Anti-Obesity Effect of Ginkgo Vinegar, a Fermented Product of Ginkgo Seed Coat, in Mice Fed a High-Fat Diet and 3T3-L1 Preadipocyte Cells

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Histochemical Analysis
2.4. Cell Culture
2.5. Evaluation of Cytotoxicity and Differentiation
2.6. Western Blotting
2.7. Real-Time PCR
2.8. Statistical Analysis
3. Results
3.1. Suppressive Effect of Ginkgo Vinegar on Weight Gain in Mice
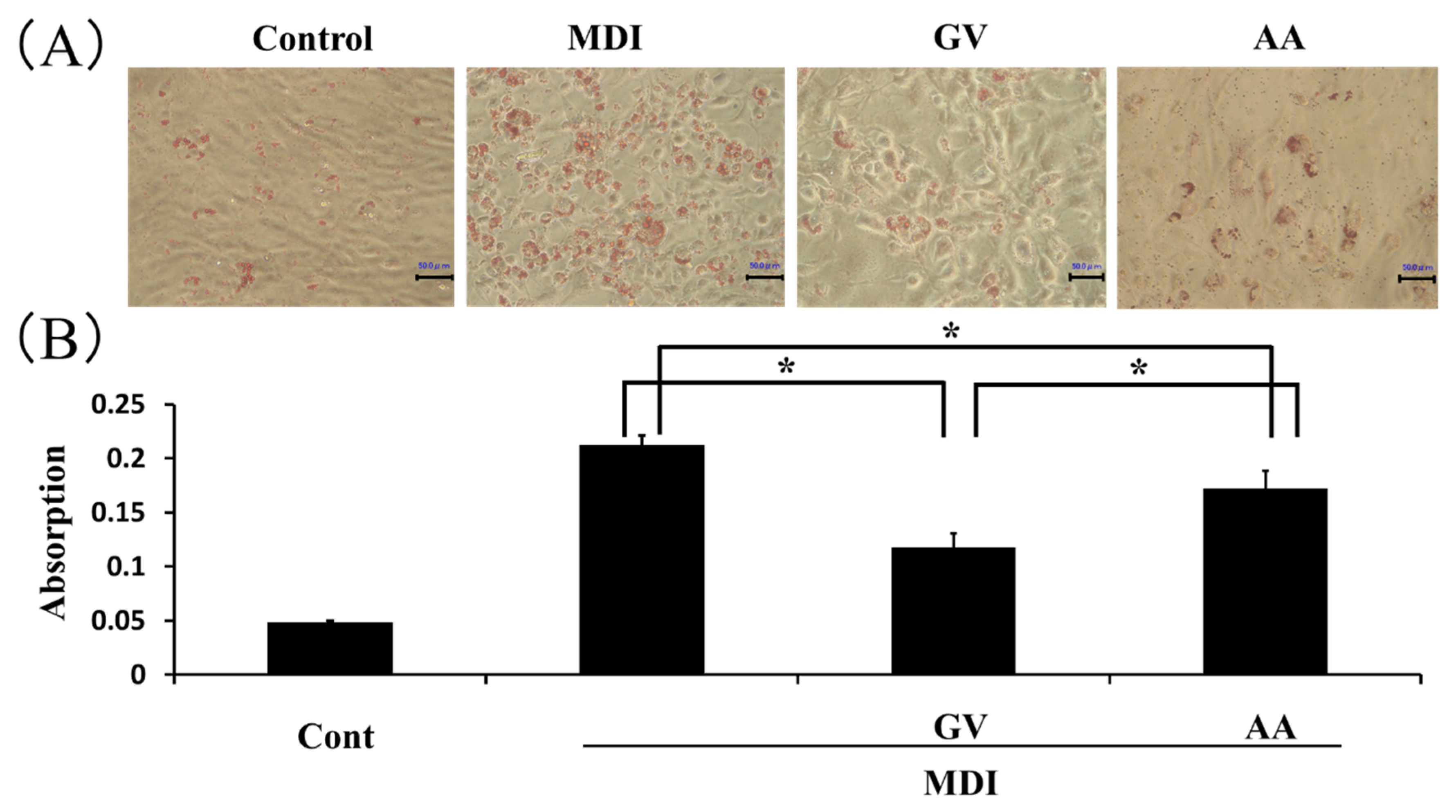
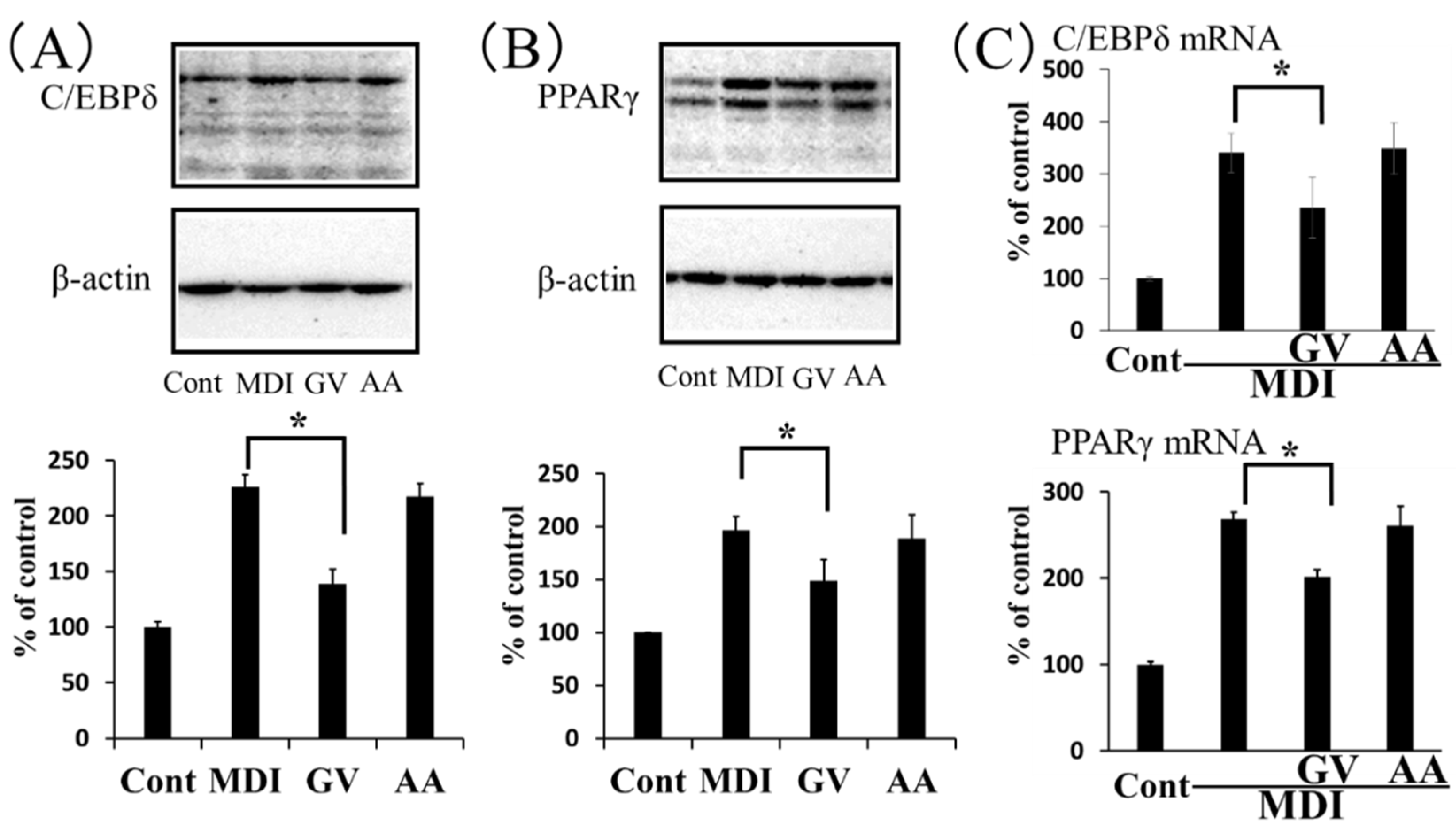
3.2. Effect of Ginkgo Vinegar on Adipocyte Differentiation In Vitro
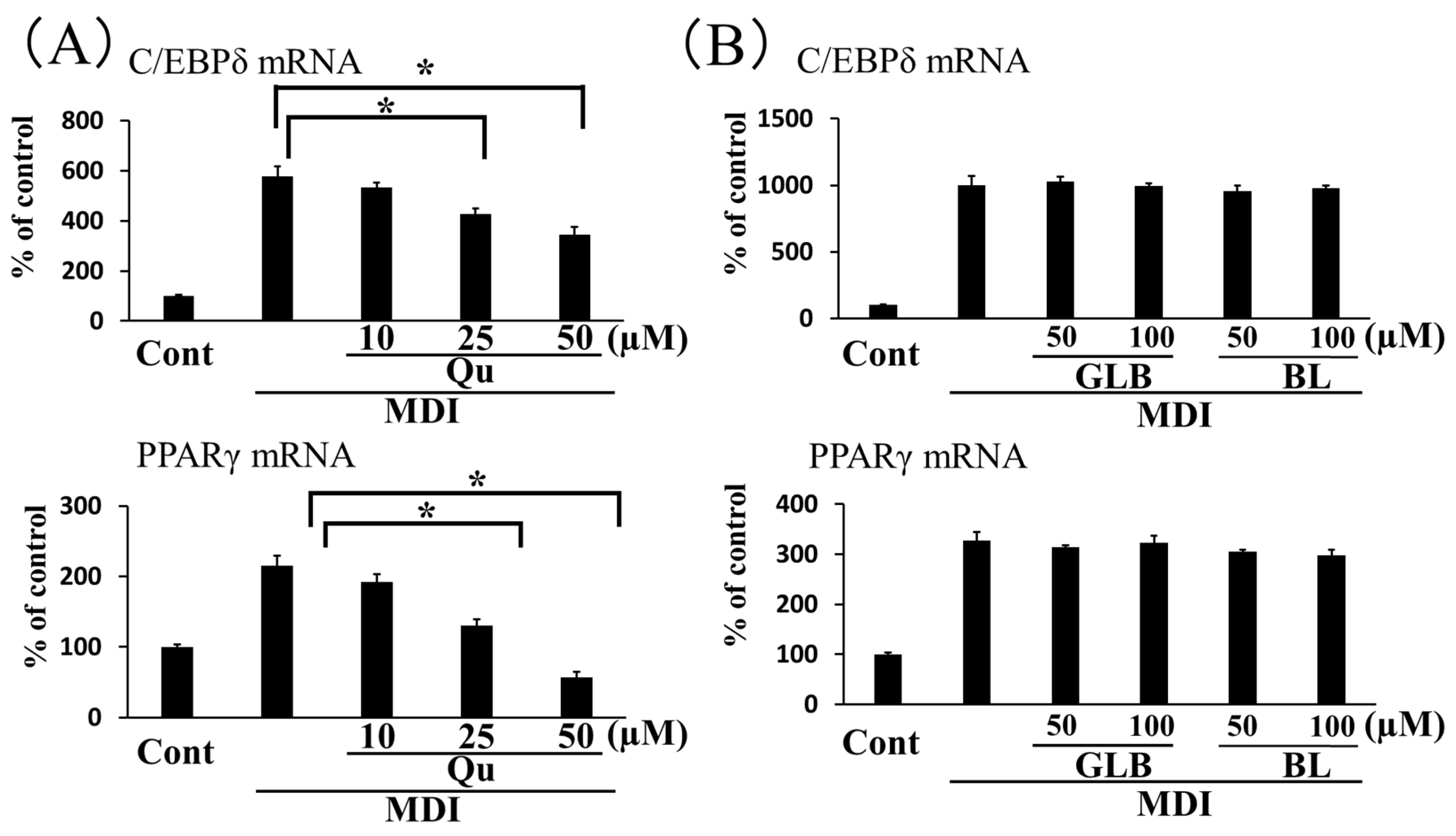
3.3. Effect of Components in Ginkgo Vinegar on Adipogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schötz, K. Quantification of allergenic urushiols in extracts of Ginkgo biloba leaves, in simple one-step extracts and refined manufactured material (EGb 761). Phytochem. Anal. 2004, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, D.; Yoshimura, T.; Johno, A.; Ishikawa, M.; Sasaki, K.; Wada, K. Decrease in pyridoxal-5′-phosphate concentration and increase in pyridoxal concentration in rat plasma by 4′-O-methylpyridoxine administration. Nutr. Res. 2015, 35, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Sado, T.; Nakata, S.; Tsuno, T.; Sato, M.; Misawa, Y.; Yamauchi, S.; Inaba, Y.; Kobayashi, D.; Wada, K. Concentrations of various forms of vitamin B6 in ginkgo seed poisoning. Brain Dev. 2019, 41, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.S.; Park, Y.J.; Kim, C.H.; Cho, Y.K.; Jang, M.J. Effect of seed disinfection on Bakanae disease in Ginkgo biloba outer seed coat 6 extract. Emir. J. Food Agric. 2016, 28, 671–675. [Google Scholar] [CrossRef]
- Forest, F.; Moat, J.; Baloch, E.; Brummitt, N.A.; Bachman, S.P.; Ickert-Bond, S.; Hollingsworth, P.M.; Liston, A.; Little, D.P.; Mathews, S.; et al. Gymnosperms on the EDGE. Sci. Rep. 2018, 8, 6053. [Google Scholar] [CrossRef]
