Role of Vitamin A in Mammary Gland Development and Lactation

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Vitamin A Dietary Sources, Availability, and Requirements
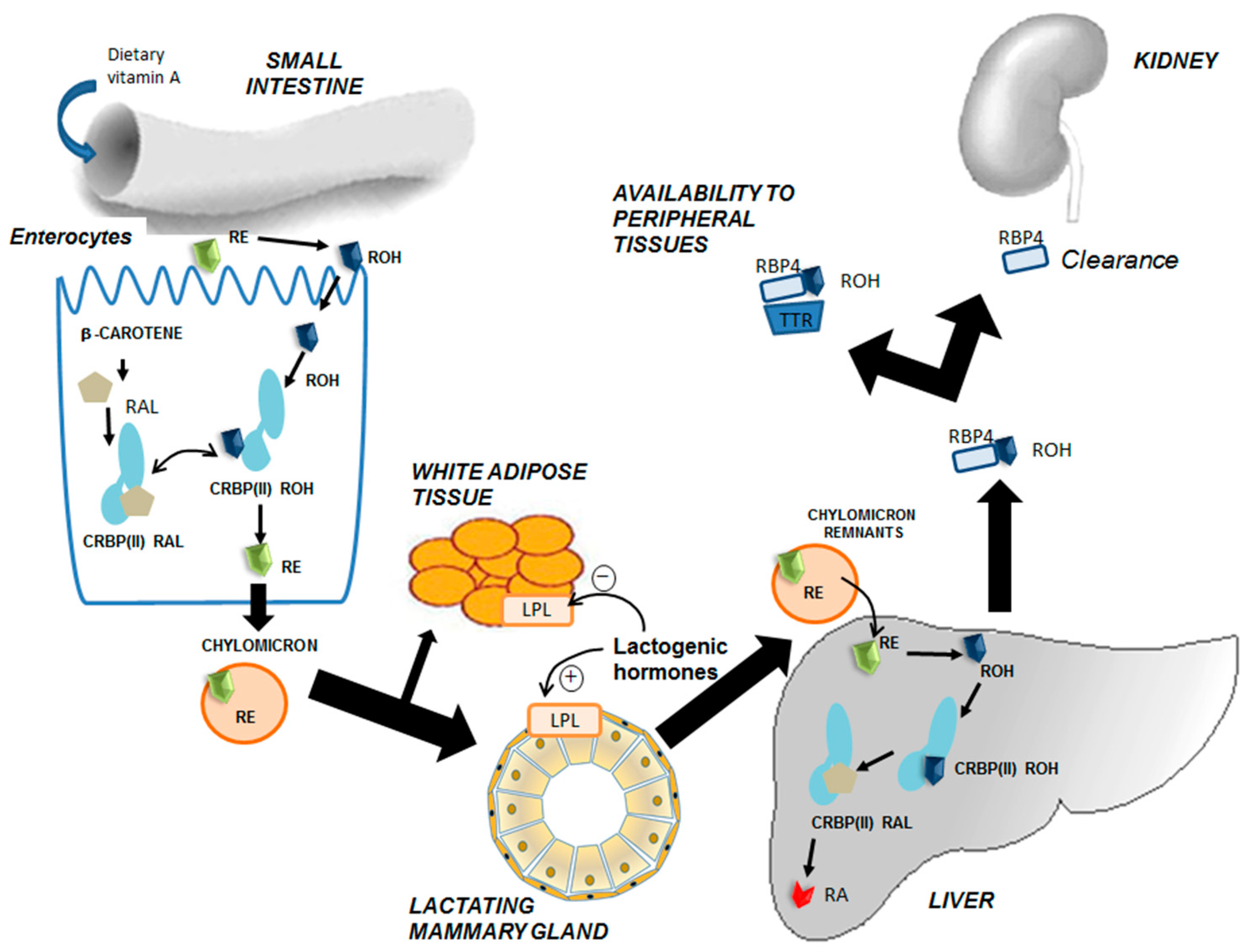
3. Intertissue Flux and Metabolic Transformations of Vitamin A
4. Role of Vitamin A during Development and Regression of the Mammary Gland
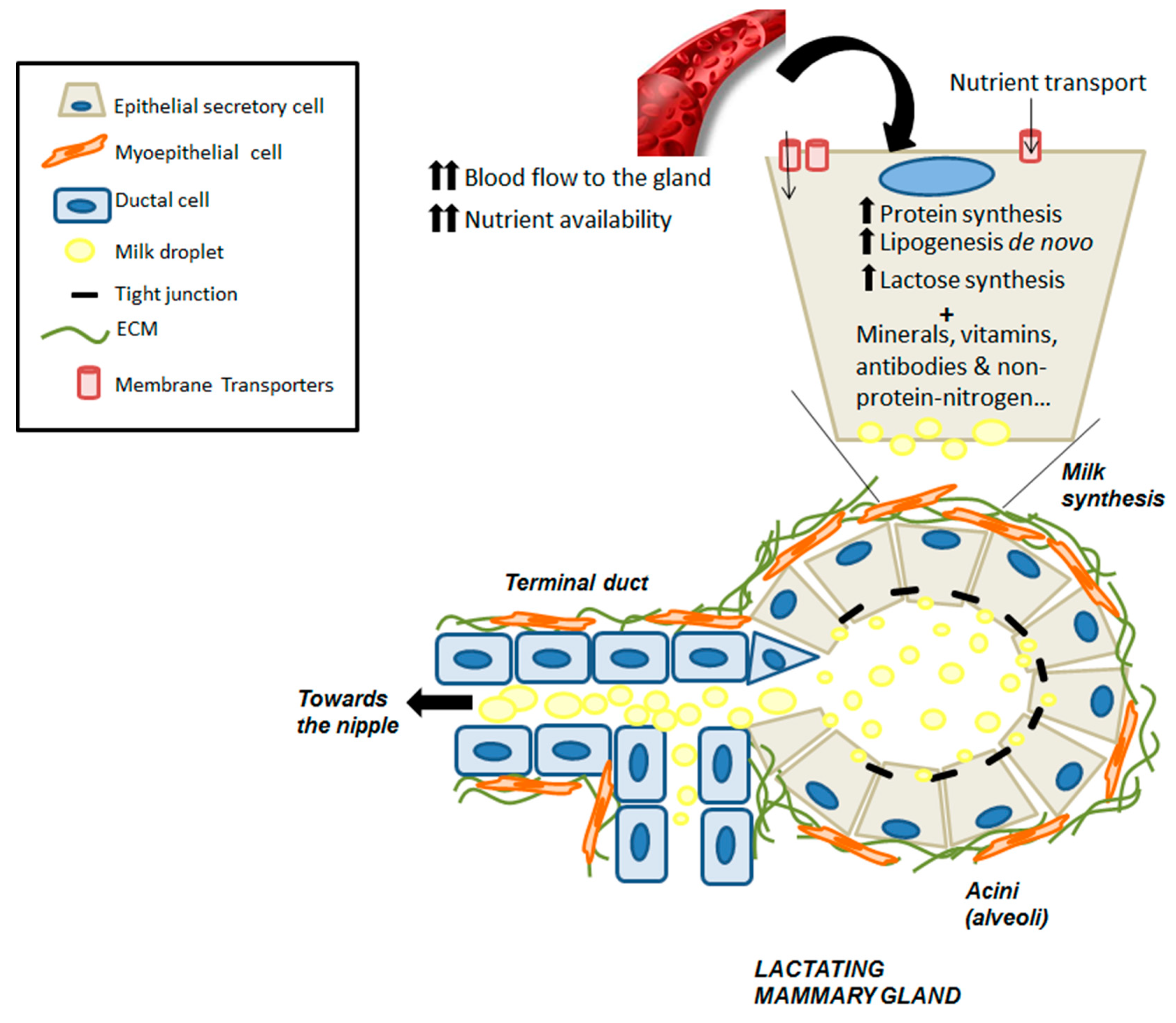
4.1. An Overview of Mammary Gland Development
4.2. Retinoids and Mammary Gland Development
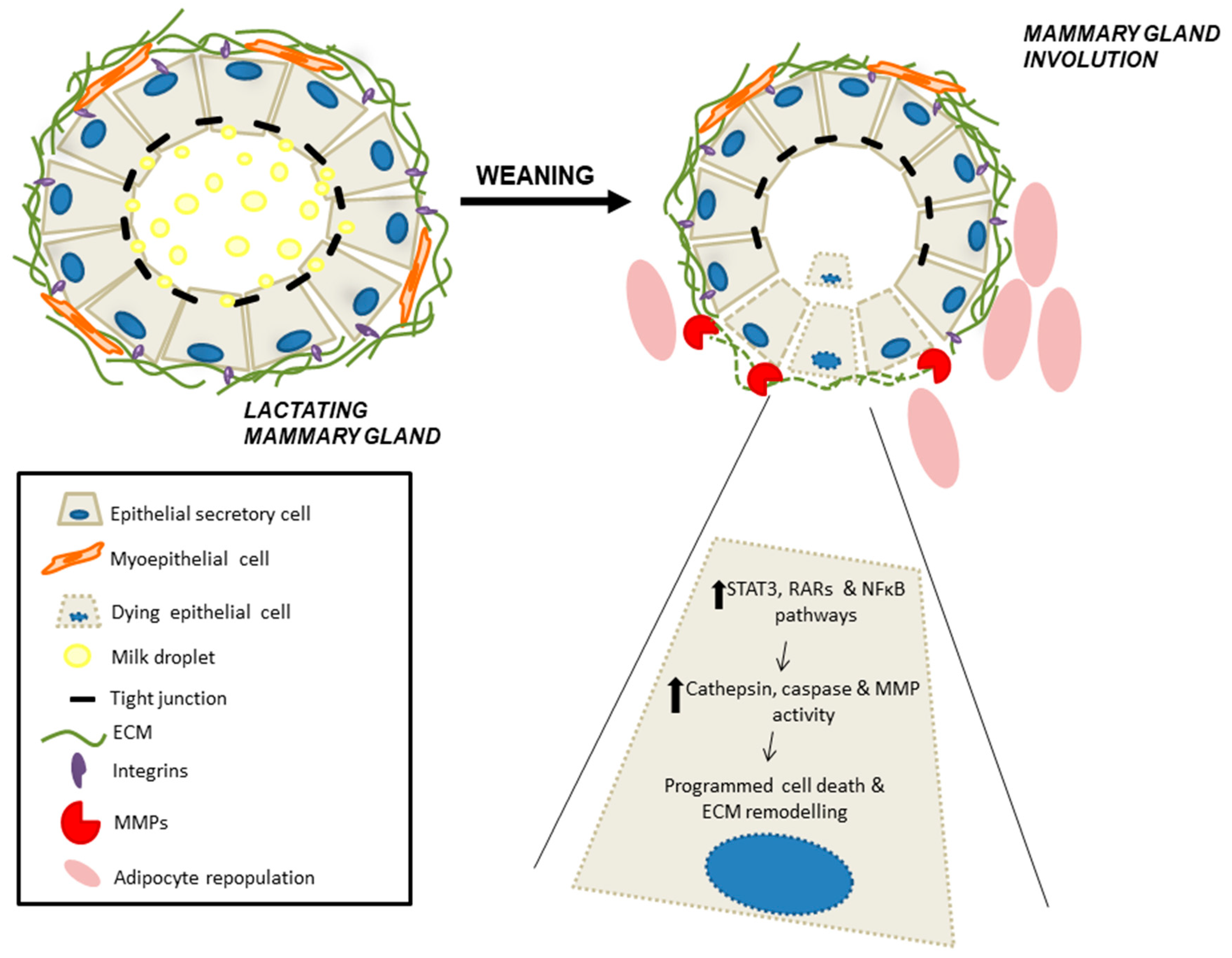
4.3. Retinoid Signaling During Mammary Gland Involution
5. Vitamin A and Carotenoid Uptake by the Mammary Gland during Lactation
6. Vitamin A Concentration in Milk
7. Deficiency of Vitamin A and Carotenoids during Pregnancy and Lactation
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McLaren, D.S.; Kraemer, K. Vitamin A in nature. World Rev. Nutr. Diet 2012, 103, 7–17. [Google Scholar] [CrossRef] [PubMed]
- De Luca, L.M. Retinoids and their receptors in differentiation, embryogenesis and neoplasia. FASEB J. 1991, 5, 2924–2933. [Google Scholar] [CrossRef] [PubMed]
- Livrea, M.A.; Tesoriere, L. Antioxidant activity of vitamin A within lipid environments. In Fat-Soluble Vitamins; Subcellular Biochemistry; Springer: Boston, MA, USA, 1998; Volume 30, pp. 113–143. [Google Scholar] [CrossRef]
- Barber, T.; Borrás, E.; Torres, L.; García, C.; Cabezuelo, F.; Lloret, A.; Pallardó, F.V.; Viña, J.R. Vitamin A deficiency causes oxidative damage to liver mitochondria in rats. Free Radic. Biol. Med. 2000, 29, 1–7. [Google Scholar] [CrossRef]
- Clagett-Dame, M.; Knutson, D. Vitamin A in reproduction and development. Nutrients 2011, 3, 385–428. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, A.; Zaragozá, R.; Vivó-Sesé, I.; Viña, J.R.; Miralles, V.J. Retinol, at concentration greater than the physiological limit, induces oxidative stress and apoptosis in human dermal fibroblasts. Exp. Dermatol. 2004, 13, 45–54. [Google Scholar] [CrossRef]
