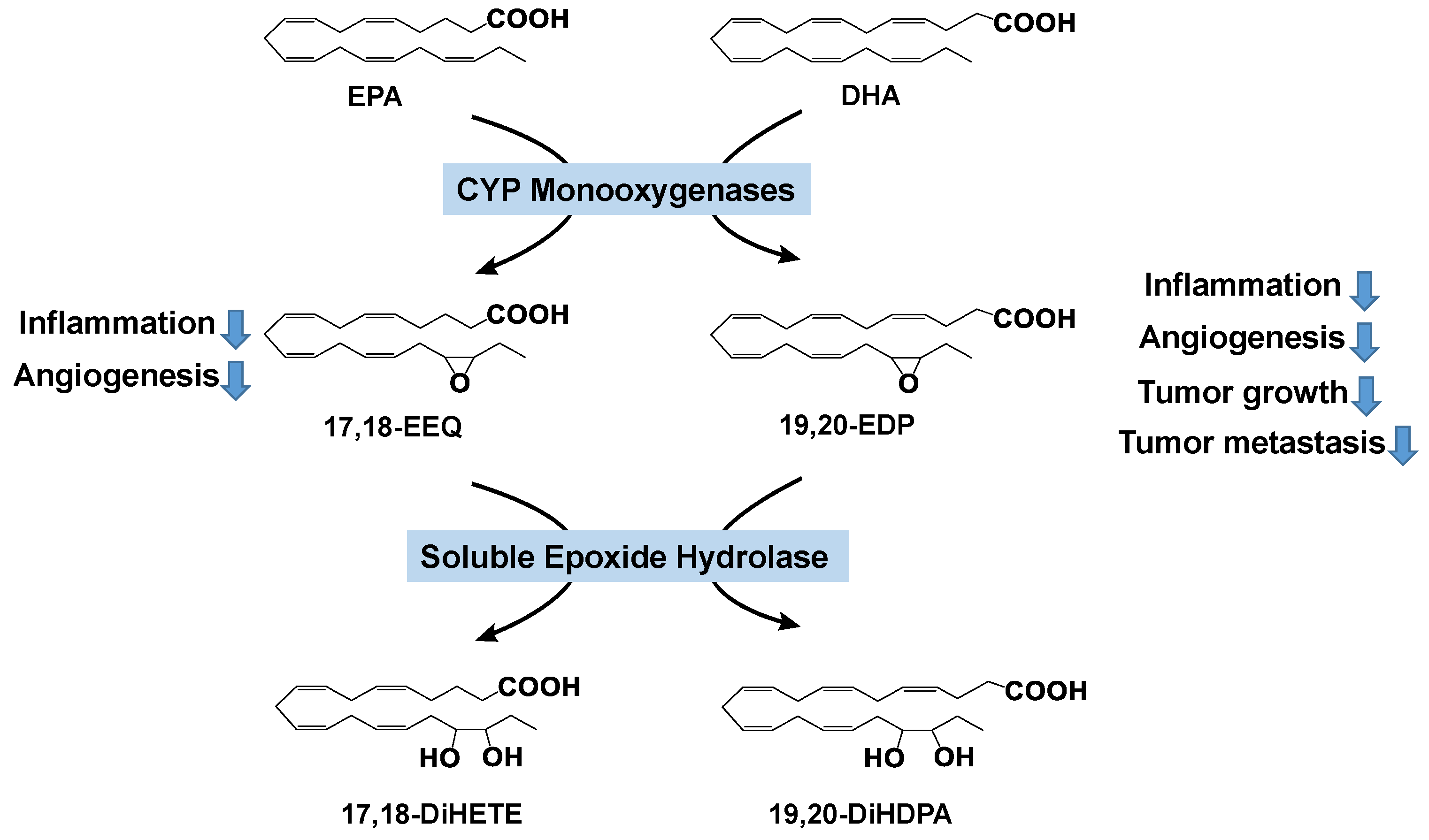
3.2. Effects of CYP-Produced ω-3 PUFA Metabolites on Inflammation and Cancer
3.3. Roles of CYP Pathway in the Pathogenesis of CRC
3.4. Therapeutic Benefit of sEH inhibitors with ω-3 PUFAs Combination in Inflammation and Cancer
4. Summary
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Key, T.J.; Allen, N.E.; Spencer, E.A.; Travis, R.C. The effect of diet on risk of cancer. Lancet (Lond. Engl.) 2002, 360, 861–868. [Google Scholar] [CrossRef]
- Sasazuki, S.; Inoue, M.; Iwasaki, M.; Sawada, N.; Shimazu, T.; Yamaji, T.; Takachi, R.; Tsugane, S. Intake of n-3 and n-6 polyunsaturated fatty acids and development of colorectal cancer by subsite: Japan Public Health Center-based prospective study. Int. J. Cancer 2011, 129, 1718–1729. [Google Scholar] [CrossRef]
- Murff, H.J.; Shrubsole, M.J.; Cai, Q.; Smalley, W.E.; Dai, Q.; Milne, G.L.; Ness, R.M.; Zheng, W. Dietary intake of PUFAs and colorectal polyp risk. Am. J. Clin. Nutr. 2012, 95, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Sandler, D.P.; Galanko, J.; Martin, C.; Sandler, R.S. Intake of polyunsaturated fatty acids and distal large bowel cancer risk in whites and African Americans. Am. J. Epidemiol. 2010, 171, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.N.; Chavarro, J.E.; Lee, I.M.; Willett, W.C.; Ma, J. A 22-year prospective study of fish, n-3 fatty acid intake, and colorectal cancer risk in men. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Schloss, I.; Kidd, M.S.; Tichelaar, H.Y.; Young, G.O.; O’Keefe, S.J. Dietary factors associated with a low risk of colon cancer in coloured west coast fishermen. S. Afr. Med. J. 1997, 87, 152–158. [Google Scholar] [PubMed]
- Pot, G.K.; Geelen, A.; van Heijningen, E.M.; Siezen, C.L.; van Kranen, H.J.; Kampman, E. Opposing associations of serum n-3 and n-6 polyunsaturated fatty acids with colorectal adenoma risk: An endoscopy-based case-control study. Int. J. Cancer 2008, 123, 1974–1977. [Google Scholar] [CrossRef] [PubMed]
- MacLean, C.H.; Newberry, S.J.; Mojica, W.A.; Khanna, P.; Issa, A.M.; Suttorp, M.J.; Lim, Y.W.; Traina, S.B.; Hilton, L.; Garland, R.; et al. Effects of omega-3 fatty acids on cancer risk: A systematic review. JAMA 2006, 295, 403–415. [Google Scholar] [CrossRef]
- Rose, D.P.; Connolly, J.M. Omega-3 fatty acids as cancer chemopreventive agents. Pharmacol. Ther. 1999, 83, 217–244. [Google Scholar]
- Serhan, C.N.; Petasis, N.A. Resolvins and protectins in inflammation resolution. Chem. Rev. 2011, 111, 5922–5943. [Google Scholar] [CrossRef] [PubMed]
- Sapieha, P.; Stahl, A.; Chen, J.; Seaward, M.R.; Willett, K.L.; Krah, N.M.; Dennison, R.J.; Connor, K.M.; Aderman, C.M.; Liclican, E.; et al. 5-Lipoxygenase metabolite 4-HDHA is a mediator of the antiangiogenic effect of omega-3 polyunsaturated fatty acids. Sci. Transl. Med. 2011, 3, 69ra12. [Google Scholar] [CrossRef] [PubMed]
- Bagga, D.; Wang, L.; Farias-Eisner, R.; Glaspy, J.A.; Reddy, S.T. Differential effects of prostaglandin derived from omega-6 and omega-3 polyunsaturated fatty acids on COX-2 expression and IL-6 secretion. Proc. Natl. Acad. Sci. USA 2003, 100, 1751–1756. [Google Scholar] [CrossRef] [PubMed]
- Nording, M.L.; Yang, J.; Georgi, K.; Hegedus Karbowski, C.; German, J.B.; Weiss, R.H.; Hogg, R.J.; Trygg, J.; Hammock, B.D.; Zivkovic, A.M. Individual variation in lipidomic profiles of healthy subjects in response to omega-3 Fatty acids. PLoS ONE 2013, 8, e76575. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Genetic variants in the metabolism of omega-6 and omega-3 fatty acids: Their role in the determination of nutritional requirements and chronic disease risk. Exp. Biol. Med. (Maywood) 2010, 235, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, S.H.; Waterland, R.A.; Ordovas, J.M.; Muoio, D.M.; Jia, W.; Fodor, A. Highlights of the 2012 Research Workshop: Using nutrigenomics and metabolomics in clinical nutrition research. J. Parenter. Enteral Nutr. 2013, 37, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Geelen, A.; Schouten, J.M.; Kamphuis, C.; Stam, B.E.; Burema, J.; Renkema, J.M.S.; Bakker, E.-J.; van’t Veer, P.; Kampman, E. Fish Consumption, n-3 Fatty Acids, and Colorectal Cancer: A Meta-Analysis of Prospective Cohort Studies. Am. J. Epidemiol. 2007, 166, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Kantor, E.D.; Lampe, J.W.; Peters, U.; Vaughan, T.L.; White, E. Long-chain omega-3 polyunsaturated fatty acid intake and risk of colorectal cancer. Nutr. Cancer 2014, 66, 716–727. [Google Scholar] [CrossRef]
- Aglago, E.K.; Huybrechts, I.; Murphy, N.; Casagrande, C.; Nicolas, G.; Pischon, T.; Fedirko, V.; Severi, G.; Boutron-Ruault, M.C.; Fournier, A.; et al. Consumption of Fish and Long-chain n-3 Polyunsaturated Fatty Acids Is Associated with Reduced Risk of Colorectal Cancer in a Large European Cohort. Clin. Gastroenterol. Hepatol. 2020, 18, 654–666 e656. [Google Scholar] [CrossRef]
- West, N.J.; Clark, S.K.; Phillips, R.K.; Hutchinson, J.M.; Leicester, R.J.; Belluzzi, A.; Hull, M.A. Eicosapentaenoic acid reduces rectal polyp number and size in familial adenomatous polyposis. Gut 2010, 59, 918–925. [Google Scholar] [CrossRef]
