Establishment of the Variation of Vitamin K Status According to Vkorc1 Point Mutations Using Rat Models

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Animal Treatment
2.3. Preparation of Liver Microsomes
2.4. Vitamin K Epoxide Reductase Activity (VKOR) Assays and Kinetics
2.5. VKORC1 and VKORC1L1 ELISA Quantitative Assays
2.6. Measurement of Vitamin K Concentrations
2.7. Undercarboxylated Osteocalcin (ucOC) and Undercarboxylated Matrix Gla Protein (ucMGP) ELISA Quantitative Assays
2.8. Tissue Calcium Measurement
2.9. Data Analysis
3. Results
3.1. Tissue VKOR Activity in Rats According to Vkorc1 Genotype
3.1.1. Kinetics Parameters of VKOR Activity
3.1.2. Ratio Between VKORC1 and VKORC1L1 Enzymes
3.2. Vitamin K Status According to the Vkorc1 Genotype in Rats fed with a Standard Rodent Diet
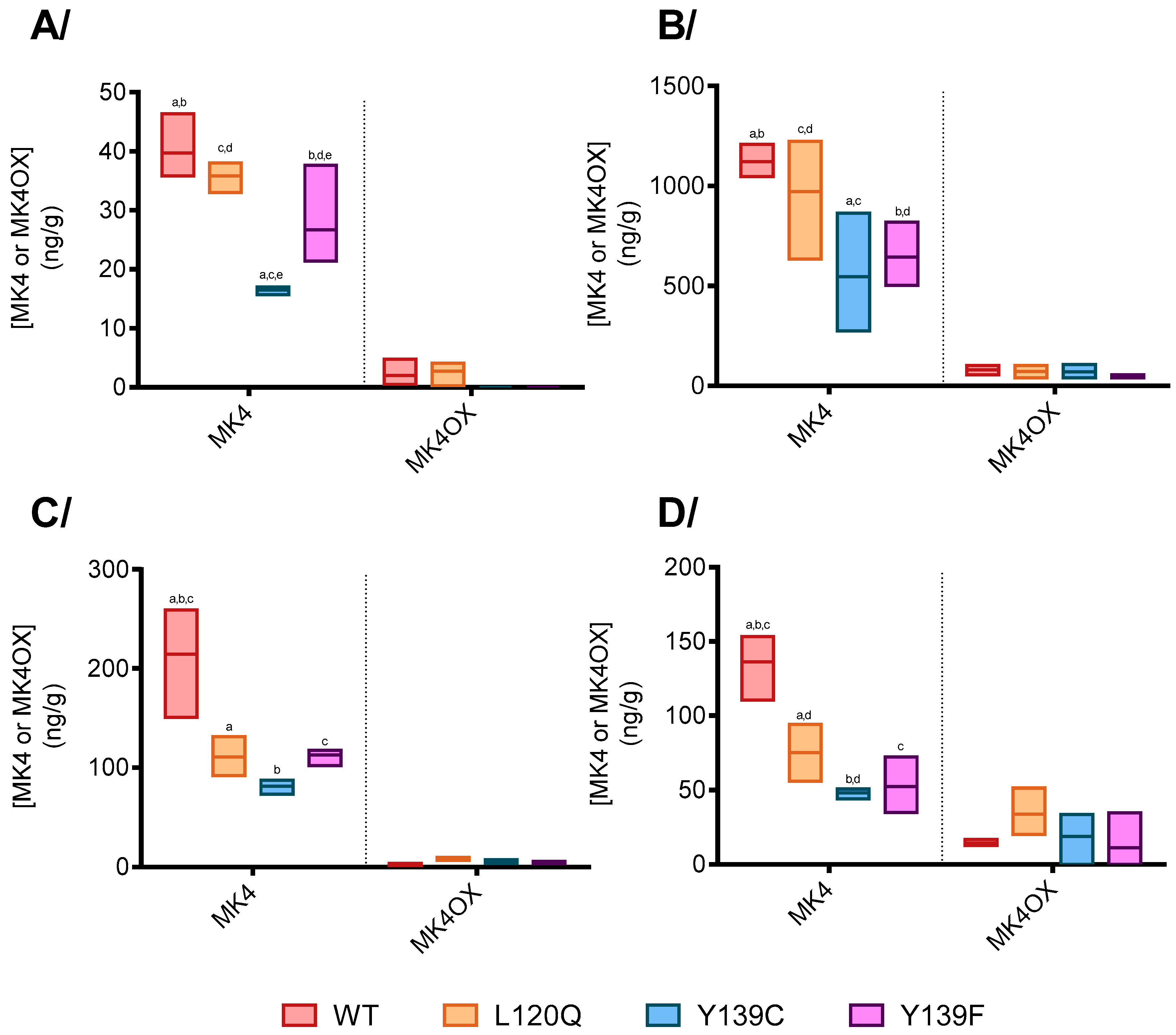
3.2.1. Vitamin K Concentrations
3.2.2. Plasma ucOC Concentration
3.2.3. Tissue Calcium Concentration
3.3. Evolution of the Vitamin K Status According to the Vkorc1 Genotype in Rats Receiving a Specific Diet-K3 for 12 Days
3.3.1. Vitamin K Concentrations
3.3.2. Prothrombin Time
3.3.3. Plasma ucOC Concentration
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Shearer, M.J.; Newman, P. Metabolism and cell biology of vitamin K. Thromb. Haemost. 2008, 100, 530–547. [Google Scholar] [PubMed]
- Stafford, D.W. The vitamin K cycle. J. Thromb. Haemost. 2005, 3, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zhang, B.L.; Liu, B.Y.; Shi, S.; Gao, D.Y.; Zhang, T.C.; Shi, H.J.; Li, Z.; Shum, W.W. Vitamin K2-Dependent GGCX and MGP Are Required for Homeostatic Calcium Regulation of Sperm Maturation. iScience 2019, 14, 210–225. [Google Scholar] [CrossRef] [PubMed]
- Van Summeren, M.J.; Braam, L.A.; Lilien, M.R.; Schurgers, L.J.; Kuis, W.; Vermeer, C. The effect of menaquinone-7 (vitamin K2) supplementation on osteocalcin carboxylation in healthy prepubertal children. Br. J. Nutr. 2009, 102, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Willems, B.A.; Vermeer, C.; Reutelingsperger, C.P.; Schurgers, L.J. The realm of vitamin K dependent proteins: Shifting from coagulation toward calcification. Mol. Nutr. Food Res. 2014, 58, 1620–1635. [Google Scholar] [CrossRef] [PubMed]
- Koshihara, Y.; Hoshi, K.; Shiraki, M. Vitamin K2 (menatetrenone) inhibits prostaglandin synthesis in cultured human osteoblast-like periosteal cells by inhibiting prostaglandin H synthase activity. Biochem. Pharmacol. 1993, 46, 1355–1362. [Google Scholar] [CrossRef]
