Alterations in the Intestinal Morphology, Gut Microbiota, and Trace Mineral Status Following Intra-Amniotic Administration (Gallus gallus) of Teff (Eragrostis tef) Seed Extracts

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Extraction of Prebiotics From Teff
2.3. Iron, Zinc, Calcium and Magnesium Analysis
2.4. Phytate Analysis
2.5. Protein and Fiber Analysis
2.6. Polyphenol Extraction
Liquid Chromatography-Mass Spectroscopy (LC-MS) Analysis of Polyphenols
2.7. Animals and Study Design
2.8. Blood Analysis and Hb Measurements
2.9. Isolation of Total RNA From Duodenum and Liver Tissue Samples
2.10. Real-Time Polymerase Chain Reaction (RT-PCR)
2.10.1. Primer Design
2.10.2. Real-Time qPCR Design
2.11. Collection of Microbial Samples and Intestinal Contents DNA Isolation
16S rRNA Gene Amplification, Sequencing and Analysis
2.12. Glycogen Analysis
2.13. Liver Ferritin Analysis
2.14. Tissue Morphology Examination
2.15. Statistical Analysis
3. Results
3.1. Concentration of Calcium, Iron, Magnesium, and Zinc in Teff Flour and in Teff Flour Extract
3.2. Polyphenol Profile of the Teff Seed Flour and Extract
3.3. Hemoglobin, Body Weight, Cecum Weight, and Cecum:Body Weight Ratio
3.4. Hepatic, Serum Fe, and Zn Concentrations: LA:DGLA Ratio
3.5. Pectoral Muscle Glycogen Concentration
3.6. Liver Ferritin Concentration
3.7. Duodenal Gene Expression of Relevant Proteins
3.7.1. Fe-Related Proteins
3.7.2. Zn-Related Proteins
3.7.3. Ca-Related Proteins
3.7.4. Mg-Related Proteins
3.7.5. Inflammatory Cytokines and BBM Proteins
3.8. Morphometric Analysis of Duodenal Villi, Depth of Crypts and Goblet Cells
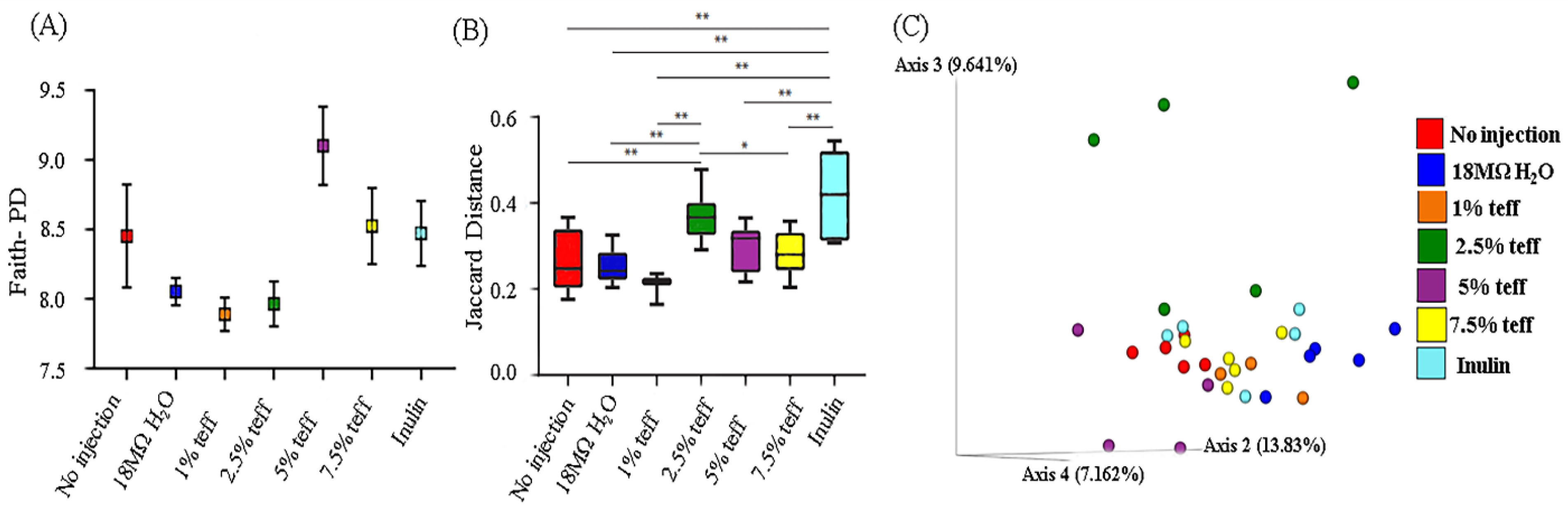
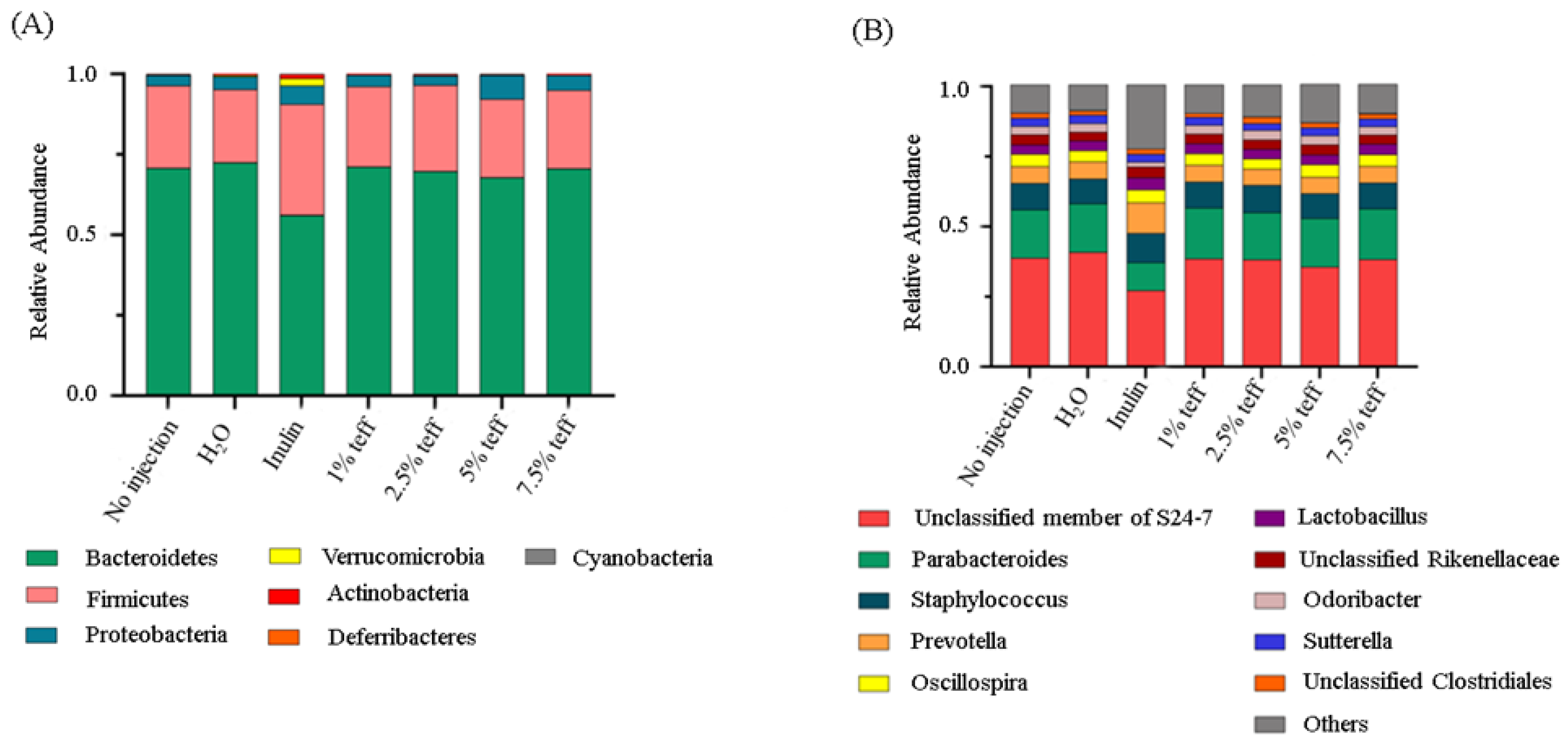
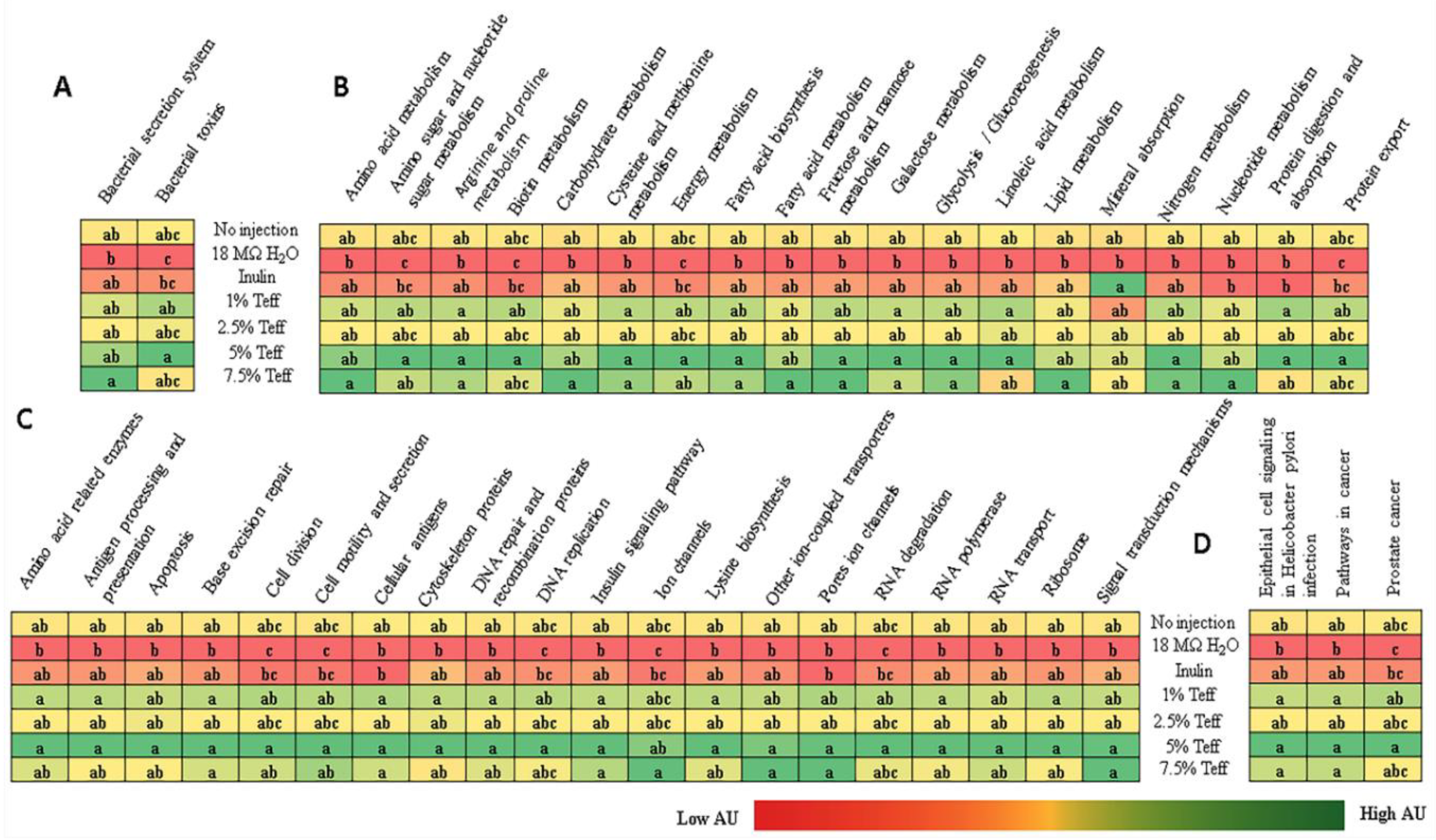
3.9. Analysis of the Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abebe, Z.; Takele, W.W.; Anlay, D.Z.; Ekubagewargies, D.T.; Getaneh, Z.; Abebe, M.; Melku, M. Prevalence of anemia and its associated factors among children in Ethiopia: A protocol for systematic review and meta-analysis. EJIFCC 2018, 29, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Gebreegziabher, T.; Stoecker, B.J. Iron deficiency was not the major cause of anemia in rural women of reproductive age in Sidama zone, southern Ethiopia: A cross-sectional study. PLoS ONE 2017, 12, e0184742. [Google Scholar] [CrossRef] [PubMed]
