Glutathione Transferase P1-1 an Enzyme Useful in Biomedicine and as Biomarker in Clinical Practice and in Environmental Pollution

 , , and
, , and 
Abstract
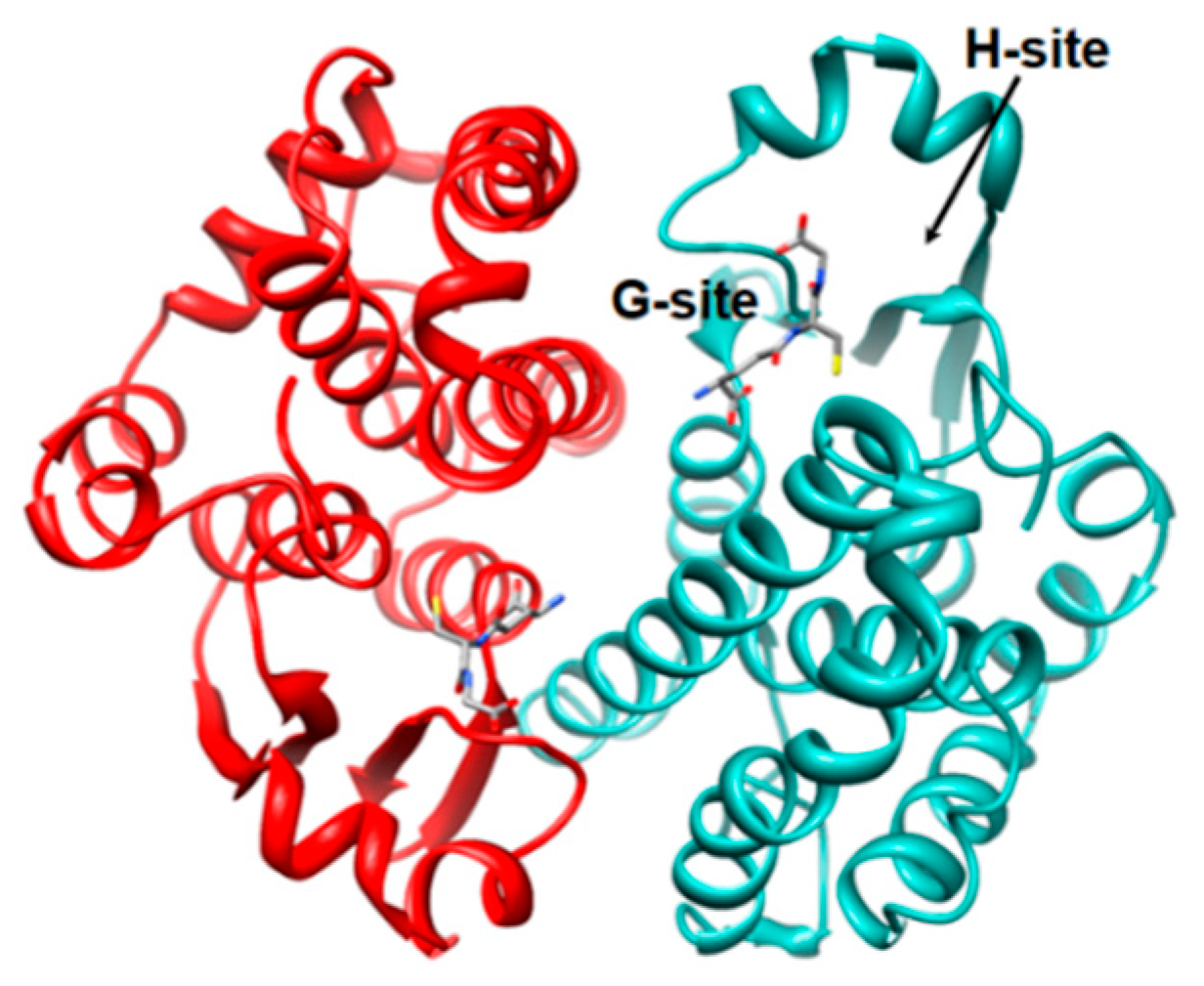
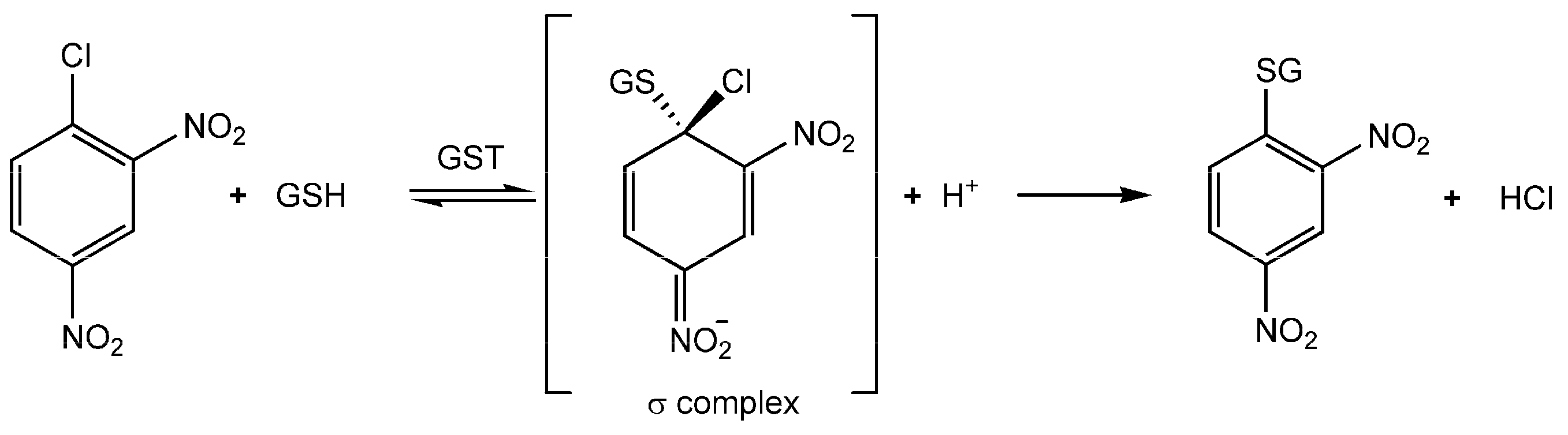
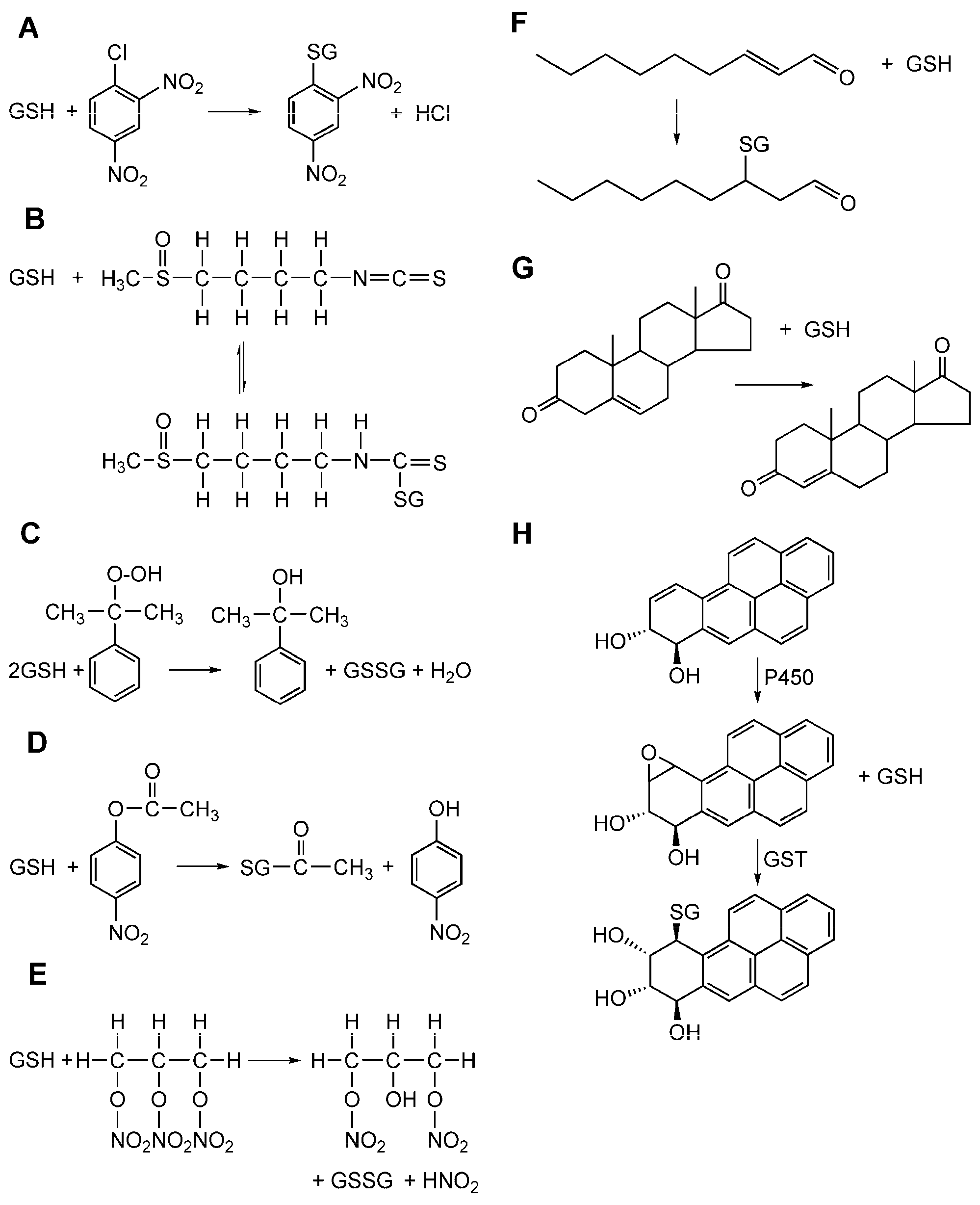
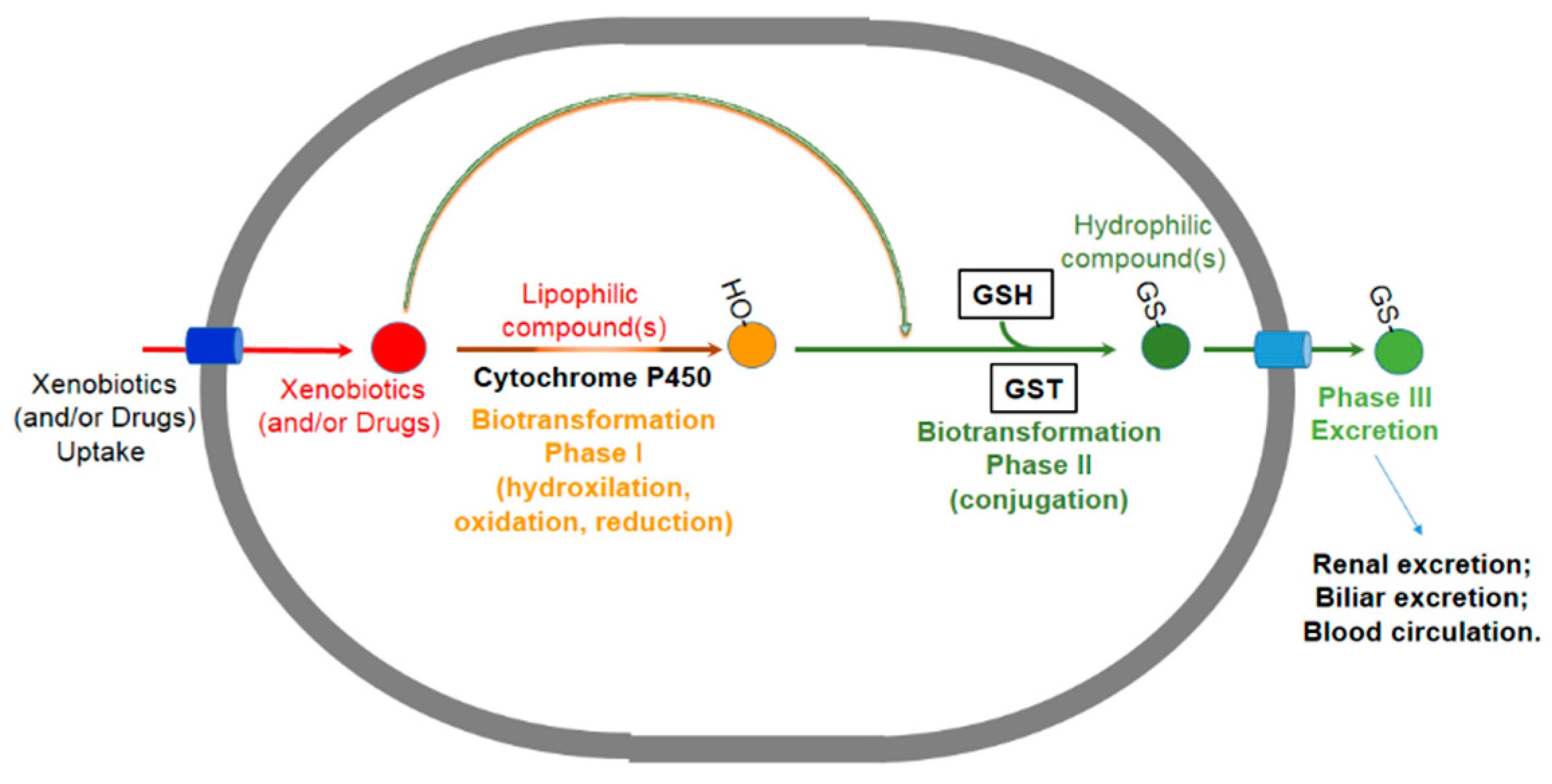
1. Introduction
1.1. The Erythrocyte GSTP1-1 (e-GST)
1.2. GSTP1-1 in Blood
2. Methods
3. Usefulness of GSTP1-1 Enzymatic Activity in Some Pathological Conditions
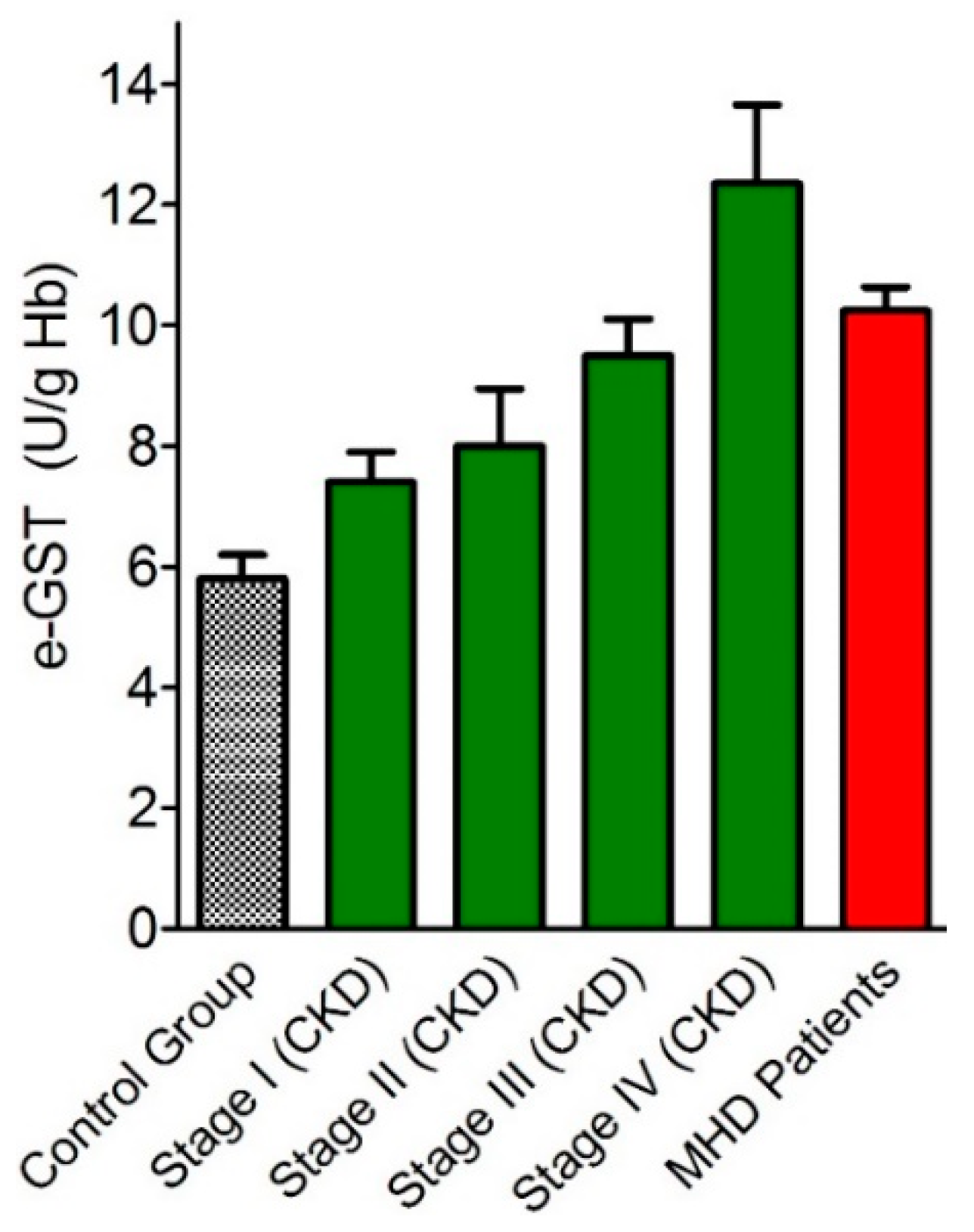
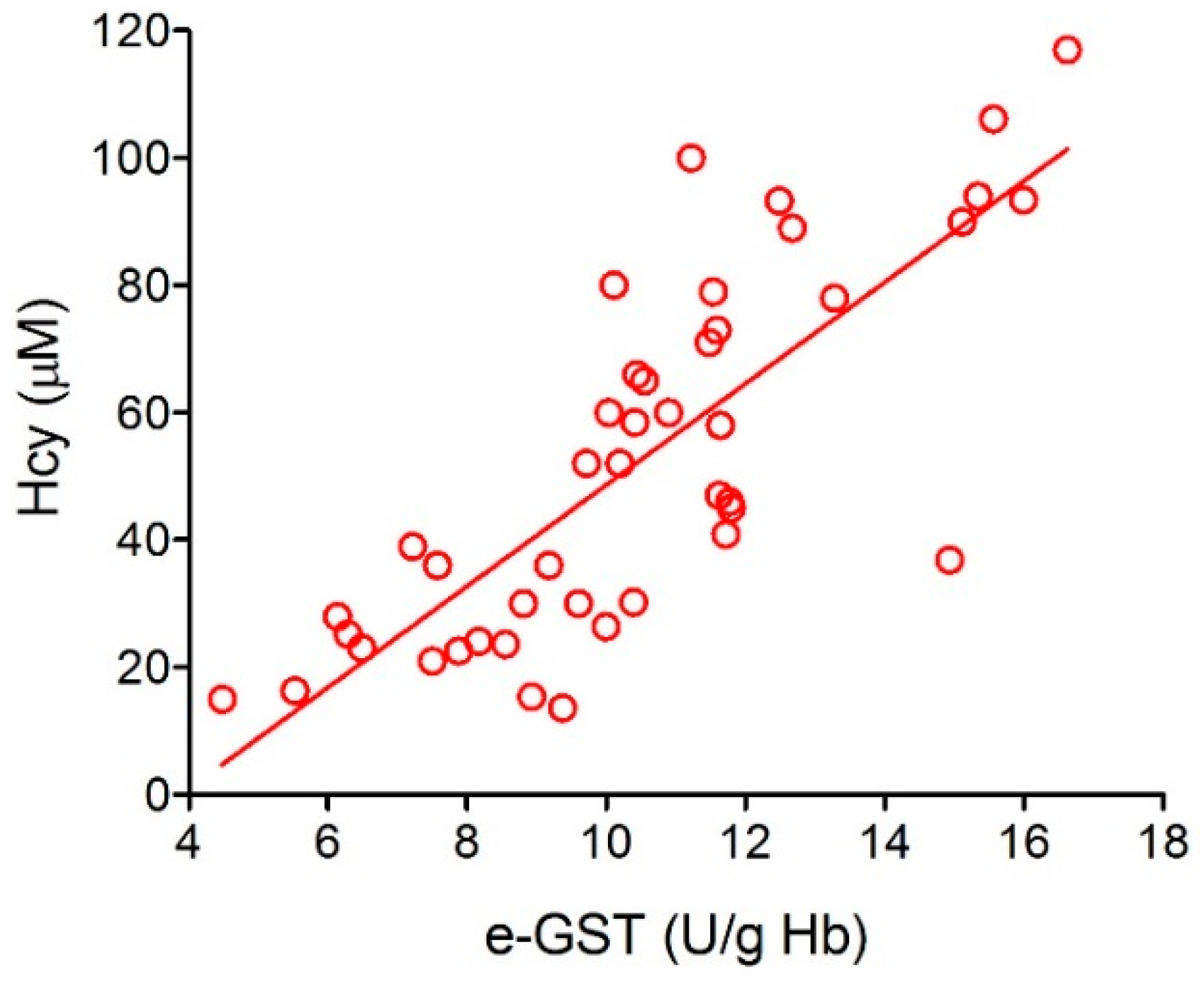
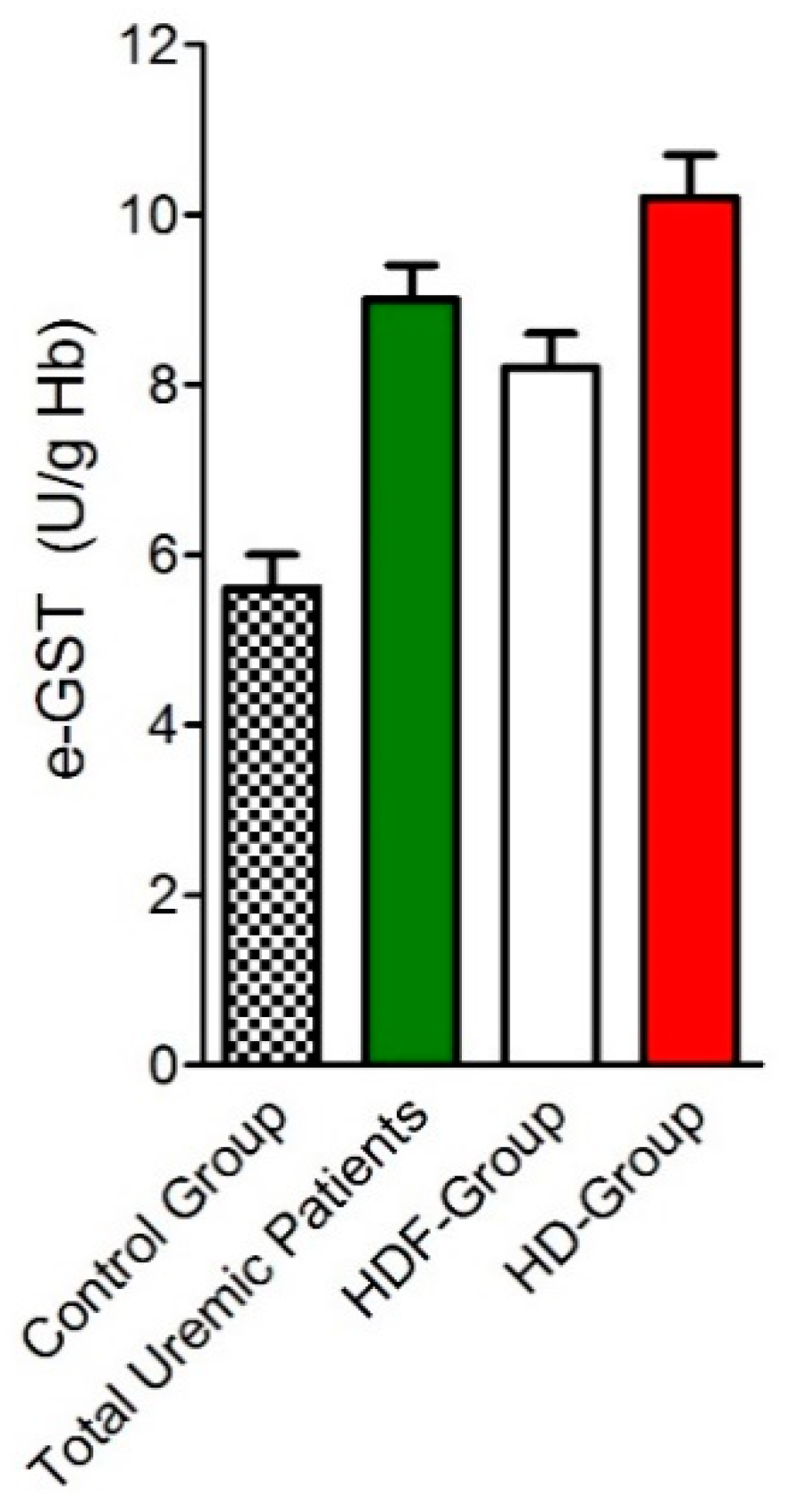
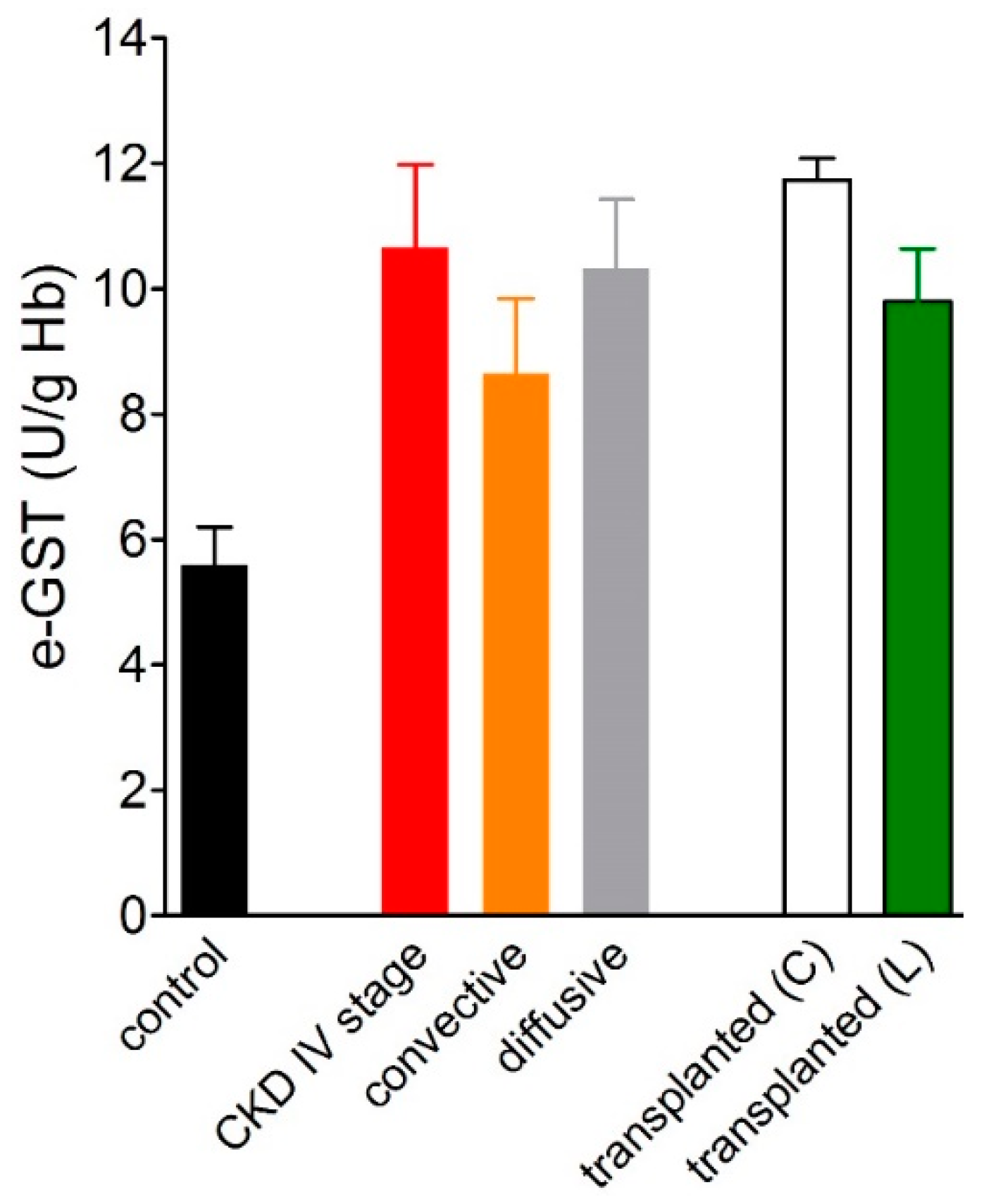
3.1. Over-Expression of e-GST in Chronic Kidney Disease
3.2. Overexpression of e-GST in Kidney Transplanted Patients
3.3. GSTP1-1 in Neurodegenerative Diseases and Psychiatric Disorders
3.4. e-GST Activity and Scleroderma
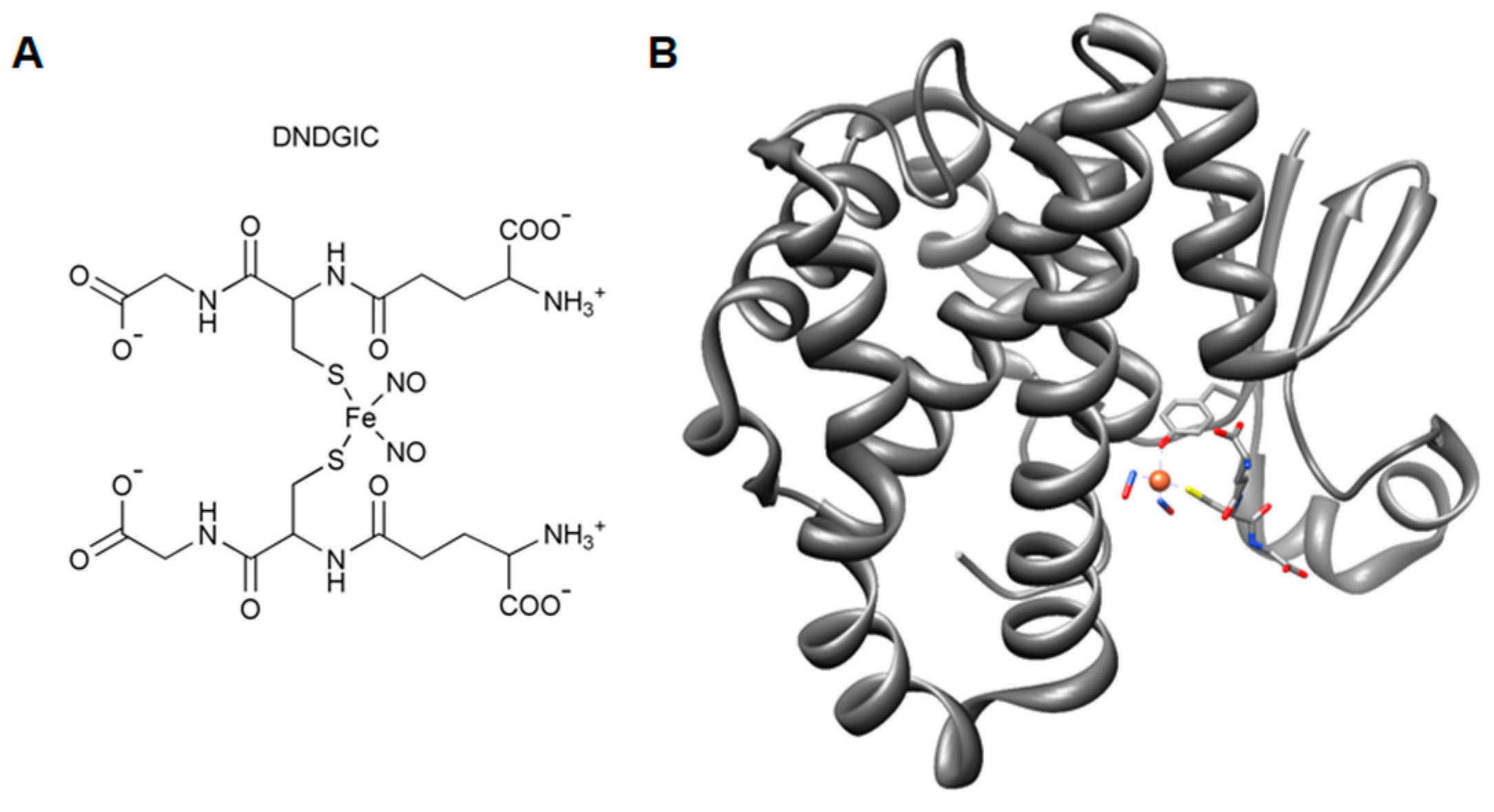
3.5. Role of e-GST in Oxidative Stress
3.6. GSTP1-1 in Cancer
3.7. GSTP1-1 and Liver Disease
4. Environmental and Endogenous Factors Affecting GST Levels in Healthy Subjects
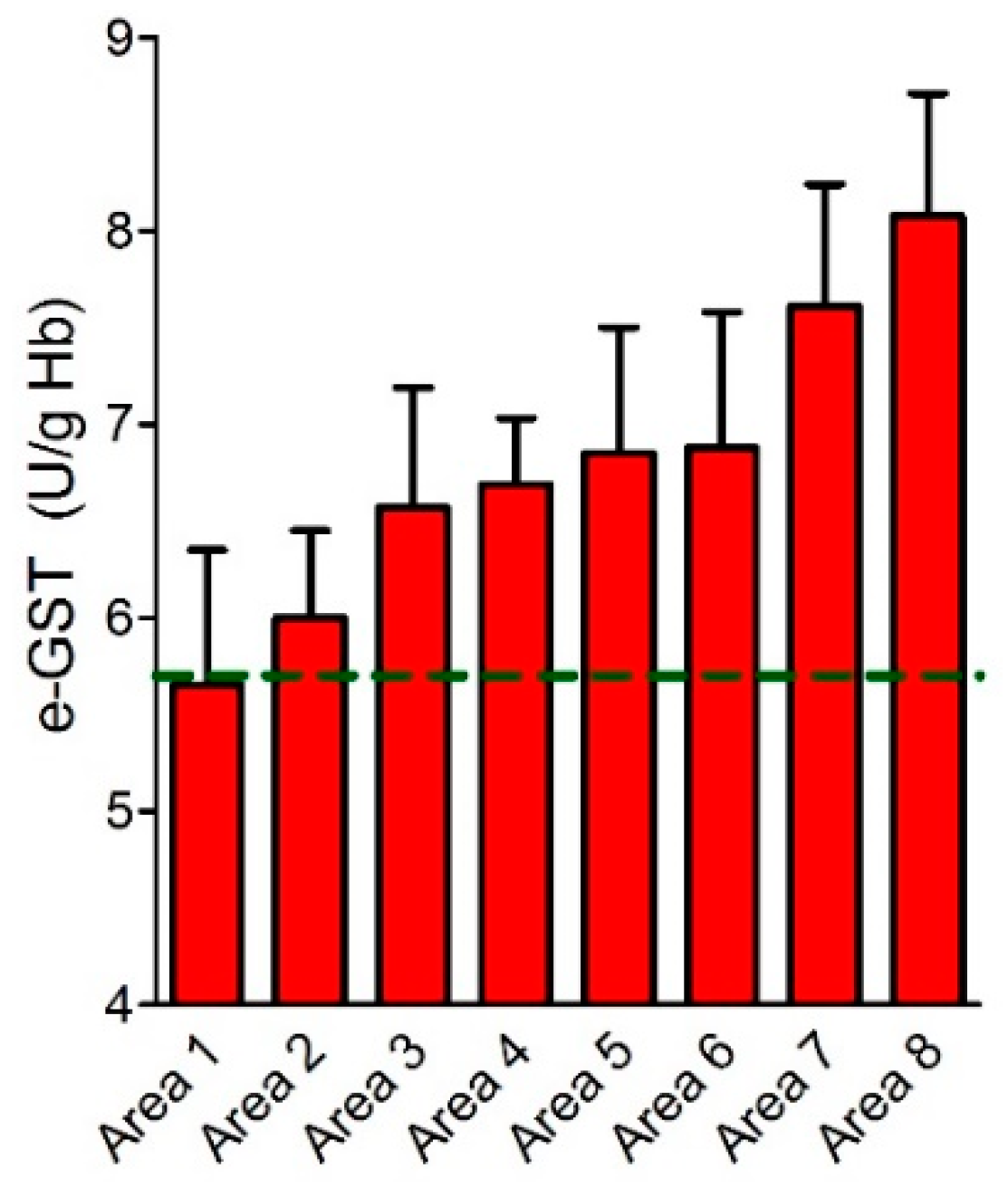
e-GST in Acute and Chronic Exposition to Contaminants: A Brief Comment
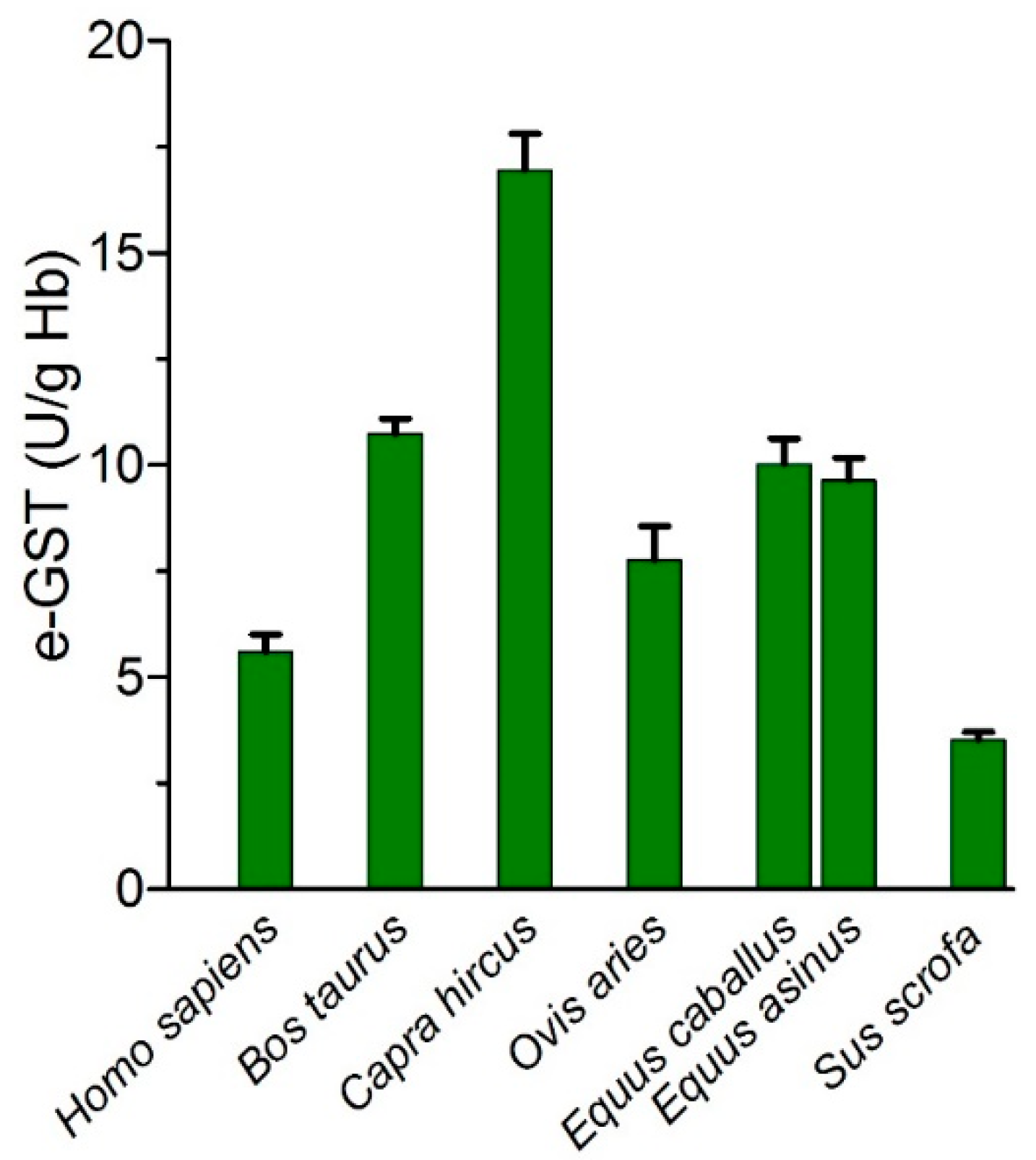
5. Utility of e-GST in Veterinary Field
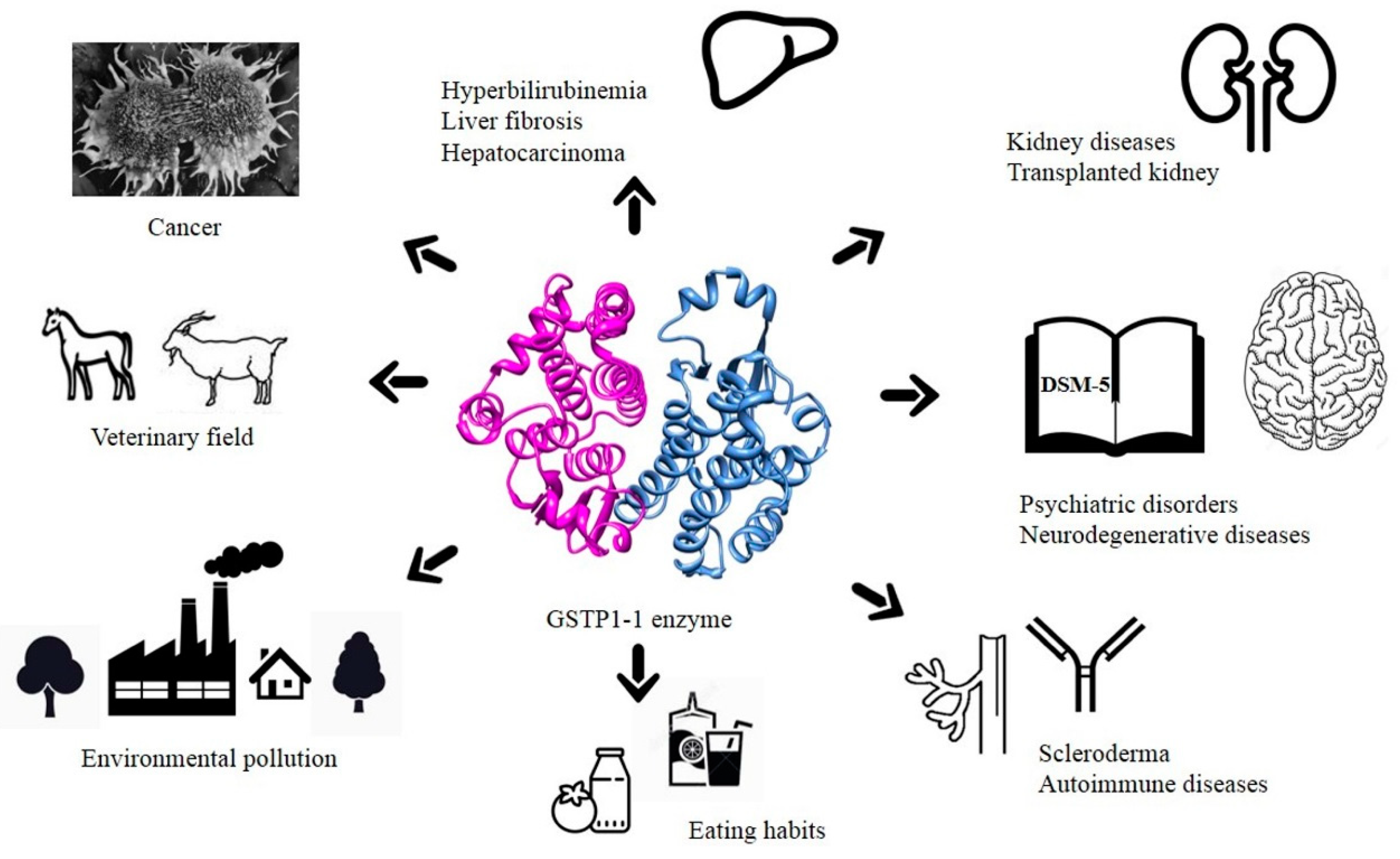
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| α-LA | Alpha-lipoic acid |
| AD | Alzheimer’s Disease |
| APC | Antigen-presenting cells |
| CAT | Catalase |
| CDAA | choline-deficient L-amino acid-defined diet |
| CDNB | 1-chloro-2,4-dinitrobenzene |
| CKD | Chronic kidney disease |
| CMD | Choline-methionine-deficient |
| DEN | Diethylnitrosamine |
| DNDGIC | dinitrosyl-diglutathionyl iron complex |
| e-GFR | Estimated-GFR |
| EGF-R | Epidermal growth factor-R |
| e-GST | Erythrocyte glutathione transferases |
| ELISA | Enzyme-linked immunosorbent assay |
| ESRD | End-stage-renal disease |
| GFR | Glomerular filtration rate |
| GR | Glutathione reductase |
| GSH | Glutathione |
| GSTP | Glutathione transferase class P |
| GSTs | Glutathione transferases |
| Hb | Hemoglobin |
| HCC | Hepatocellular carcinoma |
| Hcy | Homocysteine |
| HD | Hemodialysis |
| HDF | HemoDiaFiltration |
| MHD | Maintenance Hemodialysis |
| NBDHEX | 6-(7-nitro-2,1,3-benzoxidiazol-4-ylthio) hexanol |
| NFK K-DOQI | National Kidney Foundation Kidney- Disease Outcomes Quality Initiative |
| NO | Nitric oxide |
| NOS | Nitric oxide synthase |
| Nrf | Nuclear respiratory factor |
| OS | Oxidative stress |
| PD | Parkinson’s disease |
| PDB | Protein data bank |
| RBC | Red blood cell |
| ROS | Reactive oxygen species |
| SOCS-1 | Suppressor of cytokine signaling 1 |
| SOD | Superoxide dismutase |
| SSc | Scleroderma or Systemic sclerosis |
| T2DM | Type 2 diabetes mellitus |
| TNF-α | Tumor necrosis factor-α |
| TRAF2 | TNF-receptor factor 2 |
| TSGs | Tumor suppressor genes |
References
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem. J. 2001, 360, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Booth, J.; Boyland, E.; Sims, P. An enzyme from rat liver catalysing conjugations with glutathione. Biochem. J. 1961, 79, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.N. Structure, catalytic mechanism, and evolution of the glutathione transferases. Chem. Res. Toxicol. 1997, 10, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Oakley, A.J.; Lo Bello, M.; Battistoni, A.; Ricci, G.; Rossjohn, J.; Villar, H.O.; Parker, M.W. The structures of human glutathione transferase P1-1 in complex with glutathione and various inhibitors at high resolution. J. Mol. Biol. 1997, 274, 84–100. [Google Scholar] [CrossRef] [PubMed]
- Bocedi, A.; Fabrini, R.; Farrotti, A.; Stella, L.; Ketterman, A.J.; Pedersen, J.Z.; Allocati, N.; Lau, P.C.; Grosse, S.; Eltis, L.D.; et al. The impact of nitric oxide toxicity on the evolution of the glutathione transferase superfamily: A proposal for an evolutionary driving force. J. Biol. Chem. 2013, 288, 24936–24947. [Google Scholar] [CrossRef] [PubMed]
- Habig, W.H.; Jakoby, W.B. Assays for differentiation of glutathione S-transferases. Methods Enzymol. 1981, 77, 398–405. [Google Scholar]
- Johansson, K.; Jarvliden, J.; Gogvadze, V.; Morgenstern, R. Multiple roles of microsomal glutathione transferase 1 in cellular protection: A mechanistic study. Free Radic. Biol. Med. 2010, 49, 1638–1645. [Google Scholar] [CrossRef]
- Hubatsch, I.; Ridderstrom, M.; Mannervik, B. Human glutathione transferase A4-4: An alpha class enzyme with high catalytic efficiency in the conjugation of 4-hydroxynonenal and other genotoxic products of lipid peroxidation. Biochem. J. 1998, 330, 175–179. [Google Scholar] [CrossRef]
- Raffalli-Mathieu, F.; Mannervik, B. Human glutathione transferase A3-3 active as steroid double-bond isomerase. Methods Enzymol. 2005, 401, 265–278. [Google Scholar] [CrossRef]
- Bartling, D.; Radzio, R.; Steiner, U.; Weiler, E.W. A glutathione S-transferase with glutathione-peroxidase activity from Arabidopsis thaliana. Molecular cloning and functional characterization. Eur. J. Biochem. 1993, 216, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Cesareo, E.; Parker, L.J.; Pedersen, J.Z.; Nuccetelli, M.; Mazzetti, A.P.; Pastore, A.; Federici, G.; Caccuri, A.M.; Ricci, G.; Adams, J.J.; et al. Nitrosylation of human glutathione transferase P1-1 with dinitrosyl diglutathionyl iron complex in vitro and in vivo. J. Biol. Chem. 2005, 280, 42172–42180. [Google Scholar] [CrossRef] [PubMed]
- Fabrini, R.; Bocedi, A.; Dawood, K.F.; Turella, P.; Stella, L.; Parker, M.W.; Pedersen, J.Z.; Federici, G.; Antonini, G.; Ricci, G. The extended catalysis of glutathione transferase. FEBS Lett. 2011, 585, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, Y.C.; Singh, S.V. Purification and characterization of a new form of glutathione S-transferase from human erythrocytes. Biochem. Biophys. Res. Commun. 1984, 125, 1053–1060. [Google Scholar] [CrossRef]
- Ivanetich, K.M.; Goold, R.D. A rapid equilibrium random sequential bi-bi mechanism for human placental glutathione S-transferase. Biochim. Biophys. Acta 1989, 998, 7–13. [Google Scholar] [CrossRef]
- Ricci, G.; Lo Bello, M.; Caccurri, A.M.; Pastore, A.; Nuccetelli, M.; Parker, M.W.; Federici, G. Site-directed mutagenesis of human glutathione transferase P1-1. Mutation of Cys-47 induces a positive cooperativity in glutathione transferase P1-1. J. Biol. Chem. 1995, 270, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Lo Bello, M.; Parker, M.W.; Desideri, A.; Polticelli, F.; Falconi, M.; Del Boccio, G.; Pennelli, A.; Federici, G.; Ricci, G. Peculiar spectroscopic and kinetic properties of Cys-47 in human placental glutathione transferase. Evidence for an atypical thiolate ion pair near the active site. J. Biol. Chem. 1993, 268, 19033–19038. [Google Scholar] [PubMed]
