The Influence of Early Nutrition on Brain Growth and Neurodevelopment in Extremely Preterm Babies: A Narrative Review


Abstract
1. Introduction
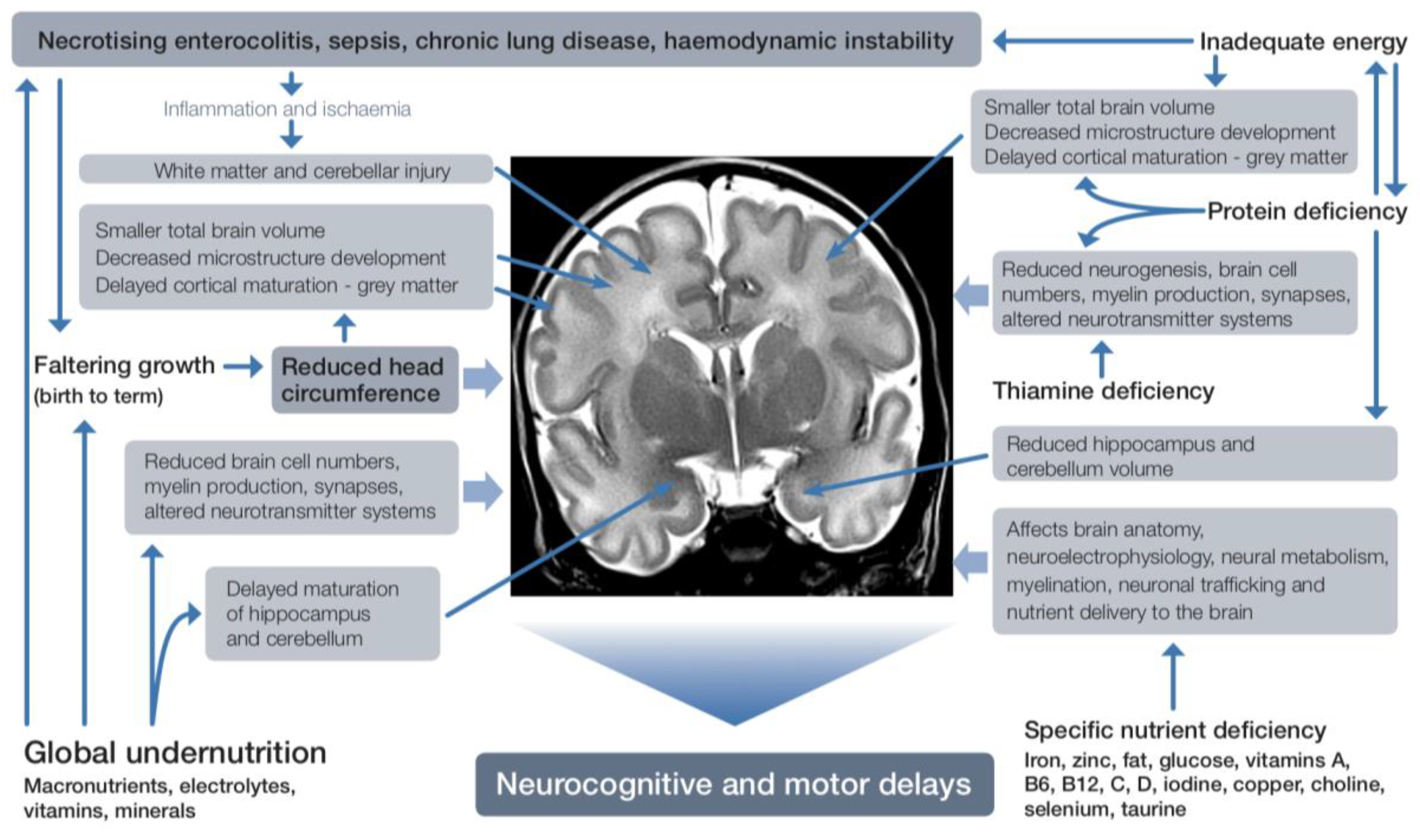
2. Effects of Preterm Birth on the Brain
3. Postnatal Factors that Play a Role in Neurodevelopment
4. Early Nutrition Affects Morbidity and Neurodevelopment
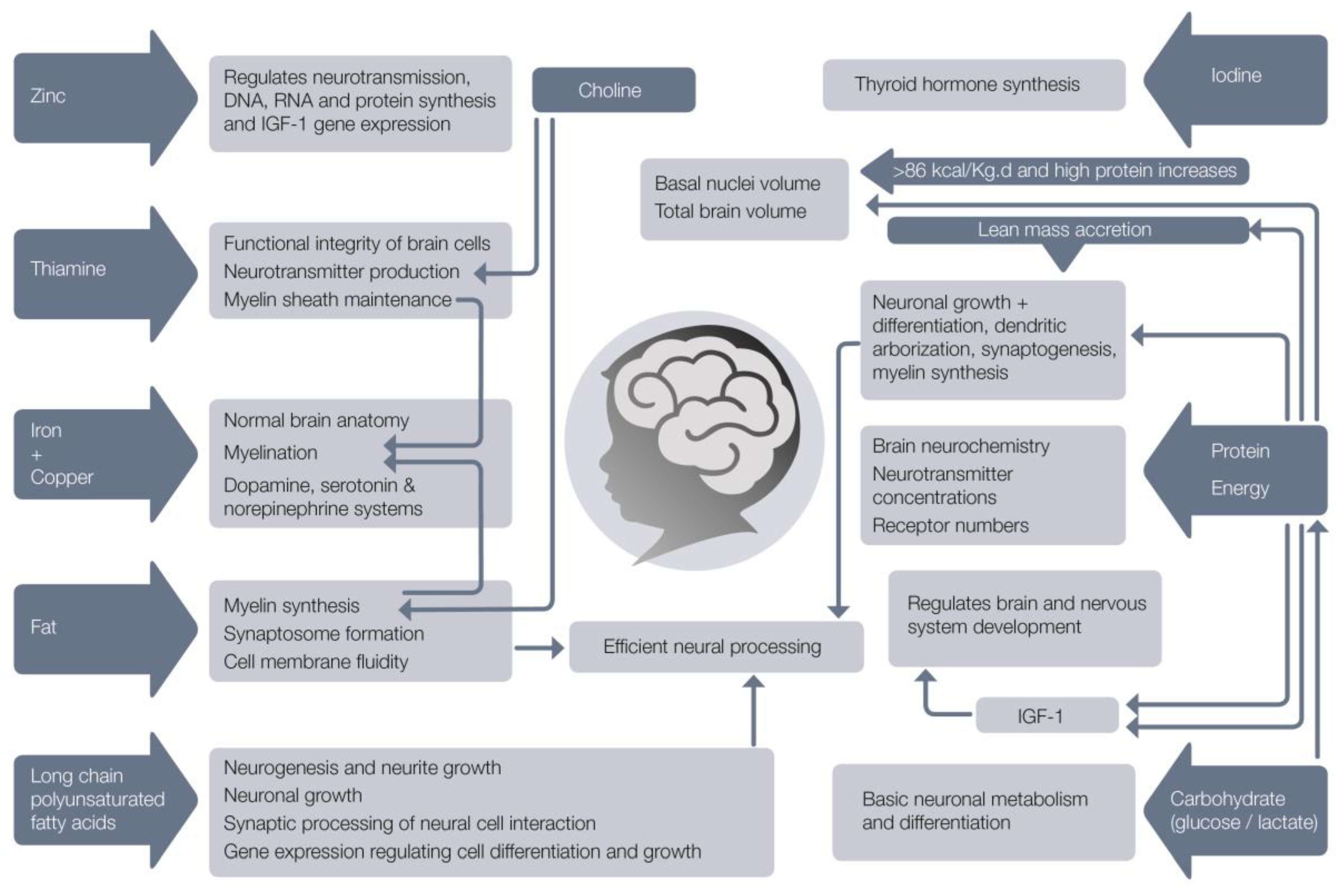
5. The Role of Specific Nutrients in Brain Development
6. Early Growth Is Related to Neurodevelopment
7. Evidence of Nutritional Effects on the Brain from Brain Imaging
8. Protein and Amino Acid Requirements
9. Direct Effects of Protein on Neurodevelopment
10. Studies of Higher vs. Lower Protein Intakes
11. Possible Reasons Why Research Has Not Yet Shown Whether We Can Improve Neurodevelopment with Nutrition Interventions
12. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clouchoux, C.; Guizard, N.; Evans, A.C.; du Plessis, A.J.; Limperopoulos, C. Normative fetal brain growth by quantitative in vivo magnetic resonance imaging. Am. J. Obs. Gynecol. 2012, 206, 173.e1–173.e8. [Google Scholar] [CrossRef]
- Dubois, J.; Benders, M.; Cachia, A.; Lazeyras, F.; Ha-Vinh Leuchter, R.; Sizonenko, S.V.; Borradori-Tolsa, C.; Mangin, J.F.; Huppi, P.S. Mapping the early cortical folding process in the preterm newborn brain. Cereb. Cortex 2008, 18, 1444–1454. [Google Scholar] [CrossRef] [PubMed]
- Volpe, J.J. Brain injury in premature infants: A complex amalgam of destructive and developmental disturbances. Lancet Neurol. 2009, 8, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Back, S.A.; Miller, S.P. Brain injury in premature neonates: A primary cerebral dysmaturation disorder? Ann. Neurol. 2014, 75, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Z.; Ashburner, J.; Andersson, J.; Jbabdi, S.; Draganski, B.; Skare, S.; Bohm, B.; Smedler, A.C.; Forssberg, H.; Lagercrantz, H. Structural correlates of preterm birth in the adolescent brain. Pediatrics 2009, 124, e964–e972. [Google Scholar] [CrossRef]
- Lubsen, J.; Vohr, B.; Myers, E.; Hampson, M.; Lacadie, C.; Schneider, K.C.; Katz, K.H.; Constable, R.T.; Ment, L.R. Microstructural and functional connectivity in the developing preterm brain. Semin. Perinatol. 2011, 35, 34–43. [Google Scholar] [CrossRef]
- Bassi, L.; Chew, A.; Merchant, N.; Ball, G.; Ramenghi, L.; Boardman, J.; Allsop, J.M.; Doria, V.; Arichi, T.; Mosca, F.; et al. Diffusion tensor imaging in preterm infants with punctate white matter lesions. Pediatr. Res. 2011, 69, 561–566. [Google Scholar] [CrossRef]
- Moore, T.; Hennessy, E.M.; Myles, J.; Johnson, S.J.; Draper, E.S.; Costeloe, K.L.; Marlow, N. Neurological and developmental outcome in extremely preterm children born in England in 1995 and 2006: The EPICure studies. BMJ 2012, 345, e7961. [Google Scholar] [CrossRef]
- Aarnoudse-Moens, C.S.; Weisglas-Kuperus, N.; van Goudoever, J.B.; Oosterlaan, J. Meta-analysis of neurobehavioral outcomes in very preterm and/or very low birth weight children. Pediatrics 2009, 124, 717–728. [Google Scholar] [CrossRef]
- Aarnoudse-Moens, C.S.; Smidts, D.P.; Oosterlaan, J.; Duivenvoorden, H.J.; Weisglas-Kuperus, N. Executive function in very preterm children at early school age. J. Abnorm. Child Psychol. 2009, 37, 981–993. [Google Scholar] [CrossRef]
