Nordic Seaweed and Diabetes Prevention: Exploratory Studies in KK-Ay Mice

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Seaweed Diets
2.2. Design
2.3. Oral Glucose Tolerance Test
2.4. Analytical Procedures and Statistics
3. Results
3.1. Animal Bodyweight
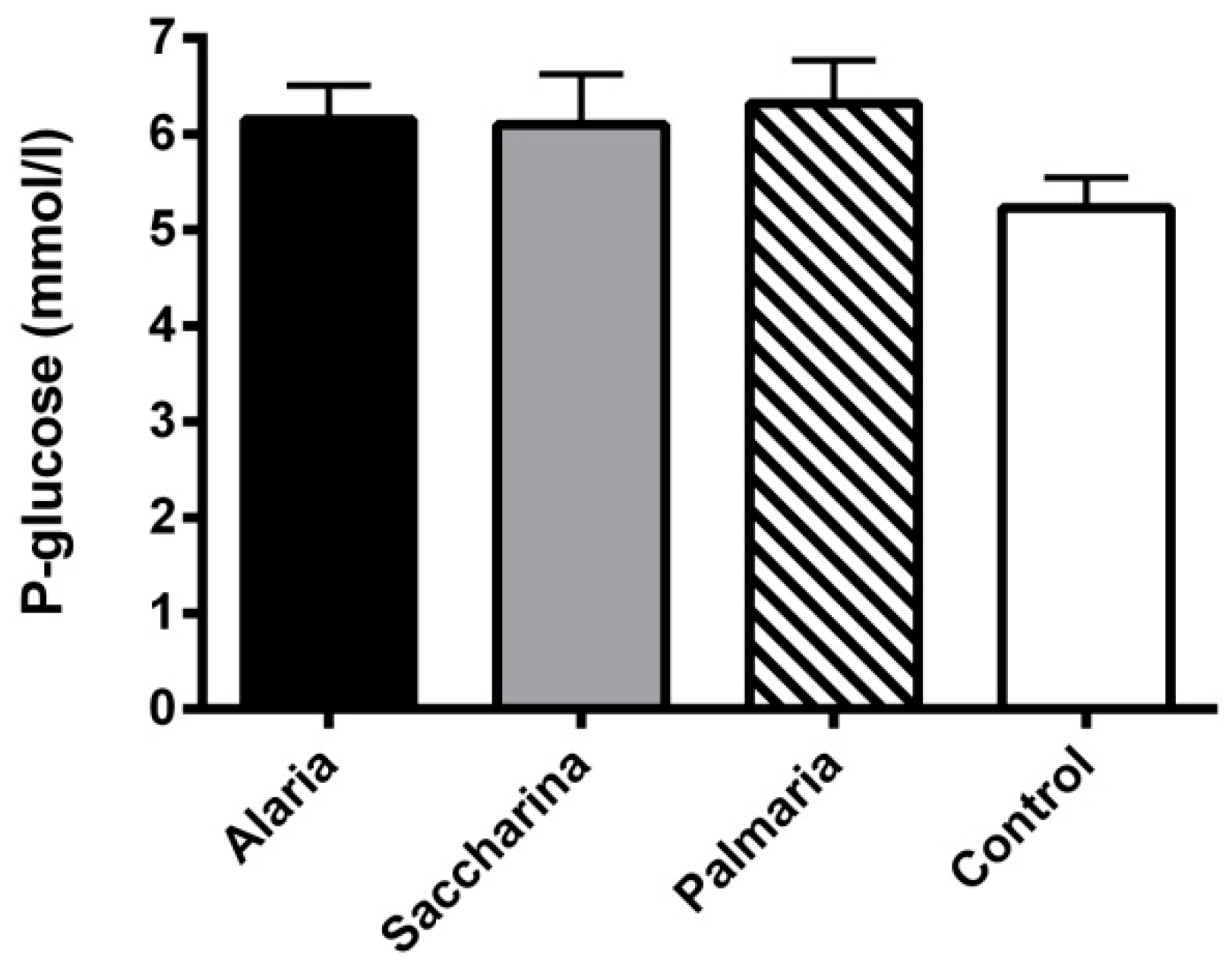
3.2. Plasma Glucose
3.3. Oral Glucose Tolerance Test
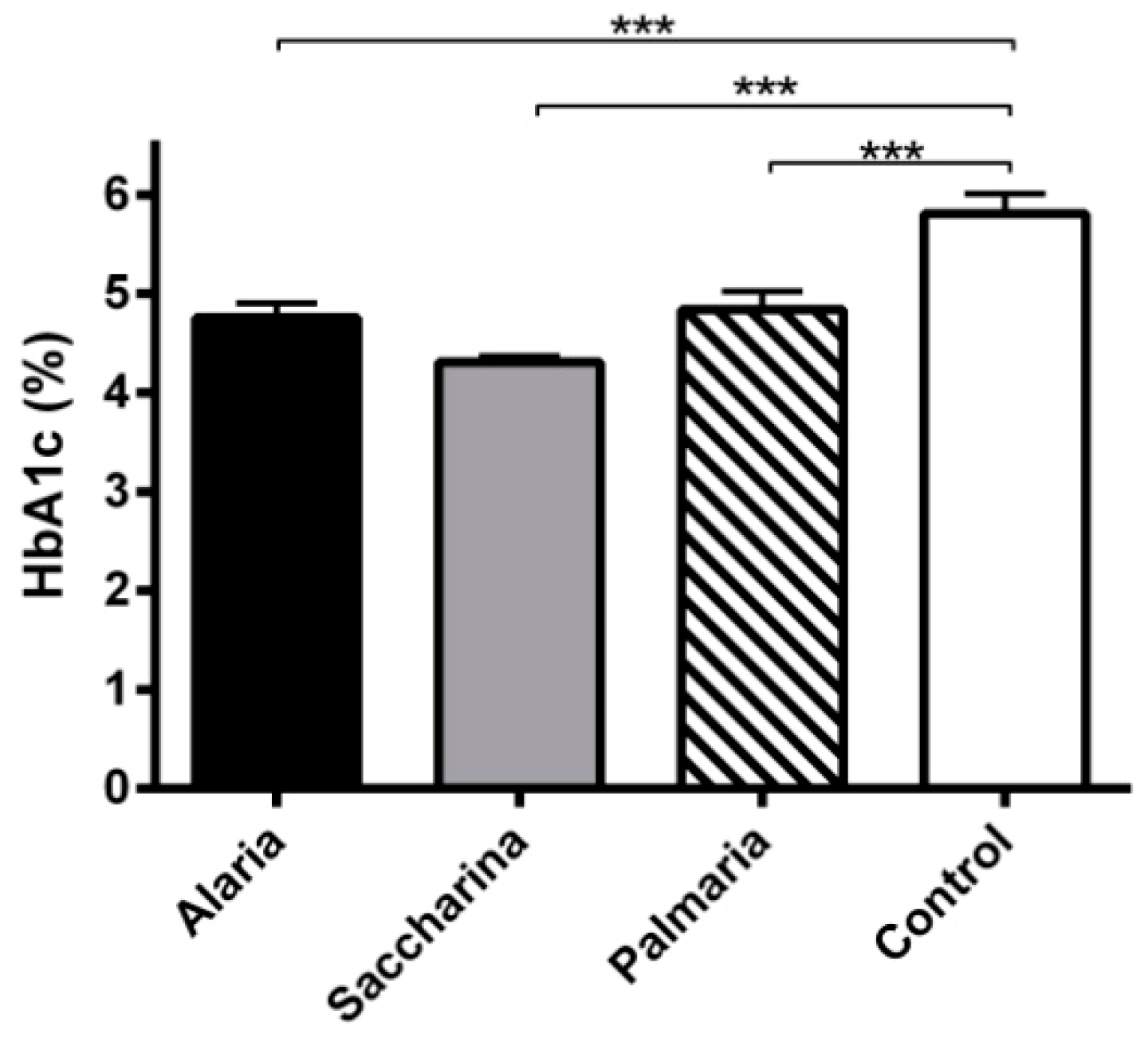
3.4. Glycated Hemoglobin
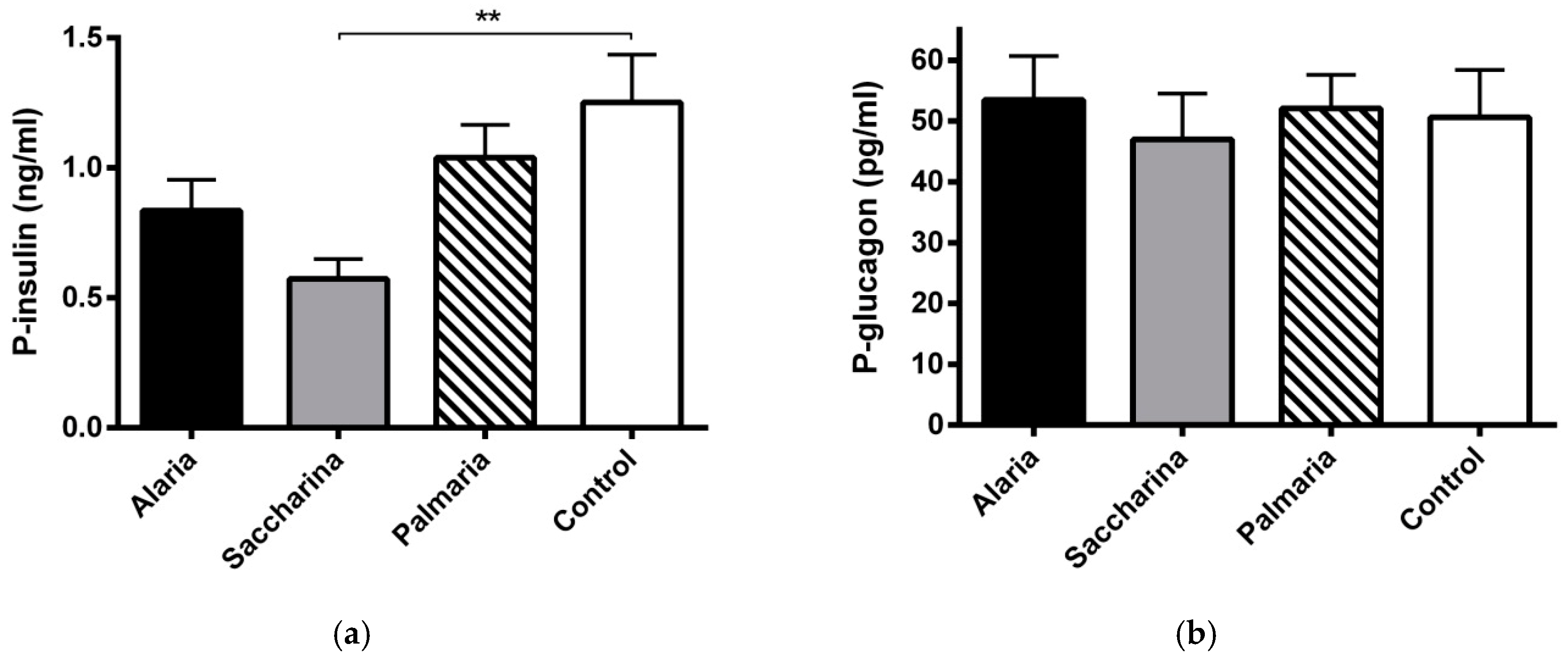
3.5. Plasma Insulin and Glucagon
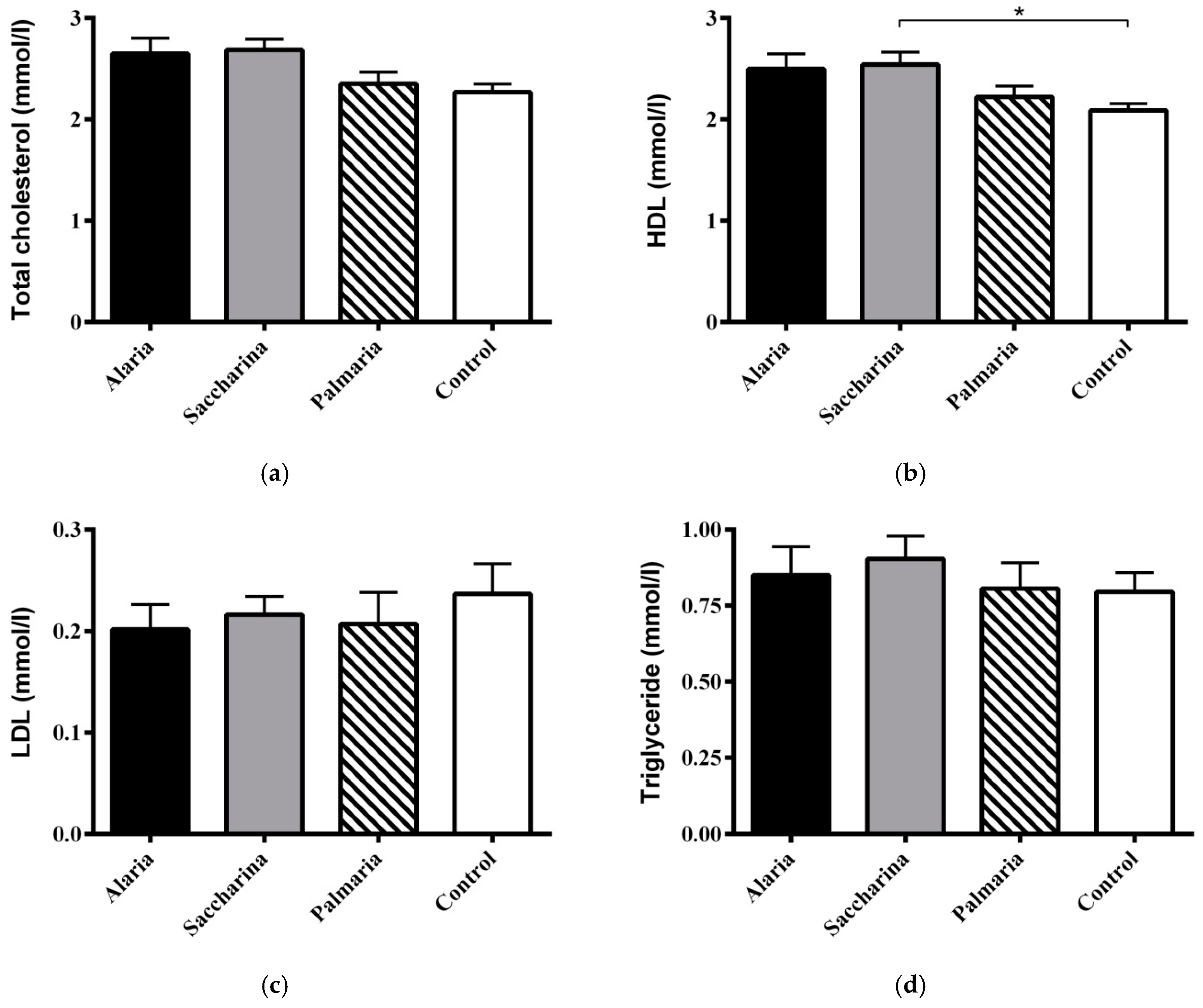
3.6. Circulating Cholesterol and Triglyceride
3.7. Thyroid Stimulating Hormone
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Tuomilehto, J.; Lindström, J.; Eriksson, J.G.; Valle, T.T.; Hämäläinen, H.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Laakso, M.; Louheranta, A.; Rastas, M.; et al. Finnish Diabetes Prevention Study Group. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. N. Engl. J. Med. 2001, 344, 1343–1350. [Google Scholar] [CrossRef] [PubMed]
- Galassi, A.; Reynolds, K.; He, J. Metabolic syndrome and risk of cardiovascular disease: A meta-analysis. Am. J. Med. 2006, 119, 812–819. [Google Scholar] [CrossRef] [PubMed]
- Cornier, M.A.; Dabelea, D.; Hernandez, T.L.; Lindstrom, R.C.; Steig, A.J.; Stob, N.R.; Van Pelt, R.E.; Wang, H.; Eckel, R.H. The metabolic syndrome. Endocr. Rev. 2008, 29, 777–822. [Google Scholar] [CrossRef] [PubMed]