- Hara, K.; Akiyama, Y.; Nakamura, T.; Murota, S.; Morita, I. The inhibitory effect of vitamin K2 (menatetrenone) on bone resorption may be related to its side chain. Bone 1995, 16, 179–184. [Google Scholar] [CrossRef]
- Reddi, K.; Henderson, B.; Meghji, S.; Wilson, M.; Poole, S.; Hopper, C.; Harris, M.; Hodges, S.J. Interleukin 6 production by lipopolysaccharide-stimulated human fibroblasts is potently inhibited by naphthoquinone (vitamin K) compounds. Cytokine 1995, 7, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lin, J.C.; Wang, H.; Peterson, J.W.; Furie, B.C.; Furie, B.; Booth, S.L.; Volpe, J.J.; Rosenberg, P.A. Novel role of vitamin k in preventing oxidative injury to developing oligodendrocytes and neurons. J. Neurosci. 2003, 23, 5816–5826. [Google Scholar] [CrossRef] [PubMed]
- Lev, M.; Milford, A.F. Effect of vitamin K depletion and restoration on sphingolipid metabolism in Bacteroides melaninogenicus. J. Lipid Res. 1972, 13, 364–370. [Google Scholar]
- Thomas, M.W.; Gebara, B.M. Difficulty breathing. Late onset vitamin K deficiency bleeding. Clin. Pediatr. 2004, 43, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Shearer, M.J.; Fu, X.; Booth, S.L. Vitamin K nutrition, metabolism, and requirements: Current concepts and future research. Adv. Nutr. 2012, 3, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Chatrou, M.L.; Reutelingsperger, C.P.; Schurgers, L.J. Role of vitamin K-dependent proteins in the arterial vessel wall. Hamostaseologie 2011, 31, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Crosier, M.D.; Peter, I.; Booth, S.L.; Bennett, G.; Dawson-Hughes, B.; Ordovas, J.M. Association of sequence variations in vitamin K epoxide reductase and gamma-glutamyl carboxylase genes with biochemical measures of vitamin K status. J. Nutr. Vitaminol. 2009, 55, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Stepien, E.; Branicka, A.; Ciesla-Dul, M.; Undas, A. A vitamin K epoxide reductase-oxidase complex gene polymorphism (−1639G > A) and interindividual variability in the dose-effect of vitamin K antagonists. J. Appl. Genet. 2009, 50, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Rost, S.; Fregin, A.; Ivaskevicius, V.; Conzelmann, E.; Hörtnagel, K.; Pelz, H.J.; Lappegard, K.; Seifried, E.; Scharrer, I.; Tuddenham, E.G.; et al. Mutations in VKORC1 cause warfarin resistance and multiple coagulation factor deficiency type 2. Nature 2004, 427, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Bodin, L.; Perdu, J.; Diry, M.; Horellou, M.H.; Loriot, M.A. Multiple genetic alterations in vitamin K epoxide reductase complex subunit 1 gene (VKORC1) can explain the high dose requirement during oral anticoagulation in humans. J. Thromb. Haemost. 2008, 6, 1436–1439. [Google Scholar] [CrossRef]