- De Kieviet, J.F.; Piek, J.P.; Aarnoudse-Moens, C.S.; Oosterlaan, J. Motor development in very preterm and very low-birth-weight children from birth to adolescence: A meta-analysis. J. Am. Med. Assoc. 2009, 302, 2235–2242. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.; Doyle, L.W.; Victorian Infant Collaborative Study Group. Neurobehavioral outcomes of school-age children born extremely low birth weight or very preterm in the 1990s. J. Am. Med. Assoc. 2003, 289, 3264–3272. [Google Scholar] [CrossRef] [PubMed]
- Baron, I.S.; Erickson, K.; Ahronovich, M.D.; Baker, R.; Litman, F.R. Neuropsychological and behavioral outcomes of extremely low birth weight at age three. Dev. Neuropsychol. 2011, 36, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Gidley Larson, J.C.; Baron, I.S.; Erickson, K.; Ahronovich, M.D.; Baker, R.; Litman, F.R. Neuromotor outcomes at school age after extremely low birth weight: Early detection of subtle signs. Neuropsychology 2011, 25, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Marlow, N.; Hennessy, E.M.; Bracewell, M.A.; Wolke, D.; EPICure Study Group. Motor and executive function at 6 years of age after extremely preterm birth. Pediatrics 2007, 120, 793–804. [Google Scholar] [CrossRef]
- Mikkola, K.; Ritari, N.; Tommiska, V.; Salokorpi, T.; Lehtonen, L.; Tammela, O.; Paakkonen, L.; Olsen, P.; Korkman, M.; Fellman, V. Neurodevelopmental outcome at 5 years of age of a national cohort of extremely low birth weight infants who were born in 1996–1997. Pediatrics 2005, 116, 1391–1400. [Google Scholar] [CrossRef]
- Farooqi, A.; Adamsson, M.; Serenius, F.; Hagglof, B. Executive functioning and learning skills of adolescent children born at fewer than 26 weeks of gestation. PLoS ONE 2016, 11, e0151819. [Google Scholar] [CrossRef]
- Stoll, B.J.; Hansen, N.I.; Adams-Chapman, I.; Fanaroff, A.A.; Hintz, S.R.; Vohr, B.; Higgins, R.D. Neurodevelopmental and growth impairment among extremely low-birth-weight infants with neonatal infection. JAMA 2004, 292, 2357–2365. [Google Scholar] [CrossRef]
- Johnson, S.; Fawke, J.; Hennessy, E.; Rowell, V.; Thomas, S.; Wolke, D.; Marlow, N. Neurodevelopmental disability through 11 years of age in children born before 26 weeks of gestation. Pediatrics 2009, 124, e249–e257. [Google Scholar] [CrossRef]
- Adams-Chapman, I.; Heyne, R.J.; DeMauro, S.B.; Duncan, A.F.; Hintz, S.R.; Pappas, A.; Vohr, B.R.; McDonald, S.A.; Das, A.; Newman, J.E.; et al. Neurodevelopmental impairment among extremely preterm infants in the neonatal research network. Pediatrics 2018, 141, e20173091. [Google Scholar] [CrossRef]
- Leversen, K.T.; Sommerfelt, K.; Ronnestad, A.; Kaaresen, P.I.; Farstad, T.; Skranes, J.; Stoen, R.; Bircow Elgen, I.; Rettedal, S.; Egil Eide, G.; et al. Prediction of neurodevelopmental and sensory outcome at 5 years in Norwegian children born extremely preterm. Pediatrics 2011, 127, e630–e638. [Google Scholar] [CrossRef] [PubMed]
- Serenius, F.; Ewald, U.; Farooqi, A.; Fellman, V.; Hafstrom, M.; Hellgren, K.; Marsal, K.; Ohlin, A.; Olhager, E.; Stjernqvist, K.; et al. Neurodevelopmental outcomes among extremely preterm infants 6.5 years after active perinatal care in Sweden. JAMA Pediatr. 2016, 170, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, H.; Suenaga, H.; Uchiyama, A.; Kono, Y.; Kusuda, S.; Neonatal Research Network, Japan. Trends in the neurodevelopmental outcomes among preterm infants from 2003–2012: A retrospective cohort study in Japan. J. Perinatol. 2018, 38, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Chow, S.S.W.; Creighton, P.; Kander, V.; Haslam, R.R.; Lui, K. 2016 Report of the Australian and New Zealand Neonatal Network. Available online: https://anznn.net/annualreports (accessed on 31 July 2018).