- Rhinn, M.; Dollé, P. Retinoic acid signaling during development. Development 2012, 139, 843–858. [Google Scholar] [CrossRef]
- Ross, A.C. Vitamin A and retinoic acid in T cell-related immunity. Am. J. Clin. Nutr. 2012, 96, 1166S–1172S. [Google Scholar] [CrossRef]
- Sommer, A.; Vyas, K.S. A global clinical view on vitamin A and carotenoids. Am. J. Clin. Nutr. 2012, 96, 1204S–1206S. [Google Scholar] [CrossRef]
- Esteban-Pretel, G.; Marín, M.P.; Renau-Piqueras, J.; Barber, T.; Timoneda, J. Vitamin A deficiency alters rat lung alveolar basement membrane: Reversibility by retinoic acid. J. Nutr. Biochem. 2010, 21, 227–236. [Google Scholar] [CrossRef]
- Zielinska, M.A.; Wesołowska, A.; Pawlus, B.; Hamułka, J. Health Effects of Carotenoids during Pregnancy and Lactation. Nutrients 2017, 9, 838. [Google Scholar] [CrossRef]
- Chambon, P. A decade of molecular biology of retinoic acid receptors. FASEB J. 1996, 10, 940–954. [Google Scholar] [CrossRef] [PubMed]
- Ziouzenkova, O.; Orasanu, G.; Sharlach, M.; Akiyama, T.E.; Berger, J.P.; Viereck, J.; Hamilton, J.A.; Tang, G.; Dolnokowski, G.G.; Vogel, S.; et al. Retinaldehyde represses adipogenesis and diet-induced obesity. Nat. Med. 2007, 13, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.C.; Noy, N. Signaling by Vitamin A and Retinol-Binding Protein in Regulation of Insulin Responses and Lipid Homeostasis. Biochim. Biophys. Acta 2012, 1821, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Al Tanoury, Z.; Piskunov, A.; Rochette-Egly, C. Vitamin A and Retinoid Signaling: Genomic and Nongenomic Effects. J. Lipid Res. 2013, 54, 1761–1775. [Google Scholar] [CrossRef] [PubMed]