- Knowler, W.C.; Barrett-Connor, E.; Fowler, S.E.; Hamman, R.F.; Lachin, J.M.; Walker, E.A.; Nathan, D.M. Diabetes Prevention Program Research Group. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med. 2002, 346, 393–403. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, H.C.; Vitek, L.; Nam, C.M. Algae consumption and risk of type 2 diabetes: Korean National Health and Nutrition Examination Survey in 2005. J. Nutr. Sci. Vitaminol. (Tokyo) 2010, 56, 13–18. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, J.Y.; Choi, W.H.; Lee, S.S. Effects of seaweed supplementation on blood glucose concentration, lipid profile, and antioxidant enzyme activities in patients with type 2 diabetes mellitus. Nutr. Res. Pract. 2008, 2, 62–67. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Torsdottir, I.; Alpsten, M.; Holm, G.; Sandberg, A.S.; Tölli, J. A small dose of soluble alginate-fiber affects postprandial glycemia and gastric emptying in humans with diabetes. J. Nutr. 1991, 121, 795–799. [Google Scholar] [CrossRef]
- Lakshmana, S.; Chandrasekaran, R.; Arjun, A.; Anantharaman, P. In vitro and in silico inhibition properties of fucoidan against α-amylase and α-D-glucosidase with relevance to type 2 diabetes mellitus. Carbohydr. Polym. 2019, 209, 350–355. [Google Scholar] [CrossRef]
- Kim, K.J.; Yoon, K.Y.; Lee, B.Y. Fucoidan regulate blood glucose homeostasis in C57BL/KSJ m+/+db and C57BL/KSJ db/db mice. Fitoterapia 2012, 83, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Sakai, C.; Abe, S.; Kouzuki, M.; Shimohiro, H.; Ota, Y.; Sakinada, H.; Takeuchi, T.; Okura, T.; Kasagi, T.; Hanaki, K. A Randomized Placebo-controlled Trial of an Oral Preparation of High Molecular Weight Fucoidan in Patients with Type 2 Diabetes with Evaluation of Taste Sensitivity. Yonago Acta Med. 2019, 62, 14–23. [Google Scholar] [CrossRef]