- Pascal, A.; Govaert, P.; Oostra, A.; Naulaers, G.; Ortibus, E.; Van den Broeck, C. Neurodevelopmental outcome in very preterm and very-low-birthweight infants born over the past decade: A meta-analytic review. Dev. Med. Child Neurol. 2018, 60, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Himpens, E.; Van den Broeck, C.; Oostra, A.; Calders, P.; Vanhaesebrouck, P. Prevalence, type, distribution, and severity of cerebral palsy in relation to gestational age: A meta-analytic review. Dev. Med. Child Neurol. 2008, 50, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Synnes, A.R.; Anson, S.; Arkesteijn, A.; Butt, A.; Grunau, R.E.; Rogers, M.; Whitfield, M.F. School entry age outcomes for infants with birth weight ≤800 grams. J. Pediatr. 2010, 157, 989–994.e1. [Google Scholar] [CrossRef] [PubMed]
- Bhutta, A.T.; Cleves, M.A.; Casey, P.H.; Cradock, M.M.; Anand, K.J. Cognitive and behavioral outcomes of school-aged children who were born preterm: A meta-analysis. JAMA 2002, 288, 728–737. [Google Scholar] [CrossRef]
- Gonzalez, F.F.; Miller, S.P. Does perinatal asphyxia impair cognitive function without cerebral palsy? Arch. Dis. Child. Fetal Neonatal Ed. 2006, 91, F454–F459. [Google Scholar] [CrossRef]
- Marlow, N.; Wolke, D.; Bracewell, M.A.; Samara, M.; EPICure Study Group. Neurologic and developmental disability at six years of age after extremely preterm birth. N. Engl. J. Med. 2005, 352, 9–19. [Google Scholar] [CrossRef]
- Anderson, P.J.; Doyle, L.W. Executive functioning in school-aged children who were born very preterm or with extremely low birth weight in the 1990s. Pediatrics 2004, 114, 50–57. [Google Scholar] [CrossRef]
- Grunau, R.E.; Whitfield, M.F.; Davis, C. Pattern of learning disabilities in children with extremely low birth weight and broadly average intelligence. Arch. Pediatr. Adolesc. Med. 2002, 156, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.G.; Minich, N.M.; Klein, N.; Hack, M. Longitudinal outcomes of very low birth weight: Neuropsychological findings. J. Int. Neuropsychol. Soc. 2004, 10, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, E.B.; Edmonds, C.J.; Chong, W.K.; Lucas, A.; Morley, R.; Gadian, D.G. Brain morphometry and IQ measurements in preterm children. Brain 2004, 127, 2595–2607. [Google Scholar] [CrossRef] [PubMed]
- Grunau, R.E.; Whitfield, M.F.; Fay, T.B. Psychosocial and academic characteristics of extremely low birth weight (<or =800 g) adolescents who are free of major impairment compared with term-born control subjects. Pediatrics 2004, 114, e725–e732. [Google Scholar] [CrossRef] [PubMed]
- Hack, M.; Flannery, D.J.; Schluchter, M.; Cartar, L.; Borawski, E.; Klein, N. Outcomes in young adulthood for very-low-birth-weight infants. N. Engl. J. Med. 2002, 346, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Nosarti, C.; Giouroukou, E.; Micali, N.; Rifkin, L.; Morris, R.G.; Murray, R.M. Impaired executive functioning in young adults born very preterm. J. Int. Neuropsychol. Soc. 2007, 13, 571–581. [Google Scholar] [CrossRef]
- Hack, M.; Taylor, H.G.; Schluchter, M.; Andreias, L.; Drotar, D.; Klein, N. Behavioral outcomes of extremely low birth weight children at age 8 years. J. Dev. Behav. Pediatr. 2009, 30, 122–130. [Google Scholar] [CrossRef]
- Farooqi, A.; Hagglof, B.; Sedin, G.; Gothefors, L.; Serenius, F. Mental health and social competencies of 10- to 12-year-old children born at 23 to 25 weeks of gestation in the 1990s: A Swedish national prospective follow-up study. Pediatrics 2007, 120, 118–133. [Google Scholar] [CrossRef]
- Scott, M.N.; Taylor, H.G.; Fristad, M.A.; Klein, N.; Espy, K.A.; Minich, N.; Hack, M. Behavior disorders in extremely preterm/extremely low birth weight children in kindergarten. J. Dev. Behav. Pediatr. 2012, 33, 202–213. [Google Scholar] [CrossRef]
- Botting, N.; Powls, A.; Cooke, R.W.; Marlow, N. Attention deficit hyperactivity disorders and other psychiatric outcomes in very low birthweight children at 12 years. J. Child Psychol. Psychiatr. 1997, 38, 931–941. [Google Scholar]
- Nadeau, L.; Tessier, R.; Lefebvre, F.; Robaey, P. Victimization: A newly recognized outcome of prematurity. Dev. Med. Child Neurol. 2004, 46, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Pierrat, V.; Marchand-Martin, L.; Arnaud, C.; Kaminski, M.; Resche-Rigon, M.; Lebeaux, C.; Bodeau-Livinec, F.; Morgan, A.S.; Goffinet, F.; Marret, S.; et al. Neurodevelopmental outcome at 2 years for preterm children born at 22 to 34 weeks’ gestation in France in 2011: EPIPAGE-2 cohort study. BMJ 2017, 358, j3448. [Google Scholar] [CrossRef] [PubMed]
- Van Haastert, I.C.; Groenendaal, F.; Uiterwaal, C.S.; Termote, J.U.; van der Heide-Jalving, M.; Eijsermans, M.J.; Gorter, J.W.; Helders, P.J.; Jongmans, M.J.; de Vries, L.S. Decreasing incidence and severity of cerebral palsy in prematurely born children. J. Pediatr. 2011, 159, 86–91.e1. [Google Scholar] [CrossRef] [PubMed]
- Hamrick, S.E.; Miller, S.P.; Leonard, C.; Glidden, D.V.; Goldstein, R.; Ramaswamy, V.; Piecuch, R.; Ferriero, D.M. Trends in severe brain injury and neurodevelopmental outcome in premature newborn infants: The role of cystic periventricular leukomalacia. J. Pediatr. 2004, 145, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Soltirovska Salamon, A.; Groenendaal, F.; van Haastert, I.C.; Rademaker, K.J.; Benders, M.J.; Koopman, C.; de Vries, L.S. Neuroimaging and neurodevelopmental outcome of preterm infants with a periventricular haemorrhagic infarction located in the temporal or frontal lobe. Dev. Med. Child Neurol. 2014, 56, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Woodward, L.J.; Edgin, J.O.; Thompson, D.; Inder, T.E. Object working memory deficits predicted by early brain injury and development in the preterm infant. Brain 2005, 128, 2578–2587. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, T.; Diba, P.; Dean, J.M.; Banine, F.; Shaver, D.; Hagen, M.; Gong, X.; Su, W.; Emery, B.; Marks, D.L.; et al. A TLR/AKT/FoxO3 immune tolerance-like pathway disrupts the repair capacity of oligodendrocyte progenitors. J. Clin. Investig. 2018, 128, 2025–2041. [Google Scholar] [CrossRef] [PubMed]
- Ball, G.; Boardman, J.P.; Aljabar, P.; Pandit, A.; Arichi, T.; Merchant, N.; Rueckert, D.; Edwards, A.D.; Counsell, S.J. The influence of preterm birth on the developing thalamocortical connectome. Cortex 2013, 49, 1711–1721. [Google Scholar] [CrossRef]
- Guo, T.; Duerden, E.G.; Adams, E.; Chau, V.; Branson, H.M.; Chakravarty, M.M.; Poskitt, K.J.; Synnes, A.; Grunau, R.E.; Miller, S.P. Quantitative assessment of white matter injury in preterm neonates: Association with outcomes. Neurology 2017, 88, 614–622. [Google Scholar] [CrossRef]
- Chau, V.; Synnes, A.; Grunau, R.E.; Poskitt, K.J.; Brant, R.; Miller, S.P. Abnormal brain maturation in preterm neonates associated with adverse developmental outcomes. Neurology 2013, 81, 2082–2089. [Google Scholar] [CrossRef]
- Counsell, S.J.; Boardman, J.P. Differential brain growth in the infant born preterm: Current knowledge and future developments from brain imaging. Semin. Fetal Neonatal Med. 2005, 10, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Limperopoulos, C.; Benson, C.B.; Bassan, H.; Disalvo, D.N.; Kinnamon, D.D.; Moore, M.; Ringer, S.A.; Volpe, J.J.; du Plessis, A.J. Cerebellar hemorrhage in the preterm infant: Ultrasonographic findings and risk factors. Pediatrics 2005, 116, 717–724. [Google Scholar] [CrossRef]
- Srinivasan, L.; Allsop, J.; Counsell, S.J.; Boardman, J.P.; Edwards, A.D.; Rutherford, M. Smaller cerebellar volumes in very preterm infants at term-equivalent age are associated with the presence of supratentorial lesions. Am. J. Neuroradiol. 2006, 27, 573–579. [Google Scholar] [PubMed]