- Van Blarigan, E.L.; Fuchs, C.S.; Niedzwiecki, D.; Ye, X.; Zhang, S.; Song, M.; Saltz, L.B.; Mayer, R.J.; Mowat, R.B.; Whittom, R.; et al. Marine omega-3 Polyunsaturated Fatty Acid and Fish Intake after Colon Cancer Diagnosis and Survival: CALGB 89803 (Alliance). Cancer Epidemiol. Biomarkers Prev. 2018, 27, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Cockbain, A.J.; Volpato, M.; Race, A.D.; Munarini, A.; Fazio, C.; Belluzzi, A.; Loadman, P.M.; Toogood, G.J.; Hull, M.A. Anticolorectal cancer activity of the omega-3 polyunsaturated fatty acid eicosapentaenoic acid. Gut 2014, 63, 1760–1768. [Google Scholar] [CrossRef]
- Song, M.; Chan, A.T.; Fuchs, C.S.; Ogino, S.; Hu, F.B.; Mozaffarian, D.; Ma, J.; Willett, W.C.; Giovannucci, E.L.; Wu, K. Dietary intake of fish, omega-3 and omega-6 fatty acids and risk of colorectal cancer: A prospective study in U.S. men and women. Int. J. Cancer 2014, 135, 2413–2423. [Google Scholar] [CrossRef] [PubMed]
- Bakker, N.; van den Helder, R.S.; Stoutjesdijk, E.; van Pelt, J.; Houdijk, A.P.J. Effects of perioperative intravenous omega-3 fatty acids in colon cancer patients: A randomized, double-blind, placebo-controlled clinical trial. Am. J. Clin. Nutr. 2020, 111, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Notarnicola, M.; Tutino, V.; De Nunzio, V.; Dituri, F.; Caruso, M.G.; Giannelli, G. Dietary omega-3 Polyunsaturated Fatty Acids Inhibit Tumor Growth in Transgenic Apc(Min/+) Mice, Correlating with CB1 Receptor Up-Regulation. Int. J. Mol. Sci. 2017, 18, 485. [Google Scholar] [CrossRef]
- Fini, L.; Piazzi, G.; Ceccarelli, C.; Daoud, Y.; Belluzzi, A.; Munarini, A.; Graziani, G.; Fogliano, V.; Selgrad, M.; Garcia, M.; et al. Highly purified eicosapentaenoic acid as free fatty acids strongly suppresses polyps in Apc(Min/+) mice. Clin. Cancer Res. 2010, 16, 5703–5711. [Google Scholar] [CrossRef] [PubMed]
- Piazzi, G.; D’Argenio, G.; Prossomariti, A.; Lembo, V.; Mazzone, G.; Candela, M.; Biagi, E.; Brigidi, P.; Vitaglione, P.; Fogliano, V.; et al. Eicosapentaenoic acid free fatty acid prevents and suppresses colonic neoplasia in colitis-associated colorectal cancer acting on Notch signaling and gut microbiota. Int. J. Cancer 2014, 135, 2004–2013. [Google Scholar] [CrossRef]
- Moreira, A.P.; Sabarense, C.M.; Dias, C.M.; Lunz, W.; Natali, A.J.; Gloria, M.B.; Peluzio, M.C. Fish oil ingestion reduces the number of aberrant crypt foci and adenoma in 1,2-dimethylhydrazine-induced colon cancer in rats. Braz. J. Med. Biol. Res. 2009, 42, 1167–1172. [Google Scholar] [CrossRef]
- Reddy, B.S.; Patlolla, J.M.; Simi, B.; Wang, S.H.; Rao, C.V. Prevention of colon cancer by low doses of celecoxib, a cyclooxygenase inhibitor, administered in diet rich in omega-3 polyunsaturated fatty acids. Cancer Res. 2005, 65, 8022–8027. [Google Scholar] [CrossRef]
- Han, Y.M.; Park, J.M.; Cha, J.Y.; Jeong, M.; Go, E.J.; Hahm, K.B. Endogenous conversion of omega-6 to omega-3 polyunsaturated fatty acids in fat-1 mice attenuated intestinal polyposis by either inhibiting COX-2/beta-catenin signaling or activating 15-PGDH/IL-18. Int. J. Cancer 2016, 138, 2247–2256. [Google Scholar] [CrossRef]
- Han, Y.M.; Jeong, M.; Park, J.M.; Kim, M.Y.; Go, E.J.; Cha, J.Y.; Kim, K.J.; Hahm, K.B. The omega-3 polyunsaturated fatty acids prevented colitis-associated carcinogenesis through blocking dissociation of beta-catenin complex, inhibiting COX-2 through repressing NF-kappaB, and inducing 15-prostaglandin dehydrogenase. Oncotarget 2016, 7, 63583–63595. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.; Weylandt, K.H.; Habbel, P.; Wang, J.; Dignass, A.; Glickman, J.N.; Kang, J.X. Colitis-associated colon tumorigenesis is suppressed in transgenic mice rich in endogenous n-3 fatty acids. Carcinogenesis 2007, 28, 1991–1995. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, J.; Nimiya, Y.; Lee, K.S.S.; Sanidad, K.; Qi, W.; Sukamtoh, E.; Park, Y.; Liu, Z.; Zhang, G. omega-3 Polyunsaturated fatty acids and their cytochrome P450-derived metabolites suppress colorectal tumor development in mice. J. Nutr. Biochem. 2017, 48, 29–35. [Google Scholar] [CrossRef]
- Bathen, T.F.; Holmgren, K.; Lundemo, A.G.; Hjelstuen, M.H.; Krokan, H.E.; Gribbestad, I.S.; Schonberg, S.A. Omega-3 fatty acids suppress growth of SW620 human colon cancer xenografts in nude mice. Anticancer Res. 2008, 28, 3717–3723. [Google Scholar]
- Jeong, S.; Kim, D.Y.; Kang, S.H.; Yun, H.K.; Kim, J.L.; Kim, B.R.; Park, S.H.; Na, Y.J.; Jo, M.J.; Jeong, Y.A.; et al. Docosahexaenoic Acid Enhances Oxaliplatin-Induced Autophagic Cell Death via the ER Stress/Sesn2 Pathway in Colorectal Cancer. Cancers (Basel) 2019, 11, 982. [Google Scholar] [CrossRef]
- Fluckiger, A.; Dumont, A.; Derangere, V.; Rebe, C.; de Rosny, C.; Causse, S.; Thomas, C.; Apetoh, L.; Hichami, A.; Ghiringhelli, F.; et al. Inhibition of colon cancer growth by docosahexaenoic acid involves autocrine production of TNFalpha. Oncogene 2016, 35, 4611–4622. [Google Scholar] [CrossRef] [PubMed]
- Hawcroft, G.; Volpato, M.; Marston, G.; Ingram, N.; Perry, S.L.; Cockbain, A.J.; Race, A.D.; Munarini, A.; Belluzzi, A.; Loadman, P.M.; et al. The omega-3 polyunsaturated fatty acid eicosapentaenoic acid inhibits mouse MC-26 colorectal cancer cell liver metastasis via inhibition of PGE2-dependent cell motility. Br. J. Pharmacol. 2012, 166, 1724–1737. [Google Scholar] [CrossRef]