- Jung, F.; Morwietz, C.; Kiesewetter, H.; Wenzel, E. Effect of Ginkgo biloba on fluidity of blood and peripheral microcirculation in volunteers. Drug Res. 1990, 40, 589–593. [Google Scholar]
- Guinot, P.; Caffrey, E.; Lambe, R.; Darragh, A. Tanakan inhibits platelet-activating-factor-induced platelet aggregation in healthy male volunteers. Haemostasis 1989, 19, 219–223. [Google Scholar] [CrossRef]
- Nunez, D.; Chignard, M.; Korth, R.; Le Couedic, J.P.; Norel, X.; Spinnewyn, B.; Braquet, P.; Benveniste, J. Specific inhibition of PAF-acether-induced platelet activation by BN 52021 and comparison with the PAF-acether inhibitors kadsurenone and CV 3988. Eur. J. Pharmacol. 1986, 123, 197–205. [Google Scholar] [CrossRef]
- Haramaki, N.; Aggarwal, S.; Kawabata, T.; DroyLefaix, M.T.; Packer, L. Effects of natural antioxidant Ginkgo biloba extract (EGB 761) on myocardial ischemia-reperfusion injury. Free Radic. Biol. Med. 1994, 16, 789–794. [Google Scholar] [CrossRef]
- Pietri, S.; Maurelli, E.; Drieu, K.; Culcasi, M. Cardioprotective and anti-oxidant effects of the terpenoid constituents of Ginkgo biloba extract (EGb761). J. Mol. Cell. Cardiol. 1997, 29, 733–742. [Google Scholar] [CrossRef]
- Friesen, J.B.; McAlpine, J.B.; Chen, S.N.; Pauli, G.F. Countercurrent separation of natural products. J. Nat. Prod. 2015, 78, 1765–1796. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.; Yan, F.; Zhang, B.; Li, J.; Mei, D. Advances in the studies of Ginkgo biloba leaves extract on aging-related diseases. Aging Dis. 2017, 8, 812–826. [Google Scholar] [CrossRef] [PubMed]
- Akemoto, N.; Sawada, K.; Shiba, H.; Matsumura, K. Vinegar Using Ginkgo, Production Method Therefor and Production Method for Alcohol Using Ginkgo. JP2013208113A, 2 March 2013. [Google Scholar]