- Limperopoulos, C.; Bassan, H.; Gauvreau, K.; Robertson, R.L., Jr.; Sullivan, N.R.; Benson, C.B.; Avery, L.; Stewart, J.; Soul, J.S.; Ringer, S.A.; et al. Does cerebellar injury in premature infants contribute to the high prevalence of long-term cognitive, learning, and behavioral disability in survivors? Pediatrics 2007, 120, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Patra, K.; Wilson-Costello, D.; Taylor, H.G.; Mercuri-Minich, N.; Hack, M. Grades I-II intraventricular hemorrhage in extremely low birth weight infants: Effects on neurodevelopment. J. Pediatr. 2006, 149, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Franz, A.R.; Pohlandt, F.; Bode, H.; Mihatsch, W.A.; Sander, S.; Kron, M.; Steinmacher, J. Intrauterine, early neonatal, and postdischarge growth and neurodevelopmental outcome at 5.4 years in extremely preterm infants after intensive neonatal nutritional support. Pediatrics 2009, 123, e101–e109. [Google Scholar] [CrossRef] [PubMed]
- Ehrenkranz, R.A.; Younes, N.; Lemons, J.A.; Fanaroff, A.A.; Donovan, E.F.; Wright, L.L.; Katsikiotis, V.; Tyson, J.E.; Oh, W.; Shankaran, S.; et al. Longitudinal growth of hospitalized very low birth weight infants. Pediatrics 1999, 104, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Regev, R.H.; Arnon, S.; Litmanovitz, I.; Bauer-Rusek, S.; Boyko, V.; Lerner-Geva, L.; Reichman, B.; Israel Neonatal Network. Association between neonatal morbidities and head growth from birth until discharge in very-low-birthweight infants born preterm: A population-based study. Dev. Med. Child Neurol. 2016, 58, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Benavente-Fernandez, I.; Synnes, A.; Grunau, R.E.; Chau, V.; Ramraj, C.; Glass, T.; Cayam-Rand, D.; Siddiqi, A.; Miller, S.P. Association of socioeconomic status and brain Injury with neurodevelopmental outcomes of very preterm children. JAMA Netw. Open 2019, 2, e192914. [Google Scholar] [CrossRef]
- Weisglas-Kuperus, N.; Hille, E.T.; Duivenvoorden, H.J.; Finken, M.J.; Wit, J.M.; van Buuren, S.; van Goudoever, J.B.; Verloove-Vanhorick, S.P. Intelligence of very preterm or very low birthweight infants in young adulthood. Arch. Dis. Child. Fetal Neonatal Ed. 2009, 94, F196–F200. [Google Scholar] [CrossRef]
- Allen, M.C. Outcomes after brain injury in the preterm infant. In Acquired Brain Injury in the Fetus and Newborn; Shevel, M., Miller, S.P., Eds.; Mac Keith Press: London, UK, 2012; pp. 99–120. [Google Scholar]
- Schlapbach, L.J.; Aebischer, M.; Adams, M.; Natalucci, G.; Bonhoeffer, J.; Latzin, P.; Nelle, M.; Bucher, H.U.; Latal, B.; Swiss Neonatal Network Follow-Up Group. Impact of sepsis on neurodevelopmental outcome in a Swiss National Cohort of extremely premature infants. Pediatrics 2011, 128, e348–e357. [Google Scholar] [CrossRef] [PubMed]
- Rees, C.M.; Pierro, A.; Eaton, S. Neurodevelopmental outcomes of neonates with medically and surgically treated necrotizing enterocolitis. Arch. Dis. Child. Fetal Neonatal Ed. 2007, 92, F193–F198. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.K.; Doyle, L.W.; Anderson, P.J.; Bear, M.; Daley, A.J.; Hunt, R.W.; Inder, T.E. Adverse neurodevelopment in preterm infants with postnatal sepsis or necrotizing enterocolitis is mediated by white matter abnormalities on magnetic resonance imaging at term. J. Pediatr. 2008, 153, 170–175.e1. [Google Scholar] [CrossRef] [PubMed]
- Volpe, J.J.; Kinney, H.C.; Jensen, F.E.; Rosenberg, P.A. The developing oligodendrocyte: Key cellular target in brain injury in the premature infant. Int. J. Dev. Neurosci. 2011, 29, 423–440. [Google Scholar] [CrossRef] [PubMed]
- Hansen-Pupp, I.; Hellstrom-Westas, L.; Cilio, C.M.; Andersson, S.; Fellman, V.; Ley, D. Inflammation at birth and the insulin-like growth factor system in very preterm infants. Acta Paediatr. 2007, 96, 830–836. [Google Scholar] [CrossRef]
- Twilhaar, E.S.; Wade, R.M.; de Kieviet, J.F.; van Goudoever, J.B.; van Elburg, R.M.; Oosterlaan, J. Cognitive outcomes of children born extremely or very preterm since the 1990s and associated risk factors: A meta-analysis and meta-regression. JAMA Pediatr. 2018, 172, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef]
- Malikiwi, A.I.; Lee, Y.M.; Davies-Tuck, M.; Wong, F.Y. Postnatal nutritional deficit is an independent predictor of bronchopulmonary dysplasia among extremely premature infants born at or less than 28 weeks gestation. Early Hum. Dev. 2019, 131, 29–35. [Google Scholar] [CrossRef]
- Berrington, J.E.; Stewart, C.J.; Embleton, N.D.; Cummings, S.P. Gut microbiota in preterm infants: Assessment and relevance to health and disease. Arch. Dis. Child. Fetal Neonatal Ed. 2013, 98, F286–F290. [Google Scholar] [CrossRef]
- Ronnestad, A.; Abrahamsen, T.G.; Medbo, S.; Reigstad, H.; Lossius, K.; Kaaresen, P.I.; Egeland, T.; Engelund, I.E.; Irgens, L.M.; Markestad, T. Late-onset septicemia in a Norwegian national cohort of extremely premature infants receiving very early full human milk feeding. Pediatrics 2005, 115, e269–e276. [Google Scholar] [CrossRef]
- Shim, S.Y.; Ahn, H.M.; Cho, S.J.; Park, E.A. Early aggressive nutrition enhances language development in very low-birthweight infants. Pediatr. Int. 2014, 56, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Dit Trolli, S.E.; Kermorvant-Duchemin, E.; Huon, C.; Bremond-Gignac, D.; Lapillonne, A. Early lipid supply and neurological development at one year in very low birth weight (VLBW) preterm infants. Early Hum. Dev. 2012, 88 (Suppl. 1), S25–S29. [Google Scholar] [CrossRef]
- Beauport, L.; Schneider, J.; Faouzi, M.; Hagmann, P.; Huppi, P.S.; Tolsa, J.F.; Truttmann, A.C.; Fischer Fumeaux, C.J. Impact of early nutritional intake on preterm brain: A magnetic resonance imaging study. J. Pediatr. 2017, 181, 29–36.e1. [Google Scholar] [CrossRef] [PubMed]
- Belfort, M.B.; Ehrenkranz, R.A. Neurodevelopmental outcomes and nutritional strategies in very low birth weight infants. Semin. Fetal Neonatal Med. 2017, 22, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Leenders, E.; de Waard, M.; van Goudoever, J.B. Low- versus high-dose and early versus late parenteral amino-acid administration in very-low-birth-weight infants: A systematic review and meta-analysis. Neonatology 2018, 113, 187–205. [Google Scholar] [CrossRef] [PubMed]
- Lapillonne, A.; Kermorvant-Duchemin, E. A systematic review of practice surveys on parenteral nutrition for preterm infants. J. Nutr. 2013, 143, 2061S–2065S. [Google Scholar] [CrossRef] [PubMed]
- Vinall, J.; Grunau, R.E.; Brant, R.; Chau, V.; Poskitt, K.J.; Synnes, A.R.; Miller, S.P. Slower postnatal growth is associated with delayed cerebral cortical maturation in preterm newborns. Sci. Transl. Med. 2013, 5, 168ra8. [Google Scholar] [CrossRef] [PubMed]
- Latal-Hajnal, B.; von Siebenthal, K.; Kovari, H.; Bucher, H.U.; Largo, R.H. Postnatal growth in VLBW infants: Significant association with neurodevelopmental outcome. J. Pediatr. 2003, 143, 163–170. [Google Scholar] [CrossRef]
- Ramel, S.E.; Georgieff, M.K. Preterm nutrition and the Brain. In Nutritional Care of Preterm Infants Scientific Basis and Practical Guidelines; Karger: Basel, Switzerland, 2014; Volume 110, pp. 190–200. [Google Scholar]
- Fuglestad, A.; Rao, R.; Georgieff, M.K. The role of nutrition in cognitive development. In Handbook in Developmental Cognitive Neuroscience; MIT Press: Cambridge, MA, USA, 2008; Volume 2, pp. 623–641. [Google Scholar]
- Georgieff, M.K. Nutrition and the developing brain: Nutrient priorities and measurement. Am. J. Clin. Nutr. 2007, 85, 614S–620S. [Google Scholar] [CrossRef]
- Jha, M.K.; Morrison, B.M. Glia-neuron energy metabolism in health and diseases: New insights into the role of nervous system metabolic transporters. Exp. Neurol. 2018, 309, 23–31. [Google Scholar] [CrossRef]