- Lo Bello, M.; Oakley, A.J.; Battistoni, A.; Mazzetti, A.P.; Nuccetelli, M.; Mazzarese, G.; Rossjohn, J.; Parker, M.W.; Ricci, G. Multifunctional role of Tyr 108 in the catalytic mechanism of human glutathione transferase P1-1. Crystallographic and kinetic studies on the Y108F mutant enzyme. Biochemistry 1997, 36, 6207–6217. [Google Scholar] [CrossRef]
- Nicotra, M.; Paci, M.; Sette, M.; Oakley, A.J.; Parker, M.W.; Lo Bello, M.; Caccuri, A.M.; Federici, G.; Ricci, G. Solution structure of glutathione bound to human glutathione transferase P1-1: Comparison of NMR measurements with the crystal structure. Biochemistry 1998, 37, 3020–3027. [Google Scholar] [CrossRef]
- Caccuri, A.M.; Antonini, G.; Ascenzi, P.; Nicotra, M.; Nuccetelli, M.; Mazzetti, A.P.; Federici, G.; Lo Bello, M.; Ricci, G. Temperature adaptation of glutathione S-transferase P1-1. A case for homotropic regulation of substrate binding. J. Biol. Chem. 1999, 274, 19276–19280. [Google Scholar] [CrossRef]
- Lo Bello, M.; Nuccetelli, M.; Chiessi, E.; Lahm, A.; Mazzetti, A.P.; Battistoni, A.; Caccuri, A.M.; Oakley, A.J.; Parker, M.W.; Tramontano, A.; et al. Mutations of Gly to Ala in human glutathione transferase P1-1 affect helix 2 (G-site) and induce positive cooperativity in the binding of glutathione. J. Mol. Biol. 1998, 284, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Lo Bello, M.; Nuccetelli, M.; Caccuri, A.M.; Stella, L.; Parker, M.W.; Rossjohn, J.; McKinstry, W.J.; Mozzi, A.F.; Federici, G.; Polizio, F.; et al. Human glutathione transferase P1-1 and nitric oxide carriers; a new role for an old enzyme. J. Biol. Chem. 2001, 276, 42138–42145. [Google Scholar] [CrossRef]
- Becker, K.; Savvides, S.N.; Keese, M.; Schirmer, R.H.; Karplus, P.A. Enzyme inactivation through sulfhydryl oxidation by physiologic NO-carriers. Nat. Struct. Biol. 1998, 5, 267–271. [Google Scholar] [CrossRef]
- Ricci, G.; Caccuri, A.M.; Lo Bello, M.; Parker, M.W.; Nuccetelli, M.; Turella, P.; Stella, L.; Di Iorio, E.E.; Federici, G. Glutathione transferase P1-1: Self-preservation of an anti-cancer enzyme. Biochem. J. 2003, 376, 71–76. [Google Scholar] [CrossRef]
- Bocedi, A.; Fabrini, R.; Lo Bello, M.; Caccuri, A.M.; Federici, G.; Mannervik, B.; Cornish-Bowden, A.; Ricci, G. Evolution of Negative Cooperativity in Glutathione Transferase Enabled Preservation of Enzyme Function. J. Biol. Chem. 2016, 291, 26739–26749. [Google Scholar] [CrossRef]
- Wang, T.; Arifoglu, P.; Ronai, Z.; Tew, K.D. Glutathione S-transferase P1-1 (GSTP1-1) inhibits c-Jun N-terminal kinase (JNK1) signaling through interaction with the C terminus. J. Biol. Chem. 2001, 276, 20999–21003. [Google Scholar] [CrossRef] [PubMed]
- Kampranis, S.C.; Damianova, R.; Atallah, M.; Toby, G.; Kondi, G.; Tsichlis, P.N.; Makris, A.M. A novel plant glutathione S-transferase/peroxidase suppresses Bax lethality in yeast. J. Biol. Chem. 2000, 275, 29207–29216. [Google Scholar] [CrossRef] [PubMed]
- Dulhunty, A.; Gage, P.; Curtis, S.; Chelvanayagam, G.; Board, P. The glutathione transferase structural family includes a nuclear chloride channel and a ryanodine receptor calcium release channel modulator. J. Biol. Chem. 2001, 276, 3319–3323. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Fan, Y.; Xue, B.; Luo, L.; Shen, J.; Zhang, S.; Jiang, Y.; Yin, Z. Human glutathione S-transferase P1-1 interacts with TRAF2 and regulates TRAF2-ASK1 signals. Oncogene 2006, 25, 5787–5800. [Google Scholar] [CrossRef] [PubMed]
- Bartolini, D.; Commodi, J.; Piroddi, M.; Incipini, L.; Sancineto, L.; Santi, C.; Galli, F. Glutathione S-transferase pi expression regulates the Nrf2-dependent response to hormetic diselenides. Free Radic. Biol. Med. 2015, 88, 466–480. [Google Scholar] [CrossRef] [PubMed]
- Fabrini, R.; Bocedi, A.; Camerini, S.; Fusetti, M.; Ottaviani, F.; Passali, F.M.; Topazio, D.; Iavarone, F.; Francia, I.; Castagnola, M.; et al. Inactivation of human salivary glutathione transferase P1-1 by hypothiocyanite: A post-translational control system in search of a role. PLoS ONE 2014, 9, e112797. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fabrini, R.; Bocedi, A.; Massoud, R.; Federici, G.; Ricci, G. Spectrophotometric assay for serum glutathione transferase: A re-examination. Clin. Biochem. 2012, 45, 668–671. [Google Scholar] [CrossRef] [PubMed]
- Carmagnol, F.; Sinet, P.M.; Rapin, J.; Jerome, H. Glutathione-S-transferase of human red blood cells; assay, values in normal subjects and in two pathological circumstances: Hyperbilirubinemia and impaired renal function. Clin. Chim. Acta 1981, 117, 209–217. [Google Scholar] [CrossRef]
- Mimic-Oka, J.; Simic, T.; Djukanovic, L.; Stefanovski, J.; Ramic, Z. Glutathione and its associated enzymes in peripheral blood cells in different stages of chronic renal insufficiency. Amino Acids 1992, 2, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Galli, F.; Rovidati, S.; Benedetti, S.; Buoncristiani, U.; Covarelli, C.; Floridi, A.; Canestrari, F. Overexpression of erythrocyte glutathione S-transferase in uremia and dialysis. Clin. Chem. 1999, 45, 1781–1788. [Google Scholar] [PubMed]
- Kopple, J.D. National kidney foundation K/DOQI clinical practice guidelines for nutrition in chronic renal failure. Am. J. Kidney Dis. 2001, 37, S66–S70. [Google Scholar] [CrossRef]
- Dessi, M.; Noce, A.; Dawood, K.F.; Galli, F.; Taccone-Gallucci, M.; Fabrini, R.; Bocedi, A.; Massoud, R.; Fucci, G.; Pastore, A.; et al. Erythrocyte glutathione transferase: A potential new biomarker in chronic kidney diseases which correlates with plasma homocysteine. Amino Acids 2012, 43, 347–354. [Google Scholar] [CrossRef]
- Pastore, A.; Noce, A.; Di Giovamberardino, G.; De Stefano, A.; Calla, C.; Zenobi, R.; Dessi, M.; Di Daniele, N. Homocysteine, cysteine, folate and vitamin B(1)(2) status in type 2 diabetic patients with chronic kidney disease. J. Nephrol. 2015, 28, 571–576. [Google Scholar] [CrossRef]
- Welch, G.N.; Loscalzo, J. Homocysteine and atherothrombosis. N. Engl. J. Med. 1998, 338, 1042–1050. [Google Scholar] [CrossRef]
- Noce, A.; Ferrannini, M.; Fabrini, R.; Bocedi, A.; Dessi, M.; Galli, F.; Federici, G.; Palumbo, R.; Di Daniele, N.; Ricci, G. Erythrocyte glutathione transferase: A new biomarker for hemodialysis adequacy, overcoming the Kt/V(urea) dogma? Cell Death Dis. 2012, 3, e377. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Galli, F.; Piroddi, M.; Bartolini, D.; Ciffolilli, S.; Buoncristiani, E.; Ricci, G.; Buoncristiani, U. Blood thiol status and erythrocyte glutathione-S-transferase in chronic kidney disease patients on treatment with frequent (daily) hemodialysis. Free Radic. Res. 2014, 48, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Noce, A.; Fabrini, R.; Dessi, M.; Bocedi, A.; Santini, S.; Rovella, V.; Pastore, A.; Tesauro, M.; Bernardini, S.; Di Daniele, N.; et al. Erythrocyte glutathione transferase activity: A possible early biomarker for blood toxicity in uremic diabetic patients. Acta Diabetol. 2014, 51, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Noce, A.; Fabrini, R.; Bocedi, A.; Di Daniele, N. Erythrocyte glutathione transferase in uremic diabetic patients: Additional data. Acta Diabetol. 2015, 52, 813–815. [Google Scholar] [CrossRef] [PubMed]