- Lee, H.; Park, H.; Heo, J.; Kang, M.; Ko, C.; Han, S.; Jeon, J. Dieckol isolated from Ecklonia cava inhibits α-glucosidase and α-amylase in vitro and alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. Food Chem. Toxicol. 2010, 48, 2633–2637. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.C.; Wijesinghe, W.A.; Lee, S.H.; Kang, S.M.; Ko, S.C.; Yang, X.; Kang, N.; Jeon, B.T.; Kim, J.; Lee, D.H.; et al. Dieckol isolated from brown seaweed Ecklonia cava attenuates type ІІ diabetes in db/db mouse model. Food Chem. Toxicol. 2013, 53, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, M.H.; Kang, S.M.; Ko, S.C.; Kang, M.C.; Cho, S.; Park, P.J.; Jeon, B.T.; Kim, S.K.; Han, J.S.; et al. Dieckol isolated from Ecklonia cava protects against high-glucose induced damage to rat insulinoma cells by reducing oxidative stress and apoptosis. Biosci. Biotechnol. Biochem. 2012, 76, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Han, J.S.; Heo, S.J.; Hwang, J.Y.; Jeon, Y.J. Protective effects of dieckol isolated from Ecklonia cava against high glucose-induced oxidative stress in human umbilical vein endothelial cells. Toxicol. In Vitro 2010, 24, 375–381. [Google Scholar] [CrossRef]
