Linseed Components Are More Effective Than Whole Linseed in Reversing Diet-Induced Metabolic Syndrome in Rats



Abstract
1. Introduction
2. Materials and Methods
2.1. Rats and Diet
2.2. Measurements in Live Rats
2.3. Measurements after Euthanasia
2.4. Statistical Analysis
3. Results
3.1. Dietary Intakes
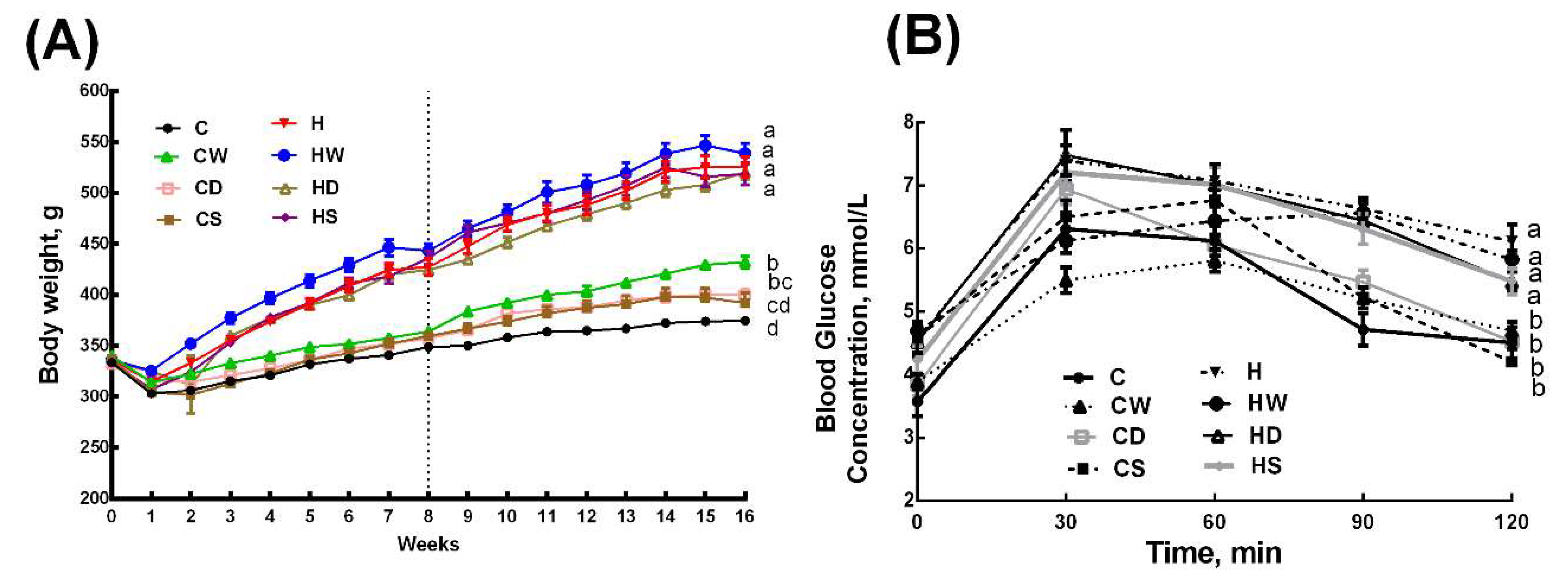
3.2. Body Composition and Organ Weights
3.3. Metabolic Parameters
3.4. Cardiovascular, Liver, and Gut Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goyal, A.; Sharma, V.; Upadhyay, N.; Gill, S.; Sihag, M. Flax and flaxseed oil: An ancient medicine & modern functional food. J. Food Sci. Technol. 2014, 51, 1633–1653. [Google Scholar] [CrossRef]
- Shim, Y.Y.; Gui, B.; Arnison, P.G.; Wang, Y.; Reaney, M.J.T. Flaxseed (Linum usitatissimum L.) bioactive compounds and peptide nomenclature: A review. Trends Food Sci. Technol. 2014, 38, 5–20. [Google Scholar] [CrossRef]
- Parikh, M.; Netticadan, T.; Pierce, G.N. Flaxseed: Its bioactive components and their cardiovascular benefits. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H146–H159. [Google Scholar] [CrossRef]
- Shafie, S.R.; Poudyal, H.; Panchal, S.K.; Brown, L. Linseed as a functional food for the management of obesity. In Omega-3 Fatty Acids: Keys to Nutritional Health; Hegde, M.V., Zanwar, A.A., Adekar, S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 173–187. [Google Scholar]
- Imran, M.; Ahmad, N.; Anjum, F.M.; Khan, M.K.; Mushtaq, Z.; Nadeem, M.; Hussain, S. Potential protective properties of flax lignan secoisolariciresinol diglucoside. Nutr. J. 2015, 14, 71. [Google Scholar] [CrossRef]
- Parikh, M.; Pierce, G.N. Dietary flaxseed: What we know and don’t know about its effects on cardiovascular disease. Can. J. Physiol. Pharmacol. 2018, 97, 75–81. [Google Scholar] [CrossRef]
- Oomah, B.D.; Mazza, G. Effect of dehulling on chemical composition and physical properties of flaxseed. LWT—Food Sci. Technol. 1997, 30, 135–140. [Google Scholar] [CrossRef]
- Kuijsten, A.; Arts, I.C.W.; van’t Veer, P.; Hollman, P.C.H. The relative bioavailability of enterolignans in humans is enhanced by milling and crushing of flaxseed. J. Nutr. 2005, 135, 2812–2816. [Google Scholar] [CrossRef]
- Waszkowiak, K.; Gliszczynska-Swiglo, A.; Barthet, V.; Skrety, J. Effect of extraction method on the phenolic and cyanogenic glucoside profile of flaxseed extracts and their antioxidant capacity. J. Am. Oil Chem. Soc. 2015, 92, 1609–1619. [Google Scholar] [CrossRef]
