Vitamin D and Phenylbutyrate Supplementation Does Not Modulate Gut Derived Immune Activation in HIV-1

 , ,
, , 
Abstract
1. Introduction
2. Material and Method
2.1. Study Design
2.2. Patients and Clinical Samples
2.3. Intervention and Randomization
2.4. Vitamin D
2.5. LL-37, sCD14 and Targeted Metabolomics
2.6. Microbiota
2.7. Statistical Analyses
3. Results
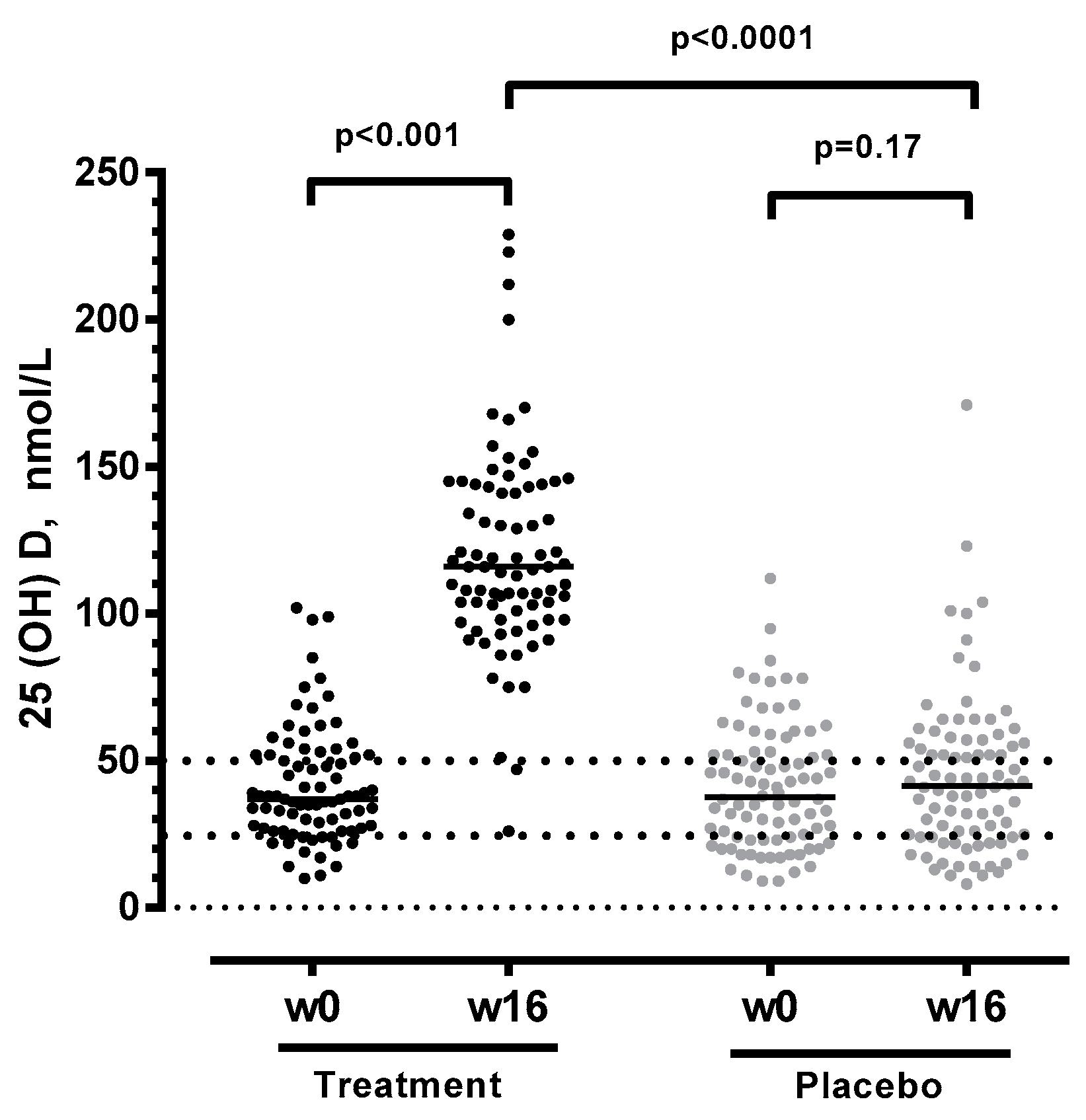
3.1. Vitamin D + PBA Supplementation Improved Vitamin D Status in ART-Naïve HIV Patients
3.2. Vitamin D + PBA Supplementation Had No Effect on Immune Activation or Microbial Metabolites in Plasma Assessed in ART-Naïve HIV Patients
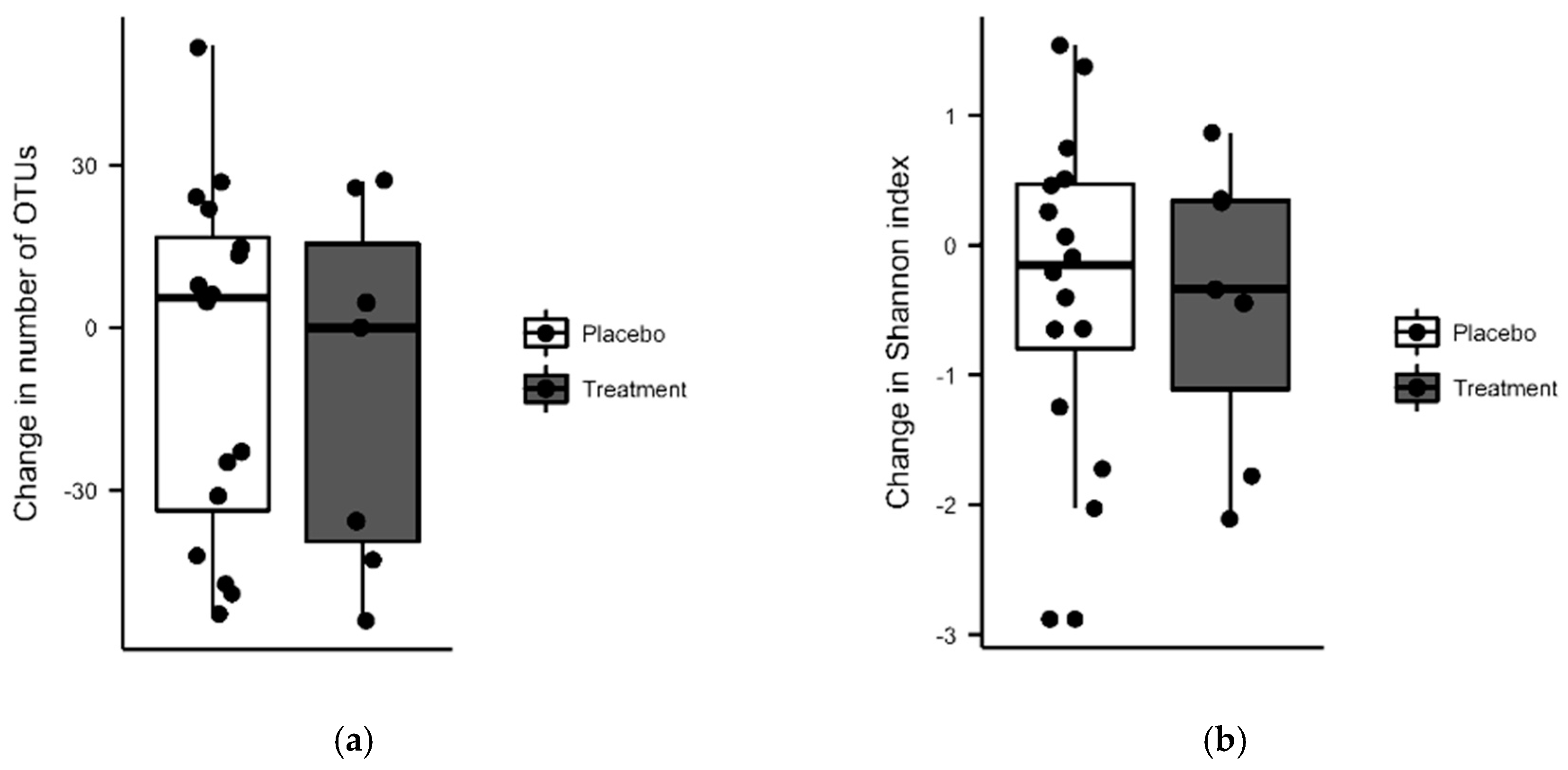
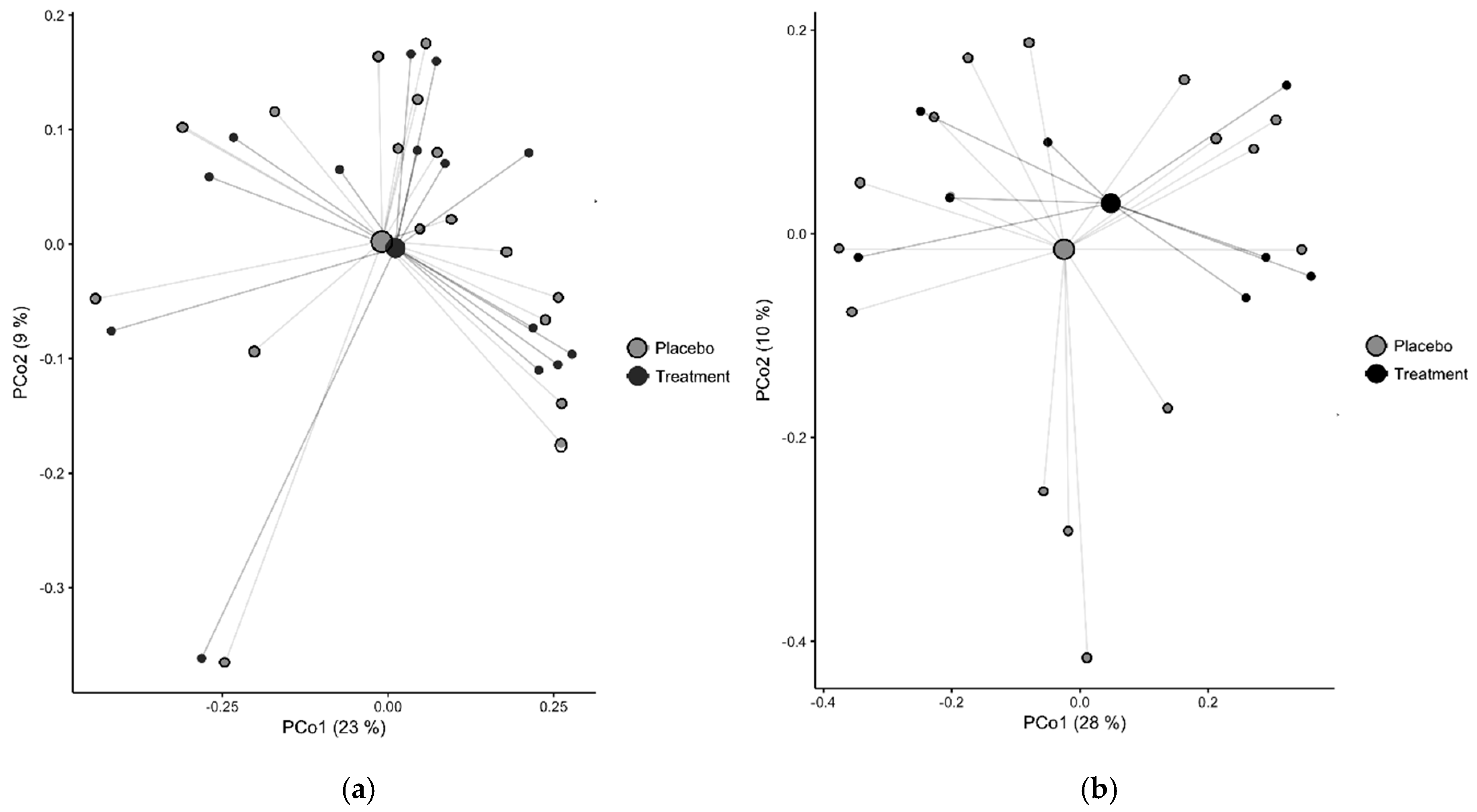
3.3. Vitamin D + PBA Supplementation Had No Effect on the Microbiota Composition of Gut Mucosa Assessed in ART-Naïve HIV Patients
3.4. Subgroup Analysis of Immune Activation and Metabolites in Vitamin D Insufficient ART-Naïve HIV Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Deeks, S.G.; Tracy, R.; Douek, D.C. Systemic effects of inflammation on health during chronic HIV infection. Immunity 2013, 39, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, J.M.; Douek, D.C. The mucosal barrier and immune activation in HIV pathogenesis. Curr. Opin. HIV AIDS 2008, 3, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic-Cvijin, I.; Dunham, R.M.; Iwai, S.; Maher, M.C.; Albright, R.G.; Broadhurst, M.J.; Hernandez, R.D.; Lederman, M.M.; Huang, Y.; Somsouk, M.; et al. Dysbiosis of the gut microbiota is associated with hiv disease progression and tryptophan catabolism. Sci. Trans. Med. 2013, 5, 193ra191. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Castellanos, J.F.; Serrano-Villar, S.; Latorre, A.; Artacho, A.; Ferrus, M.L.; Madrid, N.; Vallejo, A.; Sainz, T.; Martinez-Botas, J.; Ferrando-Martinez, S.; et al. Altered metabolism of gut microbiota contributes to chronic immune activation in HIV-infected individuals. Mucosal Immunol. 2015, 8, 760–772. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Hua, S.; Clish, C.B.; Scott, J.M.; Hanna, D.B.; Wang, T.; Haberlen, S.A.; Shah, S.J.; Glesby, M.J.; Lazar, J.M.; et al. Plasma tryptophan-kynurenine metabolites are altered in HIV infection and associated with progression of carotid artery atherosclerosis. Clin. Infect. Dis. 2018, 67, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Sandler, N.G.; Wand, H.; Roque, A.; Law, M.; Nason, M.C.; Nixon, D.E.; Pedersen, C.; Ruxrungtham, K.; Lewin, S.R.; Emery, S.; et al. Plasma levels of soluble CD14 independently predict mortality in HIV infection. J. Infect. Dis. 2011, 203, 780–790. [Google Scholar] [CrossRef]