- Gutt, C.N.; Brinkmann, L.; Mehrabi, A.; Fonouni, H.; Muller-Stich, B.P.; Vetter, G.; Stein, J.M.; Schemmer, P.; Buchler, M.W. Dietary omega-3-polyunsaturated fatty acids prevent the development of metastases of colon carcinoma in rat liver. Eur. J. Nutr. 2007, 46, 279–285. [Google Scholar] [CrossRef]
- Williams, D.; Verghese, M.; Walker, L.T.; Boateng, J.; Shackelford, L.; Chawan, C.B. Flax seed oil and flax seed meal reduce the formation of aberrant crypt foci (ACF) in azoxymethane-induced colon cancer in Fisher 344 male rats. Food Chem. Toxicol. 2007, 45, 153–159. [Google Scholar] [CrossRef]
- Tsoukas, M.A.; Ko, B.J.; Witte, T.R.; Dincer, F.; Hardman, W.E.; Mantzoros, C.S. Dietary walnut suppression of colorectal cancer in mice: Mediation by miRNA patterns and fatty acid incorporation. J. Nutr. Biochem. 2015, 26, 776–783. [Google Scholar] [CrossRef]
- Watson, H.; Mitra, S.; Croden, F.C.; Taylor, M.; Wood, H.M.; Perry, S.L.; Spencer, J.A.; Quirke, P.; Toogood, G.J.; Lawton, C.L.; et al. A randomised trial of the effect of omega-3 polyunsaturated fatty acid supplements on the human intestinal microbiota. Gut 2018, 67, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Akedo, I.; Ishikawa, H.; Nakamura, T.; Kimura, K.; Takeyama, I.; Suzuki, T.; Kameyama, M.; Sato, S.; Nakamura, T.; Matsuzawa, Y.; et al. Three cases with familial adenomatous polyposis diagnosed as having malignant lesions in the course of a long-term trial using docosahexanoic acid (DHA)-concentrated fish oil capsules. Jpn J. Clin. Oncol. 1998, 28, 762–765. [Google Scholar] [CrossRef][Green Version]
- Kobayashi, M.; Tsubono, Y.; Otani, T.; Hanaoka, T.; Sobue, T.; Tsugane, S. Fish, long-chain n-3 polyunsaturated fatty acids, and risk of colorectal cancer in middle-aged Japanese: The JPHC study. Nutr. Cancer 2004, 49, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.C.; Butler, L.M.; Corral, R.; Joshi, A.D.; Yuan, J.M.; Koh, W.P.; Yu, M.C. Polyunsaturated fatty acids, DNA repair single nucleotide polymorphisms and colorectal cancer in the Singapore Chinese Health Study. J. Nutrigenet. Nutrigenomics 2009, 2, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Woodworth, H.L.; McCaskey, S.J.; Duriancik, D.M.; Clinthorne, J.F.; Langohr, I.M.; Gardner, E.M.; Fenton, J.I. Dietary fish oil alters T lymphocyte cell populations and exacerbates disease in a mouse model of inflammatory colitis. Cancer Res. 2010, 70, 7960–7969. [Google Scholar] [CrossRef]
- Lee, J.Y.; Sim, T.B.; Lee, J.E.; Na, H.K. Chemopreventive and Chemotherapeutic Effects of Fish Oil derived Omega-3 Polyunsaturated Fatty Acids on Colon Carcinogenesis. Clin. Nutr. Res. 2017, 6, 147–160. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, B.L.; Shang, B.; Chen, A.S.; Liu, S.Q.; Sun, W.; Yin, H.Z.; Yin, J.Q.; Su, Q. Nutrition support in surgical patients with colorectal cancer. World J. Gastroenterol. 2011, 17, 1779–1786. [Google Scholar] [CrossRef]
- Dahlhamer, J.M.; Zammitti, E.P.; Ward, B.W.; Wheaton, A.G.; Croft, J.B. Prevalence of Inflammatory Bowel Disease among Adults Aged > 18 Years—United States, 2015. Morb. Mortal. Wkly. Rep. 2016, 65, 1166–1169. [Google Scholar] [CrossRef]
- Nguyen, G.C.; Chong, C.A.; Chong, R.Y. National estimates of the burden of inflammatory bowel disease among racial and ethnic groups in the United States. J. Crohns Colitis 2014, 8, 288–295. [Google Scholar] [CrossRef]
- Cohen, B.L.; Sachar, D.B. Update on anti-tumor necrosis factor agents and other new drugs for inflammatory bowel disease. BMJ (Clin. Res. Ed.) 2017, 357, j2505. [Google Scholar] [CrossRef]
- Almallah, Y.Z.; Richardson, S.; O’Hanrahan, T.; Mowat, N.A.; Brunt, P.W.; Sinclair, T.S.; Ewen, S.; Heys, S.D.; Eremin, O. Distal procto-colitis, natural cytotoxicity, and essential fatty acids. Am. J. Gastroenterol. 1998, 93, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Aslan, A.; Triadafilopoulos, G. Fish oil fatty acid supplementation in active ulcerative colitis: A double-blind, placebo-controlled, crossover study. Am. J. Gastroenterol. 1992, 87, 432–437. [Google Scholar] [PubMed]
- McCall, T.B.; O’Leary, D.; Bloomfield, J.; O’Morain, C.A. Therapeutic potential of fish oil in the treatment of ulcerative colitis. Aliment. Pharmacol. Ther. 1989, 3, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Whiting, C.V.; Bland, P.W.; Tarlton, J.F. Dietary n-3 polyunsaturated fatty acids reduce disease and colonic proinflammatory cytokines in a mouse model of colitis. Inflamm. Bowel Dis. 2005, 11, 340–349. [Google Scholar] [CrossRef]
- Kunisawa, J.; Arita, M.; Hayasaka, T.; Harada, T.; Iwamoto, R.; Nagasawa, R.; Shikata, S.; Nagatake, T.; Suzuki, H.; Hashimoto, E.; et al. Dietary omega3 fatty acid exerts anti-allergic effect through the conversion to 17,18-epoxyeicosatetraenoic acid in the gut. Sci. Rep. 2015, 5, 9750. [Google Scholar] [CrossRef]
- Sharma, M.; Kaur, R.; Kaushik, K.; Kaushal, N. Redox modulatory protective effects of omega-3 fatty acids rich fish oil against experimental colitis. Toxicol. Mech. Methods 2019, 29, 244–254. [Google Scholar] [CrossRef]