- Tesauro, M.; Nistico, S.; Noce, A.; Tarantino, A.; Marrone, G.; Costa, A.; Rovella, V.; Di Cola, G.; Campia, U.; Lauro, D.; et al. The possible role of glutathione-S-transferase activity in diabetic nephropathy. Int. J. Immunopathol. Pharmacol. 2015, 28, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Velladath, S.U.; Das, A.; Rishikesh, K.; Nalini, K. Erythrocyte Glutathione-S-Transferase Activity In Diabetics And Its Association With HBA1c. Clin. Biochem. 2011, 2. [Google Scholar] [CrossRef]
- Wolfe, R.A.; Ashby, V.B.; Milford, E.L.; Ojo, A.O.; Ettenger, R.E.; Agodoa, L.Y.; Held, P.J.; Port, F.K. Comparison of mortality in all patients on dialysis, patients on dialysis awaiting transplantation, and recipients of a first cadaveric transplant. N. Engl. J. Med. 1999, 341, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Laupacis, A.; Keown, P.; Pus, N.; Krueger, H.; Ferguson, B.; Wong, C.; Muirhead, N. A study of the quality of life and cost-utility of renal transplantation. Kidney Int. 1996, 50, 235–242. [Google Scholar] [CrossRef]
- Bocedi, A.; Noce, A.; Rovella, V.; Marrone, G.; Cattani, G.; Iappelli, M.; De Paolis, P.; Iaria, G.; Sforza, D.; Gallu, M.; et al. Erythrocyte glutathione transferase in kidney transplantation: A probe for kidney detoxification efficiency. Cell Death Dis. 2018, 9, 288. [Google Scholar] [CrossRef]
- Rovcanin, B.; Medic, B.; Kocic, G.; Cebovic, T.; Ristic, M.; Prostran, M. Molecular Dissection of Renal Ischemia-Reperfusion: Oxidative Stress and Cellular Events. Curr. Med. Chem. 2016, 23, 1965–1980. [Google Scholar] [CrossRef]
- Guimaraes, J.; Araujo, A.M.; Santos, F.; Nunes, C.S.; Casal, M. Living-donor and Deceased-donor Renal Transplantation: Differences in Early Outcome--A Single-center Experience. Transpl. Proc. 2015, 47, 958–962. [Google Scholar] [CrossRef] [PubMed]
- Morozova, N.; Khrapko, K.; Panee, J.; Liu, W.; Harney, J.W.; Berry, M.J. Glutathione depletion in hippocampal cells increases levels of H and L ferritin and glutathione S-transferase mRNAs. Genes Cells 2007, 12, 561–567. [Google Scholar] [CrossRef]
- Johnson, W.M.; Wilson-Delfosse, A.L.; Mieyal, J.J. Dysregulation of glutathione homeostasis in neurodegenerative diseases. Nutrients 2012, 4, 1399–1440. [Google Scholar] [CrossRef] [PubMed]
- Mazzetti, A.P.; Fiorile, M.C.; Primavera, A.; Lo Bello, M. Glutathione transferases and neurodegenerative diseases. Neurochem. Int. 2015, 82, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Hollman, A.L.; Tchounwou, P.B.; Huang, H.C. The Association between Gene-Environment Interactions and Diseases Involving the Human GST Superfamily with SNP Variants. Int. J. Environ. Res. Public Health 2016, 13, 379. [Google Scholar] [CrossRef]
- Lescuyer, P.; Allard, L.; Zimmermann-Ivol, C.G.; Burgess, J.A.; Hughes-Frutiger, S.; Burkhard, P.R.; Sanchez, J.C.; Hochstrasser, D.F. Identification of post-mortem cerebrospinal fluid proteins as potential biomarkers of ischemia and neurodegeneration. Proteomics 2004, 4, 2234–2241. [Google Scholar] [CrossRef] [PubMed]
- Allocati, N.; Masulli, M.; Di Ilio, C.; Federici, L. Glutathione transferases: Substrates, inihibitors and pro-drugs in cancer and neurodegenerative diseases. Oncogenesis 2018, 7, 8. [Google Scholar] [CrossRef]
- Deng, Y.; Newman, B.; Dunne, M.P.; Silburn, P.A.; Mellick, G.D. Case-only study of interactions between genetic polymorphisms of GSTM1, P1, T1 and Z1 and smoking in Parkinson’s disease. Neurosci. Lett. 2004, 366, 326–331. [Google Scholar] [CrossRef]
- Longo, G.S.; Pinhel, M.S.; Sado, C.L.; Gregorio, M.L.; Amorim, G.S.; Florim, G.S.; Mazeti, C.M.; Martins, D.P.; Oliveira, F.N.; Tognola, W.A.; et al. Exposure to pesticides and heterozygote genotype of GSTP1-Alw26I are associated to Parkinson’s disease. Arq. Neuropsiquiatr. 2013, 71, 446–452. [Google Scholar] [CrossRef]
- Sun, K.H.; Chang, K.H.; Clawson, S.; Ghosh, S.; Mirzaei, H.; Regnier, F.; Shah, K. Glutathione-S-transferase P1 is a critical regulator of Cdk5 kinase activity. J. Neurochem. 2011, 118, 902–914. [Google Scholar] [CrossRef]
- Bernardini, S.; Bellincampi, L.; Ballerini, S.; Federici, G.; Iori, R.; Trequattrini, A.; Ciappi, F.; Baldinetti, F.; Bossu, P.; Caltagirone, C.; et al. Glutathione S-transferase P1 *C allelic variant increases susceptibility for late-onset Alzheimer disease: Association study and relationship with apolipoprotein E epsilon4 allele. Clin. Chem. 2005, 51, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Pinhel, M.A.; Nakazone, M.A.; Cacao, J.C.; Piteri, R.C.; Dantas, R.T.; Godoy, M.F.; Godoy, M.R.; Tognola, W.A.; Conforti-Froes, N.D.; Souza, D. Glutathione S-transferase variants increase susceptibility for late-onset Alzheimer’s disease: Association study and relationship with apolipoprotein E epsilon4 allele. Clin. Chem. Lab. Med. 2008, 46, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.S. Redefining epilepsy. Curr. Opin. Neurol. 2015, 28, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Shang, W.; Liu, W.H.; Zhao, X.H.; Sun, Q.J.; Bi, J.Z.; Chi, Z.F. Expressions of glutathione S-transferase alpha, mu, and pi in brains of medically intractable epileptic patients. BMC Neurosci. 2008, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Dhull, D.K.; Gupta, V.; Channana, P.; Singh, A.; Bhardwaj, M.; Ruhal, P.; Mittal, R. Role of Glutathione-S-transferases in neurological problems. Expert Opin. Ther. Pat. 2017, 27, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Kiernan, M.C.; Vucic, S.; Cheah, B.C.; Turner, M.R.; Eisen, A.; Hardiman, O.; Burrell, J.R.; Zoing, M.C. Amyotrophic lateral sclerosis. Lancet 2011, 377, 942–955. [Google Scholar] [CrossRef]
- Eum, K.D.; Seals, R.M.; Taylor, K.M.; Grespin, M.; Umbach, D.M.; Hu, H.; Sandler, D.P.; Kamel, F.; Weisskopf, M.G. Modification of the association between lead exposure and amyotrophic lateral sclerosis by iron and oxidative stress related gene polymorphisms. Amyotroph. Lateral Scler. Front. Degener. 2015, 16, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Usarek, E.; Gajewska, B.; Kazmierczak, B.; Kuzma, M.; Dziewulska, D.; Baranczyk-Kuzma, A. A study of glutathione S-transferase pi expression in central nervous system of subjects with amyotrophic lateral sclerosis using RNA extraction from formalin-fixed, paraffin-embedded material. Neurochem. Res. 2005, 30, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.J.; Yucel, M.; Pantelis, C.; Berk, M. Neurobiology of schizophrenia spectrum disorders: The role of oxidative stress. Ann. Acad Med. Singap. 2009, 38, 396. [Google Scholar]