- Schouten, J.; Wit, F.W.; Stolte, I.G.; Kootstra, N.; van der Valk, M.; Geerlings, S.G.; Prins, M.; Reiss, P. Cross-sectional comparison of the prevalence of age-associated comorbidities and their risk factors between HIV-infected and uninfected individuals. the AGEhIV Cohort Study. Clin. Infect. Dis. 2014, 59, 1787–1797. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef]
- Zhu, W.; Gregory, J.C.; Org, E.; Buffa, J.A.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M.; et al. Gut Microbial Metabolite TMAO Enhances Platelet Hyperreactivity and Thrombosis Risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef]
- Shan, Z.; Clish, C.B.; Hua, S.; Scott, J.M.; Hanna, D.B.; Burk, R.D.; Haberlen, S.A.; Shah, S.J.; Margolick, J.B.; Sears, C.L.; et al. Gut Microbial-Related Choline Metabolite Trimethylamine-N-Oxide Is Associated With Progression of Carotid Artery Atherosclerosis in HIV Infection. J. Infect. Dis. 2018, 218, 1474–1479. [Google Scholar] [CrossRef]
- Mutlu, E.A.; Keshavarzian, A.; Losurdo, J.; Swanson, G.; Siewe, B.; Forsyth, C.; French, A.; Demarais, P.; Sun, Y.; Koenig, L.; et al. A compositional look at the human gastrointestinal microbiome and immune activation parameters in HIV infected subjects. PLoS Pathog 2014, 10, e1003829. [Google Scholar] [CrossRef] [PubMed]
- Nowak, P.; Troseid, M.; Avershina, E.; Barqasho, B.; Neogi, U.; Holm, K.; Hov, J.R.; Noyan, K.; Vesterbacka, J.; Svard, J.; et al. Gut microbiota diversity predicts immune status in HIV-1 infection. AIDS 2015, 29, 2409–2418. [Google Scholar] [CrossRef]
- Chen, J.; Waddell, A.; Lin, Y.D.; Cantorna, M.T. Dysbiosis caused by vitamin D receptor deficiency confers colonization resistance to Citrobacter rodentium through modulation of innate lymphoid cells. Mucosal Immunol. 2015, 8, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Assa, A.; Vong, L.; Pinnell, L.J.; Avitzur, N.; Johnson-Henry, K.C.; Sherman, P.M. Vitamin D Deficiency Promotes Epithelial Barrier Dysfunction and Intestinal Inflammation. J. Infect. Dis. 2014, 210, 1296–1305. [Google Scholar] [CrossRef]
- Liu, W.; Chen, Y.; Golan, M.A.; Annunziata, M.L.; Du, J.; Dougherty, U.; Kong, J.; Musch, M.; Huang, Y.; Pekow, J.; et al. Intestinal epithelial vitamin D receptor signaling inhibits experimental colitis. J. Clin. Investig. 2013, 123, 3983–3996. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, H.; Wu, H.; Li, H.; Liu, L.; Guo, J.; Li, C.; Shih, D.Q.; Zhang, X. Protective role of 1,25(OH)2 vitamin D3 in the mucosal injury and epithelial barrier disruption in DSS-induced acute colitis in mice. BMC Gastroenterol. 2012, 12, 57. [Google Scholar] [CrossRef]
