Modulation of Milk Allergenicity by Baking Milk in Foods: A Proteomic Investigation

 , ,
, ,  ,
, 
Abstract
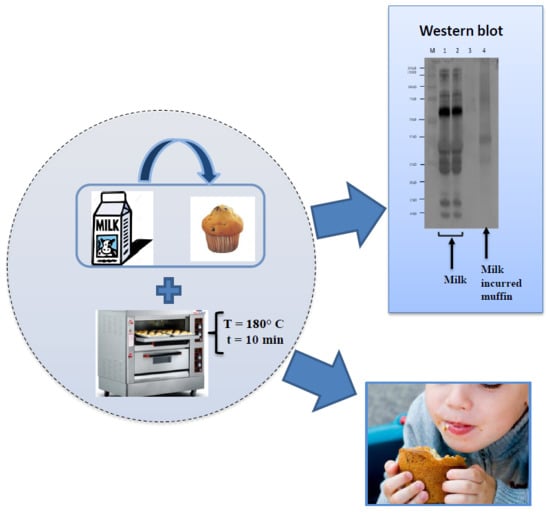
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sera of Milk Allergic Patients
2.3. Samples Preparation
2.4. Protein Extraction and Quantification
2.5. SDS-PAGE Analysis
2.6. In-Gel Protein Digestion
2.7. Protein Identification by Untargeted HR MS/MS Analysis
2.8. In Silico Analysis to Assess the Immunoreactivity of Milk Proteins after Baking Process
2.9. Immunoblot for IgE-Binding Assay
3. Results and Discussion
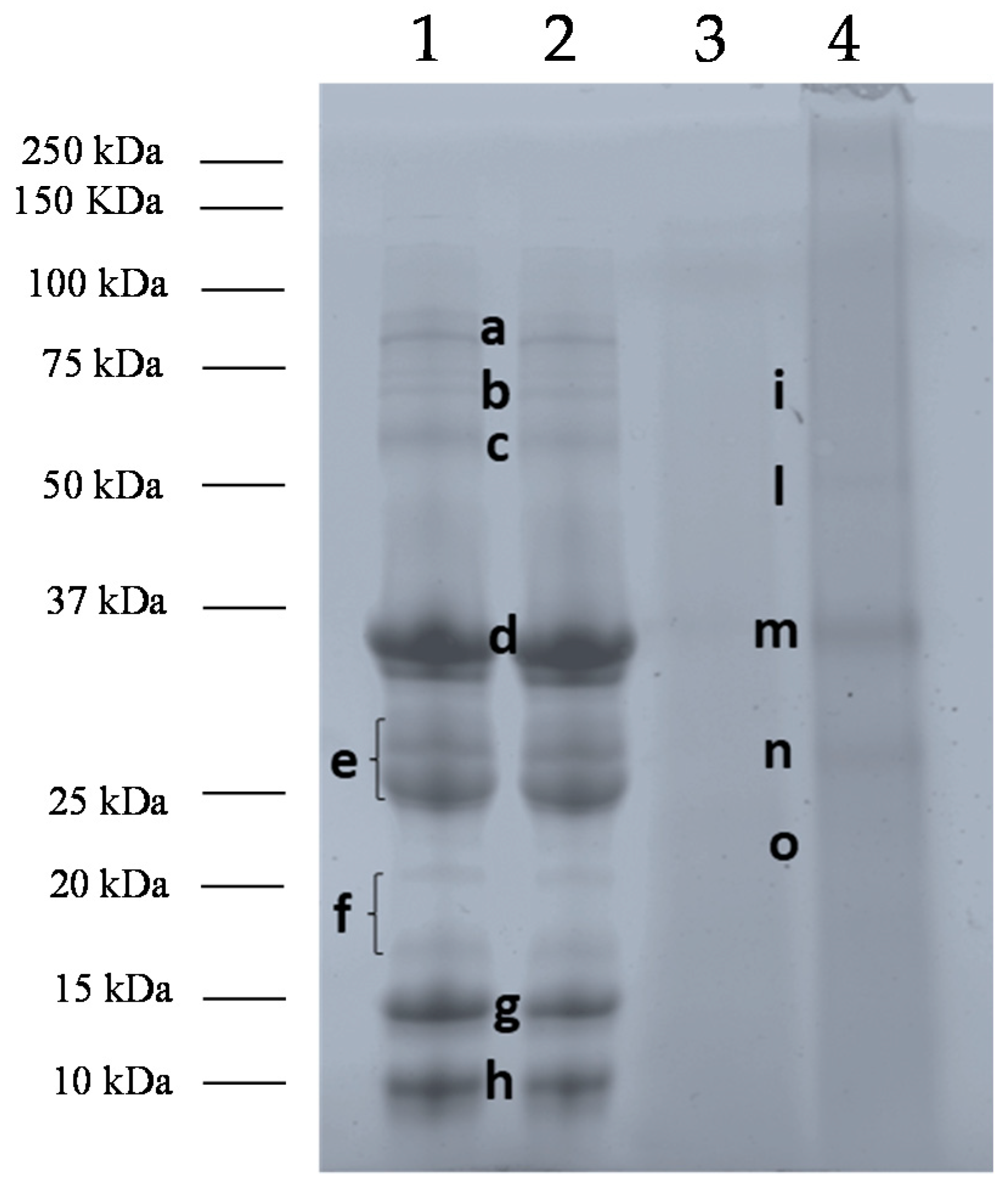
3.1. Effects of Baking and Matrix on Milk Protein Profiles
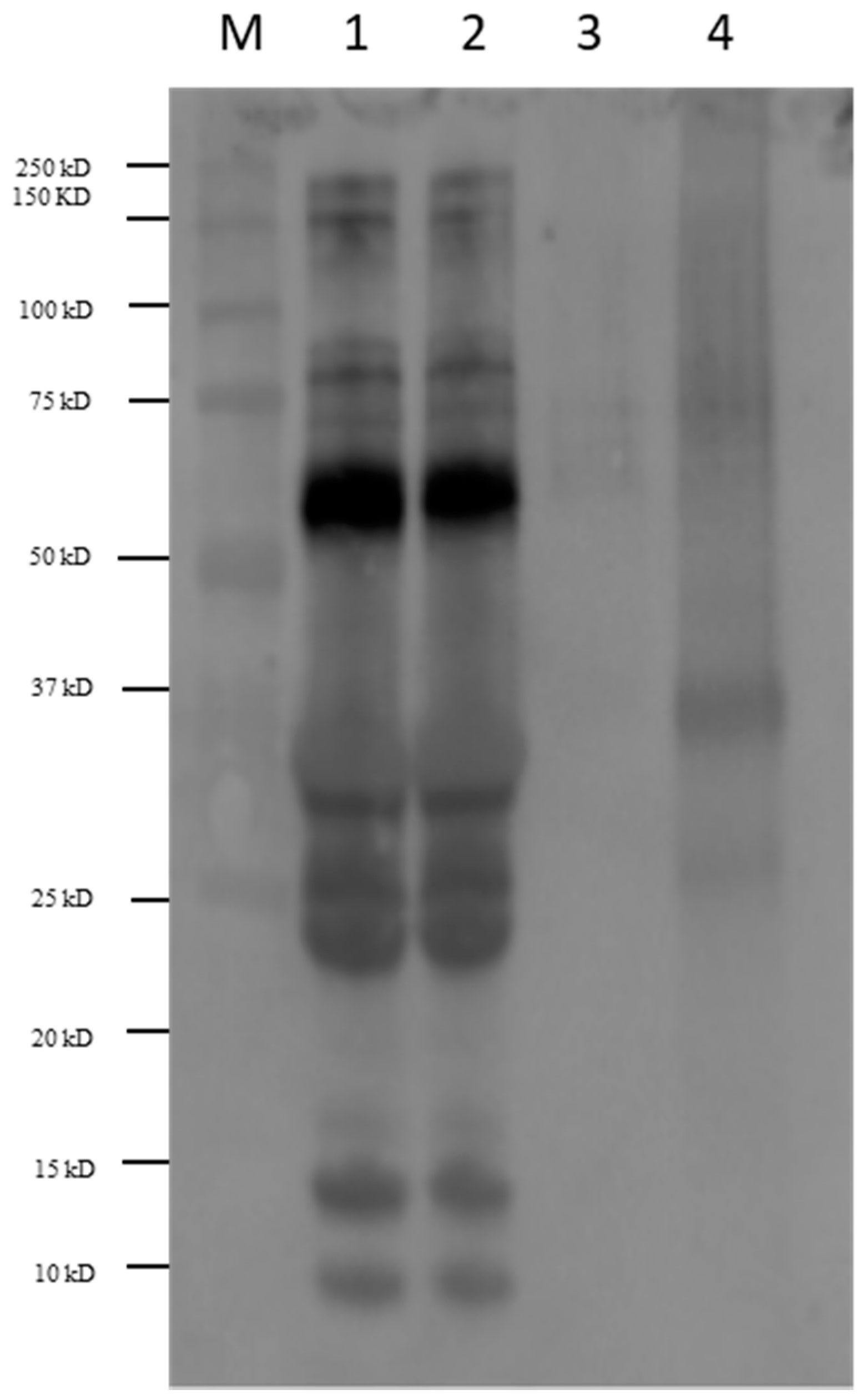
3.2. Immunoblot of Milk Products with Sera of Allergic Patients
3.3. In Silico Analyses to Assess IgE Binding Capacity of Milk Products
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hochwallner, H.; Schulmeister, U.; Swoboda, I.; Spitzauer, S.; Valenta, R. Cow’s Milk Allergy: From Allergens to New Forms of Diagnosis, Therapy and Prevention. Methods 2014, 66, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Venter, C.; Arshad, S.H. Epidemiology of Food Allergy. Pediatr. Clin. N. Am. 2011, 58, 327–349. [Google Scholar] [CrossRef] [PubMed]
- Nwaru, B.I.; Hickstein, L.; Panesar, S.S.; Roberts, G.; Muraro, A.; Sheikh, A. Prevalence of Common Food Allergies in Europe: A Systematic Review and Meta-Analysis. Allergy Eur. J. Allergy Clin. Immunol. 2014, 69, 992–1007. [Google Scholar] [CrossRef] [PubMed]
- Fiocchi, A.; Brozek, J.; Schunemann, H.; Bahna, S.L.; Von Berg, A.; Beyer, K.; Bozzola, M.; Bradsher, J.B.; Compalati, E.; Ebisawa, M.; et al. World Allergy Organization (WAO) Diagnosis and Rationale for Action Against Cow’s Milk Allergy (DRACMA) Guidelines. World Allergy Organ. J. 2010, 3, 57–161. [Google Scholar] [CrossRef] [PubMed]
- Skripak, J.M.; Matsui, E.C.; Mudd, K.; Wood, R.A. The Natural History of IGE-mediated Cow’s Milk Allergy. J. Allergy Clin. Immunol. 2007, 120, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Martorell, A.; Alonso, E.; Bone, J.; Echeverria, L.; Lopez, M.C.; Martin, F.; Nevot, S.; Plaza, A.M. Position Document: IGE-mediated Allergy to Egg Protein Food Allergy Committee of SEICAP. Allergol Immunopathol. (Madr) 2013, 41, 320–336. [Google Scholar] [CrossRef] [PubMed]