- WHO, World Health Organization. Global prevalence of vitamin A deficiency in populations at risk 1995–2005. In WHO Global Database on Vitamin A Deficiency; World Health Organization: Geneva, Switzerland, 2009; ISBN 978-92-4-159801-9. [Google Scholar]
- Emmett, P.M.; Rogers, I.S. Properties of human milk and their relationship with maternal nutrition. Early Hum. Dev. 1997, 9, S7–S28. [Google Scholar] [CrossRef]
- Strobel, M.; Tinz, J.; Biesalski, H.K. The importance of β-carotene as a source of vitamin A with special regard to pregnant and breastfeeding women. Eur. J. Nutr. 2007, 46, 1–20. [Google Scholar] [CrossRef]
- Cruz, S.; Pereira da Cruz, S.; Ramalho, A. Impact of Vitamin A Supplementation on Pregnant Women and on Women Who Have Just Given Birth: A Systematic Review. J. Am. Coll. Nutr. 2018, 37, 243–250. [Google Scholar] [CrossRef]
- Borel, P.; Desmarchelier, C. Genetic Variations Associated with Vitamin A Status and Vitamin A Bioavailability. Nutrients 2017, 9, 246. [Google Scholar] [CrossRef]
- Das, B.C.; Thapa, P.; Karki, R.; Das, S.; Mahapatra, S.; Liu, T.C.; Torregroza, I.; Wallace, D.P.; Kambhampati, S.; Van Veldhuizen, P.; et al. Retinoic Acid Signaling Pathways in Development and Diseases. Bioorg. Med. Chem. 2014, 22, 673–683. [Google Scholar] [CrossRef]
- Harrison, E.H.; Curley, R.W., Jr. Carotenoids and Retinoids: Nomenclature, Chemistry, and Analysis. In The Biochemistry of Retinoid Signaling II; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2016; Volume 81, pp. 1–19. [Google Scholar] [CrossRef]
- Eroglu, A.; Harrison, E.H. Carotenoid metabolism in mammals, including man: Formation, occurrence, and function of apocarotenoids. J. Lipid Res. 2013, 54, 1719–1730. [Google Scholar] [CrossRef]
- Kim, Y.K.; Wassef, L.; Chung, S.; Jiang, H.; Wyss, A.; Blaner, W.S.; Quadro, L. β-Carotene and its Cleavage Enzyme β-Carotene-15,15′-oxygenase (CMOI) Affect Retinoid Metabolism in Developing Tissues. FASEB J. 2011, 25, 1641–1652. [Google Scholar] [CrossRef] [PubMed]
