Associations between Dietary Patterns and Bile Acids—Results from a Cross-Sectional Study in Vegans and Omnivores

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Fecal Samples
2.2. Analysis of Bile Acids in Stool and Serum
2.3. Dietary Assessment
2.4. Assessment of Lifestyle Characteristics
2.5. Statistics
3. Results
3.1. Study Population
3.2. Intake of Macro-Nutrients and Food Groups Derived from Three-Day-Weighing Records
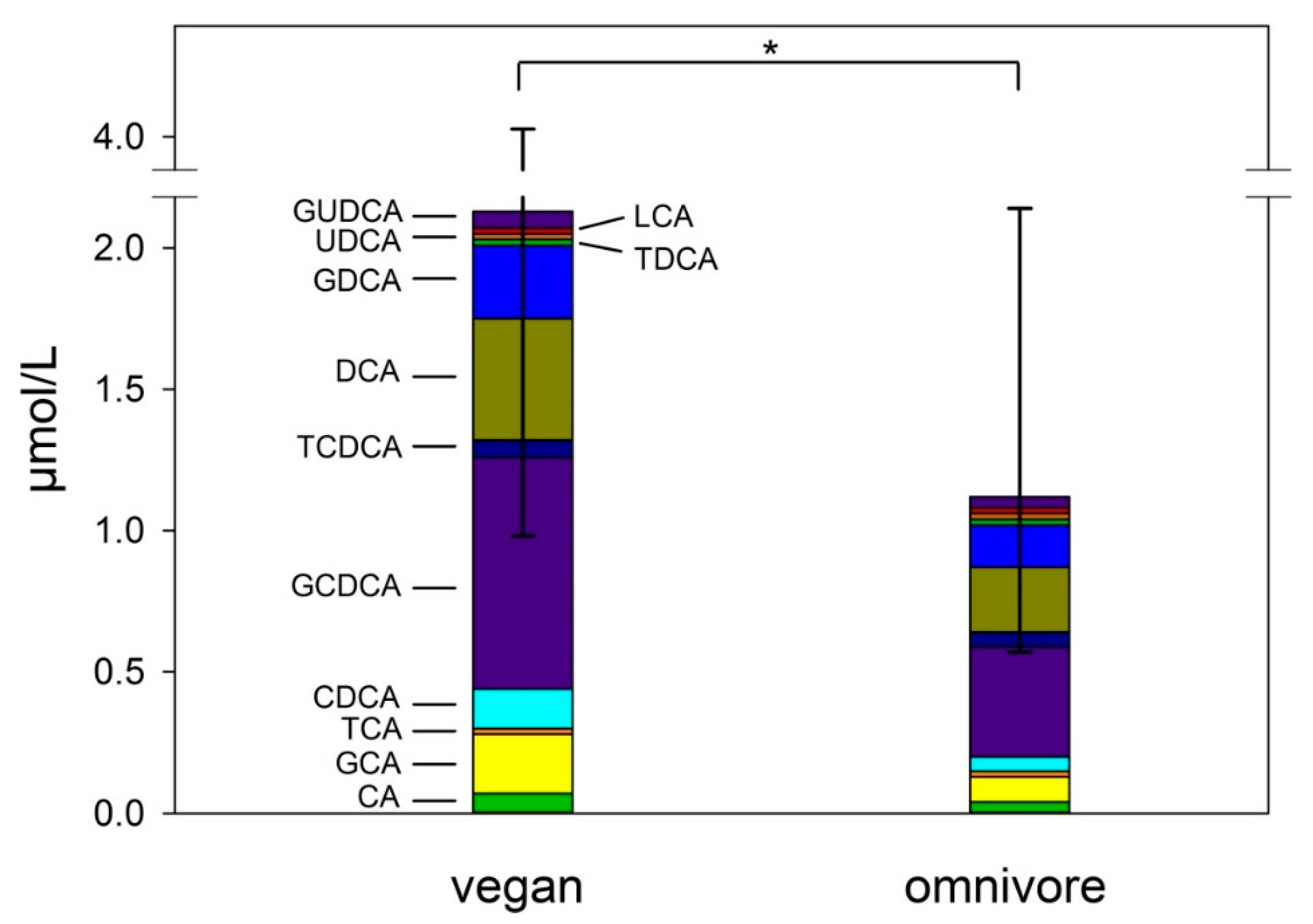
3.3. Concentrations of Bile Acids in Feces and Serum
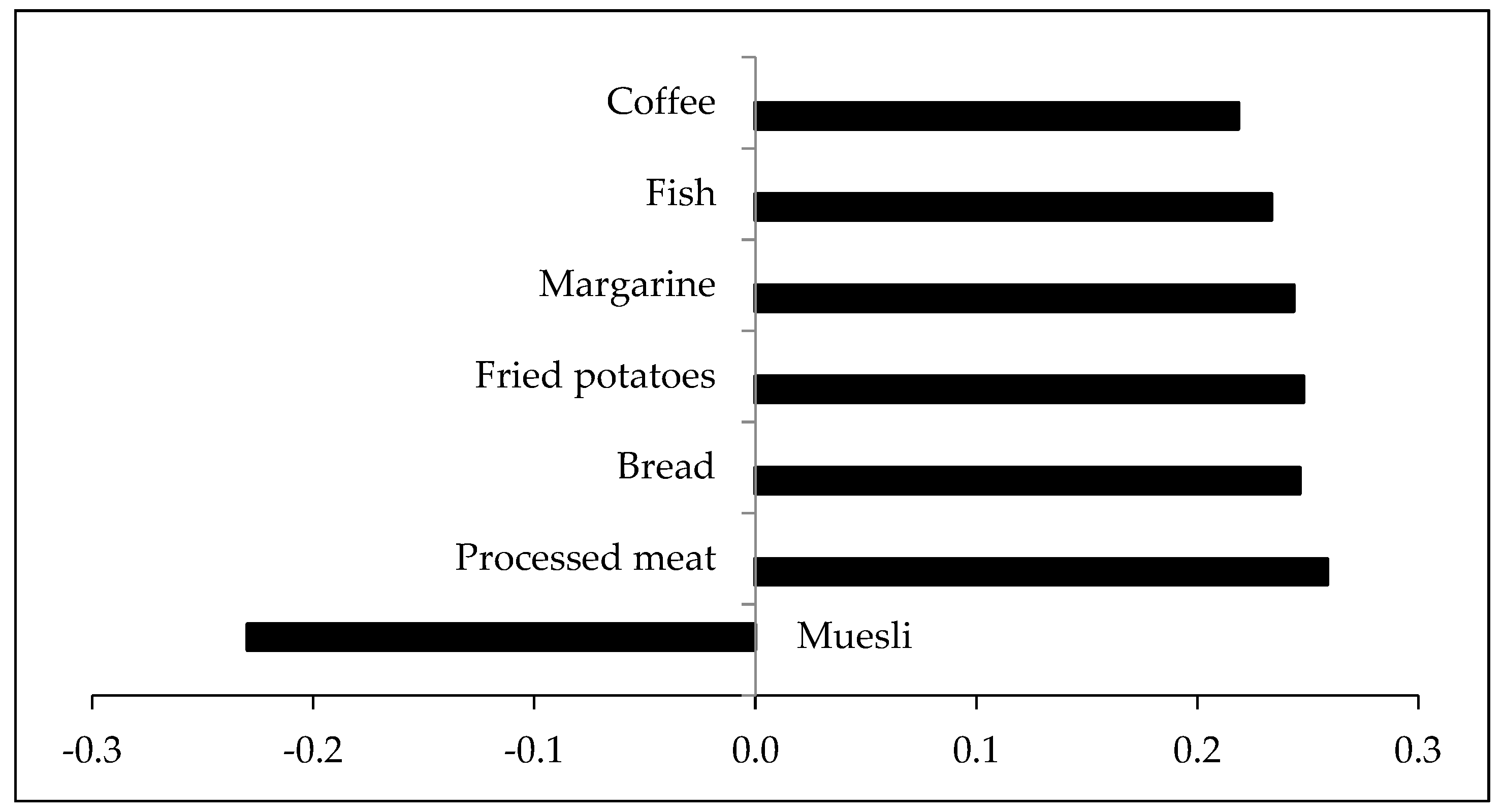
3.4. Dietary Pattern Explaining Variance in Fecal Bile Acids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mensink, G.; Lage Barbosa, C.; Brettschneider, A. Verbreitung der vegetarischen Ernährungsweise in Deutschland. J. Health Monit. 2016, 1, 2–15. [Google Scholar]
- Janssen, M.; Busch, C.; Rodiger, M.; Hamm, U. Motives of consumers following a vegan diet and their attitudes towards animal agriculture. Appetite 2016, 105, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Tonstad, S.; Butler, T.; Yan, R.; Fraser, G.E. Type of vegetarian diet, body weight, and prevalence of type 2 diabetes. Diabetes Care 2009, 32, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Le, L.T.; Sabate, J. Beyond meatless, the health effects of vegan diets: Findings from the Adventist cohorts. Nutrients 2014, 6, 2131–2147. [Google Scholar] [CrossRef] [PubMed]
- Appleby, P.N.; Key, T.J. The long-term health of vegetarians and vegans. Proc. Nutr. Soc. 2016, 75, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Benatar, J.R.; Stewart, R.A.H. Cardiometabolic risk factors in vegans; A meta-analysis of observational studies. PLoS ONE 2018, 13, e0209086. [Google Scholar] [CrossRef]
- Dinu, M.; Abbate, R.; Gensini, G.F.; Casini, A.; Sofi, F. Vegetarian, vegan diets and multiple health outcomes: A systematic review with meta-analysis of observational studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3640–3649. [Google Scholar] [CrossRef]
- World Cancer Research Fund International. Colorectal Cancer Statistics. Available online: https://www.wcrf.org/dietandcancer/cancer-trends/colorectal-cancer-statistics (accessed on 3 April 2019).
- Thanikachalam, K.; Khan, G. Colorectal cancer and nutrition. Nutrients 2019, 11, 164. [Google Scholar] [CrossRef]