- Peoch, K.; Pruvot, S.; Gourmel, C.; Sollier, C.B.; Drouet, L. A new VKORC1mutation leading to an isolated resistance to fluindione. Br. J. Haematol. 2009, 145, 841–843. [Google Scholar] [CrossRef]
- Watzka, M.; Geisen, C.; Bevans, C.G.; Sittinger, K.; Spohn, G.; Rost, S.; Seifried, E.; Müller, C.R.; Oldenburg, J. Thirteen novel VKORC1 mutations associated with oral anticoagulant resistance: Insights into improved patient diagnosis and treatment. J. Throm. Haemost. 2011, 9, 109–118. [Google Scholar] [CrossRef]
- Harrington, D.J.; Siddiq, S.; Allford, S.L.; Shearer, M.J.; Mumford, A.D. More on: Endoplasmic reticulum loop VKORC1 substitutions cause warfarin resistance but do not diminish gamma-carboxylation of the vitamin K-dependent coagulation factors. J. Thromb. Haemost. 2011, 9, 1093–1095. [Google Scholar] [CrossRef]
- Chen, X.; Jin, D.Y.; Stafford, D.W.; Tie, J.K. Evaluation of oral anticoagulants with vitamin K epoxide reductase in its native milieu. Blood 2018, 132, 1974–1984. [Google Scholar] [CrossRef] [PubMed]
- Rost, S.; Pelz, H.J.; Menzel, S.; MacNicoll, A.D.; León, V.; Song, K.J.; Jäkel, T.; Oldenburg, J.; Müller, C.R. Novel mutations in the VKORC1 gene of wild rats and mice—A response to 50 years of selection pressure by warfarin? BMC Genet. 2009, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Michaux, A.; Matagrin, B.; Debaux, J.V.; Schurgers, L.J.; Benoit, E.; Lattard, V. Missense mutation of VKORC1 leads to medial arterial calcification in rats. Sci. Rep. 2018, 8, 13733. [Google Scholar] [CrossRef] [PubMed]
- Grandemange, A.; Kohn, M.H.; Lasseur, R.; Longin-Sauvageon, C.; Berny, P.; Benoit, E. Consequences of the Y139F Vkorc1 mutation on resistance to AVKs: In-vivo investigation in a 7th generation of congenic Y139F strain of rats. Pharm. Genom. 2009, 19, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Pelz, H.J.; Rost, S.; Hünerberg, M.; Fregin, A.; Heiberg, A.C.; Baert, K.; MacNicoll, A.D.; Prescott, C.V.; Walker, A.S.; Oldenburg, J.; et al. The genetic basis of resistance to anticoagulants in rodents. Genetics 2005, 170, 1839–1847. [Google Scholar] [CrossRef] [PubMed]
- Boitet, M.; Hammed, A.; Chatron, N.; Debaux, J.V.; Benoit, E.; Lattard, V. Elevated difenacoum metabolism is involved in the difenacoum-resistant phenotype observed in Berkshire rats homozygous for the L120Q mutation in the vitamin K epoxide reductase complex subunit 1 (Vkorc1) gene. Pest Manag. Sci. 2018, 74, 1328–1334. [Google Scholar] [CrossRef] [PubMed]