- Kajla, P.; Sharma, A.; Sood, D.R. Flaxseed-a potential functional food source. J. Food Sci. Technol. 2015, 52, 1857–1871. [Google Scholar] [CrossRef]
- Austria, J.A.; Richard, M.N.; Chahine, M.N.; Edel, A.L.; Malcolmson, L.J.; Dupasquier, C.M.; Pierce, G.N. Bioavailability of a-linolenic acid in subjects after ingestion of three different forms of flaxseed. J. Am. Coll. Nutr. 2008, 27, 214–221. [Google Scholar] [CrossRef]
- Mukker, J.K.; Singh, R.S.; Muir, A.D.; Krol, E.S.; Alcorn, J. Comparative pharmacokinetics of purified flaxseed and associated mammalian lignans in male Wistar rats. Br. J. Nutr. 2015, 113, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Panchal, S.K.; Poudyal, H.; Iyer, A.; Nazer, R.; Alam, M.A.; Diwan, V.; Kauter, K.; Sernia, C.; Campbell, F.; Ward, L.; et al. High-carbohydrate, high-fat diet-induced metabolic syndrome and cardiovascular remodeling in rats. J. Cardiovasc. Pharmacol. 2011, 57, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Kumar, S.A.; Iyer, A.; Waanders, J.; Ward, L.C.; Brown, L. Responses to oleic, linoleic and a-linolenic acids in high-carbohydrate, high-fat diet-induced metabolic syndrome in rats. J. Nutr. Biochem. 2013, 24, 1381–1392. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, H.; Panchal, S.K.; Waanders, J.; Ward, L.; Brown, L. Lipid redistribution by a-linolenic acid-rich chia seed inhibits stearoyl-CoA desaturase-1 and induces cardiac and hepatic protection in diet-induced obese rats. J. Nutr. Biochem. 2012, 23, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Khattab, R.Y.; Zeitoun, M.A. Quality evaluation of flaxseed oil obtained by different extraction techniques. LWT—Food Sci. Technol. 2013, 53, 338–345. [Google Scholar] [CrossRef]
- Ghattamaneni, N.K.R.; Panchal, S.K.; Brown, L. An improved rat model for chronic inflammatory bowel disease. Pharmacol. Rep. 2018, 71, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Poudyal, H.; Panchal, S.K. Functional foods as potential therapeutic options for metabolic syndrome. Obes. Rev. 2015, 16, 914–941. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi-Sartang, M.; Mazloom, Z.; Raeisi-Dehkordi, H.; Barati-Boldaji, R.; Bellissimo, N.; Totosy de Zepetnek, J.O. The effect of flaxseed supplementation on body weight and body composition: A systematic review and meta-analysis of 45 randomized placebo-controlled trials. Obes. Rev. 2017, 18, 1096–1107. [Google Scholar] [CrossRef]
- Ursoniu, S.; Sahebkar, A.; Andrica, F.; Serban, C.; Banach, M. Effects of flaxseed supplements on blood pressure: A systematic review and meta-analysis of controlled clinical trial. Clin. Nutr. 2016, 35, 615–625. [Google Scholar] [CrossRef]