- Barrea, L.; Muscogiuri, G.; Annunziata, G.; Laudisio, D.; Pugliese, G.; Salzano, C.; Colao, A.; Savastano, S. From gut microbiota dysfunction to obesity: Could short-chain fatty acids stop this dangerous course? Hormones 2019, 18, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, G.; Maisto, M.; Schisano, C.; Ciampaglia, R.; Daliu, P.; Narciso, V.; Tenore, G.C.; Novellino, E. Colon bioaccessibility and antioxidant activity of white, green and black tea polyphenols extract after in vitro simulated gastrointestinal digestion. Nutrients 2018, 10, 1711. [Google Scholar] [CrossRef]
- Yan, Z.; Fan, R.; Yin, S.; Zhao, X.; Liu, J.; Li, L.; Zhang, W.; Ge, L. Protective effects of Ginkgo biloba leaf polysaccharide on nonalcoholic fatty liver disease and its mechanisms. Int. J. Biol. Macromol. 2015, 80, 573–580. [Google Scholar] [CrossRef]
- Tian, J.; Popal, M.S.; Liu, Y.; Gao, R.; Lyu, S.; Chen, K.; Liu, Y. Ginkgo biloba leaf extract attenuates atherosclerosis in streptozotocin-induced diabetic ApoE-/- mice by inhibiting endoplasmic reticulum stress via restoration of autophagy through the mTOR signaling pathway. Oxid. Med. Cell. Longev. 2019, 2019, 8134678. [Google Scholar]
- Banin, R.M.; Hirata, B.K.; Andrade, I.S.; Zemdegs, J.C.; Clemente, A.P.; Dornellas, A.P.; Boldarine, V.T.; Estadella, D.; Albuquerque, K.T.; Oyama, L.M.; et al. Beneficial effects of Ginkgo biloba extract on insulin signaling cascade, dyslipidemia, and body adiposity of diet-induced obese rats. Braz. J. Med. Biol. Res. 2014, 47, 780–788. [Google Scholar] [CrossRef]
- Kondo, T.; Kishi, M.; Fushimi, T.; Ugajin, S.; Kaga, T. Vinegar intake reduces body weight, body fat mass, and serum triglyceride levels in obese Japanese subjects. Biosci. Biotechnol. Biochem. 2009, 73, 1837–1843. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef]