- Chowdari, K.V.; Bamne, M.N.; Nimgaonkar, V.L. Genetic association studies of antioxidant pathway genes and schizophrenia. Antioxid Redox Signal. 2011, 15, 2037–2045. [Google Scholar] [CrossRef]
- Sarkhel, S.; Alcott Sadoc, V. Kaplan and Sadock’s Synopsis of Psychiatry: Behavioral Sciences/Clinical Psychiatry, 10th ed.; Philadelphia Indian: Philadelphia, PA, USA, 2009; Volume 51, pp. 1–1470. [Google Scholar]
- Rund, B.R. Is schizophrenia a neurodegenerative disorder? Nord. J. Psychiatry 2009, 63, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Prokhorova, T.A.; Tereshkina, E.B.; Savushkina, O.K.; Boksha, I.S.; Vorobyeva, E.A.; Omel’chenko, M.A.; Pomytkin, A.N.; Kaleda, V.G.; Burbaeva, G.S. [The activity of enzymes of glutathione metabolism in blood cells of patients with a high risk of manifestation of endogenous psychoses and patients with the first psychotic episode]. Zh. Nevrol. Psikhiatr. Im. S S Korsakova 2019, 119, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Savushkina, O.K.; Boksha, I.S.; Prokhorova, T.A.; Tereshkina, E.B.; Burminskiy, D.S.; Morozova, M.A.; Vorobyeva, E.A.; Burbaeva, G.S. [The activity of erythrocyte and platelet glutathione reductase and glutathione-S-transferase in paranoid schizophrenia]. Zh. Nevrol. Psikhiatr. Im. S S Korsakova 2018, 118, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Gravina, P.; Spoletini, I.; Masini, S.; Valentini, A.; Vanni, D.; Paladini, E.; Bossu, P.; Caltagirone, C.; Federici, G.; Spalletta, G.; et al. Genetic polymorphisms of glutathione S-transferases GSTM1, GSTT1, GSTP1 and GSTA1 as risk factors for schizophrenia. Psychiatry Res. 2011, 187, 454–456. [Google Scholar] [CrossRef] [PubMed]
- Pae, C.U.; Kim, J.J.; Lee, S.J.; Lee, C.U.; Lee, C.; Paik, I.H.; Park, H.R.; Yang, S.; Serretti, A. Association study between glutathione S-transferase P1 polymorphism and schizophrenia in the Korean population. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 519–523. [Google Scholar] [CrossRef]
- Lord, C.; Cook, E.H.; Leventhal, B.L.; Amaral, D.G. Autism spectrum disorders. Neuron 2000, 28, 355–363. [Google Scholar] [CrossRef]
- Rahbar, M.H.; Samms-Vaughan, M.; Ma, J.; Bressler, J.; Loveland, K.A.; Hessabi, M.; Dickerson, A.S.; Grove, M.L.; Shakespeare-Pellington, S.; Beecher, C.; et al. Interaction between GSTT1 and GSTP1 allele variants as a risk modulating-factor for autism spectrum disorders. Res. Autism Spectr. Disord. 2015, 12, 1–9. [Google Scholar] [CrossRef]
- LeRoy, E.C.; Black, C.; Fleischmajer, R.; Jablonska, S.; Krieg, T.; Medsger, T.A., Jr.; Rowell, N.; Wollheim, F. Scleroderma (systemic sclerosis): Classification, subsets and pathogenesis. J. Rheumatol. 1988, 15, 202–205. [Google Scholar]
- Broen, J.C.; Coenen, M.J.; Radstake, T.R. Genetics of systemic sclerosis: An update. Curr. Rheumatol. Rep. 2012, 14, 11–21. [Google Scholar] [CrossRef]
- Fietta, A.; Bardoni, A.; Salvini, R.; Passadore, I.; Morosini, M.; Cavagna, L.; Codullo, V.; Pozzi, E.; Meloni, F.; Montecucco, C. Analysis of bronchoalveolar lavage fluid proteome from systemic sclerosis patients with or without functional, clinical and radiological signs of lung fibrosis. Arthritis Res. Ther. 2006, 8, R160. [Google Scholar] [CrossRef]
- Medsger, T.A., Jr.; Bombardieri, S.; Czirjak, L.; Scorza, R.; Della Rossa, A.; Bencivelli, W. Assessment of disease severity and prognosis. Clin. Exp. Rheumatol. 2003, 21, S42–S46. [Google Scholar] [PubMed]
- Valentini, G.; Silman, A.J.; Veale, D. Assessment of disease activity. Clin. Exp. Rheumatol. 2003, 21, S39–S41. [Google Scholar] [PubMed]
- Fabrini, R.; Rosato, E.; Gigante, A.; Bocedi, A.; Cianci, R.; Barbano, B.; Del Grosso, E.; Ricci, F.; Zingaretti, V.; Salsano, F.; et al. Erythrocyte glutathione transferase: A non-antibody biomarker for systemic sclerosis, which correlates with severity and activity of the disease. Cell Death Dis. 2013, 4, e736. [Google Scholar] [CrossRef] [PubMed]
- Rose, N.R. Prediction and Prevention of Autoimmune Disease in the 21st Century: A Review and Preview. Am. J. Epidemiol. 2016, 183, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Glesse, N.; Rohr, P.; Monticielo, O.A.; Rech, T.F.; Brenol, J.C.; Xavier, R.M.; Kvitko, K.; Chies, J.A. Genetic polymorphisms of glutathione S-transferases and cytochrome P450 enzymes as susceptibility factors to systemic lupus erythematosus in southern Brazilian patients. Mol. Biol. Rep. 2014, 41, 6167–6179. [Google Scholar] [CrossRef] [PubMed]
- Song, G.G.; Bae, S.C.; Lee, Y.H. The glutathione S-transferase M1 and P1 polymorphisms and rheumatoid arthritis: A meta-analysis. Mol. Biol. Rep. 2012, 39, 10739–10745. [Google Scholar] [CrossRef]
- Agundez, J.A.; Garcia-Martin, E.; Martinez, C.; Benito-Leon, J.; Millan-Pascual, J.; Diaz-Sanchez, M.; Calleja, P.; Pisa, D.; Turpin-Fenoll, L.; Alonso-Navarro, H.; et al. The GSTP1 gene variant rs1695 is not associated with an increased risk of multiple sclerosis. Cell Mol. Immunol. 2015, 12, 777–779. [Google Scholar] [CrossRef]
- Salminen, L.E.; Paul, R.H. Oxidative stress and genetic markers of suboptimal antioxidant defense in the aging brain: A theoretical review. Rev. Neurosci. 2014, 25, 805–819. [Google Scholar] [CrossRef]
- Neefjes, V.M.; Evelo, C.T.; Baars, L.G.; Blanco, C.E. Erythrocyte glutathione S transferase as a marker of oxidative stress at birth. Arch. Dis. Child. Fetal Neonatal Ed. 1999, 81, F130–F133. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Glutathione and modulation of cell apoptosis. Biochim. Biophys. Acta 2012, 1823, 1767–1777. [Google Scholar] [CrossRef]
- Milic, M.; Frustaci, A.; Del Bufalo, A.; Sanchez-Alarcon, J.; Valencia-Quintana, R.; Russo, P.; Bonassi, S. DNA damage in non-communicable diseases: A clinical and epidemiological perspective. Mutat. Res. 2015, 776, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Walker, S.O.; Hong, X.; Bartell, T.R.; Wang, X. Epigenetics and early life origins of chronic noncommunicable diseases. J. Adolesc. Health 2013, 52, S14–S21. [Google Scholar] [CrossRef] [PubMed]
- Gouda, M.; Moustafa, A.; Hussein, L.; Hamza, M. Three week dietary intervention using apricots, pomegranate juice or/and fermented sour sobya and impact on biomarkers of antioxidative activity, oxidative stress and erythrocytic glutathione transferase activity among adults. Nutr. J. 2016, 15, 52. [Google Scholar] [CrossRef] [PubMed]
- Noce, A.; Marrone, G.; Di Daniele, F.; Ottaviani, E.; Wilson Jones, G.; Bernini, R.; Romani, A.; Rovella, V. Impact of Gut Microbiota Composition on Onset and Progression of Chronic Non-Communicable Diseases. Nutrients 2019, 11, 1073. [Google Scholar] [CrossRef] [PubMed]