- Combs, G.F., Jr.; McClung, J.P. The Vitamins: Fundamental Aspects in Nutrition and Health; Academic Press, Elsevier: New York, NY, USA, 2017. [Google Scholar] [CrossRef]
- Wieringa, F.T.; Dijkhuizen, M.A.; Berger, J. Micronutrient deficiencies and their public health implications for South-East Asia. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 479–482. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Qin, J.; Dolnikowski, G.G.; Russell, R.M.; Grusak, M.A. Golden rice is an effective source of vitamin A. Am. J. Clin. Nutr. 2009, 89, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Diretto, G.; Al-Babili, S.; Tavazza, R.; Scossa, F.; Papacchioli, V.; Migliore, M.; Beyer, P.; Giuliano, G. Transcriptional-metabolic networks in beta-carotene-enriched potato tubers: The long and winding road to the golden phenotype. Plant Physiol. 2010, 154, 899–912. [Google Scholar] [CrossRef]
- Moghissi, A.A.; Pei, S.; Liu, Y. Golden Rice: Scientific, Regulatory and Public Information Processes of a Genetically Modified Organism. Crit. Rev. Biotechnol. 2016, 36, 535–541. [Google Scholar] [CrossRef]
- Govender, L.; Pillay, K.; Siwela, M.; Modi, A.T.; Mabhaudhi, T. Improving the Dietary Vitamin A Content of Rural Communities in South Africa by Replacing Non-Biofortified white Maize and Sweet Potato with Biofortified Maize and Sweet Potato in Traditional Dishes. Nutrients 2019, 11, 1198. [Google Scholar] [CrossRef]
- NAS, National Academy of Sciences. Dietary References Intakes: The Essential Guide to Nutrient Requirements. In Food and Nutrition Board; National Academic Press: Washington, DC, USA, 2006; Available online: http://www.nap.edu/catalog.php?record_id511537#toc (accessed on 25 June 2019).