- Lee, H.; Jeon, J. Efficacy and safety of a dieckol-rich extract (AG-dieckol) of brown algae, Ecklonia cava, in pre-diabetic individuals: A double-blind, randomized, placebo-controlled clinical trial. Food Funct. 2015, 6, 853–858. [Google Scholar] [CrossRef]
- Kawamura-Konishi, Y.; Watanabe, N.; Saito, M.; Nakajima, N.; Sakaki, T.; Katayama, T.; Enomoto, T. Isolation of a new phlorotannin, a potent inhibitor of carbohydrate-hydrolyzing enzymes, from the brown alga Sargassum patens. J. Agric. Food Chem. 2012, 60, 5565–5570. [Google Scholar] [CrossRef]
- Apostolidis, E.; Lee, C.M. In vitro potential of Ascophyllumnodosum phenolic antioxidant-mediated alpha-glucosidase and alpha-amylase inhibition. J. Food Sci. 2010, 75, 97–102. [Google Scholar] [CrossRef]
- Zhang, J.; Tiller, C.; Shen, J.; Wang, C.; Girouard, G.S.; Dennis, D.; Barrow, C.J.; Miao, M.; Ewart, H.S. Antidiabetic properties of polysaccharide- and polyphenolic-enriched fractions from the brown seaweed Ascophyllumnodosum. Can. J. Physiol. Pharmacol. 2007, 85, 1116–1123. [Google Scholar] [CrossRef]
- Hertog, M.G.; Feskens, E.J.; Hollman, P.C.; Katan, M.B.; Kromhout, D. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Jeong, S.M.; Kang, M.J.; Choi, H.N.; Kim, J.H.; Kim, J.I. Quercetin ameliorates hyperglycemia and dyslipidemia and improves antioxidant status in type 2 diabetic db/db mice. Nutr. Res. Pract. 2012, 6, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Ding, Y.; Zhang, Z.; Cai, X.; Bao, L.; Li, Y. Quercetin but not quercitrin ameliorates tumor necrosis factor- alpha -induced insulin resistance in C2C12 skeletal muscle