- World Bank. World Development Report 2019: The Changing Nature of Work.; License: Creative Commons Attribution CC BY 3.0 IGO; World Bank: Washington, DC, USA, 2019. [Google Scholar] [CrossRef]
- Satheesh, N.; Fanta, S.W. Review on structural, nutritional and anti-nutritional composition of Teff (Eragrostis tef) in comparison with Quinoa (Chenopodium quinoa Willd.). Cogent Food Agric. 2018, 4. [Google Scholar] [CrossRef]
- Umeta, M.; West, C.E.; Fufa, H. Content of zinc, iron, calcium and their absorption inhibitors in foods commonly consumed in Ethiopia. J. Food Compos. Anal. 2005, 18, 803–817. [Google Scholar] [CrossRef]
- Tako, E.; Glahn, R.P.; Knez, M.; Stangoulis, J.C. The effect of wheat prebiotics on the gut bacterial population and iron status of iron deficient broiler chickens. Nutr. J. 2014, 13, 58. [Google Scholar] [CrossRef]
- Baye, K. Teff: Nutrient Composition and Health Benefits; The International Food Policy Research Institute: Washington, DC, USA, 2014. [Google Scholar]
- Baye, K.; Mouquet-Rivier, C.; Icard-Vernière, C.; Picq, C.; Guyot, J.P. Changes in mineral absorption inhibitors consequent to fermentation of Ethiopian injera: Implications for predicted iron bioavailability and bioaccessibility. Int. J. Food Sci. Technol. 2014, 49, 174–180. [Google Scholar] [CrossRef]
- Harika, R.; Faber, M.; Samuel, F.; Mulugeta, A.; Kimiywe, J.; Eilander, A. Are low intakes and deficiencies in iron, vitamin A, zinc, and iodine of public health concern in Ethiopian, Kenyan, Nigerian, and South African children and adolescents? Food Nutr. Bull. 2017, 38, 405–427. [Google Scholar] [CrossRef]
- Preidis, G.A. Targeting the human microbiome with antibiotics, probiotics, and prebiotics: Gastroenterology enters the metagenomics era. Gastroenterology 2009, 136, 2015–2031. [Google Scholar] [CrossRef]
- Dias, D.M.; Kolba, N.; Hart, J.J.; Ma, M.; Sha, S.T.; Lakshmanan, N.; Nutti, M.R.; Martino, H.S.D.; Glahn, R.P.; Tako, E. Soluble extracts from carioca beans (Phaseolus vulgaris L.) affect the gut microbiota and iron related brush border membrane protein expression in vivo (Gallus gallus). Food Res. Int. 2019, 123, 172–180. [Google Scholar] [CrossRef]