- Singh, J.; Metrani, R.; Shivanagoudra, S.R.; Jayaprakasha, G.K.; Patil, B.S. Review on bile acids: Effects of the gut microbiome, interactions with dietary fiber, and alterations in the bioaccessibility of bioactive compounds. J. Agric. Food Chem. 2019, 67, 9124–9138. [Google Scholar] [CrossRef]
- O’Keefe, S.J.; Li, J.V.; Lahti, L.; Ou, J.; Carbonero, F.; Mohammed, K.; Posma, J.M.; Kinross, J.; Wahl, E.; Ruder, E.; et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 2015, 6, 6342. [Google Scholar] [CrossRef]
- Ou, J.; Carbonero, F.; Zoetendal, E.G.; DeLany, J.P.; Wang, M.; Newton, K.; Gaskins, H.R.; O’Keefe, S.J.D. Diet, microbiota, and microbial metabolites in colon cancer risk in rural Africans and African Americans. Am. J. Clin. Nutr. 2013, 98, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Thorning, T.K.; Raziani, F.; Bendsen, N.T.; Astrup, A.; Tholstrup, T.; Raben, A. Diets with high-fat cheese, high-fat meat, or carbohydrate on cardiovascular risk markers in overweight postmenopausal women: A randomized crossover trial. Am. J. Clin. Nutr. 2015, 102, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S. Diet and excretion of bile acids. Cancer Res. 1981, 41, 3766–3768. [Google Scholar] [PubMed]
- Chiang, J.Y. Recent advances in understanding bile acid homeostasis. F1000Research 2017, 6, 2029. [Google Scholar] [CrossRef] [PubMed]
- Hahn, C.; Reichel, C.; von Bergmann, K. Serum concentration of 7 alpha-hydroxycholesterol as an indicator of bile acid synthesis in humans. J. Lipid Res. 1995, 36, 2059–2066. [Google Scholar] [PubMed]
- Ridlon, J.M.; Wolf, P.G.; Gaskins, H.R. Taurocholic acid metabolism by gut microbes and colon cancer. Gut Microbes 2016, 7, 201–215. [Google Scholar] [CrossRef]
- Wahlstrom, A.; Sayin, S.I.; Marschall, H.U.; Backhed, F. Intestinal crosstalk between bile acids and microbiota and its impact on host metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef]
- Fiamoncini, J.; Yiorkas, A.M.; Gedrich, K.; Rundle, M.; Alsters, S.I.; Roeselers, G.; van den Broek, T.J.; Clavel, T.; Lagkouvardos, I.; Wopereis, S.; et al. Determinants of postprandial plasma bile acid kinetics in human volunteers. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 313, G300–G312. [Google Scholar] [CrossRef]
- De Aguiar Vallim, T.Q.; Tarling, E.J.; Edwards, P.A. Pleiotropic roles of bile acids in metabolism. Cell Metab. 2013, 17, 657–669. [Google Scholar] [CrossRef]