- Lagishetty, V.; Misharin, A.V.; Liu, N.Q.; Lisse, T.S.; Chun, R.F.; Ouyang, Y.; McLachlan, S.M.; Adams, J.S.; Hewison, M. Vitamin D deficiency in mice impairs colonic antibacterial activity and predisposes to colitis. Endocrinology 2010, 151, 2423–2432. [Google Scholar] [CrossRef]
- Del Pinto, R.; Ferri, C.; Cominelli, F. Vitamin D Axis in Inflammatory Bowel Diseases: Role, Current Uses and Future Perspectives. Int. J. Mol. Sci. 2017, 18, 2360. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef]
- Wang, T.T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting edge. 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef]
- Vandamme, D.; Landuyt, B.; Luyten, W.; Schoofs, L. A comprehensive summary of LL-37, the factotum human cathelicidin peptide. Cell. Immunol. 2012, 280, 22–35. [Google Scholar] [CrossRef]
- Campbell, G.R.; Spector, S.A. Autophagy induction by vitamin D inhibits both Mycobacterium tuberculosis and human immunodeficiency virus type 1. Autophagy 2012, 8, 1523–1525. [Google Scholar] [CrossRef]
- Wang, G.; Watson, K.M.; Buckheit, R.W., Jr. Anti-human immunodeficiency virus type 1 activities of antimicrobial peptides derived from human and bovine cathelicidins. Antimicrob. Agents Chemother. 2008, 52, 3438–3440. [Google Scholar] [CrossRef]
- Aguilar-Jimenez, W.; Zapata, W.; Rugeles, M.T. Antiviral molecules correlate with vitamin D pathway genes and are associated with natural resistance to HIV-1 infection. Microbes Infect. 2016, 18, 510–516. [Google Scholar] [CrossRef]
- Wong, J.H.; Legowska, A.; Rolka, K.; Ng, T.B.; Hui, M.; Cho, C.H.; Lam, W.W.; Au, S.W.; Gu, O.W.; Wan, D.C. Effects of cathelicidin and its fragments on three key enzymes of HIV-1. Peptides 2011, 32, 1117–1122. [Google Scholar] [CrossRef]
- Kong, J.; Zhang, Z.; Musch, M.W.; Ning, G.; Sun, J.; Hart, J.; Bissonnette, M.; Li, Y.C. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G208–G216. [Google Scholar] [CrossRef]
- Kanhere, M.; He, J.; Chassaing, B.; Ziegler, T.R.; Alvarez, J.A.; Ivie, E.A.; Hao, L.; Hanfelt, J.; Gewirtz, A.T.; Tangpricha, V. Bolus Weekly Vitamin D3 Supplementation Impacts Gut and Airway Microbiota in Adults With Cystic Fibrosis. A Double-Blind, Randomized, Placebo-Controlled Clinical Trial. J. Clin. Endocrinol. Metab. 2018, 103, 564–574. [Google Scholar] [CrossRef]
- Bashir, M.; Prietl, B.; Tauschmann, M.; Mautner, S.I.; Kump, P.K.; Treiber, G.; Wurm, P.; Gorkiewicz, G.; Hogenauer, C.; Pieber, T.R. Effects of high doses of vitamin D3 on mucosa-associated gut microbiome vary between regions of the human gastrointestinal tract. Eur J. Nutr. 2016, 55, 1479–1489. [Google Scholar] [CrossRef]