- Hou, T.; Tako, E. The in ovo feeding administration (Gallus Gallus)—An emerging in vivo approach to assess bioactive compounds with potential nutritional benefits. Nutrients 2018, 10, 418. [Google Scholar] [CrossRef]
- Wang, X.; Kolba, N.; Liang, J.; Tako, E. Alterations in gut microflora populations and brush border functionality following intra-amniotic administration (Gallus gallus) of wheat bran prebiotic extracts. Food Funct. 2019, 10, 4834–4843. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, S.; Song, J.; Zhang, C.; Wang, Q.; Glahn, R.P.; Kolba, N.; Tako, E. Intra amniotic administration of raffinose and stachyose affects the intestinal brush border functionality and alters gut microflora populations. Nutrients 2017, 9, 304. [Google Scholar] [CrossRef] [PubMed]
- Hartono, K.; Reed, S.; Ankrah, N.A.; Glahn, R.P.; Tako, E. Alterations in gut microflora populations and brush border functionality following intra-amniotic daidzein administration. RSC Adv. 2014, 5, 6407–6412. [Google Scholar] [CrossRef]
- Yeung, C.K.; Glahn, R.E.; Welch, R.M.; Miller, D.D. Prebiotics and iron bioavailability—Is there a connection? J. Food Sci. 2005, 70, 88–92. [Google Scholar] [CrossRef]
- Rocchetti, G.; Lucini, L.; Giuberti, G.; Bhumireddy, S.R.; Mandal, R.; Trevisan, M.; Wishart, D.S. Transformation of polyphenols found in pigmented gluten-free flours during in vitro large intestinal fermentation. Food Chem. 2019, 298, 125068. [Google Scholar] [CrossRef]
- Gebremariam, M.M.; Zarnkow, M.; Becker, T. Teff (Eragrostis tef) as a raw material for malting, brewing and manufacturing of gluten-free foods and beverages: A review. J. Food Sci. Technol. 2014, 51, 2881–2895. [Google Scholar] [CrossRef]
- Reed, S.; Knez, M.; Uzan, A.; Stangoulis, J.C.R.; Glahn, R.P.; Koren, O.; Tako, E. Alterations in the gut (Gallus gallus) microbiota following the consumption of zinc biofortified wheat (Triticum aestivum)-based diet. J. Agric. Food Chem. 2018, 66, 6291–6299. [Google Scholar] [CrossRef]
- Tako, E.; Rutzke, M.A.; Glahn, R.P. Using the domestic chicken (Gallus gallus) as an in vivo model for iron bioavailability. Poultry Science 2010, 89, 514–521. [Google Scholar] [CrossRef]