- Wyss, M.T.; Jolivet, R.; Buck, A.; Magistretti, P.J.; Weber, B. In vivo evidence for lactate as a neuronal energy source. J. Neurosci. 2011, 31, 7477–7485. [Google Scholar] [CrossRef]
- Sobieski, C.; Warikoo, N.; Shu, H.J.; Mennerick, S. Ambient but not local lactate underlies neuronal tolerance to prolonged glucose deprivation. PLoS ONE 2018, 13, e0195520. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.L.; Weston, P.J.; Harding, J.E. Lactate, rather than ketones, may provide alternative cerebral fuel in hypoglycaemic newborns. Arch. Dis. Child. Fetal Neonatal Ed. 2015, 100, F161–F164. [Google Scholar] [CrossRef] [PubMed]
- Cusick, S.E.; Georgieff, M.K. The role of nutrition in brain development: The golden opportunity of the “first 1000 days”. J. Pediatr. 2016, 175, 16–21. [Google Scholar] [CrossRef] [PubMed]
- American Academy of Pediatrics, Committee on Nutrition. Nutritional needs of low-birth-weight infants. Pediatrics 1977, 60, 519–530. [Google Scholar]
- Cole, T.J.; Statnikov, Y.; Santhakumaran, S.; Pan, H.; Modi, N. Postnatal weight gain after very preterm birth: A UK population study. Arch. Dis. Child. 2011, 96, A3–A4. [Google Scholar] [CrossRef]
- Horbar, J.D.; Ehrenkranz, R.A.; Badger, G.J.; Edwards, E.M.; Morrow, K.A.; Soll, R.F.; Buzas, J.S.; Bertino, E.; Gagliardi, L.; Bellù, R. Weight growth velocity and postnatal growth failure in infants 501 to 1500 grams: 2000–2013. Pediatrics 2015, 136, e84–e92. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Viana, E.; Dworzynski, K.; Murphy, M.S.; Peek, R. Faltering growth in children: Summary of NICE guidance. BMJ 2017, 358, j4219. [Google Scholar] [CrossRef] [PubMed]
- Senterre, T.; Rigo, J. Reduction in postnatal cumulative nutritional deficit and improvement of growth in extremely preterm infants. Acta Paediatr. 2012, 101, e64–e70. [Google Scholar] [CrossRef] [PubMed]
- Cormack, B.E.; Bloomfield, F.H. Increased protein intake decreases postnatal growth faltering in ELBW babies. Arch. Dis. Child. Fetal Neonatal Ed. 2013, 98, F399–F404. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I.E.; Harris, C.L.; Lawson, M.L.; Berseth, C.L. Higher protein intake improves length, not weight, z scores in preterm infants. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Ehrenkranz, R.A.; Dusick, A.M.; Vohr, B.R.; Wright, L.L.; Wrage, L.A.; Poole, W.K. Growth in the neonatal intensive care unit influences neurodevelopmental and growth outcomes of extremely low birth weight infants. Pediatrics 2006, 117, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Belfort, M.B.; Rifas-Shiman, S.L.; Sullivan, T.; Collins, C.T.; McPhee, A.J.; Ryan, P.; Kleinman, K.P.; Gillman, M.W.; Gibson, R.A.; Makrides, M. Infant growth before and after term: Effects on neurodevelopment in preterm infants. Pediatrics 2011, 128, e899–e906. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.S.; Wong, K.Y.; Merko, S.; Bishara, R.; Dunn, M.; Asztalos, E.; Darling, P.B. Postnatal growth failure in preterm infants: Ascertainment and relation to long-term outcome. J. Perinat. Med. 2006, 34, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Zozaya, C.; Diaz, C.; Saenz de Pipaon, M. How should we define postnatal growth restriction in preterm infants? Neonatology 2018, 114, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Claas, M.J.; de Vries, L.S.; Koopman, C.; Uniken Venema, M.M.; Eijsermans, M.J.; Bruinse, H.W.; Stuart, A.A. Postnatal growth of preterm born children ≤750g at birth. Early Hum. Dev. 2011. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, E.B.; Morley, R.; Lucas, A. Early diet and general cognitive outcome at adolescence in children born at or below 30 weeks gestation. J. Pediatr. 2009, 155, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Sammallahti, S.; Pyhala, R.; Lahti, M.; Lahti, J.; Pesonen, A.K.; Heinonen, K.; Hovi, P.; Eriksson, J.G.; Strang-Karlsson, S.; Andersson, S.; et al. Infant growth after preterm birth and neurocognitive abilities in young adulthood. J. Pediatr. 2014, 165, 1109–1115.e3. [Google Scholar] [CrossRef]
- Ramel, S.E.; Demerath, E.W.; Gray, H.L.; Younge, N.; Boys, C.; Georgieff, M.K. The relationship of poor linear growth velocity with neonatal illness and two-year neurodevelopment in preterm infants. Neonatology 2012, 102, 19–24. [Google Scholar] [CrossRef]
- Simon, L.; Theveniaut, C.; Flamant, C.; Frondas-Chauty, A.; Darmaun, D.; Roze, J.C. In preterm infants, length growth below expected growth during hospital stay predicts poor neurodevelopment at 2 years. Neonatology 2018, 114, 135–141. [Google Scholar] [CrossRef]
- Scharf, R.J.; Stroustrup, A.; Conaway, M.R.; DeBoer, M.D. Growth and development in children born very low birthweight. Arch. Dis. Child. Fetal Neonatal Ed. 2016, 101, F433–F438. [Google Scholar] [CrossRef]
- Skullerud, K. Variations in the size of the human brain. Influence of age, sex, body length, body mass index, alcoholism, Alzheimer changes, and cerebral atherosclerosis. Acta Neurol. Scand. Suppl. 1985, 102, 1–94. [Google Scholar]
- Goldberg, D.L.; Becker, P.J.; Brigham, K.; Carlson, S.; Fleck, L.; Gollins, L.; Sandrock, M.; Fullmer, M.; Van Poots, H.A. Identifying malnutrition in preterm and neonatal populations: Recommended indicators. J. Acad. Nutr. Diet. 2018, 118, 1571–1582. [Google Scholar] [CrossRef] [PubMed]
- Sammallahti, S.; Heinonen, K.; Andersson, S.; Lahti, M.; Pirkola, S.; Lahti, J.; Pesonen, A.K.; Lano, A.; Wolke, D.; Eriksson, J.G.; et al. Growth after late-preterm birth and adult cognitive, academic, and mental health outcomes. Pediatr. Res. 2017, 81, 767–774. [Google Scholar] [CrossRef]
- Cheong, J.L.; Hunt, R.W.; Anderson, P.J.; Howard, K.; Thompson, D.K.; Wang, H.X.; Bear, M.J.; Inder, T.E.; Doyle, L.W. Head growth in preterm infants: Correlation with magnetic resonance imaging and neurodevelopmental outcome. Pediatrics 2008, 121, e1534–e1540. [Google Scholar] [CrossRef]
- Lind, A.; Parkkola, R.; Lehtonen, L.; Munck, P.; Maunu, J.; Lapinleimu, H.; Haataja, L.; PIPARI Study Group. Associations between regional brain volumes at term-equivalent age and development at 2 years of age in preterm children. Pediatr. Radiol. 2011, 41, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Van Kooij, B.J.; Benders, M.J.; Anbeek, P.; Van Haastert, I.C.; De Vries, L.S.; Groenendaal, F. Cerebellar volume and proton magnetic resonance spectroscopy at term, and neurodevelopment at 2 years of age in preterm infants. Dev. Med. Child Neurol. 2012, 54, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Hansen-Pupp, I.; Hovel, H.; Lofqvist, C.; Hellstrom-Westas, L.; Fellman, V.; Huppi, P.S.; Hellstrom, A.; Ley, D. Circulatory insulin-like growth factor-I and brain volumes in relation to neurodevelopmental outcome in very preterm infants. Pediatr. Res. 2013, 74, 564–569. [Google Scholar] [CrossRef]
- Lee, K.A.; Hayes, B.C. Head size and growth in the very preterm infant: A literature review. Res. Rep. Neonatol. 2015, 5, 1. [Google Scholar] [CrossRef]
- Kan, E.; Roberts, G.; Anderson, P.J.; Doyle, L.W.; Victorian Infant Collaborative Study Group. The association of growth impairment with neurodevelopmental outcome at eight years of age in very preterm children. Early Hum. Dev. 2008, 84, 409–416. [Google Scholar] [CrossRef]
- Leppanen, M.; Lapinleimu, H.; Lind, A.; Matomaki, J.; Lehtonen, L.; Haataja, L.; Rautava, P.; Group, P.S.; Aarimaa, T.; Ekblad, M.; et al. Antenatal and postnatal growth and 5-year cognitive outcome in very preterm infants. Pediatrics 2014, 133, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.; McGowan, P.; Herwitker, S.; Hart, A.E.; Turner, M.A. Postnatal head growth in preterm infants: A randomized controlled parenteral nutrition study. Pediatrics 2014, 133, e120–e128. [Google Scholar] [CrossRef] [PubMed]