- Da Costa, C.A.; da Silva, P.C.; Ribeiro, D.C.; Pereira, A.D.; dos Santos Ade, S.; de Abreu, M.D.; Pessoa, L.R.; Boueri, B.F.; Pessanha, C.R.; do Nascimento-Saba, C.C.; et al. Effects of diet containing flaxseed flour (Linum usitatissimum) on body adiposity and bone health in young male rats. Food Funct. 2016, 7, 698–703. [Google Scholar] [CrossRef]
- Kristensen, M.; Knudsen, K.E.; Jorgensen, H.; Oomah, D.; Bugel, S.; Toubro, S.; Tetens, I.; Astrup, A. Linseed dietary fibers reduce apparent digestibility of energy and fat and weight gain in growing rats. Nutrients 2013, 5, 3287–3298. [Google Scholar] [CrossRef] [PubMed]
- Sammour, R.H. Proteins of linseed (Linum usitatissimum L.), extraction and characterization by electrophoresis. Bot. Bull. Acad. Sin. 1999, 40, 121–126. [Google Scholar]
- Chung, M.W.Y.; Lei, B.; Li-Chan, E.C.Y. Isolation and structural characterization of the major protein fraction from NorMan flaxseed (Linum usitatissimum L.). Food Chem. 2005, 90, 271–279. [Google Scholar] [CrossRef]
- Borsheim, E.; Bui, Q.U.; Tissier, S.; Kobayashi, H.; Ferrando, A.A.; Wolfe, R.R. Effect of amino acid supplementation on muscle mass, strength and physical function in elderly. Clin. Nutr. 2008, 27, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Pahlavani, N.; Entezari, M.H.; Nasiri, M.; Miri, A.; Rezaie, M.; Bagheri-Bidakhavidi, M.; Sadeghi, O. The effect of L-arginine supplementation on body composition and performance in male athletes: A double-blinded randomized clinical trial. Eur. J. Clin. Nutr. 2017, 71, 544–548. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Kauter, K.; Withers, K.; Sernia, C.; Brown, L. Chronic L-arginine treatment improves metabolic, cardiovascular and liver complications in diet-induced obesity in rats. Food Funct. 2013, 4, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Fukumitsu, S.; Aida, K.; Ueno, N.; Ozawa, S.; Takahashi, Y.; Kobori, M. Flaxseed lignan attenuates high-fat diet-induced fat accumulation and induces adiponectin expression in mice. Br. J. Nutr. 2008, 100, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Park, J.; Kim, H.L.; Jung, Y.; Youn, D.H.; Lim, S.; Song, G.; Park, H.; Jin, J.S.; Kwak, H.J.; et al. Secoisolariciresinol diglucoside inhibits adipogenesis through the AMPK pathway. Eur. J. Pharmacol. 