- Reed, S.; Qin, X.; Ran-Ressler, R.; Brenna, J.T.; Glahn, R.P.; Tako, E. Dietary zinc deficiency affects blood linoleic acid: Dihomo-ihlinolenic acid (LA:DGLA) ratio; a sensitive physiological marker of zinc status in vivo (Gallus gallus). Nutrients 2014, 6, 1164. [Google Scholar] [CrossRef]
- Wiesinger, J.A.; Glahn, R.P.; Cichy, K.A.; Kolba, N.; Hart, J.J.; Tako, E. An in vivo (Gallus gallus) feeding trial demonstrating the enhanced iron bioavailability properties of the fast cooking manteca yellow bean (Phaseolus vulgaris L.). Nutrients 2019, 11, 1768. [Google Scholar] [CrossRef]
- Da Silva, B.P.; Kolba, N.; Stampini Duarte Martino, H.; Hart, J.; Tako, E. Soluble extracts from chia seed (Salvia hispanica L.) affect brush border membrane functionality, morphology and intestinal bacterial populations in vivo (Gallus gallus). Nutrients 2019, 11, 2457. [Google Scholar] [CrossRef] [PubMed]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Yosef, S.; Pandya-Lorch, R. Agriculture for Improved Nutrition: Seizing the Momentum; CABI: Boston, MA, USA, 2019. [Google Scholar]
- Petry, N.; Egli, I.; Zeder, C.; Walczyk, T.; Hurrell, R. Polyphenols and phytic acid contribute to the low iron bioavailability from common beans in young women. J. Nutr. 2010, 140, 1977–1982. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Egli, I.; Gahutu, J.B.; Tugirimana, P.L.; Boy, E.; Hurrell, R. Phytic acid concentration influences iron bioavailability from biofortified beans in Rwandese women with low iron status. J. Nutr. 2014, 144, 1681–1687. [Google Scholar] [CrossRef]
- Bouis, H.E.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Sec. 2017, 12, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Knez, M.; Stangoulis, J.C.R.; Glibetic, M.; Tako, E. The linoleic acid: Dihomo-γ-linolenic acid ratio (LA:DGLA)—An emerging biomarker of Zn status. Nutrients 2017, 9, 825. [Google Scholar] [CrossRef]
- Hou, T.; Kolba, N.; Glahn, R.P.; Tako, E. Intra-amniotic administration (Gallus gallus) of Cicer arietinum and Lens culinaris prebiotics extracts and duck egg white peptides affects calcium status and intestinal functionality. Nutrients 2017, 9, 785. [Google Scholar] [CrossRef]