- Noce, A.; Vidiri, M.F.; Marrone, G.; Moriconi, E.; Bocedi, A.; Capria, A.; Rovella, V.; Ricci, G.; De Lorenzo, A.; Di Daniele, N. Is low-protein diet a possible risk factor of malnutrition in chronic kidney disease patients? Cell Death Discov. 2016, 2, 16026. [Google Scholar] [CrossRef] [PubMed]
- Mariani, E.; Polidori, M.C.; Cherubini, A.; Mecocci, P. Oxidative stress in brain aging, neurodegenerative and vascular diseases: An overview. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 827, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Muller, F.L.; Lustgarten, M.S.; Jang, Y.; Richardson, A.; Van Remmen, H. Trends in oxidative aging theories. Free Radic. Biol. Med. 2007, 43, 477–503. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.D.; Sokoloff, L. Ch. 31 Circulation and Energy Metabolism of the Brain. In Basic Neurochemistry: Molecular, Cellular, and Medical Aspects, 6th ed.; Siegel, G.J., Agranoff, B.W., Albers, R.W., Fisher, S.K., Uhler, M.D., Eds.; Philadelphia Indian: Philadelphia, PA, USA, 1999; pp. 637–671. [Google Scholar]
- Dringen, R. Metabolism and functions of glutathione in brain. Prog. Neurobiol. 2000, 62, 649–671. [Google Scholar] [CrossRef]
- Hensley, K.; Robinson, K.A.; Gabbita, S.P.; Salsman, S.; Floyd, R.A. Reactive oxygen species, cell signaling, and cell injury. Free Radic. Biol. Med. 2000, 28, 1456–1462. [Google Scholar] [CrossRef]
- Storz, P. Mitochondrial ROS--radical detoxification, mediated by protein kinase D. Trends Cell Biol. 2007, 17, 13–18. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Strange, R.C. Glutathione S-transferase polymorphisms and their biological consequences. Pharmacology 2000, 61, 154–166. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, C.C.; Townsend, D.M.; Tew, K.D. Glutathione S-transferase polymorphisms: cancer incidence and therapy. Oncogene 2006, 25, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Gupta, S. The multifaceted role of glutathione S-transferases in cancer. Cancer Lett. 2018, 433, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Tew, K.D.; Manevich, Y.; Grek, C.; Xiong, Y.; Uys, J.; Townsend, D.M. The role of glutathione S-transferase P in signaling pathways and S-glutathionylation in cancer. Free Radic. Biol. Med. 2011, 51, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Pasello, M.; Michelacci, F.; Scionti, I.; Hattinger, C.M.; Zuntini, M.; Caccuri, A.M.; Scotlandi, K.; Picci, P.; Serra, M. Overcoming glutathione S-transferase P1-related cisplatin resistance in osteosarcoma. Cancer Res. 2008, 68, 6661–6668. [Google Scholar] [CrossRef] [PubMed]
- Sau, A.; Pellizzari Tregno, F.; Valentino, F.; Federici, G.; Caccuri, A.M. Glutathione transferases and development of new principles to overcome drug resistance. Arch. Biochem. Biophys. 2010, 500, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Mignani, S.; El Brahmi, N.; El Kazzouli, S.; Eloy, L.; Courilleau, D.; Caron, J.; Bousmina, M.M.; Caminade, A.M.; Cresteil, T.; Majoral, J.P. A novel class of ethacrynic acid derivatives as promising drug-like potent generation of anticancer agents with established mechanism of action. Eur. J. Med. Chem. 2016, 122, 656–673. [Google Scholar] [CrossRef]
- Wu, J.H.; Batist, G. Glutathione and glutathione analogues; therapeutic potentials. Biochim. Biophys. Acta 2013, 1830, 3350–3353. [Google Scholar] [CrossRef]
- Galal, A.M.; Walker, L.A.; Khan, I.A. Induction of GST and related events by dietary phytochemicals: Sources, chemistry, and possible contribution to chemoprevention. Curr. Top. Med. Chem. 2015, 14, 2802–2821. [Google Scholar] [CrossRef]
- Harshbarger, W.; Gondi, S.; Ficarro, S.B.; Hunter, J.; Udayakumar, D.; Gurbani, D.; Singer, W.D.; Liu, Y.; Li, L.; Marto, J.A.; et al. Structural and Biochemical Analyses Reveal the Mechanism of Glutathione S-Transferase Pi 1 Inhibition by the Anti-cancer Compound Piperlongumine. J. Biol. Chem. 2017, 292, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Johansson, K.; Ito, M.; Schophuizen, C.M.; Mathew Thengumtharayil, S.; Heuser, V.D.; Zhang, J.; Shimoji, M.; Vahter, M.; Ang, W.H.; Dyson, P.J.; et al. Characterization of new potential anticancer drugs designed to overcome glutathione transferase mediated resistance. Mol. Pharm. 2011, 8, 1698–1708. [Google Scholar] [CrossRef]
- Qu, Y.; Dang, S.; Hou, P. Gene methylation in gastric cancer. Clin. Chim. Acta 2013, 424, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Gurioli, G.; Martignano, F.; Salvi, S.; Costantini, M.; Gunelli, R.; Casadio, V. GSTP1 methylation in cancer: A liquid biopsy biomarker? Clin. Chem. Lab. Med. 2018, 56, 702–717. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, G.J.; Neri, I.A.; Sforni, V.C.; Kameo, R.; Lorand-Metze, I.; Lima, C.S. Polymorphisms of glutathione S-transferase Mu 1, glutathione S-transferase theta 1 and glutathione S-transferase Pi 1 genes in Hodgkin’s lymphoma susceptibility and progression. Leuk. Lymphoma 2009, 50, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Sailaja, K.; Surekha, D.; Rao, D.N.; Rao, D.R.; Vishnupriya, S. Association of the GSTP1 gene (Ile105Val) polymorphism with chronic myeloid leukemia. Asian Pac. J. Cancer Prev. 2010, 11, 461–464. [Google Scholar] [PubMed]
- Dunna, N.R.; Vuree, S.; Kagita, S.; Surekha, D.; Digumarti, R.; Rajappa, S.; Satti, V. Association of GSTP1 gene (I105V) polymorphism with acute leukaemia. J. Genet. 2012, 91, e60–e63. [Google Scholar] [CrossRef]
- Schnekenburger, M.; Karius, T.; Diederich, M. Regulation of epigenetic traits of the glutathione S-transferase P1 gene: From detoxification toward cancer prevention and diagnosis. Front. Pharmacol. 2014, 5, 170. [Google Scholar] [CrossRef]
- Dasgupta, R.K.; Adamson, P.J.; Davies, F.E.; Rollinson, S.; Roddam, P.L.; Ashcroft, A.J.; Dring, A.M.; Fenton, J.A.; Child, J.A.; Allan, J.M.; et al. Polymorphic variation in GSTP1 modulates outcome following therapy for multiple myeloma. Blood 2003, 102, 2345–2350. [Google Scholar] [CrossRef]
- Kilburn, L.; Okcu, M.F.; Wang, T.; Cao, Y.; Renfro-Spelman, A.; Aldape, K.D.; Gilbert, M.R.; Bondy, M. Glutathione S-transferase polymorphisms are associated with survival in anaplastic glioma patients. Cancer 2010, 116, 2242–2249. [Google Scholar] [CrossRef]
- Lai, R.; Crevier, L.; Thabane, L. Genetic polymorphisms of glutathione S-transferases and the risk of adult brain tumors: A meta-analysis. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1784–1790. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, Z.; Cui, D.; Liu, H.; Hao, X. Glutathione S-transferase P1 Ile105Val polymorphism and breast cancer risk: A meta-analysis involving 34,658 subjects. Breast Cancer Res. Treat. 2011, 125, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Shu, X.O.; Ruan, Z.X.; Cai, Q.Y.; Jin, F.; Gao, Y.T.; Zheng, W. Genetic polymorphisms in glutathione-S-transferase genes (GSTM1, GSTT1, GSTP1) and survival after chemotherapy for invasive breast carcinoma. Cancer 2005, 103, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Gao, B.; Li, R.; Li, W.; Chen, W.; Yu, Z.; Zhang, J. Expression levels of resistant genes affect cervical cancer prognosis. Mol. Med. Rep. 2017, 15, 2802–2806. [Google Scholar] [CrossRef] [PubMed]