- Moroni, C.; Longin-Sauvageon, C.; Benoit, E. The Flavin-containing monooxygenase in rat liver: Evidence for the expression of a second form different from FMO1. Biochem. Biophys. Res. Commun. 1995, 212, 820–826. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of proteins utilizing the principle of protein dye binding. Nature 1976, 227, 248–254. [Google Scholar] [CrossRef]
- Goulois, J.; Lambert, V.; Legros, L.; Benoit, E.; Lattard, V. Adaptative evolution of the Vkorc1 gene in Mus musculus domesticus is influenced by the selective pressure of anticoagulant rodenticides. Ecol. Evol. 2017, 7, 2767–2776. [Google Scholar] [CrossRef]
- Hodroge, A.; Matagrin, B.; Moreau, C.; Fourel, I.; Hammed, A.; Benoit, E.; Lattard, V. VKORC1 mutations detected in patients resistant to vitamin K antagonists are not all associated with a resistant VKOR activity. J. Thromb. Haemost. 2012, 10, 2535–2543. [Google Scholar] [CrossRef]
- Chatron, N.; Chalmond, B.; Trouvé, A.; Benoit, E.; Caruel, H.; Lattard, V.; Tchertanov, L. dentification of the functional states of human vitamin K epoxide reductase from molecular dynamics simulations. RSC Adv. 2017, 82, 52071–52090. [Google Scholar] [CrossRef]
- Booth, S.L.; Peterson, J.W.; Smith, D.; Shea, M.K.; Chamberland, J.; Crivello, N. Age and dietary form of vitamin K affect menaquinone-4 concentrations in male Fisher 344 rats. J. Nutr. 2008, 138, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Matagrin, B.; Hodroge, A.; Montagut-Romans, A.; Andru, J.; Fourel, I.; Besse, S.; Benoit, E.; Lattard, V. New insights into the catalytic mechanism of vitamin K epoxide reductase (VKORC1)—The catalytic properties of the major mutations of rVKORC1 explain the biological cost associated to mutations. FEBS Open Biol. 2013, 16, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Hammed, A.; Matagrin, B.; Spohn, G.; Prouillac, C.; Benoit, E.; Lattard, V. VKORC1L1, an enzyme rescuing the vitamin K2,3-epoxide reductase activity in some extrahepatic tissues during anticoagulation therapy. J. Biol. Chem. 2013, 288, 28733–28742. [Google Scholar] [CrossRef] [PubMed]
- Hauschka, P.V.; Lian, J.B.; Cole, D.E.; Gundberg, C.M. Osteocalcin and matrix Gla protein: Vitamin K-dependent proteins in bone. Physiol. Rev. 1989, 69, 990–1047. [Google Scholar] [CrossRef]