- Mansueto, P.; Seidita, A.; Vitale, G.; Gangemi, S.; Iaria, C.; Cascio, A. Vitamin D Deficiency in HIV Infection. Not Only a Bone Disorder. Biomed. Res. Int. 2015, 2015, 735615. [Google Scholar] [CrossRef]
- Ansemant, T.; Mahy, S.; Piroth, C.; Ornetti, P.; Ewing, S.; Guilland, J.C.; Croisier, D.; Duvillard, L.; Chavanet, P.; Maillefert, J.F.; et al. Severe hypovitaminosis D correlates with increased inflammatory markers in HIV infected patients. BMC Infect. Dis. 2013, 13, 7. [Google Scholar] [CrossRef]
- Legeai, C.; Vigouroux, C.; Souberbielle, J.C.; Bouchaud, O.; Boufassa, F.; Bastard, J.P.; Carlier, R.; Capeau, J.; Goujard, C.; Meyer, L.; et al. Associations between 25-hydroxyvitamin D and immunologic, metabolic, inflammatory markers in treatment-naive HIV-infected persons. the ANRS CO9 cohort study. PLoS ONE 2013, 8, e74868. [Google Scholar] [CrossRef]
- Ashenafi, S.; Amogne, W.; Kassa, E.; Gebreselassie, N.; Bekele, A.; Aseffa, G.; Getachew, M.; Aseffa, A.; Worku, A.; Hammar, U.; et al. Daily Nutritional Supplementation with Vitamin D3 and Phenylbutyrate to Treatment-Naive HIV Patients Tested in a Randomized Placebo-Controlled Trial. Nutrients 2019, 11, 133. [Google Scholar] [CrossRef]
- Mily, A.; Rekha, R.S.; Kamal, S.M.; Akhtar, E.; Sarker, P.; Rahim, Z.; Gudmundsson, G.H.; Agerberth, B.; Raqib, R. Oral intake of phenylbutyrate with or without vitamin D3 upregulates the cathelicidin LL-37 in human macrophages. a dose finding study for treatment of tuberculosis. BMC Pulm. Med. 2013, 13, 23. [Google Scholar] [CrossRef]
- Missailidis, C.; Hallqvist, J.; Qureshi, A.R.; Barany, P.; Heimburger, O.; Lindholm, B.; Stenvinkel, P.; Bergman, P. Serum Trimethylamine-N-Oxide Is Strongly Related to Renal Function and Predicts Outcome in Chronic Kidney Disease. PLoS ONE 2016, 11, e0141738. [Google Scholar] [CrossRef]
- Edgar, R.C.; Flyvbjerg, H. Error filtering, pair assembly and error correction for next-generation sequencing reads. Bioinformatics 2015, 31, 3476–3482. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur. open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project. data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12, 87. [Google Scholar] [CrossRef]
- Stevens, D.A.; Hamilton, J.R.; Johnson, N.; Kim, K.K.; Lee, J.S. Halomonas, a newly recognized human pathogen causing infections and contamination in a dialysis center. three new species. Medicine (Baltimore) 2009, 88, 244–249. [Google Scholar] [CrossRef]
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H. Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef]