- Coughlin, S.S.; Hall, I.J. Glutathione S-transferase polymorphisms and risk of ovarian cancer: a HuGE review. Genet. Med. 2002, 4, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Padilla, I.; Amir, E.; Marsh, S.; Liu, G.; Mackay, H. Genetic polymorphisms as predictive and prognostic biomarkers in gynecological cancers: A systematic review. Gynecol. Oncol. 2012, 124, 354–365. [Google Scholar] [CrossRef]
- Chan, Q.K.; Khoo, U.S.; Ngan, H.Y.; Yang, C.Q.; Xue, W.C.; Chan, K.Y.; Chiu, P.M.; Ip, P.P.; Cheung, A.N. Single nucleotide polymorphism of pi-class glutathione s-transferase and susceptibility to endometrial carcinoma. Clin. Cancer Res. 2005, 11, 2981–2985. [Google Scholar] [CrossRef]
- Pernar, C.H.; Ebot, E.M.; Wilson, K.M.; Mucci, L.A. The Epidemiology of Prostate Cancer. Cold Spring Harb. Perspect. Med. 2018, 8. [Google Scholar] [CrossRef]
- Kanwal, R.; Pandey, M.; Bhaskaran, N.; Maclennan, G.T.; Fu, P.; Ponsky, L.E.; Gupta, S. Protection against oxidative DNA damage and stress in human prostate by glutathione S-transferase P1. Mol. Carcinog. 2014, 53, 8–18. [Google Scholar] [CrossRef]
- Simic, T.; Savic-Radojevic, A.; Pljesa-Ercegovac, M.; Matic, M.; Mimic-Oka, J. Glutathione S-transferases in kidney and urinary bladder tumors. Nat. Rev. Urol. 2009, 6, 281–289. [Google Scholar] [CrossRef]
- Lafuente, A.; Rodriguez, A.; Gibanel, R.; Lafuente, M.J.; Alonso, M.; Molina, R.; Ballesta, A.; Carretero, P. Limitations in the use of glutathione S-transferase P1 in urine as a marker for bladder cancer. Anticancer. Res. 1998, 18, 3771–3772. [Google Scholar] [PubMed]
- Berendsen, C.L.; Mulder, T.P.; Peters, W.H. Plasma glutathione S-transferase pi 1-1 AND alpha 1-1 levels in patients with bladder cancer. J. Urol. 2000, 164, 2126–2128. [Google Scholar] [CrossRef]
- Jankova, L.; Robertson, G.; Chan, C.; Tan, K.L.; Kohonen-Corish, M.; Fung, C.L.; Clarke, C.; Lin, B.P.; Molloy, M.; Chapuis, P.H.; et al. Glutathione S-transferase Pi expression predicts response to adjuvant chemotherapy for stage C colon cancer: A matched historical control study. BMC Cancer 2012, 12, 196. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Pan, X.; Su, T.; Mo, Z.; Cao, Y.; Gao, F. Glutathione S-transferase P1 Ile105Val polymorphism and colorectal cancer risk: A meta-analysis and HuGE review. Eur. J. Cancer 2009, 45, 3303–3314. [Google Scholar] [CrossRef] [PubMed]
- Economopoulos, K.P.; Sergentanis, T.N. GSTM1, GSTT1, GSTP1, GSTA1 and colorectal cancer risk: A comprehensive meta-analysis. Eur. J. Cancer 2010, 46, 1617–1631. [Google Scholar] [CrossRef] [PubMed]
- Crew, K.D.; Neugut, A.I. Epidemiology of gastric cancer. World J. Gastroenterol. 2006, 12, 354–362. [Google Scholar] [CrossRef] [PubMed]
- de Araujo, R.M.; de Melo, C.F.; Neto, F.M.; da Silva, J.N.; Soares, L.F.; de Arruda Cardoso Smith, M.; Sousa, E.C., Jr.; Burbano, R.M.; de Medeiros, A.C.; Lima, E.M. Association study of SNPs of genes IFNGR1 (rs137854905), GSTT1 (rs71748309), and GSTP1 (rs1695) in gastric cancer development in samples of patient in the northern and northeastern Brazil. Tumour. Biol. 2014, 35, 4983–4986. [Google Scholar] [CrossRef]
- Xu, Z.; Zhu, H.; Luk, J.M.; Wu, D.; Gu, D.; Gong, W.; Tan, Y.; Zhou, J.; Tang, J.; Zhang, Z.; et al. Clinical significance of SOD2 and GSTP1 gene polymorphisms in Chinese patients with gastric cancer. Cancer 2012, 118, 5489–5496. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Janssen, M.J.; van Oijen, M.G.; Bergevoet, S.M.; te Morsche, R.H.; van Asten, H.; Laheij, R.J.; Peters, W.H.; Jansent, J.B. Genetic polymorphisms in GSTA1, GSTP1, GSTT1, and GSTM1 and gastric cancer risk in a Vietnamese population. Oncol. Res. 2010, 18, 349–355. [Google Scholar] [CrossRef]
- Li, Q.F.; Yao, R.Y.; Liu, K.W.; Lv, H.Y.; Jiang, T.; Liang, J. Genetic polymorphism of GSTP1: Prediction of clinical outcome to oxaliplatin/5-FU-based chemotherapy in advanced gastric cancer. J. Korean Med. Sci. 2010, 25, 846–852. [Google Scholar] [CrossRef]
- Ogino, S.; Konishi, H.; Ichikawa, D.; Matsubara, D.; Shoda, K.; Arita, T.; Kosuga, T.; Komatsu, S.; Shiozaki, A.; Okamoto, K.; et al. Glutathione S-transferase Pi 1 is a valuable predictor for cancer drug resistance in esophageal squamous cell carcinoma. Cancer Sci. 2019, 110, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Gu, J.; Wu, T.T.; Swisher, S.G.; Liao, Z.; Correa, A.M.; Liu, J.; Etzel, C.J.; Amos, C.I.; Huang, M.; et al. Genetic variations in radiation and chemotherapy drug action pathways predict clinical outcomes in esophageal cancer. J. Clin. Oncol. 2006, 24, 3789–3798. [Google Scholar] [CrossRef] [PubMed]
- Hiyama, T.; Yoshihara, M.; Tanaka, S.; Chayama, K. Genetic polymorphisms and esophageal cancer risk. Int. J. Cancer 2007, 121, 1643–1658. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Bondy, M.L.; Hassan, M.M.; Chang, D.Z.; Abbruzzese, J.L.; Evans, D.B.; Smolensky, M.H.; Li, D. Glutathione S-transferase gene polymorphisms and risk and survival of pancreatic cancer. Cancer 2007, 109, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Tezuka, S.; Ueno, M.; Kobayashi, S.; Morimoto, M.; Ohkawa, S.; Hirotani, A.; Tozuka, Y.; Moriya, S.; Nakamura, Y.; Miyagi, Y.; et al. Predictive value of ERCC1, ERCC2, ERCC4, and glutathione S-Transferase Pi expression for the efficacy and safety of FOLFIRINOX in patients with unresectable pancreatic cancer. Am. J. Cancer Res. 2018, 8, 2096–2105. [Google Scholar] [PubMed]
- Lu, C.; Spitz, M.R.; Zhao, H.; Dong, Q.; Truong, M.; Chang, J.Y.; Blumenschein, G.R., Jr.; Hong, W.K.; Wu, X. Association between glutathione S-transferase pi polymorphisms and survival in patients with advanced nonsmall cell lung carcinoma. Cancer 2006, 106, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Ada, A.O.; Kunak, S.C.; Hancer, F.; Bilgen, S.; Suzen, S.H.; Alpar, S.; Gulhan, M.; Kurt, B.; Iscan, M. CYP and GST polymorphisms and survival in advanced non-small cell lung cancer patients. Neoplasma 2010, 57, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Song, H.; Higgins, J.P.; Pharoah, P.; Danesh, J. Five glutathione s-transferase gene variants in 23,452 cases of lung cancer and 30,397 controls: Meta-analysis of 130 studies. PLoS Med. 2006, 3, e91. [Google Scholar] [CrossRef] [PubMed]
- Linares, M.A.; Zakaria, A.; Nizran, P. Skin Cancer. Prim. Care 2015, 42, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Liu, T.; Li, X.; Xu, X.; Fan, D. Contribution of glutathione S-transferase gene polymorphisms to development of skin cancer. Int. J. Clin. Exp. Med. 2015, 8, 377–386. [Google Scholar] [PubMed]