- Haffa, A.; Krueger, D.; Bruner, J.; Engelke, J.; Gundberg, C.; Akhter, M.; Binkley, N. Diet- or warfarin-induced vitamin K insufficiency elevates circulating undercarboxylated osteocalcin without altering skeletal status in growing female rats. J. Bone Min. Res. 2000, 15, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Scheiber, D.; Veulemans, V.; Horn, P.; Chatrou, M.L.; Potthoff, S.A.; Kelm, M.; Schurgers, L.J.; Westenfeld, R. High-Dose Menaquinone-7 Supplementation Reduces Cardiovascular Calcification in a Murine Model of Extraosseous Calcification. Nutrients 2015, 7, 6991–7011. [Google Scholar] [CrossRef]
- Roumeliotis, S.; Dounousi, E.; Eleftheriadis, T.; Liakopoulos, V. Association of the inactive circulating matrix Gla Protein with vitamin K intake, calcification, mortality, and cardiovascular disease; a review. Int. J. Mol. Sci. 2019, 20, E628. [Google Scholar] [CrossRef] [PubMed]
- Edson, K.Z.; Prasad, B.; Unadkat, J.D.; Suhara, Y.; Okano, T.; Guengerich, F.P.; Rettie, A.E. Cytochrome P450-dependent catabolism of vitamin K: ω-hydroxylation catalyzed by human CYP4F2 and CYP4F11. Biochemistry 2013, 52, 8276–8285. [Google Scholar] [CrossRef]
- McDonald, M.G.; Rieder, M.J.; Nakano, M.; Hsia, C.K.; Rettie, A.E. CYP4F2 is a vitamin K1 oxidase: An explanation for altered warfarin dose in carriers of the V433M variant. Mol. Pharmacol. 2009, 75, 1337–1346. [Google Scholar] [CrossRef]
- Fasco, M.J.; Preusch, P.C.; Hildebrandt, E.; Suttie, J.W. Formation of hydroxyvitamin K by vitamin K epoxide reductase of warfarin-resistant rats. J. Biol. Chem. 1983, 258, 4372–4380. [Google Scholar] [PubMed]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature 2010, 468, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Hirota, Y.; Tsugawa, N.; Nakagawa, K.; Suhara, Y.; Tanaka, K.; Uchino, Y.; Takeuchi, A.; Sawada, N.; Kamao, M.; Wada, A.; et al. Menadione (vitamin K3) is a catabolic product of oral phylloquinone (vitamin K1) in the intestine and a circulating precursor of tissue menaquinone-4 (vitamin K2) in rats. J. Biol. Chem. 2013, 288, 33071–33080. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, F.; Collins, M.D. Vitamin K composition of anaerobic gut bacteria. FEMS Microbiol. Lett. 1987, 41, 175–180. [Google Scholar] [CrossRef]
- Lefebvre, S.; Hascoet, C.; Damin-Pernik, M.; Rannou, B.; Benoit, E.; Lattard, V. Monitoring of antivitamin K-dependent anticoagulation in rodents-Towards an evolution of the methodology to detect resistance in rodents. Pestic. Biochem. Physiol. 2017, 138, 29–36. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schurgers, L.J.; Cranenburg, E.C.; Vermeer, C. Matrix Gla-protein: The calcification inhibitor in need of vitamin K. Throm. Haemost. 2008, 100, 593–603. [Google Scholar]