- Di Ciaula, A.; Garruti, G.; Lunardi Baccetto, R.; Molina-Molina, E.; Bonfrate, L.; Wang, D.Q.; Portincasa, P. Bile acid physiology. Ann. Hepatol. 2017, 16, s4–s14. [Google Scholar] [CrossRef]
- Ajouz, H.; Mukherji, D.; Shamseddine, A. Secondary bile acids: An underrecognized cause of colon cancer. World J. Surg. Oncol. 2014, 12, 164. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, H.; Bernstein, C.; Payne, C.M.; Dvorak, K. Bile acids as endogenous etiologic agents in gastrointestinal cancer. World J. Gastroenterol. 2009, 15, 3329–3340. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B.; Bajaj, J.S. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Ridlon, J.M.; Kang, D.J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef]
- Molinaro, A.; Wahlstrom, A.; Marschall, H.U. Role of bile acids in metabolic control. Trends Endocrinol. Metab. 2018, 29, 31–41. [Google Scholar] [CrossRef]
- Shapiro, H.; Kolodziejczyk, A.A.; Halstuch, D.; Elinav, E. Bile acids in glucose metabolism in health and disease. J. Exp. Med. 2018, 215, 383–396. [Google Scholar] [CrossRef]
- Hu, F.B. Dietary pattern analysis: A new direction in nutritional epidemiology. Curr. Opin. Lipidol. 2002, 13, 3–9. [Google Scholar] [CrossRef]
- Frank, L.K.; Jannasch, F.; Kroger, J.; Bedu-Addo, G.; Mockenhaupt, F.P.; Schulze, M.B.; Danquah, I. A dietary pattern derived by reduced rank regression is associated with type 2 diabetes in an urban Ghanaian population. Nutrients 2015, 7, 5497–5514. [Google Scholar] [CrossRef]
- Schulz, M.; Hoffmann, K.; Weikert, C.; Nothlings, U.; Schulze, M.B.; Boeing, H. Identification of a dietary pattern characterized by high-fat food choices associated with increased risk of breast cancer: The European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam study. Br. J. Nutr. 2008, 100, 942–946. [Google Scholar] [CrossRef]
- Hoffmann, K.; Schulze, M.B.; Schienkiewitz, A.; Nothlings, U.; Boeing, H. Application of a new statistical method to derive dietary patterns in nutritional epidemiology. Am. J. Epidemiol. 2004, 159, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Weikert, C.; Schulze, M.B. Evaluating dietary patterns: The role of reduced rank regression. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 341–346. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Van Faassen, A.; Bol, J.; Van Dokkum, W.; Pikaar, N.A.; Ockhuizen, T.; Hermus, R.J.J. Bile acids, neutral steroids, and bacteria in feces as affected by a mixed, a lacto-ovovegetarian, and a vegan diet. Am. J. Clin. Nutr. 1987, 46, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Bognar, A. Tables on Weight Yield of Food and Retention Factors of Food Constituents for the Calculation of Nutrient Composition of Cooked Foods (Dishes); Bundesforschungsanstalt für Ernährung Karlsruhe: Karlsruhe, Germany, 2002. [Google Scholar]
- Schulze, M.B.; Hoffmann, K.; Kroke, A.; Boeing, H. Dietary patterns and their association with food and nutrient intake in the European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam study. Br. J. Nutr. 2001, 85, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Kroger, J.; Ferrari, P.; Jenab, M.; Bamia, C.; Touvier, M.; Bueno-de-Mesquita, H.B.; Fahey, M.T.; Benetou, V.; Schulz, M.; Wirfalt, E.; et al. Specific food group combinations explaining the variation in intakes of nutrients and other important food components in the European prospective investigation into cancer and nutrition: An application of the reduced rank regression method. Eur. J. Clin. Nutr. 2009, 63 (Suppl. 4), S263–S274. [Google Scholar] [CrossRef]
- Weikert, C.; Hoffmann, K.; Dierkes, J.; Zyriax, B.C.; Klipstein-Grobusch, K.; Schulze, M.B.; Jung, R.; Windler, E.; Boeing, H. A homocysteine metabolism-related dietary pattern and the risk of coronary heart disease in two independent German study populations. J. Nutr. 2005, 135, 1981–1988. [Google Scholar] [CrossRef]
- Chen, W.; Wei, Y.; Xiong, A.; Li, Y.; Guan, H.; Wang, Q.; Miao, Q.; Bian, Z.; Xiao, X.; Lian, M.; et al. Comprehensive analysis of serum and fecal bile acid profiles and interaction with gut microbiota in primary biliary cholangitis. Clin. Rev. Allergy Immunol. 2019. [Google Scholar] [CrossRef]
- Luo, L.; Aubrecht, J.; Li, D.; Warner, R.L.; Johnson, K.J.; Kenny, J.; Colangelo, J.L. Assessment of serum bile acid profiles as biomarkers of liver injury and liver disease in humans. PLoS ONE 2018, 13, e0193824. [Google Scholar] [CrossRef]
- Wewalka, M.; Patti, M.E.; Barbato, C.; Houten, S.M.; Goldfine, A.B. Fasting serum taurine-conjugated bile acids are elevated in type 2 diabetes and do not change with intensification of insulin. J. Clin. Endocrinol. Metab. 2014, 99, 1442–1451. [Google Scholar] [CrossRef]