- Ibarrola-Villava, M.; Martin-Gonzalez, M.; Lazaro, P.; Pizarro, A.; Lluch, A.; Ribas, G. Role of glutathione S-transferases in melanoma susceptibility: Association with GSTP1 rs1695 polymorphism. Br. J. Dermatol. 2012, 166, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.I.; Wu, M.M.; Wang, Y.H.; Lee, C.Y.; Yang, T.Y.; Hsiao, B.Y.; Chen, C.J. Association of Environmental Arsenic Exposure, Genetic Polymorphisms of Susceptible Genes, and Skin Cancers in Taiwan. Biomed Res. Int. 2015, 2015, 892579. [Google Scholar] [CrossRef]
- Ottaviani, G.; Jaffe, N. The epidemiology of osteosarcoma. Cancer Treat. Res. 2009, 152, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.R.; Wu, J.; Li, R. Contribution of the GSTP1 gene polymorphism to the development of osteosarcoma in a Chinese population. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, H.; He, M.; Wu, H.; Zhu, Y.; Su, Z. The association of glutathione S-transferase polymorphisms in patients with osteosarcoma: Evidence from a meta-analysis. Eur. J. Cancer Care (Engl.) 2015, 24, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yi, Z.; Ling, M.; Shi, J.; Qiu, Y.; Yang, S. Predictive potential of ABCB1, ABCC3, and GSTP1 gene polymorphisms on osteosarcoma survival after chemotherapy. Tumour Biol. 2014, 35, 9897–9904. [Google Scholar] [CrossRef]
- Yang, B.; Guo, M.; Herman, J.G.; Clark, D.P. Aberrant promoter methylation profiles of tumor suppressor genes in hepatocellular carcinoma. Am. J. Pathol. 2003, 163, 1101–1107. [Google Scholar] [CrossRef]
- Sakaida, I.; Kubota, M.; Kayano, K.; Takenaka, K.; Mori, K.; Okita, K. Prevention of fibrosis reduces enzyme-altered lesions in the rat liver. Carcinogenesis 1994, 15, 2201–2206. [Google Scholar] [CrossRef]
- Sakaida, I.; Hironaka, K.; Uchida, K.; Suzuki, C.; Kayano, K.; Okita, K. Fibrosis accelerates the development of enzyme-altered lesions in the rat liver. Hepatology 1998, 28, 1247–1252. [Google Scholar] [CrossRef]
- Sakaida, I.; Hironaka, K.; Uchida, K.; Okita, K. Loss of inhibitory growth regulation by TGF-beta1 in preneoplastic lesions in rat liver. Dig. Dis. Sci. 2000, 45, 325–333. [Google Scholar] [CrossRef]
- Bertuccio, P.; Turati, F.; Carioli, G.; Rodriguez, T.; La Vecchia, C.; Malvezzi, M.; Negri, E. Global trends and predictions in hepatocellular carcinoma mortality. J. Hepatol. 2017, 67, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Orru, C.; Szydlowska, M.; Taguchi, K.; Zavattari, P.; Perra, A.; Yamamoto, M.; Columbano, A. Genetic inactivation of Nrf2 prevents clonal expansion of initiated cells in a nutritional model of rat hepatocarcinogenesis. J. Hepatol. 2018, 69, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Kin, M.; Torimura, T.; Ueno, T.; Nakamura, T.; Ogata, R.; Sakamoto, M.; Tamaki, S.; Sata, M. Angiogenesis inhibitor TNP-470 suppresses the progression of experimentally-induced hepatocellular carcinoma in rats. Int. J. Oncol. 2000, 16, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Perra, A.; Pibiri, M.; Sulas, P.; Simbula, G.; Ledda-Columbano, G.M.; Columbano, A. Alpha-lipoic acid promotes the growth of rat hepatic pre-neoplastic lesions in the choline-deficient model. Carcinogenesis 2008, 29, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.Y.; Kim, K.T.; Han, S.Y. Expression of ErbB receptor proteins and TGF-alpha during diethylnitrosamine-induced hepatocarcinogenesis in the rat liver. Korean J. Hepatol. 2007, 13, 70–80. [Google Scholar]
- Chen, Y.L.; Tseng, H.S.; Kuo, W.H.; Yang, S.F.; Chen, D.R.; Tsai, H.T. Glutathione S-Transferase P1 (GSTP1) gene polymorphism increases age-related susceptibility to hepatocellular carcinoma. BMC Med. Genet. 2010, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, Y.; Chen, H.; Mao, L. Clinical significance and association of GSTP1 hypermethylation with hepatocellular carcinoma: A meta-analysis. J. Cancer Res. Ther. 2018, 14, S486–S489. [Google Scholar] [CrossRef]
- Wang, Z.; Qu, K.; Niu, W.; Lin, T.; Xu, X.; Huang, Z.; Liu, S.; Liu, S.; Chang, H.; Liu, Y.; et al. Glutathione S-transferase P1 gene rs4147581 polymorphism predicts overall survival of patients with hepatocellular carcinoma: Evidence from an enlarged study. Tumour Biol. 2016, 37, 943–952. [Google Scholar] [CrossRef]
- Kelishadi, R.; Mirghaffari, N.; Poursafa, P.; Gidding, S.S. Lifestyle and environmental factors associated with inflammation, oxidative stress and insulin resistance in children. Atherosclerosis 2009, 203, 311–319. [Google Scholar] [CrossRef]
- Kelishadi, R.; Poursafa, P. Air pollution and non-respiratory health hazards for children. Arch. Med. Sci. 2010, 6, 483–495. [Google Scholar] [CrossRef]
- van der Oost, R.; Beyer, J.; Vermeulen, N.P. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Rudneva, I.I.; Kuzminova, N.S.; Skuratovskaya, E.N. Glutathione-S-Transferase Activity in Tissues of Black Sea Fish Species. Asian J. Exp. Biol. Sci. 2010, 1, 141–150. [Google Scholar]
- Sen, A.; Kirikbakan, A. Biochemical characterization and distribution of glutathione S-transferases in leaping mullet (Liza saliens). Biochemistry (Mosc.) 2004, 69, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Slatinska, I.; Smutna, M.; Havelkova, M.; Svobodova, Z. Biochemical markers of aquatic pollution in fish-glutathione S-transferase. Folia Vet. Lat. 2008, 52, 129–134. [Google Scholar]
- Fabrini, R.; Bocedi, A.; Del Grosso, E.; Morici, L.; Federici, G.; Palleschi, A.; Ricci, G. Erythrocyte glutathione transferase: A novel biomarker to check environmental pollution hazardous for humans. Biochem. Biophys. Res. Commun. 2012, 426, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Ansari, G.A.; Singh, S.V.; Gan, J.C.; Awasthi, Y.C. Human erythrocyte glutathione S-transferase: A possible marker of chemical exposure. Toxicol Lett. 1987, 37, 57–62. [Google Scholar] [CrossRef]
- Singh, S.V.; Awasthi, Y.C. Inhibition of human glutathione S-transferases by 2,4-dichlorophenoxyacetate (2,4-D) and 2,4,5-trichlorophenoxyacetate (2,4,5-T). Toxicol. Appl. Pharmacol. 1985, 81, 328–336. [Google Scholar] [CrossRef]
- Kilpikari, I.; Savolainen, H. Decreased erythrocyte glutathione s-transferase activity in rubber workers. Int. Arch. Occup. Environ. Health 1984, 53, 299–302. [Google Scholar] [CrossRef]
- Primavera, A.; Fustinoni, S.; Biroccio, A.; Ballerini, S.; Urbani, A.; Bernardini, S.; Federici, G.; Capucci, E.; Manno, M.; Lo Bello, M. Glutathione transferases and glutathionylated hemoglobin in workers exposed to low doses of 1,3-butadiene. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3004–3012. [Google Scholar] [CrossRef]
- Dai, X.; Bowatte, G.; Lowe, A.J.; Matheson, M.C.; Gurrin, L.C.; Burgess, J.A.; Dharmage, S.C.; Lodge, C.J. Do Glutathione S-Transferase Genes Modify the Link between Indoor Air Pollution and Asthma, Allergies, and Lung Function? A Systematic Review. Curr. Allergy Asthma Rep. 2018, 18, 20. [Google Scholar] [CrossRef]