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine regulation of energy metabolism by the skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Strain | Km (µM) | Vmax (pmol.min−1.mg−1) | Vmax/Km (µL.min−1.mg−1) |
|---|---|---|---|---|
| Liver | WT | 29.58 [18.96–40.21] | 95.62 a,b,c [84.13–107.1] | 3.2 a,b,c [2.2–4.2] |
| L120Q | 37.98 [20.45–55.52] | 22.18 a,d,e [18.45–25.91] | 0.6 a [0.4–0.8] | |
| Y139F | 22.52 a [14.49–30.56] | 34.50 b,e [30.62–38.39] | 1.6 b [1.3–1.9] | |
| Testis | Y139C | 46.89 a [30.64–53.13] | 34.52 c,d [29.97–39.07] | 0.8 c [0.5–1.1] |
| WT | 9.92 [4.00–15.84] | 53.26 a,b,c [45.03–61.49] | 5.5 a,b,c [5.3–5.7] | |
| L120Q | 16.70 [11.18–22.22] | 8.94 a,d [8.08–9.79] | 0.5 a [0.4–0.6] | |
| Y139F | 5.43 [2.31–8.55] | 8.86 b,e [6.53–11.19] | 1.2 b [0.6–1.8] | |
| Y139C | 12.35 [3.55–21.15] | 17.05 c,d,e [12.11–21.99] | 1.4 c [1.0–1.8] | |
| Kidney | WT | 12.12 [3.79–20.45] | 20.39 a,b,c [15.42–25.36] | 1.7 a,b [1.0–2.4] |
| L120Q | 23.32 [19.80–26.84] | 4.82 a,d [4.59–5.05] | 0.2a [0.1–0.3] | |
| Y139F | 12.25 [1.48–23.03] | 5.11 b,e [3.89–6.32] | 0.5 [0.1–0.9] | |
| Y139C | 23.24 [4.98–41.5] | 8.67 c,d,e [6.06–11.28] | 0.4b [0.2–0.6] | |
| Lung | WT | 13.16 [6.017–20.29] | 6.53 a,b [5.56–7.51] | 0.5 [0.2–0.8] |
| L120Q | 19.24 [14.84–23.63] | 3.94 a,c [3.67–4.21] | 0.20 [0.1–0.3] | |
| Y139F | 10.03 [3.4–16.66] | 4.11 b [2.87–5.35] | 0.4 [0.2–0.6] | |
| Y139C | 15.78 [3.06–28.50] | 6.43 c [4.59–8.26] | 0.4 [0.2–0.6] |
| Tissue | Strain | VKORC1 (µg/g of Tissue) | VKORC1L1 (µg/g of Tissue) | [VKORC1 Protein] |
|---|---|---|---|---|
| [VKORC1L1 Protein] | ||||
| Liver | WT | 4.44 ± 0.97 | 0.27 ± 0.04 | 16.2 ± 1.7 |
| L120Q | 2.66 ± 0.38 | 0.19 ± 0.03 | 14.0 ± 1.4 | |
| Y139F | 7.13 ± 2.30 * | 0.43 ± 0.13 | 17.6 ± 5.7 | |
| Y139C | 4.31 ± 0.80 | 0.30 ± 0.05 | 14.4 ± 2.2 | |
| Testis | WT | 0.25 ± 0.07 | 0.19 ± 0.04 | 1.4 ± 0.5 |
| L120Q | 0.35 ± 0.08 | 0.21 ± 0.09 | 1.6 ± 0.5 | |
| Y139F | 0.24 ± 0.10 | 0.22 ± 0.02 | 1.1 ± 0.5 | |
| Y139C | 0.27 ± 0.06 | 0.21 ± 0.04 | 1.3 ± 0.3 | |
| Kidney | WT | 0.10 ± 0.03 | 0.18 ± 0.02 | 0.54 ± 0.14 |
| L120Q | 0.14 ± 0.02 | 0.17 ± 0.01 | 0.82 ± 0.13 | |
| Y139F | 0.08 ± 0.04 | 0.23 ± 0.02 | 0.33 ± 0.19 | |
| Y139C | 0.12 ± 0.01 | 0.18 ± 0.01 | 0.67 ± 0.06 | |
| Lung | WT | 1.25 ± 0.12 | 0.70 ± 0.06 | 1.8 ± 0.2 |
| L120Q | 1.25 ± 0.22 | 1.35 ± 0.22 * | 0.9 ± 0.2 | |
| Y139F | 1.83 ± 0.54 | 1.22 ± 0.11 * | 1.5 ± 0.2 | |
| Y139C | 1.23 ± 0.19 | 1.10 ± 0.04 * | 1.1 ± 0.2 |