- Villa, C.; Costa, J.; Oliveira, M.B.P.P.; Mafra, I. Bovine Milk Allergens: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 137–164. [Google Scholar] [CrossRef]
- Marchesseau, S.; Mani, J.-C.; Martineau, P.; Roquet, F.; Cuq, J.-L.; Pugniere, M. Casein Interactions Studied by the Surface Plasmon Resonance Technique. J. Dairy Sci. 2002, 85, 2711–2721. [Google Scholar] [CrossRef]
- Broyard, C.; Gaucheron, F. Modifications of Structures and Functions of Caseins: A Scientific and Technological Challenge. Dairy Sci. Technol. 2015, 95, 831–862. [Google Scholar] [CrossRef]
- Foegeding, E.A.; Mleko, S.W. Whey Protein Products. U.S. Patent 6,383,551, 7 May 2002. [Google Scholar]
- Kontopidis, G.; Holt, C.; Sawyer, L. Lactoglobulin: Binding Properties, Structure, and Function. J. Dairy Sci. 2004, 87, 785–796. [Google Scholar] [CrossRef]
- Mulcahy, E.M.; Fargier-Lagrange, M.; Mulvihill, D.M.; O’Mahony, J.A. Characterisation of Heat-induced Protein Aggregation in Whey Protein Isolate and the Influence of Aggregation on the Availability of Amino Groups as Measured by the ORTHO-Phthaldialdehyde (OPA) and Trinitrobenzenesulfonic Acid (TNBS) methods. Food Chem. 2017, 229, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Vitaliti, G.; Cimino, C.; Coco, A.; Pratico, A.D.; Lionetti, E. The Immunopathogenesis of Cows Milk Protein Allergy (CMPA). Ital. J. Pediatr. 2012, 38, 35. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Land, M. Baked Milk and Baked Egg Oral Immunotherapy. Immunotherapy 2017, 9, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Mori, F.; Cianferoni, A.; Brambilla, A.; Barni, S.; Sarti, L.; Pucci, N.; De Martino, M.; Novembre, E. Side Effects and their Impact on the Success of Milk Oral Immunotherapy (OIT) in Children. Int. J. Immunopathol. Pharmacol. 2017, 30, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Caubet, J.C.; Nowak-Wȩgrzyn, A.; Moshier, E.; Godbold, J.; Wang, J.; Sampson, H.A. Utility of Casein-specific IGE Levels in Predicting Reactivity to Baked Milk. J. Allergy Clin. Immunol. 2013, 131, 222–224. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Wegrzyn, A.; Bloom, K.A.; Sicherer, S.H.; Shreffler, W.G.; Noone, S.; Wanich, N.; Sampson, H.A. Tolerance to Extensively Heated Milk in Children with Cow’s Milk Allergy. J. Allergy Clin. Immunol. 2008, 122, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.L.; Lanser, B.J. The Role of Baked Egg and Milk in the Diets of Allergic Children. Immunol. Allergy Clin. 2018, 38, 65–76. [Google Scholar] [CrossRef]