- Institute of Medicine Committee. Dietary Reference Intakes for Calcium and Vitamin D; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Raftery, T.; Martineau, A.R.; Greiller, C.L.; Ghosh, S.; McNamara, D.; Bennett, K.; Meddings, J.; O’Sullivan, M. Effects of vitamin D supplementation on intestinal permeability, cathelicidin and disease markers in Crohn’s disease. Results from a randomised double-blind placebo-controlled study. United Eur. Gastroenterol. J. 2015, 3, 294–302. [Google Scholar] [CrossRef]
- Jorgensen, S.P.; Agnholt, J.; Glerup, H.; Lyhne, S.; Villadsen, G.E.; Hvas, C.L.; Bartels, L.E.; Kelsen, J.; Christensen, L.A.; Dahlerup, J.F. Clinical trial. vitamin D3 treatment in Crohn’s disease—A randomized double-blind placebo-controlled study. Aliment. Pharmacol. Ther. 2010, 32, 377–383. [Google Scholar] [CrossRef]
- Ponda, M.P.; Breslow, J.L. Vitamin D3 Repletion in Chronic Kidney Disease Stage 3. Effects on Blood Endotoxin Activity, Inflammatory Cytokines, and Intestinal Permeability. Renal. Fail. 2013, 35, 497–503. [Google Scholar] [CrossRef]
- Benguella, L.; Arbault, A.; Fillion, A.; Blot, M.; Piroth, C.; Denimal, D.; Duvillard, L.; Ornetti, P.; Chavanet, P.; Maillefert, J.F.; et al. Vitamin D supplementation, bone turnover, and inflammation in HIV-infected patients. Med. Mal. Infect. 2018, 48, 449–456. [Google Scholar] [CrossRef]
- Jorde, R.; Sneve, M.; Torjesen, P.A.; Figenschau, Y.; Goransson, L.G.; Omdal, R. No effect of supplementation with cholecalciferol on cytokines and markers of inflammation in overweight and obese subjects. Cytokine 2010, 50, 175–180. [Google Scholar] [CrossRef]
- Mousa, A.; Naderpoor, N.; Johnson, J.; Sourris, K.; de Courten, M.P.J.; Wilson, K.; Scragg, R.; Plebanski, M.; de Courten, B. Effect of vitamin D supplementation on inflammation and nuclear factor kappa-B activity in overweight/obese adults. a randomized placebo-controlled trial. Sci. Rep. 2017, 7, 15154. [Google Scholar] [CrossRef]
- Maestri, N.E.; Brusilow, S.W.; Clissold, D.B.; Bassett, S.S. Long-term treatment of girls with ornithine transcarbamylase deficiency. N. Engl. J. Med. 1996, 335, 855–859. [Google Scholar] [CrossRef]
- Camacho, L.H.; Olson, J.; Tong, W.P.; Young, C.W.; Spriggs, D.R.; Malkin, M.G. Phase I dose escalation clinical trial of phenylbutyrate sodium administered twice daily to patients with advanced solid tumors. Investig. New Drugs 2007, 25, 131–138. [Google Scholar] [CrossRef]
- Steinmann, J.; Halldorsson, S.; Agerberth, B.; Gudmundsson, G.H. Phenylbutyrate induces antimicrobial peptide expression. Antimicrob. Agents Chemother. 2009, 53, 5127–5133. [Google Scholar] [CrossRef]