- Sharma, R.K.; Upadhyay, G.; Siddiqi, N.J.; Sharma, B. Pesticides-induced biochemical alterations in occupational North Indian suburban population. Hum. Exp. Toxicol. 2013, 32, 1213–1227. [Google Scholar] [CrossRef] [PubMed]
- Orhan, H.; Evelo, C.T.; Sahin, G. Erythrocyte antioxidant defense response against cigarette smoking in humans--the glutathione S-transferase vulnerability. J. Biochem. Mol. Toxicol. 2005, 19, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Evelo, C.T.; Henderson, P.T. Biological effect monitoring. Arch. Toxicol. Suppl. 1992, 15, 268–277. [Google Scholar] [PubMed]
- Mecdad, A.; Ahmed, M.; ElHalwagy, M.; Afify, M. A Study on Oxidative Stress Biomarkers and Immunomodulatory Effects of Pesticides in Pesticide-Sprayers. Egypt. J. Forensic Sci. 2011, 1, 93–98. [Google Scholar] [CrossRef]
- Hegazi, M.M.; Attia, Z.I.; Ashour, O.A. Oxidative stress and antioxidant enzymes in liver and white muscle of Nile tilapia juveniles in chronic ammonia exposure. Aquat. Toxicol. 2010, 99, 118–125. [Google Scholar] [CrossRef]
- Vos, R.M.; Van Bladeren, P.J. Glutathione S-transferases in relation to their role in the biotransformation of xenobiotics. Chem. Biol. Interact. 1990, 75, 241–265. [Google Scholar] [CrossRef]
- Boyland, E.; Chasseaud, L.F. The role of glutathione and glutathione S-transferases in mercapturic acid biosynthesis. Adv. Enzymol. Relat. Areas Mol. Biol. 1969, 32, 173–219. [Google Scholar]
- Ketley, J.N.; Habig, W.H.; Jakoby, W.B. Binding of nonsubstrate ligands to the glutathione S-transferases. J. Biol. Chem. 1975, 250, 8670–8673. [Google Scholar]
- Awasthi, Y.C.; Garg, H.S.; Dao, D.D.; Partridge, C.A.; Srivastava, S.K. Enzymatic conjugation of erythrocyte glutathione with 1-chloro-2,4-dinitrobenzene: The fate of glutathione conjugate in erythrocytes and the effect of glutathione depletion on hemoglobin. Blood 1981, 58, 733–738. [Google Scholar]
- Hayes, J.D.; Pulford, D.J. The glutathione S-transferase supergene family: Regulation of GST and the contribution of the isoenzymes to cancer chemoprotection and drug resistance. Crit. Rev. Biochem. Mol. Biol. 1995, 30, 445–600. [Google Scholar] [CrossRef]
- Vodela, J.K.; Dalvi, R.R. Erythrocyte glutathione-S-transferase activity in animal species. Vet. Hum. Toxicol. 1997, 39, 9–11. [Google Scholar] [PubMed]
- Bocedi, A.; Fabrini, R.; Lai, O.; Alfieri, L.; Roncoroni, C.; Noce, A.; Pedersen, J.Z.; Ricci, G. Erythrocyte glutathione transferase: A general probe for chemical contaminations in mammals. Cell Death Discov. 2016, 2, 16029. [Google Scholar] [CrossRef] [PubMed]
- Turkan, F.; Huyut, Z.; Atalar, M.N. The toxicological impact of some avermectins on human erythrocytes glutathione S-transferase enzyme. J. Biochem. Mol. Toxicol. 2018, 32, e22205. [Google Scholar] [CrossRef] [PubMed]
- Kose, L.P.; Gulcin, I.; Ozdemir, H.; Atasever, A.; Alwasel, S.H.; Supuran, C.T. The effects of some avermectins on bovine carbonic anhydrase enzyme. J. Enzym. Inhib. Med. Chem. 2016, 31, 773–778. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Allele | Alterations in Gene | Amino Acids Affected |
|---|---|---|
| GSTP1 * A | A313, C341, C555 | Ile105, Ala114, Ser185 |
| GSTP1 * B | G313, C341, T555 | Val105, Ala114, Ser185 |
| GSTP1 * C | G313, T341, T555 | Val105, Val114, Ser185 |
| GSTP1 * D | A313, T341 | Ile105, Val114 |
| Selected Areas in the Frosinone District | Territorial Extension (Km2) | Geographic Features |
|---|---|---|
| Area 1 | 25 | Nearby confluence of Sacco and Liri Rivers |
| Area 2 | 10 | Close to Liri River |
| Area 3 | 90 | After confluence of Sacco and Liri Rivers |
| Area 4 | 40 | Near the Sacco River—Presence of industrial site |
| Area 5 | 40 | Liri River flows through the area—Presence of regularized landfill and compost sites |
| Area 6 | 30 | Close to an important industrial site |
| Area 7 | 60 | Sacco River flows through the area |
| Area 8 | 40 | Close to Sacco and Liri Rivers—Presence of incineration plant |
| Km | kcat (s−1) | ||
|---|---|---|---|
| GSH (mM) | CDNB (mM) | ||
| Homo sapiens | 0.11 ± 0.01 | 1.0 ± 0.1 | 79 ± 5 |
| Bos taurus | 0.12 ± 0.02 | 0.8 ± 0.2 | 83 ± 7 |
| Capra hircus | 0.14 ± 0.02 | 0.9 ± 0.1 | 85 ± 6 |
| Ovis aries | 0.10 ± 0.01 | 0.8 ± 0.2 | 77 ± 8 |
| Equus caballus | 0.10 ± 0.02 | 0.8 ± 0.2 | 82 ± 6 |
| Sus scrofa | 0.10 ± 0.02 | 0.9 ± 0.1 | 75 ± 7 |
| Kidney Disease | |
| Chronic Kidney Disease | [33,34,36,38,39,41,42,43,44,45,46] |
| Kidney Transplant | [49] |
| Neurodegenerative Disease and Psychiatric Disorder | |
| Parkinson’s Disease | [55,56,57,58,59] |
| Alzheimer’s Disease | [55,60,61,62] |
| Epilepsy | [64,65] |
| Amyotrophic lateral sclerosis | [55,67,68] |
| Schizophrenia | [70,73,74,75,76] |
| Autism | [55,78] |
| Autoimmune Disease | |
| Scleroderma | [84] |
| Others (systemic lupus erithematosus, rheumatoid arthritis, multiple sclerosis) | [86,87,88] |
| Oxidative Stress | |
| Oxidative Stress | [94,96,104] |
| Cancer | |
| Blood | [117,118,119,120,121] |
| Brain | [65,122,123] |
| Breast | [120,124,125] |
| Cervix | [126,128] |
| Ovarian | [127,128] |
| Endometrial | [129] |
| Prostate | [120,131] |
| Urinary bladder | [132,133,134] |
| Colorectal | [135,136,137] |
| Gastric | [115,139,140,141,142] |
| Esophageal | [143,144,145] |
| Pancreatic | [146,147] |
| Lung | [148,149,150] |
| Skin | [152,153,154] |
| Bone | [108,156,157,158] |
| Liver Disease | |
| Hepatocellular carcinoma | [159,164,165,166,167,168,169,170] |
| Liver fibrosis | [160,161,162] |
| Hyperbilirubinemia | [33] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocedi, A.; Noce, A.; Marrone, G.; Noce, G.; Cattani, G.; Gambardella, G.; Di Lauro, M.; Di Daniele, N.; Ricci, G. Glutathione Transferase P1-1 an Enzyme Useful in Biomedicine and as Biomarker in Clinical Practice and in Environmental Pollution. Nutrients 2019, 11, 1741. https://doi.org/10.3390/nu11081741
Bocedi A, Noce A, Marrone G, Noce G, Cattani G, Gambardella G, Di Lauro M, Di Daniele N, Ricci G. Glutathione Transferase P1-1 an Enzyme Useful in Biomedicine and as Biomarker in Clinical Practice and in Environmental Pollution. Nutrients. 2019; 11(8):1741. https://doi.org/10.3390/nu11081741
Chicago/Turabian StyleBocedi, Alessio, Annalisa Noce, Giulia Marrone, Gianluca Noce, Giada Cattani, Giorgia Gambardella, Manuela Di Lauro, Nicola Di Daniele, and Giorgio Ricci. 2019. "Glutathione Transferase P1-1 an Enzyme Useful in Biomedicine and as Biomarker in Clinical Practice and in Environmental Pollution" Nutrients 11, no. 8: 1741. https://doi.org/10.3390/nu11081741
APA StyleBocedi, A., Noce, A., Marrone, G., Noce, G., Cattani, G., Gambardella, G., Di Lauro, M., Di Daniele, N., & Ricci, G. (2019). Glutathione Transferase P1-1 an Enzyme Useful in Biomedicine and as Biomarker in Clinical Practice and in Environmental Pollution. Nutrients, 11(8), 1741. https://doi.org/10.3390/nu11081741