| Half-Life of MK4 | WT (in Days) | L120Q (in Days) | Y139F (in Days) | Y139C (in Days) |
|---|---|---|---|---|
| In testis | 12.09 a,b,c [10.03–13.82] | 3.10 a [2.22–5.11] | 3.31 b [2.07–8.21] | 2.50 c [1.69–4.74] |
| In kidney | 2.43 [1.78–3.84] | 3.85 [2.79–6.27] | 1.71 [1.31–2.47] | 1.68 [1.34–2.28] |
| In lung | 1.31 a,b [0.91–2.39] | 6.60 a [4.41–13.20] | 4.08 b [3.37–5.18] | 2.37 [1.37–8.93] |
 , decrease compared to WT strain;
, decrease compared to WT strain;  , increase for WT strain; =, similar with WT strain.
, decrease compared to WT strain; , increase for WT strain; =, similar with WT strain.
, increase for WT strain; =, similar with WT strain.
, decrease compared to WT strain; , increase for WT strain; =, similar with WT strain.| L120Q | Y139F | Y139C | |
|---|---|---|---|
| VKOR ACTIVITY | |||
| In liver | | | |
| In testis | | | |
| In kidney | | | |
| In lung | | | = |
| VKORC1 CONCENTRATION | |||
| In liver | = | | = |
| In testis | = | = | = |
| In kidney | = | = | = |
| In lung | = | = | = |
| VKORC1L1 CONCENTRATION | |||
| In liver | = | = | = |
| In testis | = | = | = |
| In kidney | = | = | = |
| In lung | | | |
| MK4 CONCENTRATION | |||
| In liver | = | | |
| In testis | = | | |
| In kidney | | | |
| In lung | | | |
| PLASMA ucOC CONCENTRATION | =to | | |
| TISSUE CALCIUM CONCENTRATION | |||
| In liver | = | = | = |
| In testis | = | = | = |
| In kidney | = | = | = |
| In lung | = | = | = |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Debaux, J.V.; Hammed, A.; Barbier, B.; Chetot, T.; Benoit, E.; Lefebvre, S.; Lattard, V. Establishment of the Variation of Vitamin K Status According to Vkorc1 Point Mutations Using Rat Models. Nutrients 2019, 11, 2076. https://doi.org/10.3390/nu11092076
Debaux JV, Hammed A, Barbier B, Chetot T, Benoit E, Lefebvre S, Lattard V. Establishment of the Variation of Vitamin K Status According to Vkorc1 Point Mutations Using Rat Models. Nutrients. 2019; 11(9):2076. https://doi.org/10.3390/nu11092076
Chicago/Turabian StyleDebaux, Jean Valéry, Abdessalem Hammed, Brigitte Barbier, Thomas Chetot, Etienne Benoit, Sébastien Lefebvre, and Virginie Lattard. 2019. "Establishment of the Variation of Vitamin K Status According to Vkorc1 Point Mutations Using Rat Models" Nutrients 11, no. 9: 2076. https://doi.org/10.3390/nu11092076
APA StyleDebaux, J. V., Hammed, A., Barbier, B., Chetot, T., Benoit, E., Lefebvre, S., & Lattard, V. (2019). Establishment of the Variation of Vitamin K Status According to Vkorc1 Point Mutations Using Rat Models. Nutrients, 11(9), 2076. https://doi.org/10.3390/nu11092076