- Lambert, R.; Grimshaw, K.E.C.; Ellis, B.; Jaitly, J.; Roberts, G. Evidence that Eating Baked Egg or Milk Influences Egg or Milk Allergy Resolution: A Systematic Review. Clin. Exp. Allergy 2017, 47, 829–837. [Google Scholar] [CrossRef]
- Kim, J.S.; Nowak-Węgrzyn, A.; Sicherer, S.H.; Noone, S.; Moshier, E.L.; Sampson, H.A. Dietary Baked Milk Accelerates the Resolution of Cow’s Milk Allergy in Children. J. Allergy Clin. Immunol. 2011, 128, 125–131. [Google Scholar] [CrossRef]
- Considine, T.; Patel, H.A.; Anema, S.G.; Singh, H.; Creamer, L.K. Interactions of Milk Proteins during Heat and High Hydrostatic Pressure Treatments-A Review. Innov. Food Sci. Emerg. Technol. 2007, 8, 1–23. [Google Scholar] [CrossRef]
- Schulten, V.; Lauer, I.; Scheurer, S.; Thalhammer, T.; Bohle, B. A Food Matrix Reduces Digestion and Absorption of Food Allergens in Vivo. Mol. Nutr. Food Res. 2011, 55, 1484–1491. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, E.; Bavaro, S.L.; Forte, G.; Pilolli, R.; Monaci, L. Heat and Pressure Treatments on Almond Protein Stability and Chane in Immunoreactiveity after Simulated Human Digestion. Nutrients 2018, 10, 1679. [Google Scholar] [CrossRef] [PubMed]
- Bavaro, S.L.; Di Stasio, L.; Mamone, G.; De Angelis, E.; Nocerino, R.; Canani Berni, R.; Logrieco, A.F.; Montemurro, N.; Monaci, L. Effect of Thermal/Pressure Processing and Simulated Human Digestion on the Immunoreactivity of Extractable Peanut Alllergens. Food Res. Int. 2018, 109, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Węgrzyn, A.; Lawson, K.; Masilamani, M.; Kattan, J.; Bahnson, H.T.; Sampson, H.A. Increased Tolerance to Less Extensively Heat-Denatured (Baked) Milk Products in Milk-Allergic Children. J. Allergy Clin. Immunol. Pract. 2018, 6, 486–495. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, E.; Pilolli, R.; Bavaro, S.L.; Monaci, L. Insight Into the Gastro-Duodenal Digestion Resistance of Soybean Proteins and Potential Implications for Residual Immunogenicity. Food Funct. 2017, 8, 1599–1610. [Google Scholar] [CrossRef] [PubMed]
- Kleber, N.; Hinrichs, J. Antigenic Response of Beta-lactoglobulin in Thermally Treated Bovine Skim Milk and Sweet Whey. Milchwissenschaft 2007, 62, 121–124. [Google Scholar]