- Stephens, B.E.; Walden, R.V.; Gargus, R.A.; Tucker, R.; McKinley, L.; Mance, M.; Nye, J.; Vohr, B.R. First-week protein and energy intakes are associated with 18-month developmental outcomes in extremely low birth weight infants. Pediatrics 2009, 123, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.J.; Cooke, R.W. Improving head growth in very preterm infants–a randomised controlled trial I: Neonatal outcomes. Arch. Dis. Child. Fetal Neonatal Ed. 2008, 93, F337–F341. [Google Scholar] [CrossRef] [PubMed]
- Hansen-Pupp, I.; Hovel, H.; Hellstrom, A.; Hellstrom-Westas, L.; Lofqvist, C.; Larsson, E.M.; Lazeyras, F.; Fellman, V.; Huppi, P.S.; Ley, D. Postnatal decrease in circulating insulin-like growth factor-I and low brain volumes in very preterm infants. J. Clin. Endocrinol. Metab. 2011, 96, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, E.B.; Gadian, D.G.; Sabatini, S.; Chong, W.K.; Quinn, B.T.; Fischl, B.R.; Lucas, A. The effect of early human diet on caudate volumes and IQ. Pediatr. Res. 2008, 63, 308–314. [Google Scholar] [CrossRef]
- Rijken, M.; Wit, J.M.; Le Cessie, S.; Veen, S.; Leiden Follow-Up Project on Prematurity. The effect of perinatal risk factors on growth in very preterm infants at 2 years of age: The Leiden Follow-Up Project on Prematurity. Early Hum. Dev. 2007, 83, 527–534. [Google Scholar] [CrossRef]
- Kuban, K.C.; Allred, E.N.; O’Shea, T.M.; Paneth, N.; Westra, S.; Miller, C.; Rosman, N.P.; Leviton, A. Developmental correlates of head circumference at birth and two years in a cohort of extremely low gestational age newborns. J. Pediatr. 2009, 155, 344–349.e3. [Google Scholar] [CrossRef]
- Rogers, C.E.; Smyser, T.; Smyser, C.D.; Shimony, J.; Inder, T.E.; Neil, J.J. Regional white matter development in very preterm infants: Perinatal predictors and early developmental outcomes. Pediatr. Res. 2016, 79, 87–95. [Google Scholar] [CrossRef]
- Taylor, D.G.; Bushell, M.C. The spatial mapping of translational diffusion coefficients by the NMR imaging technique. Phys. Med. Biol. 1985, 30, 345–349. [Google Scholar] [CrossRef]
- Thompson, D.K.; Lee, K.J.; Egan, G.F.; Warfield, S.K.; Doyle, L.W.; Anderson, P.J.; Inder, T.E. Regional white matter microstructure in very preterm infants: Predictors and 7 year outcomes. Cortex 2014, 52, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Tam, E.W.; Chau, V.; Barkovich, A.J.; Ferriero, D.M.; Miller, S.P.; Rogers, E.E.; Grunau, R.E.; Synnes, A.R.; Xu, D.; Foong, J.; et al. Early postnatal docosahexaenoic acid levels and improved preterm brain development. Pediatr. Res. 2016, 79, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Kamino, D.; Studholme, C.; Liu, M.; Chau, V.; Miller, S.P.; Synnes, A.; Rogers, E.E.; Barkovich, A.J.; Ferriero, D.M.; Brant, R.; et al. Postnatal polyunsaturated fatty acids associated with larger preterm brain tissue volumes and better outcomes. Pediatr. Res. 2018, 83, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Uauy, R.; Dangour, A.D. Nutrition in brain development and aging: Role of essential fatty acids. Nutr. Rev. 2006, 64, S24–S33. [Google Scholar] [CrossRef] [PubMed]
- Strommen, K.; Blakstad, E.W.; Moltu, S.J.; Almaas, A.N.; Westerberg, A.C.; Amlien, I.K.; Ronnestad, A.E.; Nakstad, B.; Drevon, C.A.; Bjornerud, A.; et al. Enhanced nutrient supply to very low birth weight infants is associated with improved white matter maturation and head growth. Neonatology 2015, 107, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Fischer Fumeaux, C.J.; Duerden, E.G.; Guo, T.; Foong, J.; Graz, M.B.; Hagmann, P.; Chakravarty, M.M.; Huppi, P.S.; Beauport, L.; et al. Nutrient intake in the first two weeks of life and brain growth in preterm neonates. Pediatrics 2018, 141, e20172169. [Google Scholar] [CrossRef] [PubMed]
- Vasu, V.; Durighel, G.; Thomas, L.; Malamateniou, C.; Bell, J.D.; Rutherford, M.A.; Modi, N. Preterm nutritional intake and MRI phenotype at term age: A prospective observational study. BMJ Open 2014, 4, e005390. [Google Scholar] [CrossRef]
- Coviello, C.; Keunen, K.; Kersbergen, K.J.; Groenendaal, F.; Leemans, A.; Peels, B.; Isgum, I.; Viergever, M.A.; de Vries, L.S.; Buonocore, G.; et al. Effects of early nutrition and growth on brain volumes, white matter microstructure, and neurodevelopmental outcome in preterm newborns. Pediatr. Res. 2017, 83, 102–110. [Google Scholar] [CrossRef]
- Joosten, K.; Embleton, N.; Yan, W.; Senterre, T.; ESPGHAN/ESPEN/ESPR/CSPEN Working Group on Pediatric Parenteral Nutrition. ESPGHAN/ESPEN/ESPR guidelines on pediatric parenteral nutrition: Energy. Clin. Nutr. 2018, 37, 2309–2314. [Google Scholar] [CrossRef]
- Van Goudoever, J.B.; Carnielli, V.; Darmaun, D.; Sainz de Pipaon, M. ESPGHAN/ESPEN/ESPR/]CSPEN Working Group on Pediatric Parenteral Nutrition. Guidelines on Pediatric Parenteral Nutrition: Amino acids. Clin. Nutr. 2018, 37, 2315–2323. [Google Scholar] [CrossRef]
- McCullough, D.L. Visual Evoked Potentials in Infants. In Infant EEG and Event-Related Potentials; de Haan, M., Ed.; Psychology Press: New York, NY, USA, 2013; pp. 39–64. [Google Scholar]
- Pfister, K.M.; Gray, H.L.; Miller, N.C.; Demerath, E.W.; Georgieff, M.K.; Ramel, S.E. Exploratory study of the relationship of fat-free mass to speed of brain processing in preterm infants. Pediatr. Res. 2013, 74, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Abernethy, L.; Cooke, R. Improving head growth in preterm infants--a randomised controlled trial II: MRI and developmental outcomes in the first year. Arch. Dis. Child. Fetal Neonatal Ed. 2008, 93, F342–F346. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.K.; Kennedy, K.; Castaneda-Gutierrez, E.; Forsyth, S.; Godfrey, K.M.; Koletzko, B.; Latulippe, M.E.; Ozanne, S.E.; Rueda, R.; Schoemaker, M.H.; et al. Postnatal growth in preterm infants and later health outcomes: A systematic review. Acta Paediatr. 2015, 104, 974–986. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Goulet, O.; Hunt, J.; Krohn, K.; Shamir, R.; Parenteral Nutrition Guidelines Working Group. Guidelines on paediatric parenteral nutrition of the European Society of Paediatric Gastroenterology, Hepatology and Nutrition (ESPGHAN) and the European Society for Clinical Nutrition and Metabolism (ESPEN), supported by the European Society of Paediatric Research (ESPR). J. Pediatr. Gastroenterol. Nutr. 2005, 41 (Suppl. 2), S1–S87. [Google Scholar] [CrossRef] [PubMed]
- Duffy, B.; Gunn, T.; Collinge, J.; Pencharz, P. The effect of varying protein quality and energy intake on the nitrogen metabolism of parenterally fed very low birthweight (less than 1600 g) infants. Pediatr. Res. 1981, 15, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- Pencharz, P.B.; House, J.D.; Wykes, L.J.; Ball, R.O. What are the essential amino acids for the preterm and term infant? In Recent Developments in Infant Nutrition; Bindels, J.G., Goedhart, A.C., Visser, H.K.A., Eds.; Springer: Dordrecht, The Netherlands, 1996; Volume 9, pp. 278–296. [Google Scholar]
- Morgan, C.; Burgess, L. High protein intake does not prevent low plasma levels of conditionally essential amino acids in very preterm infants receiving parenteral nutrition. J. Parenter. Enteral Nutr. 2017, 41, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Rassin, D.K. Essential and non-essential amino acids in neonatal nutrition. In Protein Metabolism During Infancy; Raiha, N.C.R., Ed.; Raven Press Ltd.: New York, NY, USA, 1994; pp. 183–195. [Google Scholar]
- Hay, N.; Sonenberg, N. Upstream and downstream of mTOR. Genes Dev. 2004, 18, 1926–1945. [Google Scholar] [CrossRef]
- Lipton, J.O.; Sahin, M. The neurology of mTOR. Neuron 2014, 84, 275–291. [Google Scholar] [CrossRef]