- Smirnov, A.; Tako, E.; Ferket, P.R.; Uni, Z. Mucin gene expression and mucin content in the chicken intestinal goblet cells are affected by in ovo feeding of carbohydrates. Poult. Sci. 2006, 85, 669–673. [Google Scholar] [CrossRef]
- Sobolewska, A.; Elminowska-Wenda, G.; Bogucka, J.; Dankowiakowska, A.; Kułakowska, A.; Szczerba, A.; Stadnicka, K.; Szpinda, M.; Bednarczyk, M. The influence of in ovo injection with the prebiotic DiNovo® on the development of histomorphological parameters of the duodenum, body mass and productivity in large-scale poultry production conditions. J. Anim. Sci. Biotechnol. 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, P.; Wu, Y.; Guo, P.; Liu, L.; Ma, N.; Levesque, C.; Chen, Y.; Zhao, J.; Zhang, J.; et al. Dietary fiber increases butyrate-producing bacteria and improves the growth performance of weaned piglets. J. Agric. Food Chem. 2018, 66, 7995–8004. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, W.; Degroote, J.; Possemiers, S.; Chen, D.; De Smet, S.; Michiels, J. Arabinoxylan in wheat is more responsible than cellulose for promoting intestinal barrier function in weaned male piglets. J. Nutr. 2015, 145, 51–58. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Forward Primer (5'→3') | Reverse Primer (5'→3') | Base Pair | GI Identifier |
|---|---|---|---|---|
| Calcium Metabolism | ||||
| TRPV6 | GCTCCCAGAACCTTCTCTATTT | CCAGGTAATCCTGAGCTCTAATG | 123 | 418307 |
| PMCA1b | TGCAGATGCTGTGGGTAAAT | CCATAAGGCTTCCGCAATAGA | 100 | 374244 |
| NXC1 | CCTGACGGAGAAATAAGGAAGA | CCCAGGAGAAGACACAGATAAA | 114 | 395760 |
| Iron Metabolism | ||||
| DMT1 | TTGATTCAGAGCCTCCCATTAG | GCGAGGAGTAGGCTTGTATTT | 101 | 206597489 |
| Ferroportin | CTCAGCAATCACTGGCATCA | ACTGGGCAACTCCAGAAATAAG | 98 | 61098365 |
| DcytB | CATGTGCATTCTCTTCCAAAGTC | CTCCTTGGTGACCGCATTAT | 103 | 20380692 |
| Inflammatory Response | ||||
| IL-1β | CTCACAGTCCTTCGACATCTTC | TGTTGAGCCTCACTTTCTGG | 119 | 88702685 |
| IL-6 | ACCTCATCCTCCGAGACTTTA | GCACTGAAACTCCTGGTCTT | 105 | 302315692 |
| TNF-α | GACAGCCTATGCCAACAAGTA | TTACAGGAAGGGCAACTCATC | 109 | 53854909 |
| Magnesium Metabolism | ||||
| MRS2 | GCTGGTAACCGGGATTATGT | GCAGGAACATGAGGAGGTAAT | 105 | 420820 |
| TRPM6 | ACAGATGCTGCTGACTGATATG | AAGATAGTGGGTGGTAGGAGAA | 99 | 100859603 |
| TRPM7 | GCGTGGGATAGAGTTGACATT | TCACAAGGGCATCCAACATAG | 100 | 427502 |