- Cao, S.S.; Zimmermann, E.M.; Chuang, B.M.; Song, B.; Nwokoye, A.; Wilkinson, J.E.; Eaton, K.A.; Kaufman, R.J. The unfolded protein response and chemical chaperones reduce protein misfolding and colitis in mice. Gastroenterology 2013, 144, 989–1000. [Google Scholar] [CrossRef]
- Jellbauer, S.; Perez Lopez, A.; Behnsen, J.; Gao, N.; Nguyen, T.; Murphy, C.; Edwards, R.A.; Raffatellu, M. Beneficial Effects of Sodium Phenylbutyrate Administration during Infection with Salmonella enterica Serovar Typhimurium. Infect. Immun. 2016, 84, 2639–2652. [Google Scholar] [CrossRef]
- Aguilar-Jimenez, W.; Saulle, I.; Trabattoni, D.; Vichi, F.; Lo Caputo, S.; Mazzotta, F.; Rugeles, M.T.; Clerici, M.; Biasin, M. High Expression of Antiviral and Vitamin D Pathway Genes Are a Natural Characteristic of a Small Cohort of HIV-1-Exposed Seronegative Individuals. Front. Immunol. 2017, 8, 136. [Google Scholar] [CrossRef]
- Chandel, N.; Ayasolla, K.S.; Lan, X.; Sultana-Syed, M.; Chawla, A.; Lederman, R.; Vethantham, V.; Saleem, M.A.; Chander, P.N.; Malhotra, A.; et al. Epigenetic Modulation of Human Podocyte Vitamin D Receptor in HIV Milieu. J. Mol. Biol. 2015, 427, 3201–3215. [Google Scholar] [CrossRef]
- Gore, S.D.; Carducci, M.A. Modifying histones to tame cancer. clinical development of sodium phenylbutyrate and other histone deacetylase inhibitors. Expert Opin. Investig. Drugs 2000, 9, 2923–2934. [Google Scholar] [CrossRef]
- Shirakawa, K.; Chavez, L.; Hakre, S.; Calvanese, V.; Verdin, E. Reactivation of latent HIV by histone deacetylase inhibitors. Trends Microbiol. 2013, 21, 277–285. [Google Scholar] [CrossRef]
- Lachmann, R.; Bevan, M.A.; Kim, S.; Patel, N.; Hawrylowicz, C.; Vyakarnam, A.; Peters, B.S. A comparative phase 1 clinical trial to identify anti-infective mechanisms of vitamin D in people with HIV infection. Aids 2015, 29, 1127–1135. [Google Scholar] [CrossRef]
- Bhan, I.; Camargo, C.A., Jr.; Wenger, J.; Ricciardi, C.; Ye, J.; Borregaard, N.; Thadhani, R. Circulating levels of 25-hydroxyvitamin D and human cathelicidin in healthy adults. J. Allergy Clin. Immunol. 2011, 127, 1302–1304. [Google Scholar] [CrossRef]
- Ashenafi, S.; Mazurek, J.; Rehn, A.; Lemma, B.; Aderaye, G.; Bekele, A.; Assefa, G.; Chanyalew, M.; Aseffa, A.; Andersson, J.; et al. Vitamin D3 Status and the Association with Human Cathelicidin Expression in Patients with Different Clinical Forms of Active Tuberculosis. Nutrients 2018, 10, 721. [Google Scholar] [CrossRef]