- Tokunaga, C.; Yoshino, K.; Yonezawa, K. mTOR integrates amino acid- and energy-sensing pathways. Biochem. Biophys. Res. Commun. 2004, 313, 443–446. [Google Scholar] [CrossRef]
- Kennedy, B.K.; Lamming, D.W. The mechanistic target of rapamycin: The grand conductor of metabolism and aging. Cell Metab. 2016, 23, 990–1003. [Google Scholar] [CrossRef]
- Wang, S.; Tsun, Z.Y.; Wolfson, R.L.; Shen, K.; Wyant, G.A.; Plovanich, M.E.; Yuan, E.D.; Jones, T.D.; Chantranupong, L.; Comb, W.; et al. Metabolism. Lysosomal amino acid transporter SLC38A9 signals arginine sufficiency to mTORC1. Science 2015, 347, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Embleton, N.D.; Cooke, R.J. Protein requirements in preterm infants: Effect of different levels of protein intake on growth and body composition. Pediatr. Res. 2005, 58, 855–860. [Google Scholar] [CrossRef] [PubMed]
- Thureen, P.; Heird, W.C. Protein and energy requirements of the preterm/low birthweight (LBW) infant. Pediatr. Res. 2005, 57, 95R–98R. [Google Scholar] [CrossRef] [PubMed]
- Van den Akker, C.H.; te Braake, F.W.; Schierbeek, H.; Rietveld, T.; Wattimena, D.J.; Bunt, J.E.; van Goudoever, J.B.; van den Akker, C.H.P.; te Braake, F.W.J.; Schierbeek, H.; et al. Albumin synthesis in premature neonates is stimulated by parenterally administered amino acids during the first days of life. Am. J. Clin. Nutr. 2007, 86, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Te Braake, F.W.; van den Akker, C.H.; Wattimena, D.J.; Huijmans, J.G.; van Goudoever, J.B. Amino acid administration to premature infants directly after birth. J. Pediatr. 2005, 147, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.M.; Jeroudi, M.A.; Baier, R.J.; Dhanireddy, R.; Krouskop, R.W. Aggressive early total parental nutrition in low-birth-weight infants. J. Perinatol. 2004, 24, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Vlaardingerbroek, H.; Vermeulen, M.J.; Rook, D.; van den Akker, C.H.; Dorst, K.; Wattimena, J.L.; Vermes, A.; Schierbeek, H.; van Goudoever, J.B. Safety and efficacy of early parenteral lipid and high-dose amino acid administration to very low birth weight infants. J. Pediatr. 2013, 163, 638–644. [Google Scholar] [CrossRef]
- Vucetic, Z.; Totoki, K.; Schoch, H.; Whitaker, K.W.; Hill-Smith, T.; Lucki, I.; Reyes, T.M. Early life protein restriction alters dopamine circuitry. Neuroscience 2010, 168, 359–370. [Google Scholar] [CrossRef]
- Chertoff, M. Protein malnutrition and brain development. Brain Disord. Ther. 2014. [Google Scholar] [CrossRef]
- Sanches, E.F.; Arteni, N.S.; Spindler, C.; Moyses, F.; Siqueira, I.R.; Perry, M.L.; Netto, C.A. Effects of pre- and postnatal protein malnutrition in hypoxic-ischemic rats. Brain Res. 2012, 1438, 85–92. [Google Scholar] [CrossRef]
- Osborn, D.A.; Schindler, T.; Jones, L.J.; Sinn, J.K.; Bolisetty, S. Higher versus lower amino acid intake in parenteral nutrition for newborn infants. Cochrane Database Syst. Rev. 2018, 3, CD005949. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, M.; Jennings, A.; Przystac, L.; Phornphutkul, C.; Tucker, R.; Vohr, B.; Stephens, B.E.; Bliss, J.M. Growth and neurodevelopmental outcomes of early, high-dose parenteral amino acid intake in very low birth weight infants: A randomized controlled trial. J. Parenter. Enteral Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.L.; Gong, A.K.; Schoolfield, J.; Green, B.K.; Daniels, W.; Liechty, E.A.; Ramamurthy, R. Impact of early and high amino acid supplementation on ELBW infants at 2 years. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Burattini, I.; Bellagamba, M.P.; Spagnoli, C.; D’Ascenzo, R.; Mazzoni, N.; Peretti, A.; Cogo, P.E.; Carnielli, V.P.; Marche Neonatal, N. Targeting 2.5 versus 4 g/kg/day of amino acids for extremely low birth weight infants: A randomized clinical trial. J. Pediatr. 2013, 163, 1278–1282.e1. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Parry, S.; Morgan, C. Neurodevelopmental outcome in very preterm infants randomised to receive two different parenteral nutrition regimens: The SCAMP nutrition study. Arch. Dis. Child. 2016, 101:A5, P8. [Google Scholar]
- Roelants, J.A.; Vlaardingerbroek, H.; van den Akker, C.H.P.; de Jonge, R.C.J.; van Goudoever, J.B.; Vermeulen, M.J. Two-year follow-up of a randomized controlled nutrition intervention trial in very low-birth-weight infants. J. Parenter. Enteral Nutr. 2018, 42, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Uthaya, S.; Liu, X.; Babalis, D.; Dore, C.J.; Warwick, J.; Bell, J.; Thomas, L.; Ashby, D.; Durighel, G.; Ederies, A.; et al. Nutritional evaluation and optimisation in neonates: A randomized, double-blind controlled trial of amino acid regimen and intravenous lipid composition in preterm parenteral nutrition. Am. J. Clin. Nutr. 2016, 103, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Bellagamba, M.P.; Carmenati, E.; D’Ascenzo, R.; Malatesta, M.; Spagnoli, C.; Biagetti, C.; Burattini, I.; Carnielli, V.P. One extra gram of protein to preterm infants from birth to 1800 g: A single-blinded randomized clinical trial. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 879–884. [Google Scholar] [CrossRef]
- Blanco, C.L.; Falck, A.; Green, B.K.; Cornell, J.E.; Gong, A.K. Metabolic responses to early and high protein supplementation in a randomized trial evaluating the prevention of hyperkalemia in extremely low birth weight infants. J. Pediatr. 2008, 153, 535–540. [Google Scholar] [CrossRef]
- Buddhavarapu, S.; Manickaraj, S.; Lodha, A.; Soraisham, A.S. Does high protein intake during first week of life improve growth and neurodevelopmental outcome at 18 months corrected age in extremely preterm infants? Indian J. Pediatr. 2016, 83, 915–921. [Google Scholar] [CrossRef]
- Cester, E.A.; Bloomfield, F.H.; Taylor, J.; Smith, S.; Cormack, B.E. Do recommended protein intakes improve neurodevelopment in extremely preterm babies? Arch. Dis. Child. Fetal Neonatal Ed. 2015, 100, F243–F247. [Google Scholar] [CrossRef] [PubMed]
- Biasini, A.; Monti, F.; Laguardia, M.C.; Stella, M.; Marvulli, L.; Neri, E. High protein intake in human/maternal milk fortification for ≤1250 gr infants: Intrahospital growth and neurodevelopmental outcome at two years. Acta Biomed. 2018, 88, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Christmann, V.; Roeleveld, N.; Visser, R.; Janssen, A.J.; Reuser, J.J.; van Goudoever, J.B.; van Heijst, A.F. The early postnatal nutritional intake of preterm infants affected neurodevelopmental outcomes differently in boys and girls at 24 months. Acta Paediatr. 2017, 106, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Tottman, A.C.; Alsweiler, J.M.; Bloomfield, F.H.; Gamble, G.D.; Jiang, Y.; Leung, M.; Poppe, T.; Thompson, B.; Wouldes, T.A.; Harding, J.E.; et al. Relationships between early neonatal nutrition and neurodevelopment at school age in children born very preterm. J. Paediatr. Gastroenterol. Nutr. 2019. [Google Scholar] [CrossRef]
- Tottman, A.C. Long-Term Neurodevelopmental and Metabolic Consequences of Neonatal Hyperglycaemia and Early Life Protein Intake. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2018. [Google Scholar]
- Lapillonne, A.; Carnielli, V.P.; Embleton, N.D.; Mihatsch, W. Quality of newborn care: Adherence to guidelines for parenteral nutrition in preterm infants in four European countries. BMJ Open 2013, 3, e003478. [Google Scholar] [CrossRef] [PubMed]
- Hay, W.W., Jr. Nutritional support strategies for the preterm infant in the neonatal intensive care unit. Pediatr. Gastroenterol. Hepatol. Nutr. 2018, 21, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Jaeger, L.A.; Bazer, F.W.; Rhoads, J.M. Arginine deficiency in preterm infants: Biochemical mechanisms and nutritional implications. J. Nutr. Biochem. 2004, 15, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Choudhri, A.F.; Sable, H.J.; Chizhikov, V.V.; Buddington, K.K.; Buddington, R.K. Parenteral nutrition compromises neurodevelopment of preterm pigs. J. Nutr. 2014, 144, 1920–1927. [Google Scholar] [CrossRef] [PubMed]