- Underwood, B.A. Vitamin A Deficiency Disorders: International Efforts to Control a Preventable “Pox”. J. Nutr. 2004, 134, 231S–236S. [Google Scholar] [CrossRef]
- Sommer, A. Vitamin A deficiency and clinical disease: An historical overview. J. Nutr. 2008, 138, 1835–1839. [Google Scholar] [CrossRef]
- Weber, D.; Grune, T. The contribution of β-carotene to vitamin A supply of humans. Mol. Nutr. Food Res. 2012, 56, 251–258. [Google Scholar] [CrossRef]
- Priyadarshani, A.M. A Review on Factors Influencing Bioaccessibility and Bioefficacy of Carotenoids. Crit. Rev. Food Sci. Nutr. 2015, 13. [Google Scholar] [CrossRef]
- Barber, T.; Esteban-Pretel, G.; Marín, M.P.; Timoneda, J. Vitamin A: An Overview. In Vitamin A and Carotenoids: Chemistry, Analysis, Function and Effects; Preedy, V.R., Ed.; The Royal Society of Chemistry: London, UK, 2012; pp. 396–416. ISBN 978-1-84973-368-7. [Google Scholar] [CrossRef]
- McCauley, M.E.; van den Broek, N.; Dou, L.; Othman, M. Vitamin A supplementation during pregnancy for maternal and newborn outcomes. Cochrane Database Syst. Rev. 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- McLaren, D.S.; Kraemer, K. Manual on Vitamin Deficiency Disorders (VADD), 3rd ed.; Sight and Life Press: Basel, Switzerland, 2012; ISBN 978-3-906412-58-0. [Google Scholar]
- Lieber, C.S. Alcohol: Its metabolism and interaction with nutrients. Annu. Rev. Nutr. 2000, 20, 395–430. [Google Scholar] [CrossRef] [PubMed]
- Timoneda, J.; Rodríguez-Fernández, L.; Zaragozá, R.; Marín, M.P.; Cabezuelo, M.T.; Torres, L.; Viña, J.R.; Barber, T. Vitamin A Deficiency and the Lung. Nutrients 2018, 10, 1132. [Google Scholar] [CrossRef] [PubMed]
- Zielinska, M.A.; Hamulka, J.; Wesolowska, A. Month of Lactation and Its Associations with Maternal Dietary Intake and Anthropometric Characteristics. Nutrients 2019, 11, 193. [Google Scholar] [CrossRef]
- Allen, L.H. Multiple micronutrients in pregnancy and lactation: An overview. Am. J. Clin. Nutr. 2005, 81, 1206S–1212S. [Google Scholar] [CrossRef]
- Bravi, F.; Wiens, F.; Decarli, A.; Dal Pont, A.; Agostoni, C.; Ferraroni, M. Impact of maternal nutrition on breast-milk composition: A systematic review. Am. J. Clin. Nutr. 2016, 104, 646–662. [Google Scholar] [CrossRef]
- Erick, M. Breast milk is conditionally perfect. Med. Hypotheses 2018, 111, 82–89. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Nohr, D. The importance of vitamin A during pregnancy and childhood: Impact on lung function. In Vitamin A and Carotenoids: Chemistry, Analysis, Function and Effects; Preedy, V.R., Ed.; The Royal Society of Chemistry: London, UK, 2012; pp. 532–554. ISBN 978-1-84973-368-7. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Black, R.E. Hidden Hunger. Malnutrition and the First 1000 Days of Life: Causes, Consequences and Solutions. World Rev. Nutr. Diet 2016, 115, 134–141. [Google Scholar] [CrossRef]
- Davisse-Paturet, C.; Adel-Patient, K.; Divaret-Chauveau, A.; Pierson, J.; Lioret, S.; Cheminat, M.; Dufourg, M.-N.; Charles, M.-A.; de Lauzon-Guillain, B. Breastfeeding Status and Duration and Infections, Hospitalizations for Infections, and Antibiotic Use in the First Two Years of Life in the ELFE Cohort. Nutrients 2019, 11, 1607. [Google Scholar] [CrossRef]
- Soares, M.M.; Silva, M.A.; Garcia, P.P.C.; Silva, L.S.D.; Costa, G.D.D.; Araújo, R.M.A.; Cotta, R.M.M. Effect of vitamin A suplementation: A systematic review. Cien. Saude Colet. 2019, 24, 827–838. [Google Scholar] [CrossRef]
- Blomhoff, R.; Blomhoff, H.K. Overview of retinoid metabolism and function. J. Neurobiol. 2006, 66, 606–630. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, D.N.; Clugston, R.D.; Blaner, W.S. Vitamin A Metabolism: An Update. Nutrients 2011, 3, 63–103. [Google Scholar] [CrossRef] [PubMed]
- Blaner, W.S.; Li, Y.; Brun, P.J.; Yueng, J.J.; Lee, S.A.; Clugston, R.D. Vitamin A Absorption, Storage and Mobilization. In The Biochemistry of Retinoid Signaling II; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2016; Volume 81, pp. 95–126. [Google Scholar] [CrossRef]
- Saeed, A.; Dullaart, R.P.F.; Schreuder, T.C.M.A.; Blokzijl, H.; Faber, K.N. Disturbed Vitamin A Metabolism in Non-Alcoholic Fatty Liver Disease (NAFLD). Nutrients 2018, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Hang, J.; Rillema, J.A. Prolactin’s effects on lipoprotein lipase (LPL) activity and on LPL mRNA levels in cultured mouse mammary gland explants. Proc. Soc. Exp. Biol. Med. 1997, 214, 161–166. [Google Scholar] [CrossRef]
- Zhao, W.S.; Hu, S.L.; Yu, K.; Wang, H.; Wang, W.; Loor, J.; Luo, J. Lipoprotein Lipase, Tissue Expression and Effects on Genes Related to Fatty Acid Synthesis in Goat Mammary Epithelial Cells. Int. J. Mol. Sci. 2014, 15, 22757–22771. [Google Scholar] [CrossRef]
- Jensen, D.R.; Gavigan, S.; Sawicki, V.; Witsell, D.L.; Eckel, R.H.; Neville, M.C. Regulation of Lipoprotein lipase activity and mRNA in the mammary gland of the lactating mouse. Biochem. J. 1994, 298, 321–327. [Google Scholar] [CrossRef]