| Zinc Metabolism | ||||
| ZnT1 | GGTAACAGAGCTGCCTTAACT | GGTAACAGAGCTGCCTTAACT | 105 | 54109718 |
| ZnT7 | GGAAGATGTCAGGATGGTTCA | CGAAGGACAAATTGAGGCAAAG | 87 | 56555152 |
| ZIP9 | CTAAGCAAGAGCAGCAAAGAAG | CATGAACTGTGGCAACGTAAAG | 100 | 237874618 |
| Δ6 desaturase | GGCGAAAGTCAGCCTATTGA | AGGTGGGAAGATGAGGAAGA | 93 | 261865208 |
| Hypertension | ||||
| ACE | CATGGCCTTGTCTGTCTCC | GAGGTATCCAAAGGGCAGG | 142 | 424059 |
| AT1R | TCATCTGGCTCCTTGCTGG | AACCTAGCCCAACCCTCAG | 138 | 396065 |
| BBM Functionality | ||||
| AP | CGTCAGCCAGTTTGACTATGTA | CTCTCAAAGAAGCTGAGGATGG | 138 | 45382360 |
| SI | CCAGCAATGCCAGCATATTG | CGGTTTCTCCTTACCACTTCTT | 95 | 2246388 |
| SGLT1 | GCATCCTTACTCTGTGGTACTG | TATCCGCACATCACACATCC | 106 | 8346783 |
| 18s rRNA | GCAAGACGAACTAAAGCGAAAG | TCGGAACTACGACGGTATCT | 100 | 7262899 |
| Treatment Group | Calcium (µg/g) | Iron (µg/g) | Magnesium (µg/g) | Zinc (µg/g) |
|---|---|---|---|---|
| Teff seed | 2259.32 ± 24.73 a | 60.64 ± 0.92 b | 2023.49 ± 18.93 a | 42.87 ± 0.42 b |
| Teff seed extract | 2052.07 ± 123.95 b | 128.23 ± 11.60 a | 1088.87 ± 42.24 b | 53.84 ± 4.50 a |
| Treatment Group | Insoluble Fiber (g/100 g) | Soluble Fiber (g/100 g) | Total Fiber (g/100 g) | Protein (g/100 g) | Phytic Acid (g/100 g) | Phytic Acid:Iron Ratio |
|---|---|---|---|---|---|---|
| Teff seed | 8.32 ± 0.23 a | 2.10 ± 0.12 b | 10.43 ± 0.11 b | 10.02± 0.39 a | 0.14± 0.02 a | 1.74 ± 0.03 a |
| Teff seed extract | 6.15 ± 0.61 b | 6.98 ± 0.91 a | 13.13 ± 0.30 a | 10.86± 0.88 a | 0.17± 0.11 a | 0.41 ± 0.05 b |
| Polyphenolic Compounds | Mass (Da) | (M + H) (Da) | (M − H) (Da) | Retention Time (min) | Found in (MS mode) | |
|---|---|---|---|---|---|---|
| Teff Seed (extract) | Teff Seed (flour) | |||||
| Protocatechuic Acid | 154.12 | 155.128 | 153.112 | 1.621 | NEG | NEG |
| Caffeic Acid | 180.16 | 181.168 | 179.152 | 2.958 | POS/NEG | ND |
| Vanillic Acid | 168.15 | 169.158 | 167.142 | 3.094 | POS | ND |
| p- Coumaric Acid | 164.16 | 165.168 | 164.152 | 3.895 | POS/NEG | ND |
| Ferulic Acid | 194.18 | 195.188 | 193.172 | 4.428 | POS/NEG | POS |
| Treatment Group | Hemoglobin (g/dL) | Body Weight Average (g) | Cecum Weight Average (g) | Cecum:Body Weight Ratio |
|---|---|---|---|---|