- Wirthgen, E.; Hoeflich, A. Endotoxin-Induced Tryptophan Degradation along the Kynurenine Pathway. The Role of Indolamine 2,3-Dioxygenase and Aryl Hydrocarbon Receptor-Mediated Immunosuppressive Effects in Endotoxin Tolerance and Cancer and Its Implications for Immunoparalysis. J. Amino. Acids 2015, 2015, 973548. [Google Scholar] [CrossRef]
- Gurav, A.; Sivaprakasam, S.; Bhutia, Y.D.; Boettger, T.; Singh, N.; Ganapathy, V. Slc5a8, a Na+-coupled high-affinity transporter for short-chain fatty acids, is a conditional tumour suppressor in colon that protects against colitis and colon cancer under low-fibre dietary conditions. Biochem. J. 2015, 469, 267–278. [Google Scholar] [CrossRef]
- Obeid, R.; Awwad, H.M.; Kirsch, S.H.; Waldura, C.; Herrmann, W.; Graeber, S.; Geisel, J. Plasma trimethylamine-N-oxide following supplementation with vitamin D or D plus B vitamins. Mol. Nutr. Food Res. 2017, 61, 1600358. [Google Scholar] [CrossRef]
- Missailidis, C.; Neogi, U.; Stenvinkel, P.; Troseid, M.; Nowak, P.; Bergman, P. The microbial metabolite trimethylamine-N-oxide in association with inflammation and microbial dysregulation in three HIV cohorts at various disease stages. Aids 2018, 32, 1589–1598. [Google Scholar] [CrossRef]
- Tang, W.H.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Placebo | p-Value | |
|---|---|---|---|
| Number of participants | 81 | 86 | |
| Female gender ** | 62 (78) | 70 (81) | 0.56 |
| Age (years *) | 32 (19–59) | 30 (19–62) | 0.63 |
| Body mass index (BMI) | 21 (18–27) | 22 (18–28) | 0.45 |
| Viral load (log10 copies/mL) | 4.0 (3.3–5.0) | 3.8 (3.2–5.1) | 0.20 |
| CD4 (cells/µL) | 410 (280–700) | 410 (270–570) | 0.37 |
| CD4/CD8 ratio | 0.42 (0.23–0.79) | 0.46 (0.24–0.81) | 0.85 |
| 25(OH)D, (nmoL/L) | 37 (22–69) | 38 (17–72) | 0.91 |
| Deficient (< 25 nmoL/L) n (%) | 15 (19) | 25 (29) | |
| Insufficient (25–50 nmoL/L) n (%) | 43 (53) | 35 (41) | |
| Sufficient (> 50 nmoL/L) n (%) | 23 (28) | 27 (30) |
| Treatment (n = 81) | Placebo (n = 86) | |||||
|---|---|---|---|---|---|---|
| Baseline | Week 16 | p-Value | Baseline | Week 16 | p-Value | |
| sCD14 (µg/mL) | 1.8 (1.0–3.7) | 1.9 (1.1–3.4) | 0.82 | 1.9 (1.2–4.0) | 2.2 (1.1–4.1) | 0.21 |
| LL37 (µg/mL) | 0.6 (0.3–1.3) | 0.9 (0.4–1.4) | <0.001 | 0.7 (0.4–1.2) | 0.8 (0.4–1.5) | 0.004 |
| Kynurenine (µmoL/L) | 2.3 (1.6–3.6) | 2.5 (1.7–3.9) | 0.34 | 2.3 (1.2–3.6) | 2.5 (1.4–3.6) | 0.09 |
| Tryptophan (µmoL/L) | 36 (22–51) | 35 (20–53) | 0.25 | 35 (23–49) | 37 (20–53) | 0.51 |
| kyn/trp ratio | 0.07 (0.04–0.14) | 0.07 (0.04–0.12) | 0.35 | 0.06 (0.04–0.12) | 0.07 (0.04–0.11) | 0.14 |
| TMAO * (µmoL/L) | 2.1 (0.5–5.9) | 2.4 (0.9–6.3) | 0.25 | 2.5 (0.5–7.0) | 3.0 (0.8–7.0) | 0.12 |