cells. Biol. Pharm. Bull. 2013, 36, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Miyashita, T.; Nishikawa, S.; Emi, S.; Tsukui, T.; Beppu, F.; Okada, T.; Miyashita, K. Fucoxanthin regulates adipocytokine mRNA expression in white adipose tissue of diabetic/obese KK-Ay mice. Arch. Biochem. Biophys. 2010, 504, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S.; Hosokawa, M.; Miyashita, K. Fucoxanthin promotes translocation and induction of glucose transporter 4 in skeletal muscles of diabetic/obese KK-A(y) mice. Phytomedicine 2012, 19, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undariapinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Mikami, N.; Hosokawa, M.; Miyashita, K.; Sohma, H.; Ito, Y.M.; Kokai, Y. Reduction of HbA1c levels by fucoxanthin-enriched akamoku oil possibly involves the thrifty allele of uncoupling protein 1 (UCP1): A randomised controlled trial in normal-weight and obese Japanese adults. J. Nutr. Sci. 2017, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, O.G.; Dawczynski, C.; Duelund, L.; Jahreis, G.; Vetter, W.; Schröder, M. On the human consumption of the red seaweed dulse (Palmaria palmata (L.) Weber & Mohr). J. Appl. Phycol. 2013, 25, 1777–1791. [Google Scholar] [CrossRef]
- Hansen, B. Determination of Nitrogen as elementary-N, an alternative to Kjeldahl. Acta Agric. Scand. 1989, 39, 113–118. [Google Scholar] [CrossRef]
- Stoldt, W. Vorschlag zur Vereinheitlichung der Fettbestimmung in Lebensmitteln. Fette Seifen Anstrichm. 1952, 54, 206–207. [Google Scholar] [CrossRef]