- Noy, N. Vitamin A transport and cell signaling by the retinol-binding protein receptor STRA6. In The Biochemistry of Retinoid Signaling II; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2016; Volume 81, pp. 77–94. [Google Scholar] [CrossRef]
- Inman, J.L.; Robertson, C.; Mott, J.D.; Bissell, M.J. Mammary gland development: Cell fate specification, stem cells and the microenvironment. Development 2015, 142, 1028–1042. [Google Scholar] [CrossRef]
- Fata, J.E.; Chaudhary, V.; Khokha, R. Cellular turnover in the mammary gland is correlated with systemic levels of progesterone and not 17beta-estradiol during the estrous cycle. Biol. Reprod. 2001, 65, 680–688. [Google Scholar] [CrossRef]
- Stingl, J.; Raouf, A.; Emerman, J.T.; Eaves, C.J. Epithelial progenitors in the normal human mammary gland. J. Mammary Gland Biol. Neoplasia 2005, 10, 49–59. [Google Scholar] [CrossRef]
- Cho, K.W.; Kwon, H.J.; Shin, J.O.; Lee, J.M.; Cho, S.W.; Tickle, C.; Jung, H.S. Retinoic acid signaling and the initiation of mammary gland development. Dev. Biol. 2012, 365, 259–266. [Google Scholar] [CrossRef]
- Wang, Y.A.; Shen, K.; Wang, Y.; Brooks, S.C. Retinoic acid signaling is required for proper morphogenesis of mammary gland. Dev. Dyn. 2005, 234, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Cohn, E.; Ossowski, L.; Bertran, S.; Marzan, C.; Farias, E.F. RARα1 control of mammary gland ductal morphogenesis and wnt1-tumorigenesis. Breast Cancer Res. 2010, 12, R79. [Google Scholar] [CrossRef] [PubMed]
- Honeth, G.; Lombardi, S.; Ginestier, C.; Hur, M.; Marlow, R.; Buchupalli, B.; Shinomiya, I.; Gazinska, P.; Bombelli, S.; Ramalingam, V.; et al. Aldehyde dehydrogenase and estrogen receptor define a hierarchy of cellular differentiation in the normal human mammary epithelium. Breast Cancer Res. 2014, 16, R52. [Google Scholar] [CrossRef] [PubMed]
- Pierzchalski, K.; Yu, J.; Norman, V.; Kane, M.A. CrbpI regulates mammary retinoic acid homeostasis and the mammary microenvironment. FASEB J. 2013, 27, 1904–1916. [Google Scholar] [CrossRef]
- Montesano, R.; Soulié, P. Retinoids induce lumen morphogenesis in mammary epithelial cells. J. Cell Sci. 2002, 115, 4419–4431. [Google Scholar] [CrossRef]
- Arisi, M.F.; Starker, R.A.; Addya, S.; Huang, Y.; Fernandez, S.V. All trans-retinoic acid (ATRA) induces re-differentiation of early transformed breast epithelial cells. Int. J. Oncol. 2014, 44, 1831–1842. [Google Scholar] [CrossRef]
- Zaragozá, R.; Gimeno, A.; Miralles, V.J.; García-Trevijano, E.R.; Carmena, R.; García, C.; Mata, M.; Puertes, I.R.; Torres, L.; Viña, J.R. Retinoids induce MMP-9 expression through RAR alpha during mammary gland remodeling. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1140–E1148. [Google Scholar] [CrossRef]
- Zaragozá, R.; García-Trevijano, E.R.; Lluch, A.; Ribas, G.; Viña, J.R. Involvement of Different networks in mammary gland involution after the pregnancy/lactation cycle: Implications in breast cancer. IUBMB Life 2015, 67, 227–238. [Google Scholar] [CrossRef]
- McNally, S.; Stein, T. Overview of Mammary Gland Development: A Comparison of Mouse and Human. Methods Mol. Biol. 2017, 1501, 1–17. [Google Scholar] [CrossRef]
- Jena, M.K.; Jaswal, S.; Kumar, S.; Mohanty, A.K. Molecular mechanism of mammary gland involution: An update. Dev. Biol. 2019, 445, 145–155. [Google Scholar] [CrossRef]
- Zile, M.H. Function of vitamin A in vertebrate embryonic development. J. Nutr. 2001, 131, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Torres, L.; Serna, E.; Bosch, A.; Zaragozá, R.; García, C.; Miralles, V.J.; Sandoval, J.; Viña, J.R.; García-Trevijano, E.R. NF-KB as a node for signal amplification during weaning. Cell Physiol. Biochem. 2011, 28, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Hughes, K.; Watson, C.J. The Multifaceted Role of STAT3 in Mammary Gland Involution and Breast Cancer. Int. J. Mol. Sci. 2018, 19, 1695. [Google Scholar] [CrossRef] [PubMed]
- Underwood, B.A. The Role of Vitamin A in Child Growth, Development and Survival. In Nutrient Regulation during Pregnancy, Lactation, and Infant Growth; Advances in Experimental Medicine and Biology; Allen, L., King, J., Lönnerdal, B., Eds.; Springer: Boston, MA, USA, 1994; Volume 352, pp. 201–208. [Google Scholar] [CrossRef]
- Report on Health and Social Subjects. 32. Present day practice in infant feeding: Third report. Report of a working party of the Panel on Child Nutrition Committee on Medical Aspects of Food Policy. Rep. Health Soc. Subj. 1988, 32, 1–66. [Google Scholar]
- De Vries, J.Y.; Pundir, S.; Mckenzie, E.; Keijer, J.; Kussmann, M. Maternal Circulating Vitamin Status and Colostrum Vitamin Composition in Healthy Lactating Women—A Systematic Approach. Nutrients 2018, 10, 687. [Google Scholar] [CrossRef] [PubMed]
- WHO, World Health Organization. Essential Nutrition. Actions: Improving Maternal, Newborn, Infant and Young Child Health and Nutrition; World Health Organization: Geneva, Switzerland, 2013; Available online: https://www.who.int/nutrition/publications/infantfeeding/essential_nutrition_actions/en/ (accessed on 20 May 2019).