2018, 820, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Biasiotto, G.; Zanella, I.; Predolini, F.; Archetti, I.; Cadei, M.; Monti, E.; Luzzani, M.; Pacchetti, B.; Mozzoni, P.; Andreoli, R.; et al. 7-Hydroxymatairesinol improves body weight, fat and sugar metabolism in C57BJ/6 mice on a high-fat diet. Br. J. Nutr. 2018, 120, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Talom, R.T.; Judd, S.A.; McIntosh, D.D.; McNeill, J.R. High flaxseed (linseed) diet restores endothelial function in the mesenteric arterial bed of spontaneously hypertensive rats. Life Sci. 1999, 64, 1415–1425. [Google Scholar] [CrossRef]
- Al-Bishri, W.M. Favorable effects of flaxseed supplemented diet on liver and kidney functions in hypertensive Wistar rats. J. Oleo Sci. 2013, 62, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, Q.; Zhang, Q.; Gu, A.; Jiang, Z.Y. Urinary enterolactone is associated with obesity and metabolic alteration in men in the US National Health and Nutrition Examination Survey 2001-10. Br. J. Nutr. 2015, 113, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Pilar, B.; Gullich, A.; Oliveira, P.; Stroher, D.; Piccoli, J.; Manfredini, V. Protective role of flaxseed oil and flaxseed lignan secoisolariciresinol diglucoside against oxidative stress in rats with metabolic syndrome. J. Food Sci. 2017, 82, 3029–3036. [Google Scholar] [CrossRef] [PubMed]
- Sawant, S.H.; Bodhankar, S.L. Flax lignan concentrate reverses alterations in blood pressure, left ventricular functions, lipid profile and antioxidant status in DOCA-salt induced renal hypertension in rats. Ren. Fail. 2016, 38, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.; Velasquez, M.T. Potential effects of lignan-enriched flaxseed powder on bodyweight, visceral fat, lipid profile, and blood pressure in rats. Fitoterapia 2012, 83, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Caligiuri, S.P.; Rodriguez-Leyva, D.; Aukema, H.M.; Ravandi, A.; Weighell, W.; Guzman, R.; Pierce, G.N. Dietary flaxseed reduces central aortic blood pressure without cardiac involvement but through changes in plasma oxylipins. Hypertension 2016, 68, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Opyd, P.M.; Jurgonski, A.; Juskiewicz, J.; Fotschki, B.; Koza, J. Comparative effects of native and defatted flaxseeds on intestinal enzyme activity and lipid metabolism in rats fed a high-fat diet containing cholic acid. Nutrients 2018, 10, 1181. [Google Scholar] [CrossRef]
- Torkan, M.; Entezari, M.H.; Siavash, M. Effect of flaxseed on blood lipid level in hyperlipidemic patients. Rev. Recent Clin. Trials 2015, 10, 61–67. [Google Scholar] [CrossRef]