- Xie, G.; Wang, Y.; Wang, X.; Zhao, A.; Chen, T.; Ni, Y.; Wong, L.; Zhang, H.; Zhang, J.; Liu, C.; et al. Profiling of serum bile acids in a healthy Chinese population using UPLC-MS/MS. J. Proteome Res. 2015, 14, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Elorinne, A.L.; Alfthan, G.; Erlund, I.; Kivimaki, H.; Paju, A.; Salminen, I.; Turpeinen, U.; Voutilainen, S.; Laakso, J. Food and nutrient intake and nutritional status of Finnish vegans and non-vegetarians. PLoS ONE 2016, 11, e0148235. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, N.B.; Madsen, M.L.; Hansen, T.H.; Allin, K.H.; Hoppe, C.; Fagt, S.; Lausten, M.S.; Gobel, R.J.; Vestergaard, H.; Hansen, T.; et al. Intake of macro- and micronutrients in Danish vegans. Nutr. J. 2015, 14, 115. [Google Scholar] [CrossRef] [PubMed]
- Schupbach, R.; Wegmuller, R.; Berguerand, C.; Bui, M.; Herter-Aeberli, I. Micronutrient status and intake in omnivores, vegetarians and vegans in Switzerland. Eur. J. Nutr. 2017, 56, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Gunness, P.; Gidley, M.J. Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides. Food Funct. 2010, 1, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Naumann, S.; Schweiggert-Weisz, U.; Eglmeier, J.; Haller, D.; Eisner, P. In Vitro interactions of dietary fibre enriched food ingredients with primary and secondary bile acids. Nutrients 2019, 11, 1424. [Google Scholar] [CrossRef]
- Yang, I.F.; Jayaprakasha, G.K.; Patil, B.S. In vitro bile acid binding capacities of red leaf lettuce and cruciferous vegetables. J. Agric. Food Chem. 2017, 65, 8054–8062. [Google Scholar] [CrossRef]
- Naumann, S.; Schweiggert-Weisz, U.; Bader-Mittermaier, S.; Haller, D.; Eisner, P. Differentiation of adsorptive and viscous effects of dietary fibres on bile acid release by means of in vitro digestion and dialysis. Int. J. Mol. Sci. 2018, 19, 2193. [Google Scholar] [CrossRef]
- Breuninger, T.A.; Wawro, N.; Meisinger, C.; Artati, A.; Adamski, J.; Peters, A.; Grallert, H.; Linseisen, J. Associations between fecal bile acids, neutral sterols, and serum lipids in the KORA FF4 study. Atherosclerosis 2019, 288, 1–8. [Google Scholar] [CrossRef]
- Prawitt, J.; Caron, S.; Staels, B. Bile acid metabolism and the pathogenesis of type 2 diabetes. Curr. Diabetes Rep. 2011, 11, 160–166. [Google Scholar] [CrossRef]