- Ross, J.S.; Harvey, P.W. Contribution of breastfeeding to vitamin A nutrition of infants: A simulation model. Bull. World Health Organ. 2003, 81, 80–86. [Google Scholar]
- Da Silva, A.G.C.L.; de Sousa Rebouças, A.; Mendonça, B.M.A.; Silva, D.C.N.E.; Dimenstein, R.; Ribeiro, K.D.D.S. Relationship between the dietary intake, serum, and breast milk concentrations of vitamin A and vitamin E in a cohort of women over the course of lactation. Matern. Child Nutr. 2019, 15, e12772. [Google Scholar] [CrossRef]
- Williamson, D.H. Regulation of metabolism during lactation in the rat. Reprod. Nutr. Dev. 1986, 26, 597–603. [Google Scholar] [CrossRef][Green Version]
- Barber, T.; García de la Asunción, J.; Puertes, I.R.; Viña, J.R. Amino acid metabolism and protein synthesis in lactating rats fed on a liquid diet. Biochem. J. 1990, 270, 77–82. [Google Scholar] [CrossRef]
- Barber, T.; Triguero, A.; Martínez-López, I.; Torres, L.; García, C.; Miralles, V.J.; Viña, J.R. Elevated expression of liver gamma-cystathionase is required for the maintenance of lactation in rats. J. Nutr. 1999, 129, 928–933. [Google Scholar] [CrossRef]
- Canul-Medina, G.; Fernandez-Mejia, C. Morphological, hormonal, and molecular changes in different maternal tissues during lactation and post-lactation. J. Physiol. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zaragozá, R.; García, C.; Rus, A.D.; Pallardó, F.V.; Barber, T.; Torres, L.; Miralles, V.J.; Viña, J.R. Inhibition of liver trans-sulphuration pathway by propargylglycine mimics gene expression changes found in the mammary gland of weaned lactating rats: Role of glutathione. Biochem. J. 2003, 373, 825–834. [Google Scholar] [CrossRef] [PubMed]
- De la Asunción, J.G.; Devesa, A.; Viña, J.R.; Barber, T. Hepatic amino acid uptake is decreased in lactating rats. In vivo and in vitro studies. J. Nutr. 1994, 124, 2163–2171. [Google Scholar] [CrossRef] [PubMed]
- Piantedosi, R.; Ghyselinck, N.; Blaner, W.S.; Vogel, S. Cellular retinol-binding protein type III is needed for retinoid incorporation into milk. J. Biol. Chem. 2005, 280, 24286–24292. [Google Scholar] [CrossRef] [PubMed]
- Victora, C.G.; Bahl, R.; Barros, A.J.; França, G.V.; Horton, S.; Krasevec, J.; Murch, S.; Sankar, M.J.; Walker, N.; Rollins, N.C.; et al. Breastfeeding in the 21st century: Epidemiology, mechanisms, and lifelong effect. Lancet 2016, 387, 475–490. [Google Scholar] [CrossRef]
- Dror, D.K.; Allen, L.H. Retinol-to-Fat Ratio and Retinol Concentration in Human Milk Show Similar Time Trends and Associations with Maternal Factors at the Population Level: A Systematic Review and Meta-Analysis. Adv. Nutr. 2018, 9 (Suppl. 1), 332S–346S. [Google Scholar] [CrossRef]
- Van de Pavert, S.A.; Ferreira, M.; Domingues, R.G.; Ribeiro, H.; Molenaar, R.; Moreira-Santos, L.; Almeida, F.F.; Ibiza, S.; Barbosa, I.; Goverse, G.; et al. Maternal Retinoids Control Type 3 Innate Lymphoid Cells and Set the Offspring Immunity. Nature 2014, 508, 123–127. [Google Scholar] [CrossRef]
- Debier, C.; Larondelle, Y. Vitamins A and E: Metabolism, roles and transfer to offspring. Br. J. Nutr. 2005, 93, 153–174. [Google Scholar] [CrossRef]
- Lima, M.S.; da Silva Ribeiro, K.D.; Pires, J.F.; Bezerra, D.F.; Bellot, P.E.; de Oliveira Weigert, L.P.; Dimenstein, R. Breast milk retinol concentration in mothers of preterm newborns. Early Hum. Dev. 2017, 106–107, 41–45. [Google Scholar] [CrossRef]
- Bastos Maia, S.; Rolland Souza, A.S.; Costa Caminha, M.F.; Lins da Silva, S.; Callou Cruz, R.S.B.L.; Carvalho Dos Santos, C.; Batista Filho, M. Vitamin A and Pregnancy: A Narrative Review. Nutrients 2019, 11, 681. [Google Scholar] [CrossRef]
- Machado, M.R.; Kamp, F.; Nunes, J.C.; El-Bacha, T.; Torres, A.G. Breast Milk Content of Vitamin A and E from Early- to Mid-Lactation is Affected by Inadequate Dietary Intake in Brazilian Adult Women. Nutrients 2019, 11, 2025. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, D.S.; Ribeiro, K.D.S.; Lima, M.S.R.; Pires Medeiros, J.F.; da Silva, A.G.C.L.; Dimenstein, R.; Osório, M.M. Retinol status and associated factors in mother-newborn pairs. J. Hum. Nutr. Diet. 2019. [Google Scholar] [CrossRef] [PubMed]