- Ren, G.Y.; Chen, C.Y.; Chen, G.C.; Chen, W.G.; Pan, A.; Pan, C.W.; Zhang, Y.H.; Qin, L.Q.; Chen, L.H. Effect of flaxseed intervention on inflammatory marker C-reactive protein: A systematic review and meta-analysis of randomized controlled trials. Nutrients 2016, 8, 136. [Google Scholar] [CrossRef]
- Kailash, P.; Arti, D. Flaxseed and diabetes. Curr. Pharm. Des. 2016, 22, 141–144. [Google Scholar] [CrossRef]
- Dyck, D.J.; Heigenhauser, G.J.; Bruce, C.R. The role of adipokines as regulators of skeletal muscle fatty acid metabolism and insulin sensitivity. Acta Physiol. 2006, 186, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Mridula, D.; Rehal, J.; Barnwal, P. Flaxseed: A potential source of food, feed and fiber. Crit. Rev. Food Sci. Nutr. 2011, 51, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Requena, T.; Martínez-Cuesta, M.C.; Peláez, C. Diet and microbiota linked in health and disease. Food Funct. 2018, 9, 688–704. [Google Scholar] [CrossRef] [PubMed]
- Pulkrabek, M.; Rhee, Y.; Gibbs, P.; Hall, C. Flaxseed- and buckwheat-supplemented diets altered Enterobacteriaceae diversity and prevalence in the cecum and feces of obese mice. J. Diet. Suppl. 2017, 14, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Lagkouvardos, I.; Kläring, K.; Heinzmann, S.S.; Platz, S.; Scholz, B.; Engel, K.-H.; Schmitt-Kopplin, P.; Haller, D.; Rohn, S.; Skurk, T.; et al. Gut metabolites and bacterial community networks during a pilot intervention study with flaxseeds in healthy adult men. Mol. Nutr. Food Res. 2015, 59, 1614–1628. [Google Scholar] [CrossRef] [PubMed]
- Birchenough, G.M.; Johansson, M.E.; Gustafsson, J.K.; Bergstrom, J.H.; Hansson, G.C. New developments in goblet cell mucus secretion and function. Mucosal Immunol. 2015, 8, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, M.; Couedelo, L.; Meugnier, E.; Plaisancie, P.; Letisse, M.; Benoit, B.; Gabert, L.; Penhoat, A.; Durand, A.; Pineau, G.; et al. Dietary emulsifiers from milk and soybean differently impact adiposity and inflammation in association with modulation of colonic goblet cells in high-fat fed mice. Mol. Nutr. Food Res. 2016, 60, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Power, K.A.; Lepp, D.; Zarepoor, L.; Monk, J.M.; Wu, W.; Tsao, R.; Liu, R. Dietary flaxseed modulates the colonic microenvironment in healthy C57Bl/6 male mice which may alter susceptibility to gut-associated diseases. J. Nutr. Biochem. 2016, 28, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, H.; Ito, H.; Sugiyama, K.; Kiriyama, S.; Morita, T. Dietary indigestible components exert different regional effects on luminal mucin secretion through their bulk-forming property and fermentability. Biosci. Biotechnol. Biochem. 2006, 70, 1188–1194. [Google Scholar] [CrossRef]