- John, G. Glycatedhemoglobin analysis. Ann. Clin. Biochem. 1997, 34, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Seca, A.; Pinto, D. Overview on the Antihypertensive and Anti-Obesity Effects of Secondary Metabolites from Seaweeds. Mar. Drugs 2018, 16, 237. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, G.; Yin, J.; Wang, Y.; Wang, J.; Xia, B.; Song, W. The fucoidan A2 from the brown seaweed Ascophyllumnodosum lowers lipid by improving reverse cholesterol transport in C57BL/6J mice fed a high-fat diet. J. Agric. Food Chem. 2019, 67, 5782–5791. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, Q.; Cui, W.; Zeng, Z.; Yang, W.; Zhang, C.; Zhao, H.; Gao, W.; Wang, X.; Luo, D. Low Molecular Weight Fucoidan Alleviates Cardiac Dysfunction in Diabetic Goto-Kakizaki Rats by Reducing Oxidative Stress and Cardiomyocyte Apoptosis. J. Diabetes Res. 2014, 2014, 420929. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alaria esculenta | Saccharina latissima | Palmaria palmata | |
|---|---|---|---|
| Carbohydrates % | 49.6 | 48.9 | 45 |
| Protein % | 16.1 | 11.4 | 20 |
| Fat % | 0.5 | 0.2 | 1 |
| Iodine mg/100g | 43 | 110 | 5–15 |
| Alaria esculenta | Saccharina latissima | Palmaria palmata | Control | |
|---|---|---|---|---|
| Energy KJ/100 g | 1324 | 1301.6 | 1326 | 1385.9 |
| Carbohydrates % | 52.3 | 52.2 | 51.4 | 53.4 |
| Protein % | 18.4 | 17.4 | 19.2 | 19.2 |
| Fat % | 3.3 | 3.2 | 3.4 | 4.1 |
| Alaria | Saccharina | Palmaria | Control | |
|---|---|---|---|---|