- Varnalidis, I.; Ioannidis, O.; Karamanavi, E.; Ampas, Z.; Poutahidis, T.; Taitzoglou, I.; Paraskevas, G.; Botsios, D. Omega 3 fatty acids supplementation has an ameliorative effect in experimental ulcerative colitis despite increased colonic neutrophil infiltration. Rev. Esp. Enferm. Dig. 2011, 103, 511–518. [Google Scholar] [CrossRef]
- Morin, C.; Blier, P.U.; Fortin, S. MAG-EPA reduces severity of DSS-induced colitis in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G808–G821. [Google Scholar] [CrossRef]
- Brahmbhatt, V.; Oliveira, M.; Briand, M.; Perrisseau, G.; Bastic Schmid, V.; Destaillats, F.; Pace-Asciak, C.; Benyacoub, J.; Bosco, N. Protective effects of dietary EPA and DHA on ischemia-reperfusion-induced intestinal stress. J. Nutr. Biochem. 2013, 24, 104–111. [Google Scholar] [CrossRef]
- Hudert, C.A.; Weylandt, K.H.; Lu, Y.; Wang, J.; Hong, S.; Dignass, A.; Serhan, C.N.; Kang, J.X. Transgenic mice rich in endogenous omega-3 fatty acids are protected from colitis. Proc. Natl. Acad. Sci. USA 2006, 103, 11276–11281. [Google Scholar] [CrossRef]
- Yum, H.W.; Kang, J.X.; Hahm, K.B.; Surh, Y.J. Constitutive omega-3 fatty acid production in fat-1 transgenic mice and docosahexaenoic acid administration to wild type mice protect against 2,4,6-trinitrobenzene sulfonic acid-induced colitis. Biochem. Biophys. Res. Commun. 2017, 487, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, H.; Hokari, R.; Kurihara, C.; Okada, Y.; Takebayashi, K.; Okudaira, K.; Watanabe, C.; Komoto, S.; Nakamura, M.; Tsuzuki, Y.; et al. Omega-3 polyunsaturated fatty acids ameliorate the severity of ileitis in the senescence accelerated mice (SAM)P1/Yit mice model. Clin. Exp. Immunol. 2009, 158, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.; Steinhart, A.H.; Griffiths, A.M. Omega 3 fatty acids (fish oil) for maintenance of remission in ulcerative colitis. Cochrane Database Syst. Rev. 2007. [Google Scholar] [CrossRef]
- Barbosa, D.S.; Cecchini, R.; El Kadri, M.Z.; Rodriguez, M.A.; Burini, R.C.; Dichi, I. Decreased oxidative stress in patients with ulcerative colitis supplemented with fish oil omega-3 fatty acids. Nutrition 2003, 19, 837–842. [Google Scholar] [CrossRef]
- Dichi, I.; Frenhane, P.; Dichi, J.B.; Correa, C.R.; Angeleli, A.Y.; Bicudo, M.H.; Rodrigues, M.A.; Victoria, C.R.; Burini, R.C. Comparison of omega-3 fatty acids and sulfasalazine in ulcerative colitis. Nutrition 2000, 16, 87–90. [Google Scholar] [CrossRef]
- Bukki, J.; Stanga, Z.; Tellez, F.B.; Duclos, K.; Kolev, M.; Krahenmann, P.; Pabst, T.; Iff, S.; Juni, P. Omega-3 poly-unsaturated fatty acids for the prevention of severe neutropenic enterocolitis in patients with acute myeloid leukemia. Nutr. Cancer 2013, 65, 834–842. [Google Scholar] [CrossRef]
- Labonte, M.E.; Couture, P.; Tremblay, A.J.; Hogue, J.C.; Lemelin, V.; Lamarche, B. Eicosapentaenoic and docosahexaenoic acid supplementation and inflammatory gene expression in the duodenum of obese patients with type 2 diabetes. Nutr. J. 2013, 12, 98. [Google Scholar] [CrossRef]
- Vieira de Barros, K.; Gomes de Abreu, G.; Xavier, R.A.; Real Martinez, C.A.; Ribeiro, M.L.; Gambero, A.; de Oliveira Carvalho, P.; Silveira, V.L. Effects of a high fat or a balanced omega 3/omega 6 diet on cytokines levels and DNA damage in experimental colitis. Nutrition 2011, 27, 221–226. [Google Scholar] [CrossRef]
- Shoda, R.; Matsueda, K.; Yamato, S.; Umeda, N. Therapeutic efficacy of N-3 polyunsaturated fatty acid in experimental Crohn’s disease. J. Gastroenterol. 1995, 30 (Suppl. 8), 98–101. [Google Scholar]
- Matsunaga, H.; Hokari, R.; Kurihara, C.; Okada, Y.; Takebayashi, K.; Okudaira, K.; Watanabe, C.; Komoto, S.; Nakamura, M.; Tsuzuki, Y.; et al. Omega-3 fatty acids exacerbate DSS-induced colitis through decreased adiponectin in colonic subepithelial myofibroblasts. Inflamm. Bowel Dis. 2008, 14, 1348–1357. [Google Scholar] [CrossRef]
- Torres, D.M.; Tooley, K.L.; Butler, R.N.; Smith, C.L.; Geier, M.S.; Howarth, G.S. Lyprinol only partially improves indicators of small intestinal integrity in a rat model of 5-fluorouracil-induced mucositis. Cancer Biol. Ther. 2008, 7, 295–302. [Google Scholar] [CrossRef]
- Fradet, V.; Cheng, I.; Casey, G.; Witte, J.S. Dietary omega-3 fatty acids, cyclooxygenase-2 genetic variation, and aggressive prostate cancer risk. Clin. Cancer Res. 2009, 15, 2559–2566. [Google Scholar] [CrossRef]
- Wang, J.; John, E.; Ingles, S. 5-lipoxygenase and 5-lipoxygenase-activating protein gene polymorphisms, dietary linoleic acid, and risk for breast cancer. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2748–2754. [Google Scholar] [CrossRef]
- Habermann, N.; Ulrich, C.M.; Lundgreen, A.; Makar, K.W.; Poole, E.M.; Caan, B.; Kulmacz, R.; Whitton, J.; Galbraith, R.; Potter, J.D.; et al. PTGS1, PTGS2, ALOX5, ALOX12, ALOX15, and FLAP SNPs: Interaction with fatty acids in colon cancer and rectal cancer. Genes Nutr. 2013, 8, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Dwyer, J.H.; Allayee, H.; Dwyer, K.M.; Fan, J.; Wu, H.; Mar, R.; Lusis, A.J.; Mehrabian, M. Arachidonate 5-lipoxygenase promoter genotype, dietary arachidonic acid, and atherosclerosis. N. Engl. J. Med. 2004, 350, 29–37. [Google Scholar] [CrossRef]