- Lucas, A.; Baker, B.A.; Morley, R.M. Hyperphenylalaninaemia and outcome in intravenously fed preterm neonates. Arch. Dis. Child. 1993, 68, 579–583. [Google Scholar] [CrossRef]
- Ross, J.R.; Finch, C.; Ebeling, M.; Taylor, S.N. Refeeding syndrome in very-low-birth-weight intrauterine growth-restricted neonates. J. Perinatol. 2013, 33, 717–720. [Google Scholar] [CrossRef]
- Mizumoto, H.; Mikami, M.; Oda, H.; Hata, D. Refeeding syndrome in a small-for-dates micro-preemie receiving early parenteral nutrition. Pediatr. Int. 2012, 54, 715–717. [Google Scholar] [CrossRef] [PubMed]
- Boubred, F.; Herlenius, E.; Bartocci, M.; Jonsson, B.; Vanpee, M. Extremely preterm infants who are small for gestational age have a high risk of early hypophosphatemia and hypokalemia. Acta Paediatr. 2015, 104, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, A.; Okuno, T.; Ohta, G.; Tokuriki, S.; Ohshima, Y. Risk factors for the development of refeeding syndrome-like hypophosphatemia in very low birth weight infants. Dis. Markers 2017, 2017, 9748031. [Google Scholar] [CrossRef] [PubMed]
- Peacock, J.L.; Marston, L.; Marlow, N.; Calvert, S.A.; Greenough, A. Neonatal and infant outcome in boys and girls born very prematurely. Pediatr. Res. 2012, 71, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Moltu, S.J.; Strommen, K.; Blakstad, E.W.; Almaas, A.N.; Westerberg, A.C.; Braekke, K.; Ronnestad, A.; Nakstad, B.; Berg, J.P.; Veierod, M.B.; et al. Enhanced feeding in very-low-birth-weight infants may cause electrolyte disturbances and septicemia–a randomized, controlled trial. Clin. Nutr. 2013, 32, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, H.J.; Strommen, K.; Lang, A.M.; Abrahamsen, T.G.; Steen, E.K.; Pripp, A.H.; Ronnestad, A.E. Early enhanced parenteral nutrition, hyperglycemia, and death among extremely low-birth-weight infants. JAMA Pediatr. 2015, 169, 1003–1010. [Google Scholar] [CrossRef]
- Kent, A.L.; Wright, I.M.; Abdel-Latif, M.E.; the New South Wales and Australian Capital Territory Neonatal Intensive Care Units Audit Group. Mortality and adverse neurologic outcomes are greater in preterm male infants. Pediatrics 2012, 129, 124–131. [Google Scholar] [CrossRef]
- Stevenson, D.K.; Verter, J.; Fanaroff, A.A.; Oh, W.; Ehrenkranz, R.A.; Shankaran, S.; Donovan, E.F.; Wright, L.L.; Lemons, J.A.; Tyson, J.E.; et al. Sex differences in outcomes of very low birthweight infants: The newborn male disadvantage. Arch. Dis. Child. Fetal Neonatal Ed. 2000, 83, F182–F185. [Google Scholar] [CrossRef]
- Linsell, L.; Malouf, R.; Morris, J.; Kurinczuk, J.J.; Marlow, N. Prognostic factors for poor cognitive development in children born very preterm or with very low birth weight: A systematic review. JAMA Pediatr. 2015, 169, 1162–1172. [Google Scholar] [CrossRef]
- Reiss, A.L.; Kesler, S.R.; Vohr, B.; Duncan, C.C.; Katz, K.H.; Pajot, S.; Schneider, K.C.; Makuch, R.W.; Ment, L.R. Sex differences in cerebral volumes of 8-year-olds born preterm. J. Pediatr. 2004, 145, 242–249. [Google Scholar] [CrossRef]
- Tottman, A.C.; Bloomfield, F.H.; Cormack, B.E.; Harding, J.E.; Taylor, J.; Alsweiler, J.M. Associations between early nutritional intakes, growth and neurodevelopment are different in boys and girls born preterm. J. Paediatr. Child Health 2018, 54, 177. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| First Author Participants Country | n | Protein Intake Control Group g·kg−1·day−1 * | Protein Intake Intervention Group g·kg−1·day−1 * | Amino Acid Solution | Primary Outcome | Outcomes, Conclusions and Other Details |
|---|---|---|---|---|---|---|
| Tan <29 weeks UK [118,137] | 114 | ~2.3 | 2.6 | Not specified | Head growth at 36 weeks | No difference in total brain volume on MRI at 40 weeks. Follow up rate 64% at 9 months. MDI and PDI correlated significantly with energy deficit at 28 days of age. Bayley II assessor not blinded to group allocation. |
| Blanco <1000 g USA [160] | 32 | 2.9 | 3.7 (day 7) | Aminosyn | Mean serum potassium during first 3 days | Follow up rate 63% at 2 years. Bayley II PDI scores similar, but lower MDI score at 18 months in the high AA group. No difference at 24 months. No differences in cerebral palsy, blindness or early childhood intervention between groups at any age. More babies ≤24 weeks’ gestation allocated to higher protein group. |
| Burattini <1250 g Italy [161] | 114 | ~2.5 | ~3.0 | TrophAmine | Weight and weight <10th centile at 36 weeks | Follow up rate 87% at 2 years. No difference in Bayley III scores. Caregivers aware of group allocation. |
| Morgan (SCAMP) <1200 g or <29 weeksUK [116,162] | 150 | 2.4 week 1 3.0 week 2 | 2.8 3.6 | Vaminolact | Change in HC at 28 days postnatal age and 36 weeks | Follow up rate 62%. No difference in Bayley III assessments at 2 to 3.5 years. High protein group had higher HC z-score at 36 weeks (difference 0.40 95% CI 0.005–0.79, p = 0.047). Parents and assessors blinded to group allocation. |
| Vlaardingerbroek <1500 g Netherlands [154,163] | 103 | 2.4 + 1.4 g lipid on day 2 or 2.4 + 2–3 g lipid from birth | 3.6 + 2–3 g lipid from birth | Primène | Composite outcome of death or major disability at 2 years | Follow-up rate 92%. No difference in Bayley III assessments and incidence of major disability between groups at 2 years. At 6 weeks, weight and HC z-scores significantly higher in high protein group. Addition of lipids and extra protein increased the amount of phenylalanine used for protein synthesis. |
| Uthaya (NEON) <31 weeks UK [164] | 133 | 1.7 to 2.7 | 3.6 from day 1 | Vaminolact | Lean body mass | Follow up rate 79%. No significant differences for brain volume. HC at term-equivalent age was smaller in the high protein group (mean difference –0.8 cm, 95% CI–1.5 to–0.1 cm; p = 0.02), but also smaller at baseline. |
| Bellagamba 500–1249 g Italy [165] | 160 | ~2.5 | ~3.5 | TrophAmine | Weight gain from birth to 1800 g | Follow-up rate >90%. No differences in Bayley III scores at 2 years. Neonatologists not blinded to group allocation. IVN + enteral intervention. Difference in protein intakes between groups reached 1 g·kg−1·day−1 on day 4 |
| Balakrishnan <1250 g USA [159] | 168 | day 1–2 day 2–2.5 day 3–3 | 3 3.8 4 | Premasol | Bayley III at 18–24 months CGA | Follow-up rate 78%. No differences in Bayley III scores at 18–24 months. Largest study to date, single centre. Achieved at least 1 g·kg−1·day−1 difference in protein intakes between groups but for only first 3 days |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cormack, B.E.; Harding, J.E.; Miller, S.P.; Bloomfield, F.H. The Influence of Early Nutrition on Brain Growth and Neurodevelopment in Extremely Preterm Babies: A Narrative Review. Nutrients 2019, 11, 2029. https://doi.org/10.3390/nu11092029
Cormack BE, Harding JE, Miller SP, Bloomfield FH. The Influence of Early Nutrition on Brain Growth and Neurodevelopment in Extremely Preterm Babies: A Narrative Review. Nutrients. 2019; 11(9):2029. https://doi.org/10.3390/nu11092029
Chicago/Turabian StyleCormack, Barbara E., Jane E. Harding, Steven P. Miller, and Frank H. Bloomfield. 2019. "The Influence of Early Nutrition on Brain Growth and Neurodevelopment in Extremely Preterm Babies: A Narrative Review" Nutrients 11, no. 9: 2029. https://doi.org/10.3390/nu11092029
APA StyleCormack, B. E., Harding, J. E., Miller, S. P., & Bloomfield, F. H. (2019). The Influence of Early Nutrition on Brain Growth and Neurodevelopment in Extremely Preterm Babies: A Narrative Review. Nutrients, 11(9), 2029. https://doi.org/10.3390/nu11092029