- Xie, H.; Wang, J.R.; Yau, L.F.; Liu, Y.; Liu, L.; Han, Q.B.; Zhao, Z.; Jiang, Z.H. Quantitative analysis of the flavonoid glycosides and terpene trilactones in the extract of Ginkgo biloba and evaluation of their inhibitory activity towards fibril formation of β-amyloid peptide. Molecules 2014, 19, 4466–4478. [Google Scholar] [CrossRef]
- Tan, M.S.; Yu, J.T.; Tan, C.C.; Wang, H.F.; Meng, X.F.; Wang, C.; Jiang, T.; Zhu, X.C.; Tan, L. Efficacy and adverse effects of ginkgo biloba for cognitive impairment and dementia: A systematic review and meta-analysis. J. Alzheimers Dis. 2015, 43, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.C.; Xia, Q.; Fu, P.P. Ginkgo biloba leave extract: Biological, medicinal, and toxicological effects. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2007, 25, 211–244. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Fujisawa, K.; Ito, E.; Idei, S.; Kawaguchi, N.; Kimoto, M.; Hiemori, M.; Tsuji, N. Involvement of obesity and glucose tolerance by acetate in type 2 diabetic Otsuka Long-Evans Tokushima Fatty (OLETF) rats. Biosci. Biotechnol. Biochem. 2007, 71, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A. Effects of quercetin supplementation on lipid profile: A systematic review and meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2017, 57, 666–676. [Google Scholar] [CrossRef]
- Chang, C.J.; Tzeng, T.F.; Liou, S.S.; Chang, Y.S.; Liu, I.M. Kaempferol regulates the lipid-profile in high-fat diet-fed rats through an increase in hepatic PPARα levels. Planta Med. 2011, 77, 1876–1882. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, M.; Cai, W.; Yu, L.; Feng, L.; Zhang, L.; Zang, Q.; Wang, Y.; Wang, D.; Chen, H.; et al. Dietary component isorhamnetin is a PPARγ antagonist and ameliorates metabolic disorders induced by diet or leptin deficiency. Sci. Rep. 2016, 6, 19288. [Google Scholar] [CrossRef]