- Stephensen, C.B.; Armstrong, P.; Newman, J.W.; Pedersen, T.L.; Legault, J.; Schuster, G.U.; Kelley, D.; Vikman, S.; Hartiala, J.; Nassir, R.; et al. ALOX5 gene variants affect eicosanoid production and response to fish oil supplementation. J. Lipid Res. 2011, 52, 991–1003. [Google Scholar] [CrossRef]
- Song, M.; Ou, F.S.; Zemla, T.J.; Hull, M.A.; Shi, Q.; Limburg, P.J.; Alberts, S.R.; Sinicrope, F.A.; Giovannucci, E.L.; Van Blarigan, E.L.; et al. Marine omega-3 fatty acid intake and survival of stage III colon cancer according to tumor molecular markers in NCCTG Phase III trial N0147 (Alliance). Int. J. Cancer 2019, 145, 380–389. [Google Scholar] [CrossRef]
- Wang, W.; Yang, H.; Johnson, D.; Gensler, C.; Decker, E.; Zhang, G. Chemistry and biology of ω-3 PUFA peroxidation-derived compounds. Prostaglandins Other Lipid Mediat. 2017, 132, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Gueraud, F.; Tache, S.; Steghens, J.P.; Milkovic, L.; Borovic-Sunjic, S.; Zarkovic, N.; Gaultier, E.; Naud, N.; Helies-Toussaint, C.; Pierre, F.; et al. Dietary polyunsaturated fatty acids and heme iron induce oxidative stress biomarkers and a cancer promoting environment in the colon of rats. Free Radic. Biol. Med. 2015, 83, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.Y.; Hoh, E.; Kang, B.; DeHamer, R.; Kim, J.Y.; Lumibao, J. Fish Oil Contaminated with Persistent Organic Pollutants Induces Colonic Aberrant Crypt Foci Formation and Reduces Antioxidant Enzyme Gene Expression in Rats. J. Nutr. 2017, 147, 1524–1530. [Google Scholar] [CrossRef]
- Ferguson, J.F.; Mulvey, C.K.; Patel, P.N.; Shah, R.Y.; Doveikis, J.; Zhang, W.; Tabita-Martinez, J.; Terembula, K.; Eiden, M.; Koulman, A.; et al. Omega-3 PUFA supplementation and the response to evoked endotoxemia in healthy volunteers. Mol. Nutr. Food Res. 2014, 58, 601–613. [Google Scholar] [CrossRef]
- Gupta, S.; Knight, A.G.; Gupta, S.; Keller, J.N.; Bruce-Keller, A.J. Saturated long-chain fatty acids activate inflammatory signaling in astrocytes. J. Neurochem. 2012, 120, 1060–1071. [Google Scholar] [CrossRef]
- Gladine, C.; Newman, J.W.; Durand, T.; Pedersen, T.L.; Galano, J.M.; Demougeot, C.; Berdeaux, O.; Pujos-Guillot, E.; Mazur, A.; Comte, B. Lipid profiling following intake of the omega 3 fatty acid DHA identifies the peroxidized metabolites F4-neuroprostanes as the best predictors of atherosclerosis prevention. PLoS ONE 2014, 9, e89393. [Google Scholar] [CrossRef]
- Rose, D.P.; Connolly, J.M.; Coleman, M. Effect of omega-3 fatty acids on the progression of metastases after the surgical excision of human breast cancer cell solid tumors growing in nude mice. Clin. Cancer Res. 1996, 2, 1751–1756. [Google Scholar]
- Szymczak, M.; Murray, M.; Petrovic, N. Modulation of angiogenesis by omega-3 polyunsaturated fatty acids is mediated by cyclooxygenases. Blood 2008, 111, 3514–3521. [Google Scholar] [CrossRef] [PubMed]
- Weylandt, K.H.; Chiu, C.-Y.; Gomolka, B.; Waechter, S.F.; Wiedenmann, B. Omega-3 fatty acids and their lipid mediators: Towards an understanding of resolvin and protectin formation. Prostaglandins Other Lipid Mediat. 2012, 97, 73–82. [Google Scholar] [CrossRef]
- Yang, P.; Jiang, Y.; Fischer, S.M. Prostaglandin E3 metabolism and cancer. Cancer Lett. 2014. [Google Scholar] [CrossRef] [PubMed]
- Hegde, S.; Kaushal, N.; Ravindra, K.C.; Chiaro, C.; Hafer, K.T.; Gandhi, U.H.; Thompson, J.T.; van den Heuvel, J.P.; Kennett, M.J.; Hankey, P.; et al. Δ12-prostaglandin J3, an omega-3 fatty acid–derived metabolite, selectively ablates leukemia stem cells in mice. Blood 2011, 118, 6909–6919. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Groeger, A.L.; Cipollina, C.; Cole, M.P.; Woodcock, S.R.; Bonacci, G.; Rudolph, T.K.; Rudolph, V.; Freeman, B.A.; Schopfer, F.J. Cyclooxygenase-2 generates anti-inflammatory mediators from omega-3 fatty acids. Nat. Chem. Biol. 2010, 6, 433–441. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, J.T.; Hu, Y.; Wooten, R.E.; Horita, D.A.; Samuel, M.P.; Thomas, M.J.; Sun, H.; Edwards, I.J. 15-lipoxygenase metabolites of docosahexaenoic acid inhibit prostate cancer cell proliferation and survival. PLoS ONE 2012, 7, e45480. [Google Scholar] [CrossRef]
- Bento, A.F.; Claudino, R.F.; Dutra, R.C.; Marcon, R.; Calixto, J.B. Omega-3 fatty acid-derived mediators 17(R)-hydroxy docosahexaenoic acid, aspirin-triggered resolvin D1 and resolvin D2 prevent experimental colitis in mice. J. Immunol. 2011, 187, 1957–1969. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Gomolka, B.; Dierkes, C.; Huang, N.R.; Schroeder, M.; Purschke, M.; Manstein, D.; Dangi, B.; Weylandt, K.H. Omega-6 docosapentaenoic acid-derived resolvins and 17-hydroxydocosahexaenoic acid modulate macrophage function and alleviate experimental colitis. Inflamm. Res. 2012, 61, 967–976. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, J.; Lyu, F.; Panigrahy, D.; Ferrara, K.W.; Hammock, B.; Zhang, G. omega-3 polyunsaturated fatty acids-derived lipid metabolites on angiogenesis, inflammation and cancer. Prostaglandins Other Lipid Mediat. 2014, 113–115, 13–20. [Google Scholar] [CrossRef]
- Arnold, C.; Markovic, M.; Blossey, K.; Wallukat, G.; Fischer, R.; Dechend, R.; Konkel, A.; von Schacky, C.; Luft, F.C.; Muller, D.N.; et al. Arachidonic acid-metabolizing cytochrome P450 enzymes are targets of {omega}-3 fatty acids. J. Biol. Chem. 2010, 285, 32720–32733. [Google Scholar] [CrossRef] [PubMed]