- Hanson, C.; Lyden, E.; Anderson-Berry, A.; Kocmich, N.; Rezac, A.; Delair, S.; Furtado, J.; Van Ormer, M.; Izevbigie, N.; Olateju, E.K.; et al. Status of Retinoids and Carotenoids and Associations with Clinical Outcomes in Maternal-Infant Pairs in Nigeria. Nutrients 2018, 10, 1286. [Google Scholar] [CrossRef] [PubMed]
- Abebe, Z.; Haki, G.D.; Schweigert, F.J.; Henkel, I.M.; Baye, K. Low breastmilk vitamin A concentration is prevalent in rural Ethiopia. Eur. J. Clin. Nutr. 2019, 73, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Wessells, K.R.; Young, R.R.; Ferguson, E.L.; Ouédraogo, C.T.; Faye, M.T.; Hess, S.Y. Assessment of dietary intake and nutrient gaps, and development of food-based recommendations, among pregnant and lactating women in Zinder, Niger: An optifood linear programming analysis. Nutrients 2019, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Kaliwile, C.; Michelo, C.; Titcomb, T.J.; Moursi, M.; Donahue Angel, M.; Reinberg, C.; Bwembya, P.; Alders, R.; Tanumihardjo, S.A. Dietary intake patterns among lactating and non-lactating women of reproductive age in rural Zambia. Nutrients 2019, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Petry, N.; Jallow, B.; Sawo, Y.; Darboe, M.K.; Barrow, S.; Sarr, A.; Ceesay, P.O.; Fofana, M.N.; Prentice, A.M.; Wegmüller, R.; et al. Micronutrient Deficiencies, Nutritional Status and the Determinants of Anemia in Children 0–59 Months of Age and Non-Pregnant Women of Reproductive Age in The Gambia. Nutrients 2019, 11, 2275. [Google Scholar] [CrossRef]
- Meyer, S.; Gortner, L.; NeoVitaA Trial investigators. Up-date on the NeoVitaA Trial: Obstacles, challenges, perspectives, and local experiences. Wien. Med. Wochenschr. 2017, 167, 264–270. [Google Scholar] [CrossRef]
- Atalhi, N.; El Hamdouchi, A.; Barkat, A.; Elkari, K.; Hamrani, A.; El Mzibri, M.; Haskell, M.J.; Mokhtar, N.; Aguenaou, H. Combined consumption of a single high-dose vitamin A supplement with provision of vitamin A fortified oil to households maintains adequate milk retinol concentrations for 6 months in lactating Moroccan women. Appl. Physiol. Nutr. Metab. 2019. [Google Scholar] [CrossRef]
- WHO, World Health Organization. Micronutrients Deficiencies; World Health Organization: Geneva, Switzerland, 2018; Available online: http://www.who.int/nutrition/topics/vad/en/ (accessed on 5 May 2019).
- Copp, K.; DeFranco, E.A.; Kleiman, J.; Rogers, L.K.; Morrow, A.L.; Valentine, C.J. Nutrition Support Team Guide to Maternal Diet for the Human-Milk-Fed Infant. Nutr. Clin. Pract. 2018, 33, 687–693. [Google Scholar] [CrossRef]
- Hanson, C.; Schumacher, M.; Lyden, E.; Furtado, J.; Van Ormer, M.; McGinn, E.; Rilett, K.; Cave, C.; Johnson, R.; Weishaar, K.; et al. Status of vitamin A and related compounds and clinical outcomes in maternal-infant pairs in the midwestern United States. Ann. Nutr. Metab. 2017, 71, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Wolbach, S.B.; Howe, P.R. Tissue changes following deprivation of fat-soluble A vitamin. J. Exp. Med. 1925, 42, 753–777. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.G.; Roth, C.B.; Warkany, J. An analysis of the syndrome of malformations induced by maternal vitamin A deficiency. Effects of restoration of vitamin A at various times during gestation. Am. J. Anat. 1953, 92, 189–217. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.I.; Smith, J.E.; Winick, M.; Goodman, D.S. Vitamin A deficiency and fetal growth and development in the rat. J. Nutr. 1975, 105, 1299–1305. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabezuelo, M.T.; Zaragozá, R.; Barber, T.; Viña, J.R. Role of Vitamin A in Mammary Gland Development and Lactation. Nutrients 2020, 12, 80. https://doi.org/10.3390/nu12010080
Cabezuelo MT, Zaragozá R, Barber T, Viña JR. Role of Vitamin A in Mammary Gland Development and Lactation. Nutrients. 2020; 12(1):80. https://doi.org/10.3390/nu12010080
Chicago/Turabian StyleCabezuelo, M. Teresa, Rosa Zaragozá, Teresa Barber, and Juan R. Viña. 2020. "Role of Vitamin A in Mammary Gland Development and Lactation" Nutrients 12, no. 1: 80. https://doi.org/10.3390/nu12010080
APA StyleCabezuelo, M. T., Zaragozá, R., Barber, T., & Viña, J. R. (2020). Role of Vitamin A in Mammary Gland Development and Lactation. Nutrients, 12(1), 80. https://doi.org/10.3390/nu12010080