- Wijayanti, H.B.; Bansal, N.; Deeth, H.C. Stability of Whey Proteins during Thermal Processing: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1235–1251. [Google Scholar] [CrossRef]
- Perticaroli, S.; Nickels, J.D.; Ehlers, G.; Mamontov, E.; Sokolov, A.P. Dynamics and Rigidity in an Intrinsically Disordered Protein, β-casein. J. Phys. Chem. B 2014, 118, 7317–7326. [Google Scholar] [CrossRef]
- Tsabouri, S.; Douros, K.; Priftis, K. Cow’s Milk Allergenicity. Endocr. Metab. Immune Disord. Targets 2014, 14, 16–26. [Google Scholar] [CrossRef]
- Bhat, M.Y.; Dar, T.A.; Singh, L.R. Casein Proteins: Structural and Functional Aspects. In Milk Proteins-From Structure to Biological Properties and Health Aspects; Gigli, I., Ed.; IntechOpen: London, UK, 2016; Chapter 1; pp. 4–18. [Google Scholar] [CrossRef]
- Bloom, K.A.; Huang, F.R.; Bencharitiwong, R.; Bardina, L.; Ross, A.; Sampson, H.A.; Nowak-Wegrzyn, A. Effect of Heat Treatment on Milk and Egg Proteins Allergenicity. Pediatr. Allergy Immunol. 2014, 25, 740–746. [Google Scholar] [CrossRef]
- Morisawa, Y.; Kitamura, A.; Ujihara, T.; Zushi, N.; Kuzume, K.; Shimanouchi, Y.; Tamura, S.; Wakiguchi, H.; Saito, H.; Matsumoto, K. Effect of Heat Treatment and Enzymatic Digestion on the B Cell Epitopes of Cow’s Milk Proteins. Clin. Exp. Allergy 2009, 39, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Chatchatee, P.; Jarvinen, K.M.; Bardina, L.; Vila, L.; Beyer, K.; Sampson, H.A. Identification of IGE and IGG Binding Epitopes on β-and κ-casein in Cow’s Milk Allergic Patients. Clin. Exp. Allergy 2001, 31, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Dupont, D.; Mandalari, G.; Molle, D.; Jardin, J.; Rolet-Repecaud, O.; Duboz, G.; Leonil, J.; Mills, C.E.N.; Mackie, A.R. Food Processing Increases Casein Resistance to Simulated Infant Digestion. Mol. Nutr. Food Res. 2010, 54, 1677–1689. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.C.; Vissers, Y.M.; Baumert, J.L.; Faludi, R.; Feys, M.; Flanagan, S.; Herouet-Guicheneyg, C.; Holzhauserh, T.; Shimojoi, R.; Van der Boltj, N.; et al. Food Processing and Allergenicity. Food Chem. Toxicol. 2015, 80, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Bu, G.; Luo, Y.; Zheng, Z.; Zheng, H. Effect of Heat Treatment on the Antigenicity of Bovine α-lactalbumin and β-lactoglobulin in Whey Protein Isolate. Food Agric. Immunol. 2009, 20, 195–206. [Google Scholar] [CrossRef]