| Choline (µmoL/L) | 62 (44–76) | 63 (46–79) | 0.71 | 56 (44–72) | 61 (47–78) | 0.10 |
| Betaine (µmoL/L) | 73 (45–130) | 69 (41–110) | 0.09 | 67 (47–100) | 72 (47–120) | 0.04 |
| Change in Dependent Variable | β | SE | p-Value |
|---|---|---|---|
| sCD14 (µg/mL) | −0.04 | 0.18 | 0.81 |
| LL37 (µg/mL) | 0.04 | 0.05 | 0.41 |
| Kynurenine (µmoL/L) | −0.09 | 0.13 | 0.45 |
| Tryptophan (µmoL/L) | 0.69 | 2.0 | 0.73 |
| kyn/trp ratio | 0.12 | 0.12 | 0.33 |
| TMAO (µmoL/L) | 0.08 | 0.83 | 0.93 |
| Choline (µmoL/L) | −2.6 | 2.4 | 0.29 |
| Betaine (µmoL/L) | −11 | 4 | 0.007 |
| Change in Variable | Treatment n = 58 | Placebo n = 60 | p-Value |
|---|---|---|---|
| 25(OH)D (nmoL/L) | 79 (51–120) | 2.0 (−9–17) | <0.001 |
| sCD14 (µg/mL) | 0.18 (−1.34–1.22) | 0.05 (−1.07–1.12) | 0.76 |
| LL37 (µg/mL) | 0.16 (−0.19–0.64) | 0.05 (−0.34–0.32) | 0.02 |
| Kynurenine (µmoL/L) | 0.05 (−0.96–1.2) | 0.17 (−0.03–0.03) | 0.50 |
| Tryptophan (µmoL/L) | 1.3 (−10–19) | 2.2 (−14–14) | 0.87 |
| kyn/trp ratio | −0.00 (−0.03–0.03) | 0.00 (−0.03–0.03) | 0.28 |
| TMAO (µmoL/L) | 0.35 (−3.1–2.7) | 0.27 (−2.7–5.6) | 0.55 |
| Choline (µmoL/L) | 1.5 (−15–16) | 9.6 (−11–19) | 0.03 |
| Betaine (µmoL/L) | −2.7 (−33–31) | 3.5 (−24–33) | 0.22 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Missailidis, C.; Sørensen, N.; Ashenafi, S.; Amogne, W.; Kassa, E.; Bekele, A.; Getachew, M.; Gebreselassie, N.; Aseffa, A.; Aderaye, G.; et al. Vitamin D and Phenylbutyrate Supplementation Does Not Modulate Gut Derived Immune Activation in HIV-1. Nutrients 2019, 11, 1675. https://doi.org/10.3390/nu11071675
Missailidis C, Sørensen N, Ashenafi S, Amogne W, Kassa E, Bekele A, Getachew M, Gebreselassie N, Aseffa A, Aderaye G, et al. Vitamin D and Phenylbutyrate Supplementation Does Not Modulate Gut Derived Immune Activation in HIV-1. Nutrients. 2019; 11(7):1675. https://doi.org/10.3390/nu11071675
Chicago/Turabian StyleMissailidis, Catharina, Nikolaj Sørensen, Senait Ashenafi, Wondwossen Amogne, Endale Kassa, Amsalu Bekele, Meron Getachew, Nebiat Gebreselassie, Abraham Aseffa, Getachew Aderaye, and et al. 2019. "Vitamin D and Phenylbutyrate Supplementation Does Not Modulate Gut Derived Immune Activation in HIV-1" Nutrients 11, no. 7: 1675. https://doi.org/10.3390/nu11071675
APA StyleMissailidis, C., Sørensen, N., Ashenafi, S., Amogne, W., Kassa, E., Bekele, A., Getachew, M., Gebreselassie, N., Aseffa, A., Aderaye, G., Andersson, J., Brighenti, S., & Bergman, P. (2019). Vitamin D and Phenylbutyrate Supplementation Does Not Modulate Gut Derived Immune Activation in HIV-1. Nutrients, 11(7), 1675. https://doi.org/10.3390/nu11071675