- Ahn, J.; Lee, H.; Kim, S.; Park, J.; Ha, T. The anti-obesity effect of quercetin is mediated by the AMPK and MAPK signaling pathways. Biochem. Biophys. Res. Commun. 2008, 373, 545–549. [Google Scholar] [CrossRef]
- Liou, C.J.; Lai, X.Y.; Chen, Y.L.; Wang, C.L.; Wei, C.H.; Huang, W.C. Ginkgolide C suppresses adipogenesis in 3T3-L1 adipocytes via the AMPK signaling pathway. Evid. Based Complement. Altern. Med. 2015, 2015, 298635. [Google Scholar] [CrossRef]
- Bu, S.; Yuan, C.Y.; Xue, Q.; Chen, Y.; Cao, F. Bilobalide suppresses adipogenesis in 3T3-L1 adipocytes via the AMPK signaling pathway. Molecules 2019, 24, 3503. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosoda, S.; Kawazoe, Y.; Shiba, T.; Numazawa, S.; Manabe, A. Anti-Obesity Effect of Ginkgo Vinegar, a Fermented Product of Ginkgo Seed Coat, in Mice Fed a High-Fat Diet and 3T3-L1 Preadipocyte Cells. Nutrients 2020, 12, 230. https://doi.org/10.3390/nu12010230
Hosoda S, Kawazoe Y, Shiba T, Numazawa S, Manabe A. Anti-Obesity Effect of Ginkgo Vinegar, a Fermented Product of Ginkgo Seed Coat, in Mice Fed a High-Fat Diet and 3T3-L1 Preadipocyte Cells. Nutrients. 2020; 12(1):230. https://doi.org/10.3390/nu12010230
Chicago/Turabian StyleHosoda, Shugo, Yumi Kawazoe, Toshikazu Shiba, Satoshi Numazawa, and Atsufumi Manabe. 2020. "Anti-Obesity Effect of Ginkgo Vinegar, a Fermented Product of Ginkgo Seed Coat, in Mice Fed a High-Fat Diet and 3T3-L1 Preadipocyte Cells" Nutrients 12, no. 1: 230. https://doi.org/10.3390/nu12010230
APA StyleHosoda, S., Kawazoe, Y., Shiba, T., Numazawa, S., & Manabe, A. (2020). Anti-Obesity Effect of Ginkgo Vinegar, a Fermented Product of Ginkgo Seed Coat, in Mice Fed a High-Fat Diet and 3T3-L1 Preadipocyte Cells. Nutrients, 12(1), 230. https://doi.org/10.3390/nu12010230