- Dumitraşcu, L.; Moschopoulou, E.; Aprodu, I.; Stanciu, S.; Rapeanu, G.; Stanciuc, N. Assessing the Heat Induced Changes in Major Cow and Non-cow Whey Proteins Conformation on Kinetic and Thermodynamic Basis. Small Rumin. Res. 2013, 111, 129–138. [Google Scholar] [CrossRef]
- Kleber, N.; Krause, I.; Illgner, S.; Hinrichs, J. The Antigenic Response of β-lactoglobulin is Modulated by Thermally Induced Aggregation. Eur. Food Res. Technol. 2004, 219, 105–110. [Google Scholar] [CrossRef]
- Jarvinen, K.M.; Chatchatee, P.; Bardina, L.; Beyer, K.; Sampson, H.A. IGE and IGG Binding Epitopes on α-lactalbumin and β-lactoglobulin in Cow’s Milk Allergy. Int. Arch. Allergy Immunol. 2001, 126, 111–118. [Google Scholar] [CrossRef]
- Rahaman, T.; Vasiljevic, T.; Ramchandran, L. Effect of Processing on Conformational Changes of Food Proteins Related to Allergenicity. Trends Food Sci. Technol. 2016, 49, 24–34. [Google Scholar] [CrossRef]
- Hochwallner, H.; Schulmeister, U.; Swoboda, I.; Balic, N.; Geller, B.; Nystrand, M.; Harlin, A.; Thalhamer, J.; Scheiblhofer, S.; Niggemann, B.; et al. Microarray and Allergenic Activity Assessment of Milk Allergens. Clin. Exp. Allergy 2010, 40, 1809–1818. [Google Scholar] [CrossRef]
- Shandilya, U.K.; Kapila, R.; Haq, R.M.; Kapila, S.; Kansal, V. Effect of Thermal Processing of Cow and Buffalo Milk on the Allergenic Response to Caseins and Whey Proteins in Mice. J. Sci. Food Agric. 2013, 93, 2287–2292. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.N.C.; Sancho, A.I.; Rigby, N.M.; Jenkins, J.A.; Mackie, A.R. Impact of Food Processing on the Structural and Allergenic Properties of Food Allergens. Mol. Nutr. Food Res. 2009, 53, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, S.; Eriksen, J.; Øysæd, L.K.; Idsøe, R.; Hill, D.J. T Cell Recognition Pattern of Bovine Milk αS1-Casein and its Peptides. Mol. Immunol. 2004, 41, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, S.; Hill, D.J.; Do, T.V. Evaluation of the Allergenicity and Antigenicity of Bovine-milk αs1-casein Using Extensively Purified Synthetic Peptides. Scand. J. Immunol. 2004, 60, 486–493. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Serum | Age (Years) | IgE Total (KU/L) | IgE to Cow’s Milk (KU/L) | IgE to Casein (KU/L) | Allergic Reaction Displayed |
|---|---|---|---|---|---|
| 1 | 8 | 5000 | 62 | 44 | anaphylaxis |
| 2 | 5 | 203 | 100 | 100 | anaphylaxis |
| 3 | 11 | 433 | 54 | 56 | anaphylaxis |
| 4 | 16 | 370 | 87 | 80 | anaphylaxis |