| NI | 8.747 ± 0.797 a | 45.4 ± 1.3 b | 0.5 ± 0.1 b | 0.010 ± 0.002 b |
| 18Ω H2O | 10.603 ± 0.591 a | 46.9 ± 1.2 a | 0.5 ± 0.1 a | 0.011 ± 0.001 a |
| 5% Inulin | 8.860 ± 0.690 a | 49.7 ± 0.9 a | 0.6 ± 0.1 a | 0.012 ± 0.001 a |
| 1% Teff | 9.575 ± 1.138 a | 47.0 ± 1.0 a | 0.6 ± 0.1 a | 0.012 ± 0.001 a |
| 2.5% Teff | 8.410 ± 0.920 a | 47.1 ± 1.1 a | 0.5 ± 0.0 b | 0.010 ± 0.001 b |
| 5% Teff | 8.501 ± 0.874 a | 46.5 ± 1.3 a | 0.7 ± 0.1 a | 0.015 ± 0.002 a |
| 7.5% Teff | 9.569 ± 0.633 a | 47.4 ± 1.4 a | 0.6 ± 0.0 a | 0.012 ± 0.001 a |
| Treatment Groups | Liver | Serum | ||
|---|---|---|---|---|
| Iron (µg/g) | Zinc (µg/g) | Iron (µg/g) | Zinc (µg/g) | |
| NI | 32.47 ± 2.83 b | 15.79 ± 0.95 c | 2.09 ± 0.24 a | 0.86 ± 0.08 a |
| 18Ω H2O | 37.93 ± 4.93 ab | 16.12 ± 0.96 c | 2.00 ± 0.27 a | 0.84 ± 0.07 a |
| 5% Inulin | 48.96 ± 4.39 a | 18.23 ± 0.88 abc | 2.76 ± 0.33 a | 0.97 ± 0.08 a |
| 1% Teff | 37.40 ± 3.67 ab | 17.61 ± 0.91 bc | 2.22 ± 0.40 a | 0.91 ± 0.08 a |
| 2.5% Teff | 36.39 ± 3.54 ab | 22.82 ± 2.42 ab | 2.09 ± 0.22 a | 0.89 ± 0.06 a |
| 5% Teff | 39.40 ± 3.89 ab | 23.37 ± 3.10 ab | 2.83 ± 0.28 a | 0.81 ± 0.07 a |
| 7.5% Teff | 46.44 ± 5.83 a | 24.14 ± 3.33 a | 2.88 ± 0.38 a | 1.02 ± 0.07 a |
| Treatment Group | Glycogen (mg/g) | Liver Ferritin (AU) |
|---|---|---|
| NI | 0.034 ± 0.011 a | 1.974 ± 0.005 a |
| 18Ω H2O | 0.022 ± 0.003 a | 1.534 ± 0.519 a |
| 5% Inulin | 0.029 ± 0.005 a | 0.148 ± 0.005 b |
| 1% Teff | 0.035 ± 0.006 a | 1.491 ± 0.299 a |
| 2.5% Teff | 0.022 ± 0.007 a | 0.275 ± 0.245 b |
| 5% Teff | 0.044 ± 0.017 a | 0.031± 0.001 b |
| 7.5% Teff | 0.021 ± 0.007 a | 0.034 ± 0.001 b |
| Treatment Group | Villus Length (µm) | Villus Diameter (µm) | Depth of Crypts (µm) |
|---|---|---|---|
| NI | 223.29 ± 3.44 ab | 53.15 ± 0.73 b | 66.30 ± 1.33 b |
| 18Ω H2O | 224.14 ± 4.10 ab | 46.84 ± 0.69 c | 53.42 ± 1.11 c |
| 5% Inulin | 274.11 ± 3.92 b | 43.66 ± 0.64 d | 51.30 ± 1.06 c |
| 1% Teff | 266.85 ± 4.39 a | 56.22 ± 0.78 a | 73.62 ± 1.40 a |
| 2.5% Teff | 263.66 ± 3.85 a | 53.44 ± 0.75 ab | 65.74 ± 1.26 b |
| 5% Teff | 302.83 ± 2.95 ab | 54.80 ± 0.67 ab | 67.75 ± 1.43 b |
| 7.5% Teff | 290.86 ± 3.12 a | 50.00 ± 0.02 ab | 66.38 ± 1.25 b |