- Rohrmann, S.; Linseisen, J. Processed meat: The real villain? Proc. Nutr. Soc. 2016, 75, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Mitry, P.; Wawro, N.; Sharma, S.; Kriebel, J.; Artati, A.; Adamski, J.; Heier, M.; Meisinger, C.; Thorand, B.; Grallert, H.; et al. Associations between usual food intake and faecal sterols and bile acids: Results from the Cooperative Health Research in the Augsburg Region (KORA FF4) study. Br. J. Nutr. 2019, 122, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Korpela, J.T.; Korpela, R.; Adlercreutz, H. Fecal bile acid metabolic pattern after administration of different types of bread. Gastroenterology 1992, 103, 1246–1253. [Google Scholar] [CrossRef]
- Vaughn, B.P.; Kaiser, T.; Staley, C.; Hamilton, M.J.; Reich, J.; Graiziger, C.; Singroy, S.; Kabage, A.J.; Sadowsky, M.J.; Khoruts, A. A pilot study of fecal bile acid and microbiota profiles in inflammatory bowel disease and primary sclerosing cholangitis. Clin. Exp. Gastroenterol. 2019, 12, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Thompson, F.E.; Subar, A.F. Chapter 1—Dietary assessment methodology. In Nutrition in the Prevention and Treatment of Disease, 4th ed.; Coulston, A.M., Boushey, C.J., Ferruzzi, M.G., Delahanty, L.M., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 5–48. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Vegan (n = 36) | Omnivore (n = 36) | p | |
|---|---|---|---|
| Male (%) | 50% | 50% | |
| Age (years) | 37.5 (32.5–44.0) | 38.5 (32.0–46.0) | 0.75 |
| Body weight (kg) | 70.1 ± 13.9 | 73.6 ± 10.3 | 0.24 |
| BMI | 22.9 ± 3.2 | 24.0 ± 2.1 | 0.08 |
| Cholesterol (mg/dL) | 165.9 ± 34.9 | 205.4 ± 41.6 | <0.0001 |
| Duration of vegan diet (years) | 4.8 (3.1–8.7) | n.a. | |
| Education (n (%)) | 0.60 | ||
| Low | 0 (0.0) | 1 (2.8) | |
| Intermediate | 11 (30.6) | 11 (30.6) | |
| High | 25 (69.5) | 24 (30.6) | |
| Physical activity (h/week) | 2.8 (0.88–3.75) | 2.3 (1.2–4.1) | 0.69 |
| Smoking behavior (n (%)) | 0.30 | ||
| Never smoker | 24 (66.7) | 21 (58.3) | |
| Ex-smoker | 8 (22.2) | 6 (16.7) | |
| Smoker | 4 (11.1) | 9 (25) | |
| Stool | |||
| Weight (mg) | 102.6 (39.9–185.9) | 97.3 (47.4–157.6) | 0.79 |
| Processing time (h:min) | 2:48 (1:49–3:45) | 2:32 (1:34–3:38) | 0.47 |
| Tertiles of Dietary Pattern Score | ||||
|---|---|---|---|---|
| Bile Acids (nmol/g) | 1 (75% Vegans) | 2 (50% Vegans) | 3 (25% Vegans) | p-Value for Trend |