- Fromel, T.; Jungblut, B.; Hu, J.; Trouvain, C.; Barbosa-Sicard, E.; Popp, R.; Liebner, S.; Dimmeler, S.; Hammock, B.D.; Fleming, I. Soluble epoxide hydrolase regulates hematopoietic progenitor cell function via generation of fatty acid diols. Proc. Natl. Acad. Sci. USA 2012, 109, 9995–10000. [Google Scholar] [CrossRef]
- Fischer, R.; Konkel, A.; Mehling, H.; Blossey, K.; Gapelyuk, A.; Wessel, N.; von Schacky, C.; Dechend, R.; Muller, D.N.; Rothe, M.; et al. Dietary Omega-3 Fatty Acids Modulate the Eicosanoid Profile in Man Primarily via the CYP-epoxygenase Pathway. J. Lipid Res. 2014. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, A.; Yang, J.; Georgi, K.; Hegedus, C.; Nording, M.; O’Sullivan, A.; German, J.B.; Hogg, R.; Weiss, R.; Bay, C.; et al. Serum oxylipin profiles in IgA nephropathy patients reflect kidney functional alterations. Metabolomics 2012, 8, 1102–1113. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Keenan, A.H.; Pedersen, T.L.; Fillaus, K.; Larson, M.K.; Shearer, G.C.; Newman, J.W. Basal omega-3 fatty acid status affects fatty acid and oxylipin responses to high-dose n3-HUFA in healthy volunteers. J. Lipid Res. 2012, 53, 1662–1669. [Google Scholar] [CrossRef]
- Shearer, G.C.; Harris, W.S.; Pedersen, T.L.; Newman, J.W. Detection of omega-3 oxylipins in human plasma and response to treatment with omega-3 acid ethyl esters. J. Lipid Res. 2010, 51, 2074–2081. [Google Scholar] [CrossRef] [PubMed]
- Jump, D.B. The biochemistry of n-3 polyunsaturated fatty acids. J. Biol. Chem. 2002, 277, 8755–8758. [Google Scholar] [CrossRef]
- Malkowski, M.G.; Thuresson, E.D.; Lakkides, K.M.; Rieke, C.J.; Micielli, R.; Smith, W.L.; Garavito, R.M. Structure of eicosapentaenoic and linoleic acids in the cyclooxygenase site of prostaglandin endoperoxide H synthase-1. J. Biol. Chem. 2001, 276, 37547–37555. [Google Scholar] [CrossRef] [PubMed]
- Laneuville, O.; Breuer, D.K.; Xu, N.; Huang, Z.H.; Gage, D.A.; Watson, J.T.; Lagarde, M.; DeWitt, D.L.; Smith, W.L. Fatty Acid Substrate Specificities of Human Prostaglandin-endoperoxide H Synthase-1 and −2: FORMATION OF 12-HYDROXY-(9Z,13E/Z,15Z)-OCTADECATRIENOIC ACIDS FROM Α-LINOLENIC ACID. J. Biol. Chem. 1995, 270, 19330–19336. [Google Scholar] [PubMed]
- Zhang, G.; Panigrahy, D.; Mahakian, L.M.; Yang, J.; Liu, J.Y.; Stephen Lee, K.S.; Wettersten, H.I.; Ulu, A.; Hu, X.; Tam, S.; et al. Epoxy metabolites of docosahexaenoic acid (DHA) inhibit angiogenesis, tumor growth, and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 6530–6535. [Google Scholar] [CrossRef] [PubMed]
- Yanai, R.; Mulki, L.; Hasegawa, E.; Takeuchi, K.; Sweigard, H.; Suzuki, J.; Gaissert, P.; Vavvas, D.G.; Sonoda, K.H.; Rothe, M.; et al. Cytochrome P450-generated metabolites derived from omega-3 fatty acids attenuate neovascularization. Proc. Natl. Acad. Sci. USA 2014, 111, 9603–9608. [Google Scholar] [CrossRef]
- Hasegawa, E.; Inafuku, S.; Mulki, L.; Okunuki, Y.; Yanai, R.; Smith, K.E.; Kim, C.B.; Klokman, G.; Bielenberg, D.R.; Puli, N.; et al. Cytochrome P450 monooxygenase lipid metabolites are significant second messengers in the resolution of choroidal neovascularization. Proc. Natl. Acad. Sci. USA 2017, 114, E7545–E7553. [Google Scholar] [CrossRef]
- Nagatake, T.; Shiogama, Y.; Inoue, A.; Kikuta, J.; Honda, T.; Tiwari, P.; Kishi, T.; Yanagisawa, A.; Isobe, Y.; Matsumoto, N.; et al. The 17,18-epoxyeicosatetraenoic acid-G protein-coupled receptor 40 axis ameliorates contact hypersensitivity by inhibiting neutrophil mobility in mice and cynomolgus macaques. J. Allergy Clin. Immunol. 2018, 142, 470–484 e412. [Google Scholar] [CrossRef]
- Wang, C.; Liu, W.; Yao, L.; Zhang, X.; Ye, C.; Jiang, H.; He, J.; Zhu, Y.; Ai, D. Hydroxyeicosapentaenoic acids and epoxyeicosatetraenoic acids attenuate early occurrence of nonalcoholic fatty liver disease. Br. J. Pharmacol. 2017, 174, 2358–2372. [Google Scholar] [CrossRef]
- Morin, C.; Sirois, M.; Echave, V.; Albadine, R.; Rousseau, E. 17,18-epoxyeicosatetraenoic acid targets PPARgamma and p38 mitogen-activated protein kinase to mediate its anti-inflammatory effects in the lung: Role of soluble epoxide hydrolase. Am. J. Respir. Cell Mol. Biol. 2010, 43, 564–575. [Google Scholar] [CrossRef]
- Khaddaj-Mallat, R.; Rousseau, E. MAG-EPA and 17,18-EpETE target cytoplasmic signalling pathways to reduce short-term airway hyperresponsiveness. Pflugers Arch. 2015, 467, 1591–1605. [Google Scholar] [CrossRef]
- Kubota, T.; Arita, M.; Isobe, Y.; Iwamoto, R.; Goto, T.; Yoshioka, T.; Urabe, D.; Inoue, M.; Arai, H. Eicosapentaenoic acid is converted via omega-3 epoxygenation to the anti-inflammatory metabolite 12-hydroxy-17,18-epoxyeicosatetraenoic acid. FASEB J. 2014, 28, 586–593. [Google Scholar] [CrossRef]
- Mochimaru, T.; Fukunaga, K.; Miyata, J.; Matsusaka, M.; Masaki, K.; Kabata, H.; Ueda, S.; Suzuki, Y.; Goto, T.; Urabe, D.; et al. 12-OH-17,18-Epoxyeicosatetraenoic acid alleviates eosinophilic airway inflammation in murine lungs. Allergy 2018, 73, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, J.; Edin, M.L.; Wang, Y.; Luo, Y.; Wan, D.; Yang, H.; Song, C.-Q.; Xue, W.; Sanidad, K.Z. Targeted metabolomics identifies the cytochrome P450 monooxygenase eicosanoid pathway as a novel therapeutic target of colon tumorigenesis. Cancer Res. 2019, 79, 1822–1830. [Google Scholar] [PubMed]