- Cressey, P.; Reeve, J. Metabolism of cyanogenic glycosides: A review. Food Chem. Toxicol. 2019, 125, 225–232. [Google Scholar] [CrossRef]
- Schulz, V.; Loffler, A.; Gheorghiu, T. Resorption of hydrocyanic acid from linseed. Leber Magen Darm 1983, 13, 10–14. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | C | CW | CD | CS | H | HW | HD | HS | p Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet | Treatment | Interaction | |||||||||
| Food intake, g/day | 35.0 ± 0.6 a | 34.6 ± 1.2 a | 33.9 ± 1.2 a | 35.5 ± 0.8 a | 25.2 ± 0.6 b | 23.4 ± 0.6 b | 24.4 ± 0.4 b | 26.3 ± 0.4 b | <0.0001 | 0.06 | 0.59 |
| Water intake, mL/day | 28.7 ± 1.8 a | 30.7 ± 2.1 a | 32.7 ± 0.9 a | 30.4 ± 2.7 a | 25.6 ± 1.0 b | 26.9 ± 0.4 ab | 25.7 ± 1.1 b | 25.2 ± 1.0 b | <0.0001 | 0.54 | 0.61 |
| Energy intake, kJ/day | 392 ± 7 b | 433 ± 14 b | 397 ± 14 b | 420 ± 13 b | 552 ± 13 a | 561 ± 14 a | 542 ± 9 a | 555 ± 8 a | <0.0001 | 0.11 | 0.37 |
| Body weight gained (week 8–16), % | 7.4 ± 1.0 d | 18.2 ± 0.8 b | 13.2 ± 1.6 c | 9.1 ± 1.7 d | 23.3 ± 0.9 a | 23.4 ± 0.6 a | 22.7 ± 1.4 a | 18.9 ± 1.3 b | <0.0001 | <0.0001 | 0.0006 |
| Final body weight (week 16) | 375 ± 4 a | 432 ± 6 b | 400 ± 5 a | 392 ± 10 a | 526 ± 10 c | 539 ± 10 c | 521 ± 8 c | 519 ± 11 c | < 0.0001 | 0.0004 | 0.06 |
| Feed conversion efficiency, % | 1.8 ± 0.2 c | 4.9 ± 0.3 b | 3.2 ± 0.4 c | 2.6 ± 0.5 c | 9.6 ± 0.6 a | 9.2 ± 0.6 a | 9.0 ± 0.6 a | 8.1 ± 0.7 a | <0.0001 | 0.004 | 0.005 |
| Bone mineral content, g | 12.0 ± 0.4 c | 12.1 ± 0.2 c | 11.9 ± 0.2 c | 13.2 ± 0.6 c | 16.5 ± 0.4 a | 17.3 ± 0.7 a | 15.3 ± 0.6 b | 16.8 ± 0.4 a | <0.0001 | 0.024 | 0.20 |
| Total fat mass, g | 98 ± 16 c | 103 ± 7 c | 77 ± 6 c | 122 ± 16 c | 256 ± 21 a | 253 ± 21 a | 189 ± 22 b | 227 ± 7 ab | <0.0001 | 0.014 | 0.25 |
| Total lean mass, g | 276 ± 14 ab | 317 ± 7 a | 309 ± 5 a | 267 ± 9 b | 268 ± 14 b | 277 ± 11 ab | 312 ± 15 a | 246 ± 7 b | 0.035 | <0.0001 | 0.24 |
| Abdominal circumference, cm | 18.4 ± 0.2 c | 19.1 ± 0.2 c | 19.1 ± 0.1 c | 18.9 ± 0.2 c | 23.0 ± 0.2 a | 23.6 ± 0.4 a | 21.2 ± 0.2 b | 22.7 ± 0.2 a | <0.0001 | 0.016 | 0.19 |
| Visceral adiposity index, % | 4.9 ± 0.4 d | 4.7 ± 0.3 d | 4.2 ± 0.1 d | 4.8 ± 0.6 d | 9.6 ± 0.7 ab | 10.4 ± 0.9 a | 7.3 ± 0.5 c | 8.5 ± 0.4 bc | <0.0001 | 0.007 | 0.09 |