| 5 | 6 | 4786 | 56 | 34 | urthicaria |
| 6 | 5 | 4662 | 100 | 100 | vomit |
| Sample | Band | Accession Number | Allergenic Proteins | Allergen Code | Coverage | Filtered Peptides |
|---|---|---|---|---|---|---|
| Pasteurized Milk/Baked milk | c | A0A140T897 | Serum albumin | Bos d 6 | 38.22 | 26 |
| d | P02662 | αS1-casein | Bos d 9 | 33.17 | 1 | |
| P02663 | αS2-casein | Bos d 10 | 40.54 | 12 | ||
| B5B3R8 | αS1-casein | Bos d 9 | 33.17 | 1 | ||
| e | A0A140T8A9 | κ-casein | Bos d 12 | 23.15 | 4 | |
| A0A1Y0KDJ6 | β-casein | Bos d 11 | 25 | 4 | ||
| J9UHS4 | β-casein | Bos d 11 | 28.57 | 1 | ||
| P02663 | αS2-casein | Bos d 10 | 22.97 | 6 | ||
| f | B5B0D4 | β-lactoglobulin | Bos d 5 | 30.89 | 6 | |
| Q28049 | α-lactalbumin | Bos d 4 | 23.,57 | 4 | ||
| g | B5B0D4 | β-lactoglobulin | Bos d 5 | 71.91 | 12 | |
| h | Q28049 | α-lactalbumin | Bos d 4 | 44.71 | 5 | |
| CM incurred muffins | i | P02662 | αS1-casein | Bos d 9 | 17.28 | 2 |
| A0A1Y0KDJ6 | β-casein | Bos d 11 | 9.37 | 3 | ||
| m | B5B3R8 | αS1-casein | Bos d 9 | 39.71 | 1 | |
| P02662 | αS1-casein | Bos d 9 | 41.58 | 2 | ||
| n | A0A1Y0KDJ6 | β-casein | Bos d 11 | 23.,21 | 4 | |
| o | P02662 | αS1-casein | Bos d 9 | 8.87 | 1 |
| Protein Band | Peptide Sequence | Epitope ID |
|---|---|---|
| c | LGEYGFQNALIVR | 235209 |
| d | LHSMK | 70444, 115236, 11860, 35531, 56749, 109484, 109828, 115343, 115476 |
| EDVPSER | 663659, 28169, 109358, 30333, 30334, 31120, 31121, 115310, 48707, 78245, 115440, 190571, 68322 | |
| ITVDDK | 78138, 115315, 115477, 115479, 115532, 606543 | |
| LNFLK | 45706, 78144, 95351, 95560, 115226, 115512 | |
| e | EAMAPK | 115216 |
| FFSDK | 15893, 30141, 6173, 78257, 115305, 115404, 115449, 115465, 115733, 229682, 229689 | |
| GPFPIIV | 115251 | |
| g | TPEVDDEALEK | 65565, 78111, 96146, 13583, 56146, 95306, 95369, 95922, 96628, 115172, 115519, 146504, 222188 |
| TKIPAVFK | 33732, 95498, 96388, 33733, 46987, 78279, 96064, 96968, 115173, 115185, 115313, 222020 | |
| VLVLDTDYKK | 96517, 69827, 95545, 95579, 222193, 223163 | |
| IPAVFK | 33732, 96388, 31382, 33733, 46987, 78279, 96064, 96968, 98849, 115173, 115185, 115313, 222020 | |
| VLVLDTDYK | 96091, 96517, 69827, 95545, 95579, 222193, 223163 | |
| LSFNPTQLEEQCHI | 39349, 2820, 115382, 24090, 95389, 95574, 98777, 98893, 115427 | |
| VYVEELKPTPEGDLEILLQK | 72178, 32907, 96569, 97098, 32908, 72177, 95347, 96219, 98752, 98760, 99028, 99036, 224315 | |