- Panigrahy, D.; Edin, M.L.; Lee, C.R.; Huang, S.; Bielenberg, D.R.; Butterfield, C.E.; Barnes, C.M.; Mammoto, A.; Mammoto, T.; Luria, A.; et al. Epoxyeicosanoids stimulate multiorgan metastasis and tumor dormancy escape in mice. J. Clin. Investig. 2012, 122, 178–191. [Google Scholar] [CrossRef]
- Enayetallah, A.E.; French, R.A.; Grant, D.F. Distribution of soluble epoxide hydrolase, cytochrome P450 2C8, 2C9 and 2J2 in human malignant neoplasms. J. Mol. Histol. 2006, 37, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Kodani, S.; Hammock, B.D. Stabilized epoxygenated fatty acids regulate inflammation, pain, angiogenesis and cancer. Prog. Lipid Res. 2014, 53, 108–123. [Google Scholar] [CrossRef]
- Zhang, W.; Li, H.; Dong, H.; Liao, J.; Hammock, B.D.; Yang, G.Y. Soluble epoxide hydrolase deficiency inhibits dextran sulfate sodium-induced colitis and carcinogenesis in mice. Anticancer Res. 2013, 33, 5261–5271. [Google Scholar] [PubMed]
- Wang, W.; Yang, J.; Zhang, J.; Wang, Y.; Hwang, S.H.; Qi, W.; Wan, D.; Kim, D.; Sun, J.; Sanidad, K.Z. Lipidomic profiling reveals soluble epoxide hydrolase as a therapeutic target of obesity-induced colonic inflammation. Proc. Natl. Acad. Sci. USA 2018, 115, 5283–5288. [Google Scholar]
- Wang, Y.; Yang, J.; Wang, W.; Sanidad, K.Z.; Cinelli, M.A.; Wan, D.; Hwang, S.H.; Kim, D.; Lee, K.S.S.; Xiao, H.; et al. Soluble epoxide hydrolase is an endogenous regulator of obesity-induced intestinal barrier dysfunction and bacterial translocation. Proc. Natl. Acad. Sci. USA 2020, 117, 8431–8436. [Google Scholar] [CrossRef]
- Bastan, I.; Ge, X.N.; Dileepan, M.; Greenberg, Y.G.; Guedes, A.G.; Hwang, S.H.; Hammock, B.D.; Washabau, R.J.; Rao, S.P.; Sriramarao, P. Inhibition of soluble epoxide hydrolase attenuates eosinophil recruitment and food allergen-induced gastrointestinal inflammation. J. Leukoc. Biol. 2018, 104, 109–122. [Google Scholar] [CrossRef]
- Reisdorf, W.C.; Xie, Q.; Zeng, X.; Xie, W.; Rajpal, N.; Hoang, B.; Burgert, M.E.; Kumar, V.; Hurle, M.R.; Rajpal, D.K.; et al. Preclinical evaluation of EPHX2 inhibition as a novel treatment for inflammatory bowel disease. PLoS ONE 2019, 14, e0215033. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, A.L.; Liao, J.; Li, H.; Dong, H.; Chung, Y.T.; Bai, H.; Matkowskyj, K.A.; Hammock, B.D.; Yang, G.Y. Soluble epoxide hydrolase gene deficiency or inhibition attenuates chronic active inflammatory bowel disease in IL-10(-/-) mice. Dig. Dis. Sci. 2012, 57, 2580–2591. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liao, J.; Li, H.; Dong, H.; Bai, H.; Yang, A.; Hammock, B.D.; Yang, G.Y. Reduction of inflammatory bowel disease-induced tumor development in IL-10 knockout mice with soluble epoxide hydrolase gene deficiency. Mol. Carcinog. 2013, 52, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vicario, C.; Alcaraz-Quiles, J.; Garcia-Alonso, V.; Rius, B.; Hwang, S.H.; Titos, E.; Lopategi, A.; Hammock, B.D.; Arroyo, V.; Claria, J. Inhibition of soluble epoxide hydrolase modulates inflammation and autophagy in obese adipose tissue and liver: Role for omega-3 epoxides. Proc. Natl. Acad. Sci. USA 2015, 112, 536–541. [Google Scholar] [CrossRef]
- Liu, J.Y.; Park, S.H.; Morisseau, C.; Hwang, S.H.; Hammock, B.D.; Weiss, R.H. Sorafenib has soluble epoxide hydrolase inhibitory activity, which contributes to its effect profile in vivo. Mol. Cancer Ther. 2009, 8, 2193–2203. [Google Scholar] [CrossRef] [PubMed]
- Leineweber, C.G.; Pietzner, A.; Zhang, I.W.; Blessin, U.B.; Rothe, M.; Schott, E.; Schebb, N.H.; Weylandt, K.H. Assessment of the Effect of Sorafenib on Omega-6 and Omega-3 Epoxyeicosanoid Formation in Patients with Hepatocellular Carcinoma. Int. J. Mol. Sci. 2020, 21, 1875. [Google Scholar] [CrossRef]
- Kim, J.; Ulu, A.; Wan, D.; Yang, J.; Hammock, B.D.; Weiss, R.H. Addition of DHA Synergistically Enhances the Efficacy of Regorafenib for Kidney Cancer Therapy. Mol. Cancer Ther. 2016, 15, 890–898. [Google Scholar] [CrossRef]
- Lazaar, A.L.; Yang, L.; Boardley, R.L.; Goyal, N.S.; Robertson, J.; Baldwin, S.J.; Newby, D.E.; Wilkinson, I.B.; Tal-Singer, R.; Mayer, R.J.; et al. Pharmacokinetics, pharmacodynamics and adverse event profile of GSK2256294, a novel soluble epoxide hydrolase inhibitor. Br. J. Clin. Pharmacol. 2016, 81, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Whitcomb, R.; MacIntyre, E.; Tran, V.; Do, Z.N.; Sabry, J.; Patel, D.V.; Anandan, S.K.; Gless, R.; Webb, H.K. Pharmacokinetics and Pharmacodynamics of AR9281, an Inhibitor of Soluble Epoxide Hydrolase, in Single- and Multiple-Dose Studies in Healthy Human Subjects. J. Clin. Pharmacol. 2012, 52, 319–328. [Google Scholar] [CrossRef]
- Serhan, C.N. Novel lipid mediators and resolution mechanisms in acute inflammation: To resolve or not? Am. J. Pathol. 2010, 177, 1576–1591. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.N.; Bondy, S.C. Oxidative and Inflammatory Events in Prion Diseases: Can They Be Therapeutic Targets? Curr. Aging Sci. 2019, 11, 216–225. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Study | Individuals | N | ω-3 PUFA Treatment | Dose | Duration | Control Treatment | Results | Reference |