| Treatment Group | Goblet Cell Diameter (µM) | Crypts Goblet Cell Number | Total Villus Goblet Cell Number | Villus Goblet Cell Number | ||
|---|---|---|---|---|---|---|
| Acidic | Neutral | Mixed | ||||
| NI | 6.50 ± 0.06 cd | 9.53 ± 0.32 b | 51.32 ± 1.34 b | 10.22 ± 0.26 a | 0.12 ± 0.03 a | 0.13 ± 0.03 c |
| 18Ω H2O | 6.37 ± 0.06 de | 10.01 ± 0.22 b | 44.83 ± 1.16 c | 9.46 ± 0.23 b | 0.00 ± 0.00 b | 0.05 ± 0.03 d |
| 5% Inulin | 6.06 ± 0.04 f | 12.27 ± 0.33 a | 54.56 ± 1.40 b | 10.65 ± 0.25 a | 0.00 ± 0.00 b | 0.02 ± 0.01 d |
| 1% Teff | 6.82 ± 0.05 b | 12.15 ± 0.28 a | 61.76 ± 1.56 a | 9.37 ± 0.24 b | 0.00 ± 0.00 b | 0.22 ± 0.03 b |
| 2.5% Teff | 7.54 ± 0.05 a | 11.81 ± 0.24 a | 54.27 ± 1.20 b | 6.21 ± 0.15 d | 0.00 ± 0.00 b | 0.32 ± 0.04 a |
| 5% Teff | 6.29 ± 0.06 e | 10.11 ± 0.27 b | 42.54 ± 0.93 c | 8.33 ± 0.20 c | 0.00 ± 0.00 b | 0.18 ± 0.03 bc |
| 7.5% Teff | 6.64 ± 0.06 bc | 9.51 ± 0.24 b | 43.12 ± 0.98 c | 7.81 ± 0.19 c | 0.00 ± 0.00 b | 0.16 ± 0.03 bc |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carboni, J.; Reed, S.; Kolba, N.; Eshel, A.; Koren, O.; Tako, E. Alterations in the Intestinal Morphology, Gut Microbiota, and Trace Mineral Status Following Intra-Amniotic Administration (Gallus gallus) of Teff (Eragrostis tef) Seed Extracts. Nutrients 2020, 12, 3020. https://doi.org/10.3390/nu12103020
Carboni J, Reed S, Kolba N, Eshel A, Koren O, Tako E. Alterations in the Intestinal Morphology, Gut Microbiota, and Trace Mineral Status Following Intra-Amniotic Administration (Gallus gallus) of Teff (Eragrostis tef) Seed Extracts. Nutrients. 2020; 12(10):3020. https://doi.org/10.3390/nu12103020
Chicago/Turabian StyleCarboni, Johnathon, Spenser Reed, Nikolai Kolba, Adi Eshel, Omry Koren, and Elad Tako. 2020. "Alterations in the Intestinal Morphology, Gut Microbiota, and Trace Mineral Status Following Intra-Amniotic Administration (Gallus gallus) of Teff (Eragrostis tef) Seed Extracts" Nutrients 12, no. 10: 3020. https://doi.org/10.3390/nu12103020
APA StyleCarboni, J., Reed, S., Kolba, N., Eshel, A., Koren, O., & Tako, E. (2020). Alterations in the Intestinal Morphology, Gut Microbiota, and Trace Mineral Status Following Intra-Amniotic Administration (Gallus gallus) of Teff (Eragrostis tef) Seed Extracts. Nutrients, 12(10), 3020. https://doi.org/10.3390/nu12103020