| h | EQLTK | 115234, 227758, 558421 |
| m | FFVAPFPEVFGK | 38207, 43705, 15930, 15931, 44794, 67707, 69660, 110049, 115396, 115467, 190478, 659427, 659428 |
| HQGLPQEVLNENLLR | 115282, 31145, 50721, 50900, 109844, 115311, 675165 | |
| EPMIGVNQELAYFYPELFR | 12961, 13714, 13715, 13716, 23078, 45538, 45539 | |
| YLGYLEQLLR | 74689, 30334, 74687, 74688, 115482, 14100, 109358, 110060, 115060, 115122, 115213, 115440, 190580, 229693 | |
| VNELSK | 68473, 115253, 12896, 20548, 68472, 70058, 70059, 78158, 108948, 110049, 115068, 115531, 115544, 190572, 229694 | |
| EGIHAQQK | 24814, 12187, 30400, 41811, 115236, 24813, 109484, 109828, 115306, 115476, 190445, 606414 | |
| HIQKEDVPSER | 68322, 78245, 28169, 31120, 31121, 48707, 109358, 115310, 190571, 663657, 663658, 663659 | |
| EDVPSER | 68322, 30334, 78245, 28169, 30333, 31120, 31121, 48707, 109358, 115310, 115440, 190571, 663659 | |
| n | GPFPIIV | 115251, 658276, 670213, 671639, 673180, 673307, 688313 |
| DMPIQAFLLYQEPVLGPVR | 42283, 75481, 115675, 115796, 115847, 115866 | |
| AVPYPQR | 51169, 70443, 115430, 115439, 115495, 115694, 657013, 657014 | |
| VLPVPQK | 52358, 52359, 115495, 115835, 161678, 227654, 679766, 735655 | |
| o | FFVAPFPEVFGK | 15930, 15931, 659427, 659428, 38207, 190478, 43705, 44794, 115396, 115467, 67707, 69660, 110049 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bavaro, S.L.; De Angelis, E.; Barni, S.; Pilolli, R.; Mori, F.; Novembre, E.M.; Monaci, L. Modulation of Milk Allergenicity by Baking Milk in Foods: A Proteomic Investigation. Nutrients 2019, 11, 1536. https://doi.org/10.3390/nu11071536
Bavaro SL, De Angelis E, Barni S, Pilolli R, Mori F, Novembre EM, Monaci L. Modulation of Milk Allergenicity by Baking Milk in Foods: A Proteomic Investigation. Nutrients. 2019; 11(7):1536. https://doi.org/10.3390/nu11071536
Chicago/Turabian StyleBavaro, Simona L., Elisabetta De Angelis, Simona Barni, Rosa Pilolli, Francesca Mori, Elio. M. Novembre, and Linda Monaci. 2019. "Modulation of Milk Allergenicity by Baking Milk in Foods: A Proteomic Investigation" Nutrients 11, no. 7: 1536. https://doi.org/10.3390/nu11071536
APA StyleBavaro, S. L., De Angelis, E., Barni, S., Pilolli, R., Mori, F., Novembre, E. M., & Monaci, L. (2019). Modulation of Milk Allergenicity by Baking Milk in Foods: A Proteomic Investigation. Nutrients, 11(7), 1536. https://doi.org/10.3390/nu11071536