| Retroperitoneal fat, mg/mm * | 189 ± 18 c | 179 ± 18 c | 151 ± 20 c | 194 ± 31 c | 531 ± 41 a | 554 ± 59 a | 407 ± 31 b | 469 ± 31 ab | <0.0001 | 0.55 | 0.34 |
| Epididymal fat, mg/mm * | 89 ± 10 d | 98 ± 8 d | 74 ± 5 d | 112 ± 20 cd | 259 ± 17 a | 278 ± 28 a | 154 ± 16 c | 211 ± 11 b | <0.0001 | 0.29 | 0.037 |
| Omental fat, mg/mm * | 114 ± 15 c | 124 ± 9 c | 102 ± 7 c | 102 ± 13 c | 240 ± 25 ab | 280 ± 18 a | 198 ± 14 b | 207 ± 8 b | <0.0001 | 0.013 | 0.26 |
| Total abdominal fat, mg/mm * | 392 ± 39 d | 401 ± 35 d | 308 ± 36 d | 408 ± 63 d | 1031 ± 79 ab | 1113 ± 105 a | 683 ± 92 c | 887 ± 45 b | <0.0001 | 0.23 | 0.18 |
| Liver, mg/mm * | 201 ± 5 b | 213 ± 12 b | 216 ± 7 b | 213 ± 8 b | 327 ± 12 a | 337 ± 15 a | 310 ± 8 a | 306 ± 9 a | <0.0001 | 0.32 | 0.22 |
| Variables | C | CW | CD | CS | H | HW | HD | HS | p Value | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Diet | Treatment | Interaction | |||||||||
| Area under the curve, mmol/L×min | 636 ± 19 bc | 625 ± 14 c | 680 ± 14 b | 687 ± 20 b | 794 ± 13 a | 735 ± 17 ab | 770 ± 17 a | 765 ± 25 a | <0.0001 | 0.037 | 0.09 |
| Plasma total cholesterol, mmol/L | 1.5 ± 0.05 a | 1.3 ± 0.10 a | 1.4 ± 0.10 a | 1.6 ± 0.06 a | 1.6 ± 0.04 a | 1.5 ± 0.10 a | 1.6 ± 0.10 a | 1.6 ± 0.06 a | 0.09 | 0.023 | 0.39 |
| Plasma nonesterified fatty acids, mmol/L | 0.9 ± 0.2 b | 1.5 ± 0.2 b | 1.5 ± 0.2 b | 1.4 ± 0.2 b | 4.3 ± 0.6 a | 4.0 ± 0.3 a | 3.7 ± 0.4 a | 3.7 ± 0.5 a | <0.0001 | 0.86 | 0.31 |
| Plasma triglycerides, mmol/L | 0.4 ± 0.06 b | 0.4 ± 0.10 b | 0.5 ± 0.01 b | 0.4 ± 0.06 b | 1.6 ± 0.30 a | 1.5 ± 0.20 a | 1.3 ± 0.20 a | 1.3 ± 0.20 a | <0.0001 | 0.68 | 0.68 |
| Plasma insulin, µmol/L | 1.3 ± 0.01 e | 2.9 ± 0.05 c | 1.7 ± 0.05 d | 1.7 ± 0.01 d | 7.7 ± 0.09 a | 5.9 ± 0.12 b | 5.8 ± 0.04 b | 6.0 ± 0.11 b | <0.0001 | <0.0001 | <0.0001 |
| Plasma leptin, µmol/L | 3.2 ± 0.40 d | 4.2 ± 0.06 d | 3.9 ± 0.03 d | 2.1 ± 0.62 d | 12.3 ± 1.54 a | 13.1 ± 0.03 a | 10.2 ± 0.08 b | 6.3 ± 0.97 c | <0.0001 | <0.0001 | 0.005 |
| Heart, mg/mm * | 22.0 ± 1.0 c | 25.4 ± 0.7 bc | 25.3 ± 0.9 bc | 21.5 ± 0.4 c | 26.8 ± 1.1 b | 31.5 ± 2.1 a | 26.9 ± 0.9 b | 23.6 ± 0.9 bc | <0.0001 | <0.0001 | 0.18 |
| Left ventricle + septum, mg/mm * | 18.1 ± 0.7 b | 21.6 ± 0.6 b | 20.8 ± 0.8 b | 17.2 ± 0.3 b | 20.8 ± 0.8 b | 27.1 ± 1.8 a | 22.3 ± 0.8 b | 18.5 ± 0.8 b | 0.0002 | <0.0001 | 0.09 |
| Right ventricle, mg/mm * | 4.0 ± 0.4 | 4.5 ± 0.4 | 4.5 ± 0.3 | 4.4 ± 0.2 | 6.3 ± 1.7 | 5.8 ± 0.4 | 4.7 ± 0.2 | 5.0 ± 0.1 | 0.03 | 0.79 | 0.53 |
| Systolic blood pressure, mmHg | 128 ± 3 bc | 129 ± 2 bc | 120 ± 3 c | 133 ± 2 b | 148 ± 2 a | 134 ± 2 b | 129 ± 5 bc | 133 ± 3 b | <0.0001 | 0.026 | 0.0004 |