| DCA | 74.23 (3.22–195.81) | 586.21 (283.57–1041.85) | 1327.30 (644.51–3037.96) | <0.0001 |
| LCA | 64.20 (13.99–93.47) | 198.93 (120.73–327.30) | 417.49 (305.10–1070.67) | <0.0001 |
| UDCA | 0.01 (0.00–0.16) | 0.16 (0.00–0.64) | 1.07 (0.00–16.91) | 0.02 |
| Total glycine-con. | 0.73 (0.32–1.52) | 3.45 (2.43–5.95) | 9.06 (5.76–18.81) | <0.0001 |
| Total taurine-con. | 0.26 (0.05–0.50) | 1.05 (0.49–2.00) | 3.25 (1.71–8.65) | <0.0001 |
| Tertiles of Dietary Pattern Score | ||||
|---|---|---|---|---|
| Macronutrients (g/day) | 1 (75% Vegans) | 2 (50% Vegans) | 3 (25% Vegans) | p-Value for Trend |
| Fiber | 37 (25–55) | 32 (27–50) | 24 (17–34) | 0.01 |
| Protein | 75 (53–122) | 90 (68–107) | 78 (69–103) | 0.43 |
| Fat | 92 (64–109) | 100 (83–124) | 98 (85–152) | 0.05 |
| Carbohydrates | 238 (203–310) | 236 (205–297) | 270 (211–356) | 0.37 |
| Sucrose | 46 (34–61) | 53 (37–68) | 60 (41–81) | 0.19 |
| Unsaturated fatty acids | 34 (23–40) | 34 (28–43) | 33 (27–49) | 0.25 |
| Saturated fatty acids | 19 (10–33) | 36 (16–44) | 34 (26–52) | 0.0005 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trefflich, I.; Marschall, H.-U.; Giuseppe, R.d.; Ståhlman, M.; Michalsen, A.; Lampen, A.; Abraham, K.; Weikert, C. Associations between Dietary Patterns and Bile Acids—Results from a Cross-Sectional Study in Vegans and Omnivores. Nutrients 2020, 12, 47. https://doi.org/10.3390/nu12010047
Trefflich I, Marschall H-U, Giuseppe Rd, Ståhlman M, Michalsen A, Lampen A, Abraham K, Weikert C. Associations between Dietary Patterns and Bile Acids—Results from a Cross-Sectional Study in Vegans and Omnivores. Nutrients. 2020; 12(1):47. https://doi.org/10.3390/nu12010047
Chicago/Turabian StyleTrefflich, Iris, Hanns-Ulrich Marschall, Romina di Giuseppe, Marcus Ståhlman, Andreas Michalsen, Alfonso Lampen, Klaus Abraham, and Cornelia Weikert. 2020. "Associations between Dietary Patterns and Bile Acids—Results from a Cross-Sectional Study in Vegans and Omnivores" Nutrients 12, no. 1: 47. https://doi.org/10.3390/nu12010047
APA StyleTrefflich, I., Marschall, H.-U., Giuseppe, R. d., Ståhlman, M., Michalsen, A., Lampen, A., Abraham, K., & Weikert, C. (2020). Associations between Dietary Patterns and Bile Acids—Results from a Cross-Sectional Study in Vegans and Omnivores. Nutrients, 12(1), 47. https://doi.org/10.3390/nu12010047