|---|---|---|---|---|---|---|---|---|
| VITAL prospective cohort | US adults | 68,109 | Fish oil supplements | N/A | 4+days/week for 3+years | no use | ↓ CRC risk | Kantor et al., 2014 [18] |
| EPIC prospective cohort | European adults | 521,324 | Highest ω-3 PUFAs intake | >470 mg/day | Median 14.9 years | lowest ω-3 PUFAs intake | ↓ CRC risk | Aglago et al., 2020 [19] |
| Randomized, double-blind, placebo-controlled trial | FAP patients | EPA-FFA (n = 28) | EPA-FFA | 2 g/day | 6 months | Placebo (n = 27) | ↓ polyp diameters | West et al., 2010 [20] |
| CALGB adjuvant chemotherapy trial | stage III colon cancer patients | 1011 | Highest marine ω-3 PUFAs intake | 0.33-0.57 g/day | >8 years | lowest marine ω-3 PUFAs intake | ↑ disease-free survival | Blarigan et al., 2018 [21] |
| Double-blind, randomised, placebo-controlled trial | CRCLM patients | EPA-FFA (n = 43) | EPA-FFA | 2 g/day | 12–65 days | Placebo (n = 45) | ↑ overall survival; no effect in disease burden and early CRC recurrence rates | Cockbain et al., 2014 [22] |
| HPFS and NHS cohort | US adults | 123,529 | Highest marine ω-3 PUFAs intake | ≥ 0.30 g/d (women) ≥ 0.41 g/d (men) | 24–26 years | lowest marine ω-3 PUFAs intake | No effect on overall CRC risk; ↑ distal colon cancer risk in men and women; ↓ rectal cancer risk in men | Song et al., 2014 [23] |
| Randomized, double-blind, placebo-controlled clinical trial | colon cancer patients | ω-3 PUFA (n = 21) | ω-3 PUFA intravenous infusion | 0.2 g/ kg/day | night before and morning after resection surgery | Saline infusions (n = 23) | ↑ infectious complications | Bakker et al., 2020 [24] |
| Model | Species | ω-3 PUFA Treatment | Dose | Duration | Control Treatment | Results | Reference |
|---|---|---|---|---|---|---|---|
| ApcMin/+ mouse | C57BL/6 mouse | Fish oil | 12% in diet | 10 weeks | Standard diet with soybean oil | ↓ intestinal polyp growth | Notarnicola et al., 2017 [25] |
| ApcMin/+ mouse | C57BL/6 mouse | EPA-FFA | 2.5% or 5% in diet | 12 weeks | AIN-93G diet with soybean oil | ↓ polyp number and load in both small intestine and colon. | Fini et al., 2010 [26] |
| ApcMin/+ mouse | C57BL/6 mouse | Endogenous ω-3 PUFA synthesis by transgene of fat-1 | 20 weeks | ApcMin/+ mice on standard diet with safflower oil | ↓ intestinal polyposis | Han et al., 2016 [30] | |
| AOM/DSS-induced CRC model | C57BL/6 mouse | Endogenous ω-3 PUFA synthesis by transgene of fat-1 | 16 weeks | Wild-type mice on standard diet | ↓ Tumor number | Han et al., 2016 [31] | |
| AOM/DSS-induced CRC model | C57BL/6 mouse | Endogenous ω-3 PUFA synthesis by transgene of fat-1 | 11 weeks | Wild-type mice on AIN-93G diet with safflower oil | ↓ incidence and growth rate | Nowak et al., 2007 [32] | |
| AOM/DSS-induced CRC model | C57BL/6 mouse | EPA-FFA | 1% in diet | 15 weeks | AIN-93G diet with corn oil | ↓ tumor multiplicity, incidence and maximum tumor size | Piazzi et al., 2014 [27] |
| DMH-induced CRC model | Wistar rat | Fish oil | 18% in diet | 36 weeks | AIN-93G diet with soybean oil | ↓ number of aberrant crypt foci; ↓ incidence of adenoma | Moreira et al., 2009 [28] |
| AOM-induced CRC model | F344 rat | Fish oil | 10% in diet | 26 weeks | AIN-93G diet with mixed lipids | ↓ colon tumor incidence and multiplicity | Reddy et al., 2005 [29] |
| MC38 cell-based xenograft model | C57BL/6 mouse | DHASCO Algae oil | 8% in diet | 5 weeks | AIN-93G diet with corn oil | ↓ tumor volume and weight | Wang et al., 2016 [33] |
| SW620 cell-based xenograft model | Nude mouse | Fish oil | 12% by calorie | 6 weeks | Standard diet | ↓ tumor growth and less aggressive | Bathen et al., 2008 [34] |
| HCT116 cell-based xenograft model | Nude mouse | DHA | 10 mg/kg | every other day for 13 days | Ethanol | ↓ tumor size | Jeong et al., 2019 [35] |
| HCT116 cell-based xenograft model | Nude mouse | DHA | 3% in diet | 14 days | Standard diet with sunflower oil | ↓ tumor growth | Fluckiger et al., 2016 [36] |
| H. hepaticus-induced CRC model | SMAD3 deficiency mouse | Fish oil | 6% in diet | 12 weeks | AIN-93G diet with corn oil | ↑ adenocarcinoma formation | Woodworth et al., 2010 [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, M.; Wang, W.; Zhang, G.; Hammock, B.D. ω-3 Polyunsaturated Fatty Acids on Colonic Inflammation and Colon Cancer: Roles of Lipid-Metabolizing Enzymes Involved. Nutrients 2020, 12, 3301. https://doi.org/10.3390/nu12113301
Tu M, Wang W, Zhang G, Hammock BD. ω-3 Polyunsaturated Fatty Acids on Colonic Inflammation and Colon Cancer: Roles of Lipid-Metabolizing Enzymes Involved. Nutrients. 2020; 12(11):3301. https://doi.org/10.3390/nu12113301
Chicago/Turabian StyleTu, Maolin, Weicang Wang, Guodong Zhang, and Bruce D. Hammock. 2020. "ω-3 Polyunsaturated Fatty Acids on Colonic Inflammation and Colon Cancer: Roles of Lipid-Metabolizing Enzymes Involved" Nutrients 12, no. 11: 3301. https://doi.org/10.3390/nu12113301
APA StyleTu, M., Wang, W., Zhang, G., & Hammock, B. D. (2020). ω-3 Polyunsaturated Fatty Acids on Colonic Inflammation and Colon Cancer: Roles of Lipid-Metabolizing Enzymes Involved. Nutrients, 12(11), 3301. https://doi.org/10.3390/nu12113301