| Diastolic stiffness constant (κ) | 23.0 ± 0.4 bc | 24.2 ± 0.7 abc | 21.9 ± 0.3 c | 23.0 ± 0.8 bc | 26.6 ± 0.9 a | 25.1 ± 0.5 ab | 23.7 ± 1.3 bc | 22.0 ± 0.5 c | 0.034 | 0.001 | 0.004 |
| Plasma alanine transaminase, U/L | 26.8 ± 2.5 b | 30.9 ± 3.8 b | 28.3 ± 2.0 b | 23.2 ± 1.6 b | 33.7 ± 1.5 a | 27.8 ± 2.5 ab | 33.8 ± 1.5 a | 33.6 ± 1.7 a | 0.017 | 0.74 | 0.017 |
| Plasma aspartate transaminase, U/L | 64.6 ± 4.6 a | 66.4 ± 5.9 a | 66.3 ± 2.8 a | 61.6 ± 1.8 a | 68.8 ± 2.6 a | 61.6 ± 3.7 a | 67.5 ± 2.9 a | 69.6 ± 6.8 a | 0.52 | 0.84 | 0.38 |
| Left ventricle collagen deposition, % | 6.51 ± 1.0 b | 6.57 ± 1.1 b | 5.29 ± 0.2 b | 4.96 ± 0.5 b | 16.1 ± 3.7 a | 13.9 ± 1.3 a | 8.2 ± 0.5 b | 8.3 ± 1.0 b | <0.0001 | <0.0001 | <0.0001 |
| Left ventricle inflammatory cells, n | 9.0 ± 2.5 b | 7.5 ± 1.2 b | 10.5 ± 1.6 b | 12.3 ± 1.1 b | 60.0 ± 4.0 a | 10.8 ± 1.4 b | 12.3 ± 1.1 b | 10.8 ± 1.1 b | <0.0001 | <0.0001 | <0.0001 |
| Liver fat vacuoles, n | 4.0 ± 0.6 c | 10.8 ± 1.6 c | 4.0 ± 1.2c | 3.5 ± 1.0 c | 75.0 ± 4.6 a | 39.7 ± 4.4 b | 3.5 ± 1.0 c | 1.5 ± 0.9 c | <0.0001 | <0.0001 | <0.0001 |
| Ileum goblet cells, n | 91.5 ± 6.0 c | 82.3 ± 6.1 d | 107.3 ± 2.1 abc | 100.3 ± 2.5 cd | 124.3 ± 3.4 ab | 118.3 ± 8.6 ab | 94.8 ± 3.4 bc | 110.8 ± 4.4 ab | 0.0005 | 0.3224 | 0.0004 |
| Colon goblet cells, n | 97.5 ± 2.9 c | 42 ± 5.1 d | 84.5 ± 2.6 c | 90.8 ± 5.2 c | 78.8 ± 3.9 c | 135.3 ± 2.3 b | 203.8 ± 14.5 a | 131.3 ± 7.3 b | <0.0001 | <0.0001 | <0.0001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shafie, S.R.; Wanyonyi, S.; Panchal, S.K.; Brown, L. Linseed Components Are More Effective Than Whole Linseed in Reversing Diet-Induced Metabolic Syndrome in Rats. Nutrients 2019, 11, 1677. https://doi.org/10.3390/nu11071677
Shafie SR, Wanyonyi S, Panchal SK, Brown L. Linseed Components Are More Effective Than Whole Linseed in Reversing Diet-Induced Metabolic Syndrome in Rats. Nutrients. 2019; 11(7):1677. https://doi.org/10.3390/nu11071677
Chicago/Turabian StyleShafie, Siti Raihanah, Stephen Wanyonyi, Sunil K. Panchal, and Lindsay Brown. 2019. "Linseed Components Are More Effective Than Whole Linseed in Reversing Diet-Induced Metabolic Syndrome in Rats" Nutrients 11, no. 7: 1677. https://doi.org/10.3390/nu11071677
APA StyleShafie, S. R., Wanyonyi, S., Panchal, S. K., & Brown, L. (2019). Linseed Components Are More Effective Than Whole Linseed in Reversing Diet-Induced Metabolic Syndrome in Rats. Nutrients, 11(7), 1677. https://doi.org/10.3390/nu11071677