| Week 1 | 33.8 ± 2 g | 33.6 ± 2 g | 33.4 ± 2 g | 33.7 ± 2 g |
| Week 3 | 36.5 ± 3 g | 33.6 ± 3 g * | 36.2 ± 5 g | 37.5 ± 2 g |
| Week 5 | 38.9 ± 4 g | 33.6 ± 3 g ** | 37.2 ± 5 g | 39.8 ± 3 g |
| Week 7 | 39.3 ± 3 g | 34.3 ± 3 g *** | 37.9 ± 5 g | 40.5 ± 3 g |
| Week 11 | 39.3 ± 3 g | 35.8 ± 3 g *** | 40.5 ± 3 g | 41.5 ± 3 g |
| Alaria | Saccharina | Palmaria | Control | |
|---|---|---|---|---|
| iAUC (mmol/L × 240 min) | 1402 ± 205 | 1112 ± 164 | 1386 ± 190 | 1156 ± 121 |
| AUC (mmol/L × 240 min) | 2805 ± 270 | 2791 ± 254 | 3009 ± 251 | 2480 ± 123 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sørensen, L.E.; Jeppesen, P.B.; Christiansen, C.B.; Hermansen, K.; Gregersen, S. Nordic Seaweed and Diabetes Prevention: Exploratory Studies in KK-Ay Mice. Nutrients 2019, 11, 1435. https://doi.org/10.3390/nu11061435
Sørensen LE, Jeppesen PB, Christiansen CB, Hermansen K, Gregersen S. Nordic Seaweed and Diabetes Prevention: Exploratory Studies in KK-Ay Mice. Nutrients. 2019; 11(6):1435. https://doi.org/10.3390/nu11061435
Chicago/Turabian StyleSørensen, Lasse E., Per B. Jeppesen, Christine B. Christiansen, Kjeld Hermansen, and Søren Gregersen. 2019. "Nordic Seaweed and Diabetes Prevention: Exploratory Studies in KK-Ay Mice" Nutrients 11, no. 6: 1435. https://doi.org/10.3390/nu11061435
APA StyleSørensen, L. E., Jeppesen, P. B., Christiansen, C. B., Hermansen, K., & Gregersen, S. (2019). Nordic Seaweed and Diabetes Prevention: Exploratory Studies in KK-Ay Mice. Nutrients, 11(6), 1435. https://doi.org/10.3390/nu11061435